
Marker Assisted Selection of Malic-Consuming Saccharomyces cerevisiae Strains for Winemaking. Efficiency and Limits of a QTL’s Driven Breeding Program


Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strain Used and Culture Methods
2.2. Spore Mating and Purification
2.3. Cell Culture and DNA Extraction in Microplates
2.4. Mass ARRAY Genotyping
2.5. Alcoholic Fermentation Assays
2.5.1. Grape Must
2.5.2. Alcoholic Fermentation Monitoring
2.6. Malolactic Fermentations Monitoring
2.7. Enzymatic Assay of Wine
2.8. Use of Previous Phenotypic Datasets
2.9. Statistical Analyses
3. Results
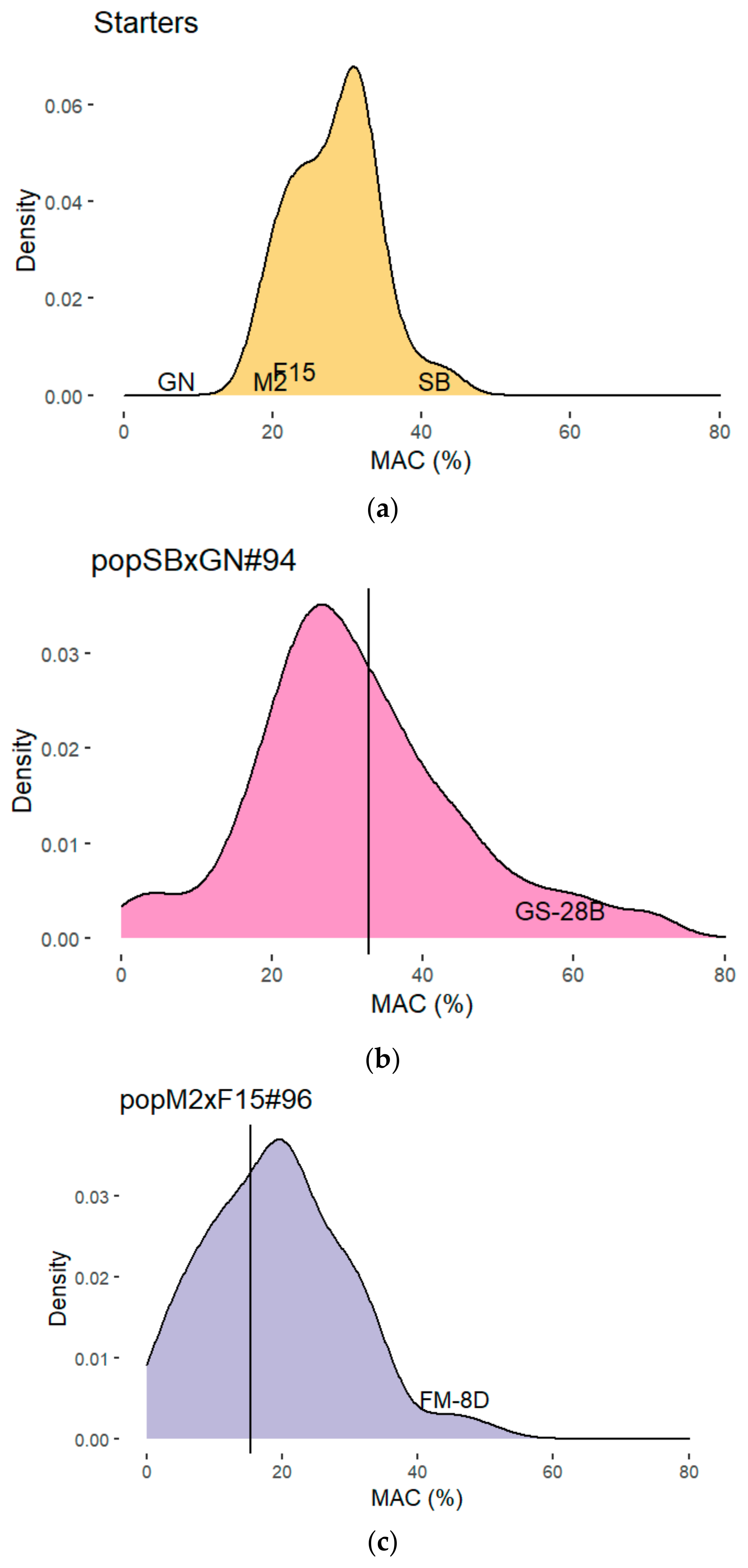
3.1. A Basic Breeding Strategy for Improving Malic Acid Consumption
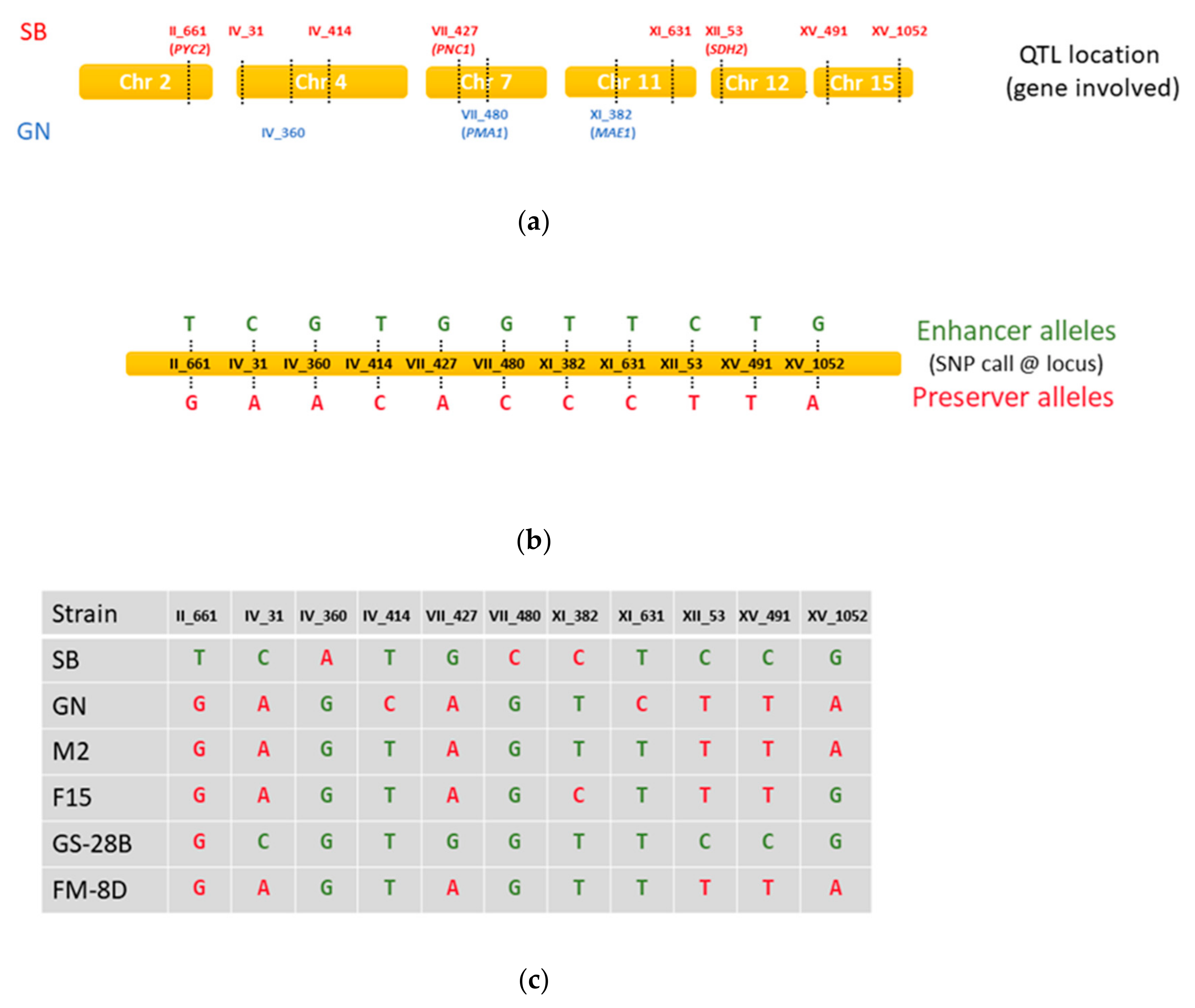
3.2. Use of Marker Assisted Selection (MAS) for Enhancing Malic Acid Consumption
3.3. Construction and Characterization of a Backcrossed FMGS-2 Population
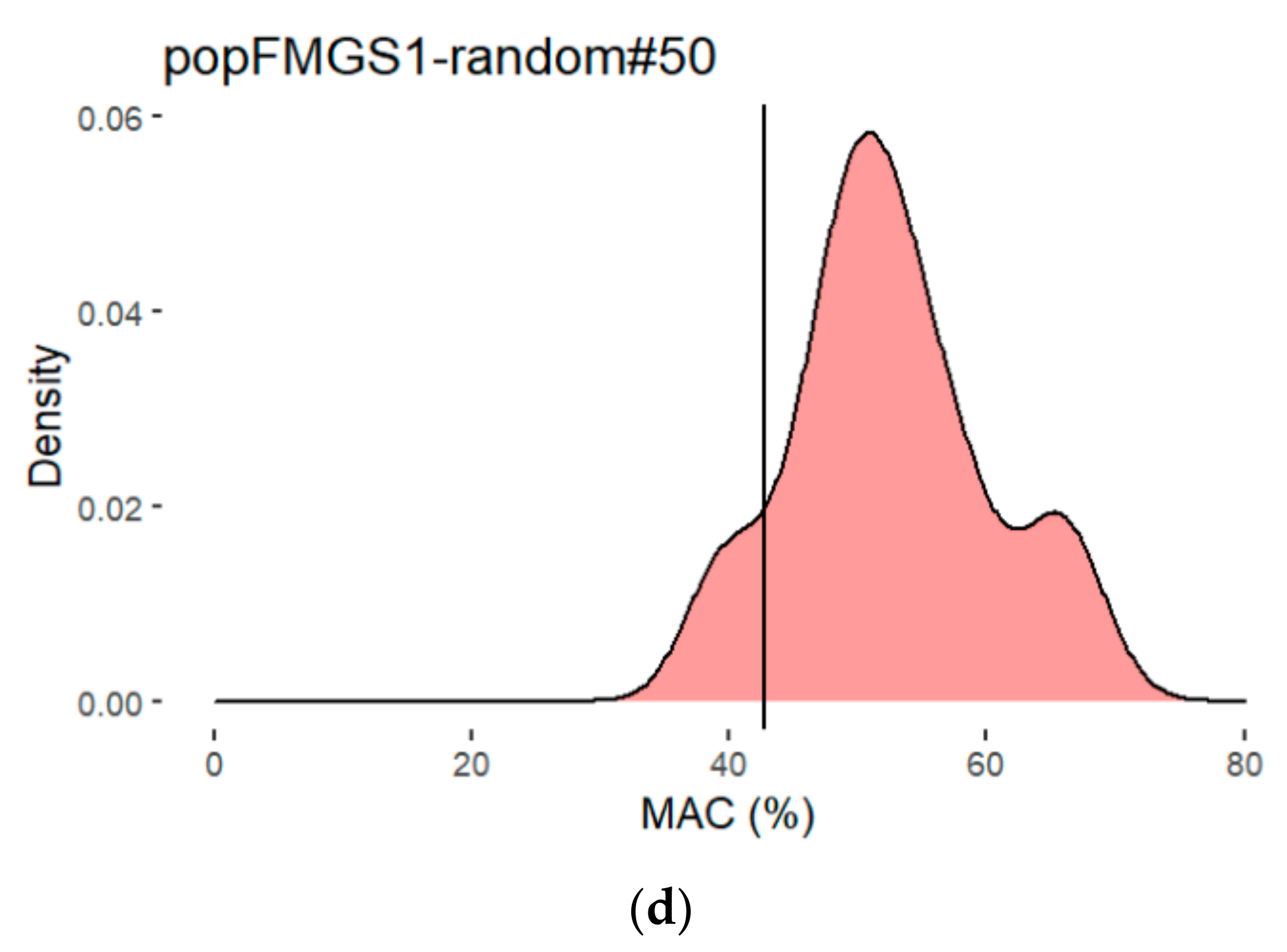
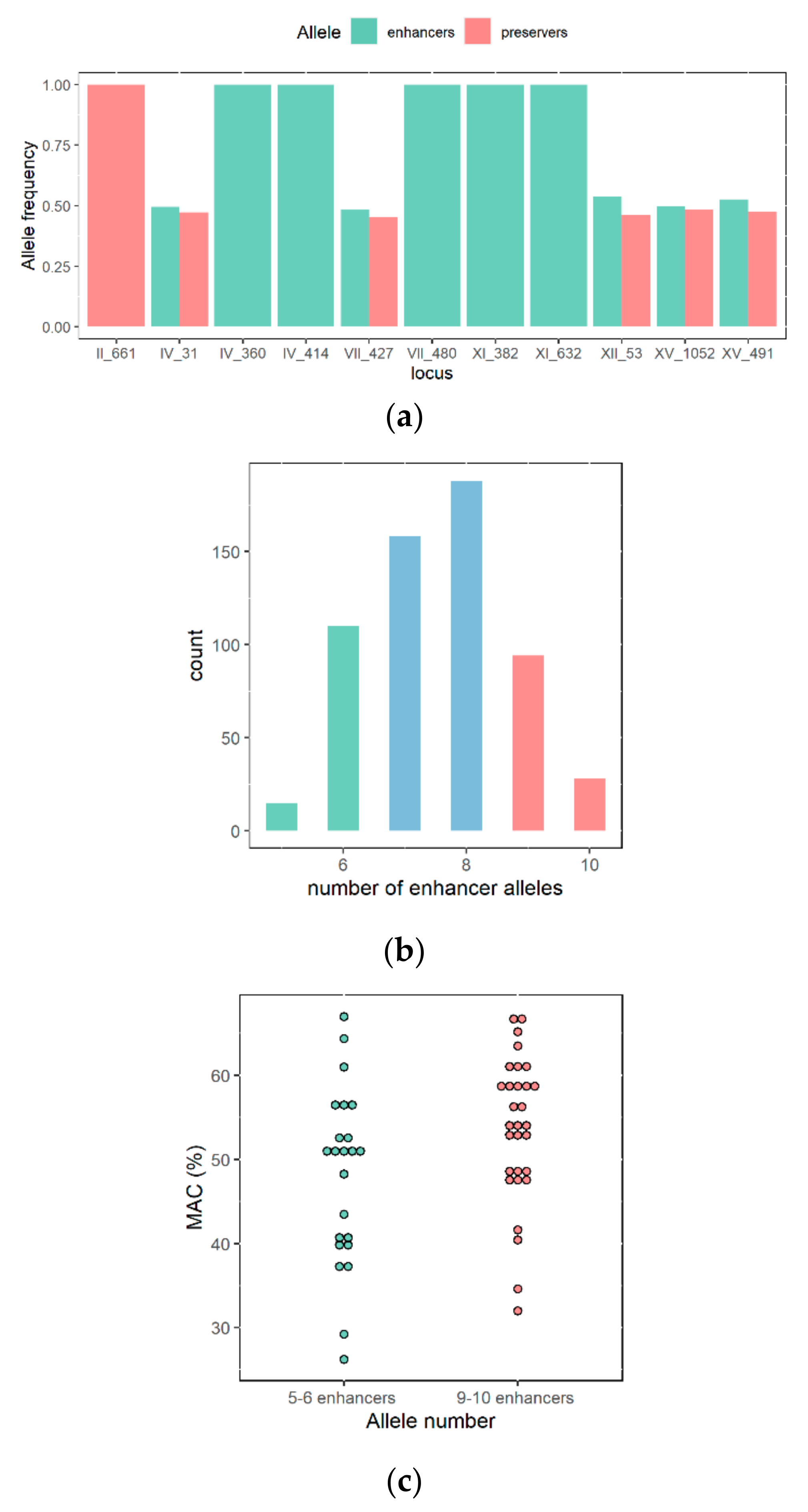
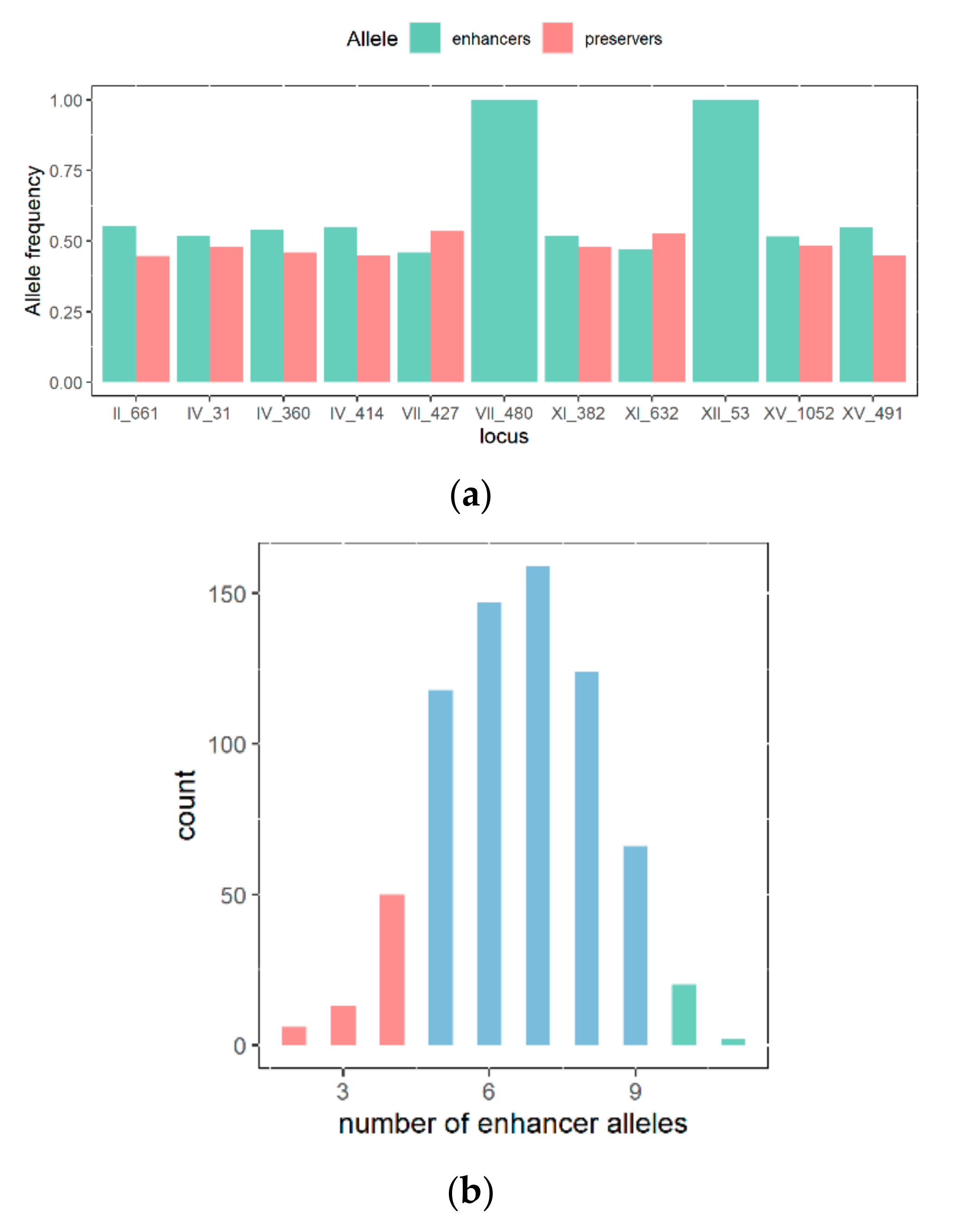
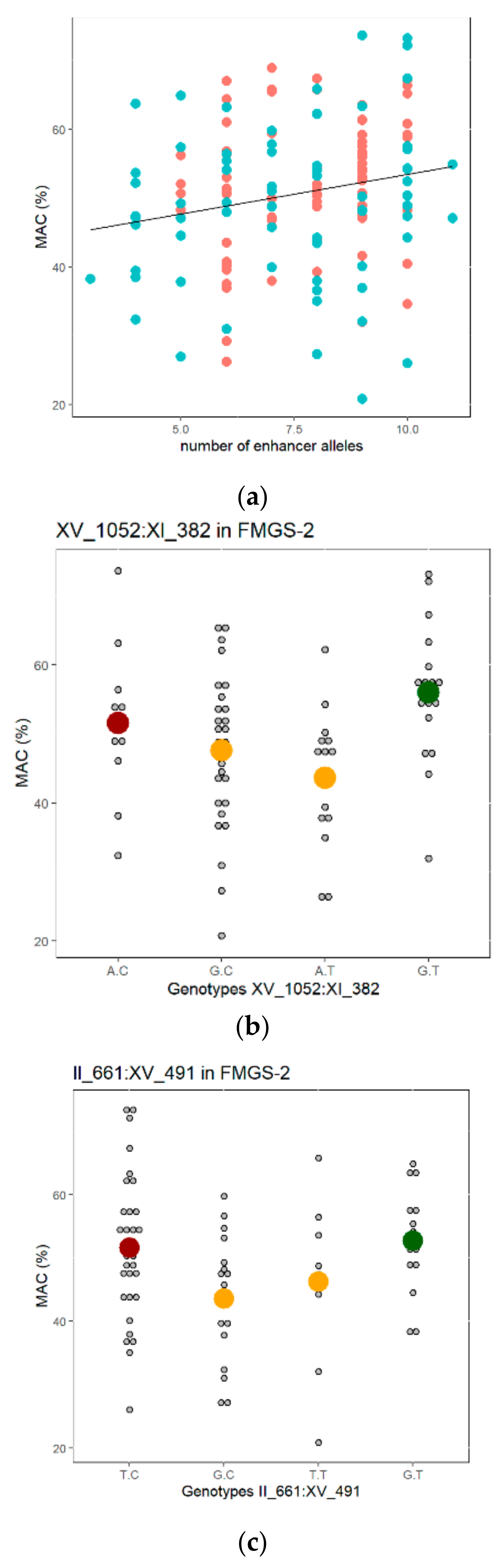
3.4. Study of QTL Penetrance in FMGS-1 and FMGS-2 Populations
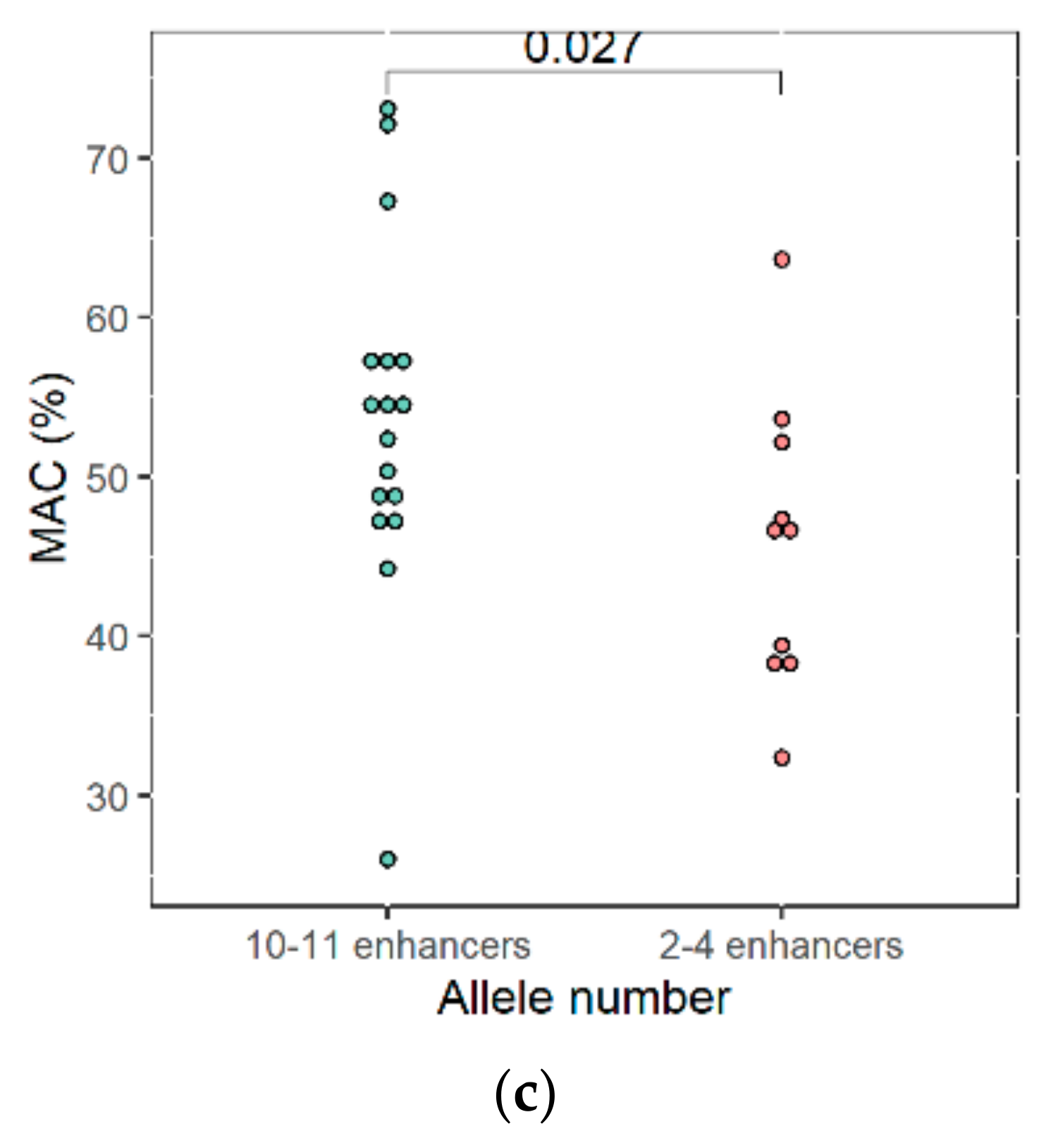
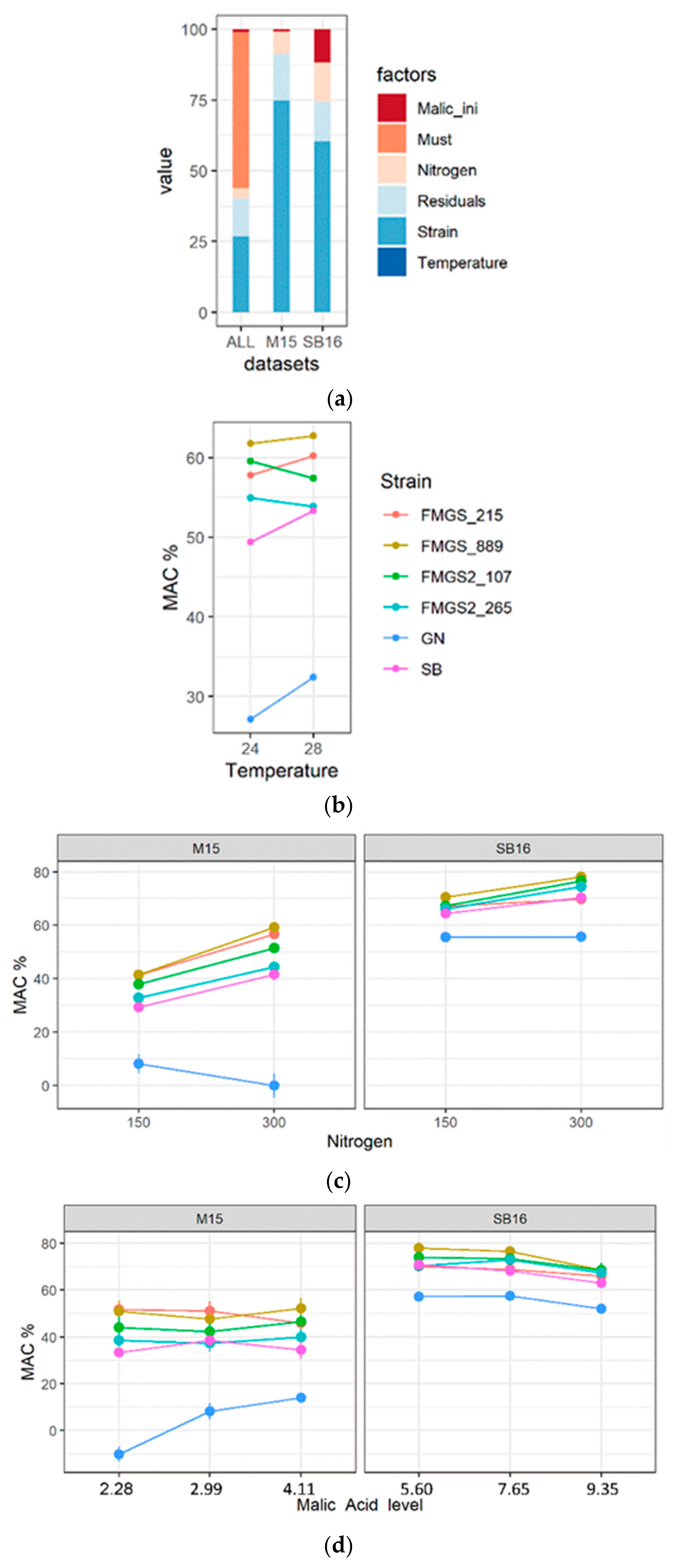
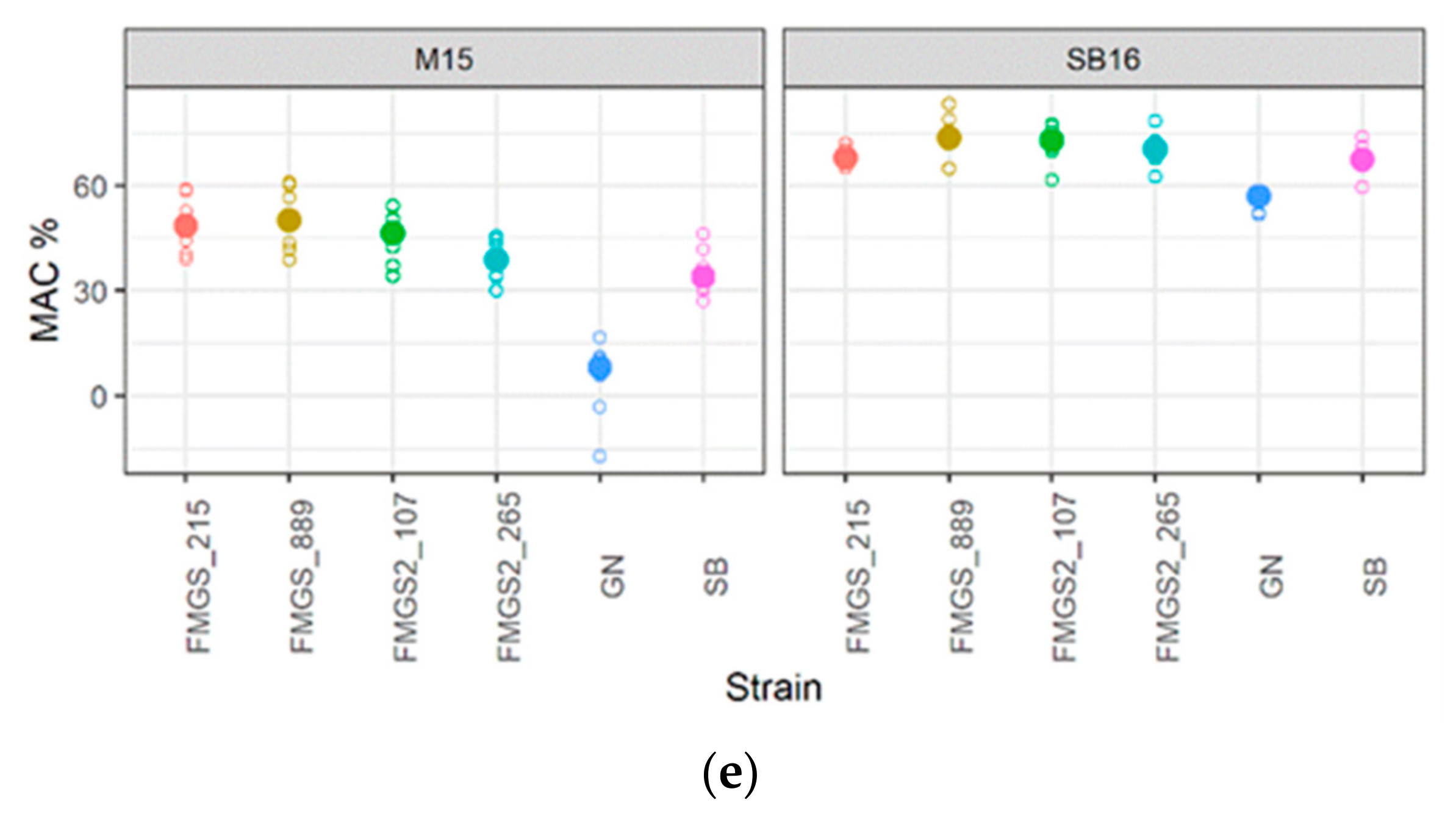
3.5. Phenotypic Characterization of High Malic Acid Consuming Strains
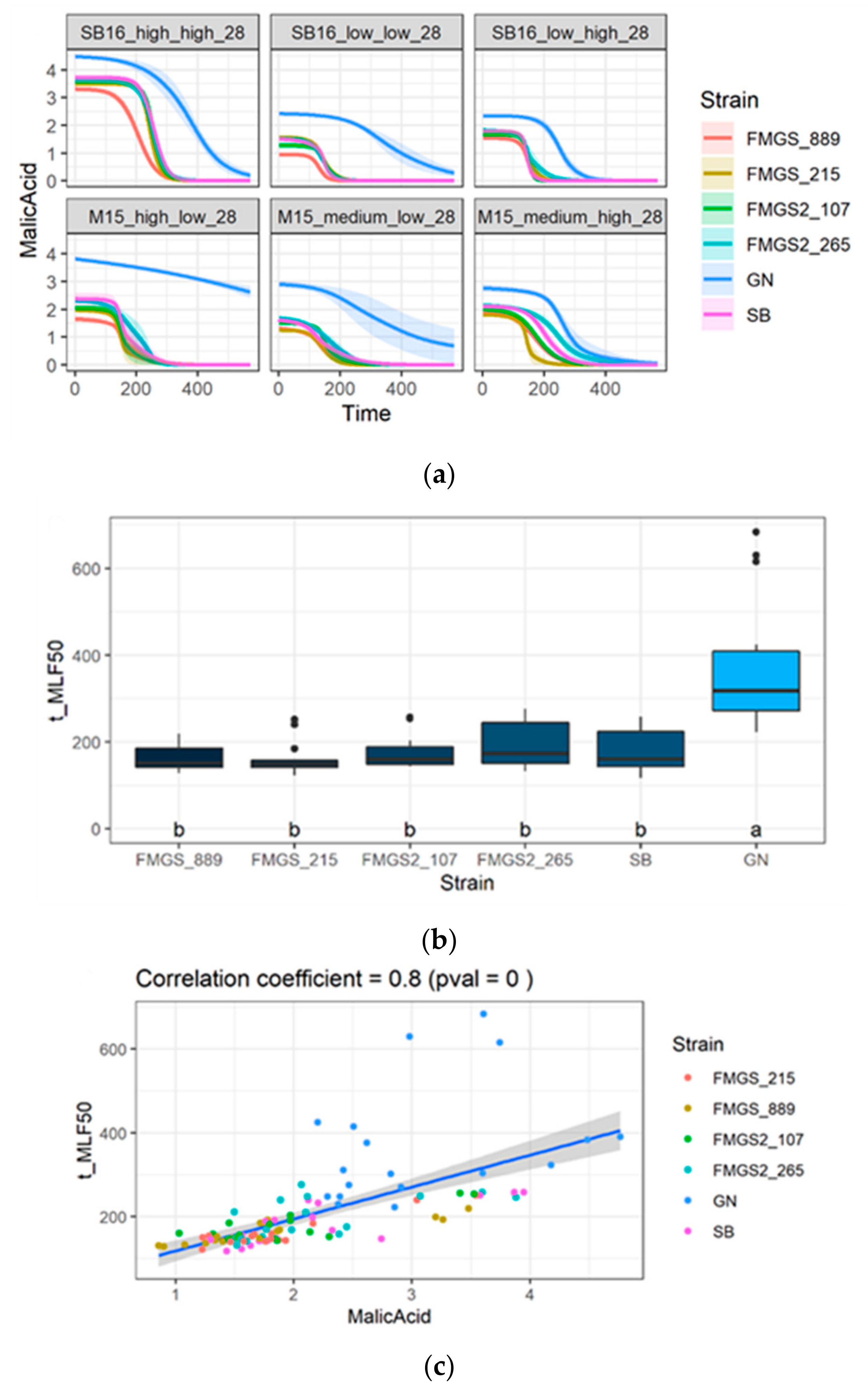
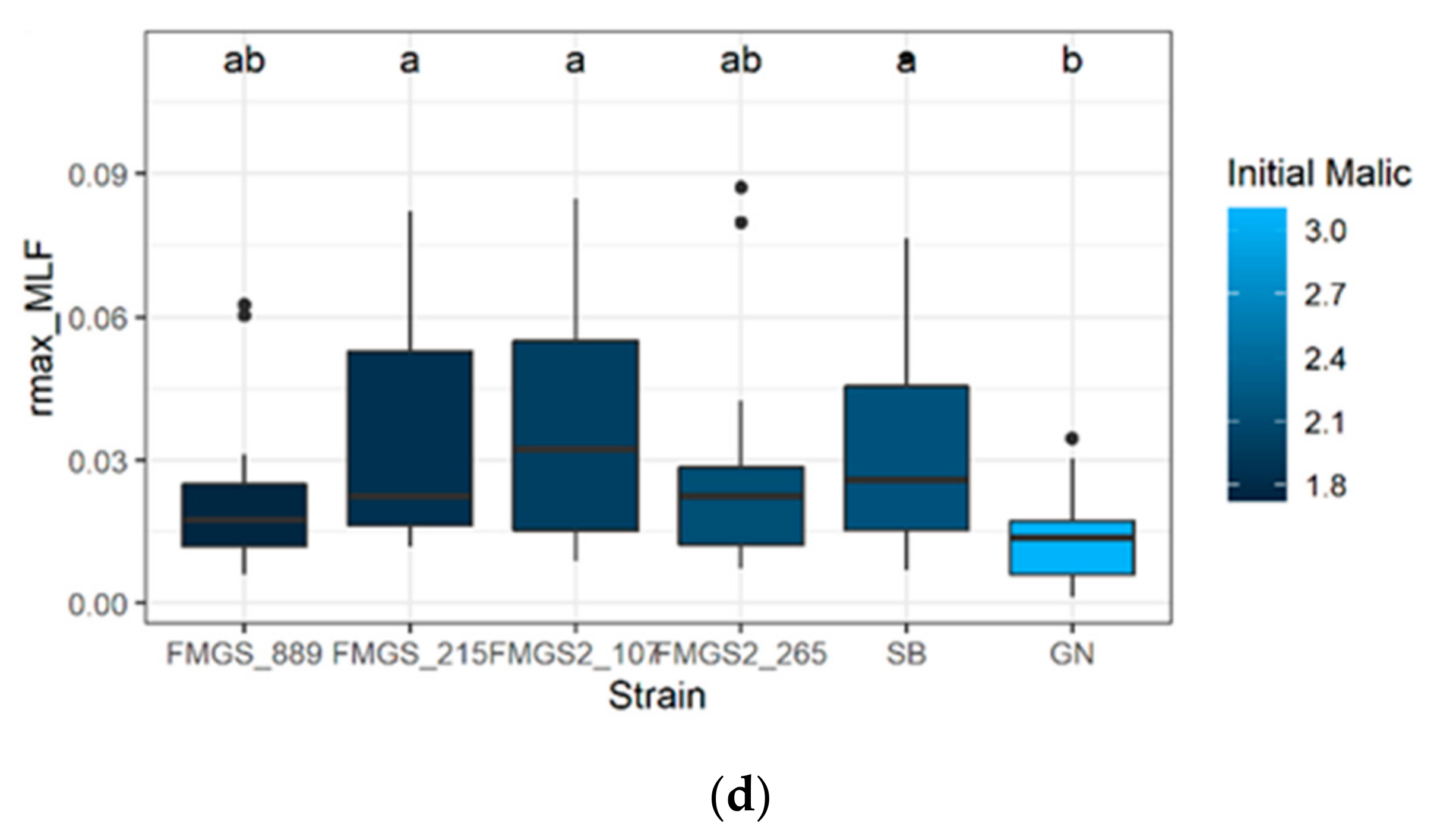
3.6. Ongoing of Malolactic Fermentation
4. Discussion
4.1. Assesment of a Wide MAS Program for Improving a Complex Trait
4.2. Possible Causes of Incomplete QTLs Penetrance
4.3. Outlier Strains for Specific Enological Applications. Lowering the Acidity of Rich Malic Wines and Shortening the MLF of Red Wines
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sicard, D.; Legras, J.L. Bread, beer and wine: Yeast domestication in the Saccharomyces sensu stricto complex. C. R. Biol. 2011, 334, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Rainieri, S.; Pretorius, I.I.S. Selection and improvement of wine yeasts. Ann. Microbiol. 2000, 50, 15–31. [Google Scholar]
- Marullo, P.; Dubourdieu, D. Yeast selection for wine flavour modulation. In Managing Wine Quality Vol 2; Reynolds, A.G., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2010; pp. 296–320. ISBN 9781845697983. [Google Scholar]
- Gammacurta, M.; Marchand, S.; Albertin, W.; Moine, V.; De Revel, G. Impact of yeast strain on ester levels and fruity aroma persistence during aging of bordeaux red wines. J. Agric. Food Chem. 2014, 62, 5378–5389. [Google Scholar] [CrossRef] [PubMed]
- Marchal, A.; Marullo, P.; Durand, C.; Moine, V.; Dubourdieu, D. Fermentative Conditions Modulating Sweetness in Dry Wines: Genetics and Environmental Factors Influencing the Expression Level of the Saccharomyces cerevisiae HSP12 Gene. J. Agric. Food Chem. 2015, 63, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Murat, M.-L.; Masneuf, I.; Darriet, P.; Lavigne, V.; Tominaga, T.; Dubourdieu, D. Effect of Saccharomyces cerevisiae Yeast Strains on the Liberation of Volatile Thiols in Sauvignon blanc Wine. Am. J. Enol. Vitic. 2001, 52, 136–139. [Google Scholar]
- Julien, A.; Roustan, J.L.; Dulau, L.; Sablayrolles, J.M. Comparison of nitrogen and oxygen demands of enological yeasts: Technological consequences. Am. J. Enol. Vitic. 2000, 51, 215–222. [Google Scholar]
- Berthels, N.J.; Otero, R.R.C.; Bauer, F.F.; Thevelein, J.M.; Pretorius, I.S. Discrepancy in glucose and fructose utilisation during fermentation by Saccharomyces cerevisiae wine yeast strains. FEMS Yeast Res. 2004, 4, 683–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marullo, P.; Mansour, C.; Dufour, M.; Albertin, W.; Sicard, D.; Bely, M.; Dubourdieu, D. Genetic improvement of thermo-tolerance in wine Saccharomyces cerevisiae strains by a backcross approach. FEMS Yeast Res. 2009, 9, 1148–1160. [Google Scholar] [CrossRef] [Green Version]
- Esteve-Zarzoso, B.; Gostıncar, A.; Bobet, R.; Uruburn, F.; Querol, A. Selection and molecular characterization of wine yeasts isolated from the “El Penedès” area (Spain). Food Microbiol. 2000, 17, 553–562. [Google Scholar] [CrossRef]
- Romano, P.; Soli, M.G.; Suzzi, G. Improvement of a wine Saccharomyces cerevisiae strain by a breeding program. Appl. Environ. Microbiol. 1985, 50, 1064–1067. [Google Scholar] [CrossRef] [Green Version]
- Marullo, P.; Bely, M.; Masneuf-Pomarède, I.; Pons, M.; Aigle, M.; Dubourdieu, D.; Masneuf-Pomarede, I.; Pons, M.; Aigle, M.; Dubourdieu, D. Breeding strategies for combining fermentative qualities and reducing off-flavor production in a wine yeast model. FEMS Yeast Res. 2006, 6, 268–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vero, L.; Solieri, L.; Giudici, P. Evolution-based strategy to generate non-genetically modified organisms Saccharomyces cerevisiae strains impaired in sulfate assimilation pathway. Lett. Appl. Microbiol. 2011, 53, 572–575. [Google Scholar] [CrossRef]
- Steensels, J.; Snoek, T.; Meersman, E.; Nicolino, M.P.; Voordeckers, K.; Verstrepen, K.J. Improving industrial yeast strains: Exploiting natural and artificial diversity. FEMS Microbiol. Rev. 2014, 38, 1–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marullo, P.; Durrens, P.; Peltier, E.; Bernard, M.; Mansour, C.; Dubourdieu, D. Natural allelic variations of Saccharomyces cerevisiae impact stuck fermentation due to the combined effect of ethanol and temperature; a QTL-mapping study. BMC Genom. 2019, 20, 680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marullo, P.; Aigle, M.; Bely, M.; Masneuf-Pomarède, I.; Durrens, P.; Dubourdieu, D.; Yvert, G. Single QTL mapping and nucleotide-level resolution of a physiologic trait in wine Saccharomyces cerevisiae strains. FEMS Yeast Res. 2007, 7, 941–952. [Google Scholar] [CrossRef] [Green Version]
- Zimmer, A.; Durand, C.; Loira, N.; Durrens, P.; Sherman, D.J.; Marullo, P. QTL dissection of lag phase in wine fermentation reveals a new translocation responsible for Saccharomyces cerevisiae adaptation to sulfite. PLoS ONE 2014, 9, e86298. [Google Scholar] [CrossRef] [Green Version]
- Martí-Raga, M.; Peltier, E.; Mas, A.; Beltran, G.; Marullo, P. Genetic Causes of Phenotypic Adaptation to the Second Fermentation of Sparkling Wines in Saccharomyces cerevisiae. G3 Genes Genomes Genet. 2017, 7, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Ambroset, C.; Petit, M.; Brion, C.; Sanchez, I.; Delobel, P.; Guerin, C.; Chiapello, H.; Nicolas, P.; Bigey, F.; Dequin, S.; et al. Deciphering the Molecular Basis of Wine Yeast Fermentation Traits Using a Combined Genetic and Genomic Approach. G3 Genes Genomes Genetics 2011, 1, 263–281. [Google Scholar] [CrossRef] [Green Version]
- Steyer, D.; Ambroset, C.; Brion, C.; Claudel, P.; Delobel, P.; Sanchez, I.; Erny, C.; Blondin, B.; Karst, F.; Legras, J.-L. QTL mapping of the production of wine aroma compounds by yeast. BMC Genom. 2012, 13, 573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cubillos, F.A.; Parts, L.; Salinas, F.; Bergström, A.; Scovacricchi, E.; Zia, A.; Illingworth, C.J.R.; Mustonen, V.; Ibstedt, S.; Warringer, J.; et al. High-resolution mapping of complex traits with a four-parent advanced intercross yeast population. Genetics 2013, 195, 1141–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jara, M.; Cubillos, F.A.; García, V.; Salinas, F.; Aguilera, O.; Liti, G.; Martínez, C. Mapping genetic variants underlying differences in the central nitrogen metabolism in fermenter yeasts. PLoS ONE 2014, 9, e86533. [Google Scholar] [CrossRef] [Green Version]
- Salinas, F.; Cubillos, F.A.; Soto, D.; Garcia, V.; Bergström, A.; Warringer, J.; Ganga, M.A.; Louis, E.J.; Liti, G.; Martinez, C. The genetic basis of natural variation in oenological traits in Saccharomyces cerevisiae. PLoS ONE 2012, 7, e49640. [Google Scholar] [CrossRef] [Green Version]
- Brice, C.; Sanchez, I.; Bigey, F.; Legras, J.-L.; Blondin, B. A genetic approach of wine yeast fermentation capacity in nitrogen-starvation reveals the key role of nitrogen signaling. BMC Genom. 2014, 15, 495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roncoroni, M.; Santiago, M.; Hooks, D.O.; Moroney, S.; Harsch, M.J.; Lee, S.A.; Richards, K.D.; Nicolau, L.; Gardner, R.C. The yeast IRC7 gene encodes a b-lyase responsible for production of the varietal thiol 4-mercapto-4-methylpentan-2-one in wine. Food Microbiol. 2011, 28, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Roncoroni, M.; Gardner, R.C. MET2 affects production of hydrogen sulfide during wine fermentation. Appl. Microbiol. Biotechnol. 2014, 98, 7125–7135. [Google Scholar] [CrossRef] [PubMed]
- Noble, J.; Sanchez, I.; Blondin, B. Identification of new Saccharomyces cerevisiae variants of the MET2 and SKP2 genes controlling the sulfur assimilation pathway and the production of undesirable sulfur compounds during alcoholic fermentation. Microb. Cell Fact. 2015, 14, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordente, A.G.; Heinrich, A.; Pretorius, I.S.; Swiegers, J.H. Isolation of sulfite reductase variants of a commercial wine yeast with significantly reduced hydrogen sulfide production. FEMS Yeast Res. 2009, 9, 446–459. [Google Scholar] [CrossRef] [Green Version]
- Linderholm, A.L.; Findleton, C.L.; Kumar, G.; Hong, Y.; Bisson, L.F. Identification of genes affecting hydrogen sulfide formation in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2008, 74, 1418–1427. [Google Scholar] [CrossRef] [Green Version]
- Linderholm, A.; Dietzel, K.; Hirst, M.; Bisson, L.F. Identification of MET10-932 and Characterization as an Allele Reducing Hydrogen Sulfide Formation in Wine Strains of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2010, 76, 7699–7707. [Google Scholar] [CrossRef] [Green Version]
- Peltier, E.; Friedrich, A.; Schacherer, J.; Marullo, P. Quantitative Trait Nucleotides Impacting the Technological Performances of Industrial Saccharomyces cerevisiae Strains. Front. Genet. 2019, 10, 683. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Crouch, J.H. Marker-Assisted Selection in Plant Breeding: From Publications to Practice. Crop Sci. 2008, 48, 391–407. [Google Scholar] [CrossRef] [Green Version]
- Dekkers, J.C.M. Marker-assisted selection for commercial crossbred performance1. J. Anim. Sci. 2007, 85, 2104–2114. [Google Scholar] [CrossRef]
- Yao, F.-J.; Lu, L.-X.; Wang, P.; Fang, M.; Zhang, Y.-M.; Chen, Y.; Zhang, W.-T.; Kong, X.-H.; Lu, J.; Honda, Y. Development of a Molecular Marker for Fruiting Body Pattern in Auricularia auricula-judae. Mycobiology 2018, 46, 72–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufour, M.; Zimmer, A.; Thibon, C.; Marullo, P. Enhancement of volatile thiol release of Saccharomyces cerevisiae strains using molecular breeding. Appl. Microbiol. Biotechnol. 2013, 97, 5893–5905. [Google Scholar] [CrossRef] [PubMed]
- Marullo, P.; Yvert, G.; Bely, M.; Aigle, M.; Dubourdieu, D. Efficient use of DNA molecular markers to construct industrial yeast strains. FEMS Yeast Res. 2007, 7, 1295–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javelot, C.; Girard, P.; Colonna-Ceccaldi, B.; Vladescu, B. Introduction of terpene-producing ability in a wine strain of Saccharomyces cerevisiae. J. Biotechnol. 1991, 21, 239–251. [Google Scholar] [CrossRef]
- Blondin, B.; Noble, J.; Sanchez, I. Méthode de Contrôle de la Production de Sulfites, D’hydrogène Sulfureux et D’acétaldéhyde par des Levures. European Patent EP2807247B1.
- Mackay, T.F.C. The Genetic Architecture of Quantitative Traits. Annu. Rev. Genet. 2001, 35, 303–339. [Google Scholar] [CrossRef]
- Lecomte, L.; Duffé, P.; Buret, M.; Servin, B.; Hospital, F.; Causse, M. Marker-assisted introgression of five QTLs controlling fruit quality traits into three tomato lines revealed interactions between QTLs and genetic backgrounds. Theor. Appl. Genet. 2004, 109, 658–668. [Google Scholar] [CrossRef] [PubMed]
- Volschenk, H.; Viljoen, M.; Grobler, J.; Petzold, B.; Bauer, F.; Subden, R.E.; Young, R.A.; Lonvaud, A.; Denayrolles, M.; van Vuuren, H.J. Engineering pathways for malate degradation in Saccharomyces cerevisiae. Nat. Biotechnol. 1997, 15, 253–257. [Google Scholar] [CrossRef]
- Redzepovic, S.; Orlic, S.; Majdak, A.; Kozina, B.; Volschenk, H.; Viljoen-Bloom, M. Differential malic acid degradation by selected strains of Saccharomyces during alcoholic fermentation. Int. J. Food Microbiol. 2003, 83, 49–61. [Google Scholar] [CrossRef]
- Delcourt, F.; Taillandier, P.; Vidal, F.; Strehaiano, P. Influence of pH, malic acid and glucose concentrations on malic acid consumption by Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 1995, 43, 321–324. [Google Scholar] [CrossRef]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud, A. Handbook of Enology, Vol. 1: The Microbiology of Wine and Vinifications, 2nd ed.; Wiley: Hoboken, NJ, USA, 2006; ISBN 9780470010341. [Google Scholar]
- Volschenk, H.; van Vuuren, H.J.J.; Viljoen–Bloom, M. Malo-ethanolic fermentation in Saccharomyces and Schizosaccharomyces. Curr. Genet. 2003, 43, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Peltier, E.; Vion, C.; Abo Saada, O.; Friedrich, A.; Schacherer, J.; Marullo, P. Flor yeast rewire the central carbon metabolism during wine alcoholic fermentation. bioRxiv 2021. [Google Scholar] [CrossRef]
- Peltier, E.; Marullo, P.; Coulon, J. Levures Œnologiques Permettant le Contrôle de la Teneur en Acide des Vins et Méthodes de Sélection de Telles Levures. Patent FR 33390, 2020. [Google Scholar]
- Peltier, E.; Sharma, V.; Raga, M.M.; Roncoroni, M.; Bernard, M.; Gibon, Y.; Marullo, P.; Jiranek, V.; Gibon, Y.; Marullo, P. Dissection of the molecular bases of genotype x environment interactions: A study of phenotypic plasticity of Saccharomyces cerevisiae in grape juices. BMC Genom. 2018, 19, 772. [Google Scholar] [CrossRef] [Green Version]
- Chernova, M.; Albertin, W.; Durrens, P.; Guichoux, E.; Sherman, D.J.; Masneuf-Pomarede, I.; Marullo, P. Many interspecific chromosomal introgressions are highly prevalent in Holarctic Saccharomyces uvarum strains found in human-related fermentations. Yeast 2018, 35, 141–156. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, S.; Ziaugra, L.; Tabbaa, D. SNP genotyping using the sequenom massARRAY iPLEX Platform. Curr. Protoc. Hum. Genet. 2009, 60, 2.12. 1–2.12. 18. [Google Scholar] [CrossRef] [PubMed]
- Den Dunnen, J.T.; Antonarakis, S.E. Mutation nomenclature extensions and suggestions to describe complex mutations: A discussion. Hum. Mutat. 2000, 15, 7–12. [Google Scholar] [CrossRef]
- Peltier, E.; Bernard, M.; Trujillo, M.; Prodhomme, D.D.; Barbe, J.-C.; Gibon, Y.; Marullo, P. Wine yeast phenomics: A standardized fermentation method for assessing quantitative traits of Saccharomyces cerevisiae strains in enological conditions. PLoS ONE 2018, 13, e0190094. [Google Scholar] [CrossRef] [Green Version]
- Stitt, M.; McC, L.R.; Gerhardt, R.; Heldt, H.W. Metabolite Levels in Specific Cells and Subcellular Compartements of Plant Leaves. Methods Enzymol. 1989, 174, 518–552. [Google Scholar]
- R Development Core Team, R. R: A Language and Environment for Statistical Computing. R Found. Stat. Comput. 2011, 1, 409. [Google Scholar]
- Taillandier, P.; Gilis, M.; Strehaiano, P.; Taillandier, P.; Gilis, M.; Strehaiano, P. Deacidification by Schizosaccharomyces: Interactions with Saccharomyces. J. Biotechnol. 1995, 40, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Yéramian, N.; Chaya, C.; Suárez Lepe, J.A. L-(-)-malic acid production by Saccharomyces spp. during the alcoholic fermentation of wine (1). J. Agric. Food Chem. 2007, 55, 912–919. [Google Scholar] [CrossRef] [PubMed]
- Vilanova, M.; Ugliano, M.; Varela, C.; Siebert, T.; Pretorius, I.S.; Henschke, P.A. Assimilable nitrogen utilisation and production of volatile and non-volatile compounds in chemically defined medium by Saccharomyces cerevisiae wine yeasts. Appl. Microbiol. Biotechnol. 2007, 77, 145–157. [Google Scholar] [CrossRef] [Green Version]
- Mortimer, R.K.; Romano, P.; Suzzi, G.; Polsinellif, M. Genome Renewal: A New Phenomenon Revealed From a Genetic Study of 43 Strains of Saccharomyces cerevisiae Derived From Natural Fermentation of Grape Musts. YEAST 1994, 10, 1543–1552. [Google Scholar] [CrossRef] [PubMed]
- Giudici, P.; Solieri, L.; Pulvirenti, A.M.; Cassanelli, S. Strategies and perspectives for genetic improvement of wine yeasts. Appl. Microbiol. Biotechnol. 2005, 66, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Salmon, J. l-Malic-acid permeation in resting cells of anaerobically grown Saccharomyces cerevisiae. Biochim. Biophys. Acta Biomembr. 1987, 901, 30–34. [Google Scholar] [CrossRef]
- Vailiant, H.; Formisyn, P.; Gerbaux, V. Malolactic fermentation of wine: Study of the influence of some physico-chemical factors by experimental design assays. J. Appl. Bacteriol. 1995, 79, 640–650. [Google Scholar] [CrossRef]
- Versari, A.; Parpinello, G.P.; Cattaneo, M. Leuconostoc oenos and malolactic fermentation in wine: A review-Journal of Industrial Microbiology and Biotechnology. J. Ind. Microbiol. Biotechnol. 1999, 23, 447–455. [Google Scholar] [CrossRef]
- Hospital, F.; Charcosset, A. Marker-assisted introgression of quantitative trait loci. Genetics 1997, 147, 1469–1485. [Google Scholar] [CrossRef]
- Bouchez, A.A.; Hospital, F.F.; Causse, M.; Gallais, A.A.; Charcosset, A. Marker-assisted introgression of favorable alleles at quantitative trait loci between maize elite lines. Genetics 2002, 162, 1945–1959. [Google Scholar] [CrossRef]
- Bloom, J.S.; Kotenko, I.; Sadhu, M.J.; Treusch, S.; Albert, F.W.; Kruglyak, L. Genetic interactions contribute less than additive effects to quantitative trait variation in yeast. Nat. Commun. 2015, 6, 8712. [Google Scholar] [CrossRef] [Green Version]
- Steinmetz, L.M.; Sinha, H.; Richards, D.R.; Spiegelman, J.I.; Oefner, P.J.; McCusker, J.H.; Davis, R.W. Dissecting the architecture of a quantitative trait locus in yeast. Nature 2002, 416, 326–330. [Google Scholar] [CrossRef]
- Sinha, H.; David, L.; Pascon, R.C.; Clauder-Münster, S.; Krishnakumar, S.; Nguyen, M.; Shi, G.; Dean, J.; Davis, R.W.; Oefner, P.J.; et al. Sequential Elimination of Major-Effect Contributors Identifies Additional Quantitative Trait Loci Conditioning High-Temperature Growth in Yeast. Genetics 2008, 180, 1661–1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liti, G.; Louis, E.J. Advances in Quantitative Trait Analysis in Yeast. PLoS Genet. 2012, 8, e1002912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnett, D.G.; Rebetzke, G.J.; Spielmeyer, W. Strategies for efficient implementation of molecular markers in wheat breeding. Mol. Breed. 2005, 15, 75–85. [Google Scholar] [CrossRef]
- Fatichenti, F.; Farris, G.A.; Deiana, P.; Ceccarelli, S. Malic acid production and consumption by selected of Saccharomyces cerevisiae under anaerobic and aerobic conditions. Appl. Microbiol. Biotechnol. 1984, 19, 427–429. [Google Scholar] [CrossRef]
- Ramon-Portugal, F.; Seiller, I.; Taillandier, P.; Favarel, J.L.; Nepveu, F.; Strehaiano, P. Kinetics of Production and Consumption of Organic Acids during Alcoholic Fermentation by Saccharomyces cerevisae. Food Technol. Biotechnol. 1999, 37, 235–240. [Google Scholar]
- Kunkee, R.E. Some roles of malic acid in the malolactic fermentation in wine making. FEMS Microbiol. Lett. 1991, 88, 55–71. [Google Scholar] [CrossRef]
- Volschenk, H.; van Vuuren, H.J.J.; Viljoen-Bloom, M. Malic Acid in Wine: Origin, Function and Metabolism during Vinification. S. Afr. J. Enol. Vitic. 2006, 27. [Google Scholar] [CrossRef] [Green Version]
- Jeppsson, H.; Yu, S.; Hahn-Hägerdal, B. Xylulose and glucose fermentation by Saccharomyces cerevisiae in chemostat culture. Appl. Environ. Microbiol. 1996, 62, 1705–1709. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Description | Reference | WDCM-791 (CRB ISVV, Bordeaux) |
|---|---|---|---|
| M2 | parental strain, a meiotic spore clone from Enoferm M2 (Lallemand, Canada) | [26] | CRBO L2010 |
| F15 | parental strain, a meiotic spore clone from Zymaflore F15 (Laffort, France) | [26] | CRBO L2011 |
| GN | parental strain, a meiotic spore clone from Zymaflore VL1 (Laffort, France) | [48] | CRBO L2002 |
| SB | parental strain, a meiotic spore clone from Zymaflore BO213 (Laffort, France) | [48] | CRBO L2001 |
| M2xF15 | F1-hybrid (M2 x F15) | [26] | CRBO L2005 |
| SBxGN | F1-hybrid (SB x GN) | [48] | CRBO L2003 |
| GS-28b | meiotic clone of SBxGN | this work | CRBO L2012 |
| GS-41b | meiotic clone of SBxGN | this work | CRBO L2013 |
| FM-8d | meiotic clone of M2xF15 | this work | CRBO L2014 |
| FMGS-1 | F1-hybrid (GS-28b x FM-8d) | this work | CRBO L2015 |
| FMGS-1-647 | meiotic clone of FMGS-1, malic acid consumer strain | this work | CRBO L2016 |
| FMGS-2 | F1-hybrid (GS-41b x FMGS-1-647) | this work | CRBO L2017 |
| FMGS-1-889 | meiotic clone of FMGS-1, malic acid consumer strain | this work | CRBO L2018 |
| FMGS-1-215 | meiotic clone of FMGS-1, Malic acid consumer strain | this work | CRBO L2019 |
| FMGS-2-107 | meiotic clone of FMGS-2, Malic acid consumer strain | this work | CRBO L2020 |
| FMGS-2-265 | meiotic clone of FMGS-2, Malic acid consumer strain | this work | CRBO L2021 |
| Populations | Number | Mean | Variance | Quantiles | % of Individuals with a MAC Above | |||
|---|---|---|---|---|---|---|---|---|
| 50% | 75% | 95% | 65% | 70% | ||||
| Starters | 31 | 28.3 | 33.3 | 28.2 | 31.5 | 36.5 | no | no |
| pop SBxGN | 93 | 31.0 | 206.6 | 28.6 | 36.9 | 58.5 | 2.2 | 2.2 |
| pop M2xF15 | 94 | 17.8 | 138.9 | 18.6 | 24.8 | 34.1 | no | no |
| random FMGS-1 | 50 | 52.6 | 58.6 | 51.7 | 57.2 | 65.7 | 12.0 | no |
| selected FMGS-1 a | 30 | 53.7 | 77.0 | 54.1 | 59.2 | 65.8 | 10 | no |
| random FMGS2-1 | 50 | 48.7 | 130.4 | 49.0 | 56.6 | 64.3 | 4.0 | 2.0 |
| selected FMGS2-2 b | 17 | 53.7 | 121.2 | 54.2 | 57.1 | 72.3 | 17.6 | 11.7 |
| Population | II_661 | IV_31 | IV_360 | IV_414 | VII_427 | VII_480 | XI_382 | XI_631 | XII_53 | XV_491 | XV_1052 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene validated | PYC2 | PNC1 | PMA1 | MAE1 | SDH2 | PTC5 | |||||
| SBxGN (n = 94) | 3.87 | 4.22 | ns | ns | ns | 8.20 | 2.97 | ns | 4.45 | 7.61 | ns |
| FMGSs (n = 154) | - | 7.14 | - | - | ns | - | - | - | - | ns | 4.92 |
| FMGS-1 (n = 82) | - | 9.31 | - | - | ns | - | - | - | ns | ns | 9.74 |
| FMGS-2 (n = 72) | ns | 6.41 | ns | ns | ns | - | ns | 3.10 | - | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vion, C.; Peltier, E.; Bernard, M.; Muro, M.; Marullo, P. Marker Assisted Selection of Malic-Consuming Saccharomyces cerevisiae Strains for Winemaking. Efficiency and Limits of a QTL’s Driven Breeding Program. J. Fungi 2021, 7, 304. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040304
Vion C, Peltier E, Bernard M, Muro M, Marullo P. Marker Assisted Selection of Malic-Consuming Saccharomyces cerevisiae Strains for Winemaking. Efficiency and Limits of a QTL’s Driven Breeding Program. Journal of Fungi. 2021; 7(4):304. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040304
Chicago/Turabian StyleVion, Charlotte, Emilien Peltier, Margaux Bernard, Maitena Muro, and Philippe Marullo. 2021. "Marker Assisted Selection of Malic-Consuming Saccharomyces cerevisiae Strains for Winemaking. Efficiency and Limits of a QTL’s Driven Breeding Program" Journal of Fungi 7, no. 4: 304. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7040304