1. Introduction
The date palm (
Phoenix dactylifera L.) is an economically important palm species grown in the arid, sub-tropical, and tropical regions. It has the tendency to tolerate not only the harsh extremely high desert temperature but also resistant to drought and salinity. Globally, Saudi Arabia stands second for dates production after Egypt. Their production in Saudi Arabia according to United Nations Food and Agriculture statistics reached 1.54 million tons/year during 2019 that is 16.97% of the total world’s dates production [
1]. However, Saudi Arabia being the major producer of dates is conscious about the loss of date palm fruits due to the infestations of various pests. The reduced yield that mainly leads to low production is mainly because of degradation of date palms due to trunk pests [
2,
3,
4], and the associated date palm fruit pests’ infestations [
5,
6]. The
Oligonychus afrasiaticus (Acari: Tetranychidae), is an economically important pest of the fruit of date palms in Saudi Arabia [
6], along with some other countries including Algeria, Egypt, Iran, Iraq, Israel, Libya, Mauritania, Oman, Tunisia, and Yemen [
7,
8,
9,
10,
11,
12,
13,
14,
15,
16]. The environmental conditions prevailing in these areas greatly favored the growth and development of
O. afrasiaticus on date palm fruits [
17]. At the advent of fruit formation,
O. afrasiaticus infestations start on the date palms by spinning the web around the bunches of dates [
18]. These webs collect the dust along with the exuviae from their different developmental stages resulting dusty appearance of the infested bunches of dates. As a consequence, they start egg laying with their full potential and their population multiply logarithmically. These webs remain intact till later stages of the development of dates resulting decline in growth and development of date palm fruit. Such damages have caused subsequent crop losses and rendering their fruits unfit for human consumption [
2,
5,
19].
The control of
O. afrasiaticus involved the application of synthetic acaricides. However, the residues associated with acaricides are harmful for the consumers and also responsible for environmental pollution. Furthermore, the indiscriminate use of synthetic acaricides is restricted due to the deleterious effects on non-target animals. These shortcomings have led to find other efficient, safe, and environmentally friendlier alternatives to control
O. afrasiaticus populations. An important component of biological control agents is the use of entomopathogenic fungi. These naturally occurring entomopathogenic fungi are considered important substitutes for acaricides. Most of the entomopathogenic fungi belong to Entomophthorales and Hypocreales. According to an estimate, around 700 taxa of fungi with more than 13,000 isolates of entomopathogenic fungal species isolated from 1300 different hosts have been reported to infect pests including aphids [
20], beetles [
21,
22], cockroaches [
23], grass-hoppers [
24], mosquitoes, lepidopterous [
25,
26], termites [
27], thrips [
28], weevils [
29], whitefly [
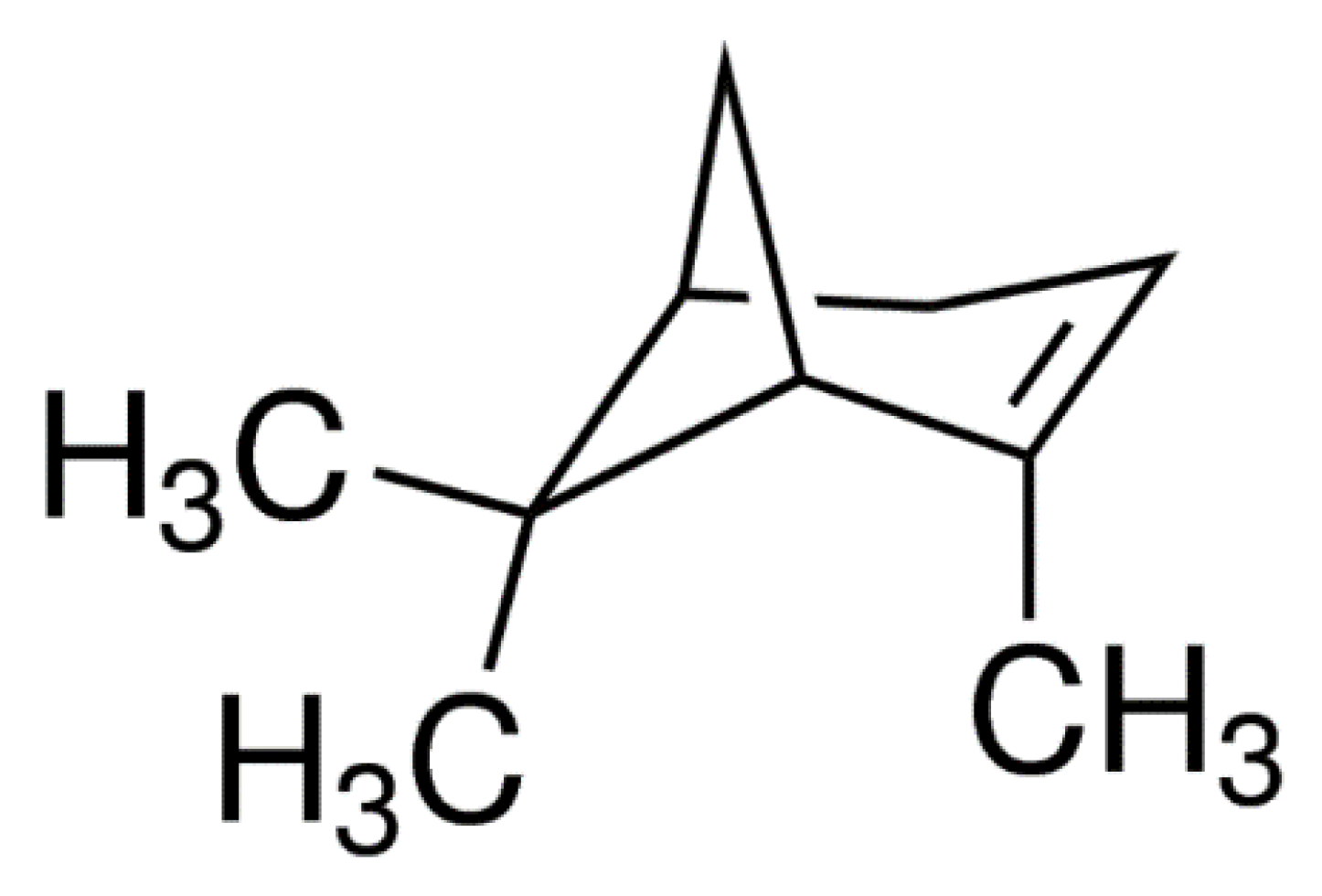
30]. However, the slow mode of action of fungal isolates along with reduced virulence over the period of time is found to be the main hurdle in the way of their commercialization. In this regard, the concept of incorporation of plant secondary metabolites into the conidial suspension is gaining momentum in the field of eco-friendly pest management. In this regard, a polyphenolic terpene hydrocarbon (+)-α-Pinene is very much pertinent to the current study due to known toxicity against various pests including head lice [
31],
Sitophilus zeamais Motschulsky [
32],
Spodoptera litura and
Achaea janata [
33]. In the past, numerous studies have been conducted on the compatibility and joint action of conidial suspensions with plant-based active natural products [
34,
35,
36], and their findings revealed enhanced joint treatment response. The addition of plant secondary metabolites for developing biorational pest management technologies is gaining interest nowadays because of numerous modes of action such as cuticle disruption, molting inhibition, respiratory failure, growth, and fecundity reduction. In addition, joint application of plant secondary metabolites with conidial suspensions is advantageous over synthetic pesticides because they are environmentally friendly and delay the development of resistance among the target pest populations.
In the past, little information is available on the joint action of plant secondary metabolites with conidial suspension against
O. afrasiaticus [
6,
37]. These preliminary studies showed the tremendous potential of plant secondary compounds and emphasized on the exploration of more isolates and plant secondary metabolites to develop the compatible formulation for the eco-friendly control of
O. afrasiaticus. The current study is mainly aimed at (1) evaluating the compatibility of conidial suspensions with (+)-α-Pinene by compatibility assays; (2) testing the acaricidal potential of a selected plant secondary metabolite (+)-α-Pinene, and
B. bassiana ARSEF 8465 by concentration mortality response bioassays; (3) calculating the joint toxicity index to sort out the nature of their interaction; (4) exploring the treatment effects on the regulation of defense-related enzymes controlling physiological mechanisms of date palm dust mites, in order to develop a synergistic interaction with improved efficacy for the effective management of
O. afrasiaticus populations infesting date palm fruits.
4. Discussion
The development of effective and stable pest management option involves the extensive toxin-pathogen compatibility and target host time- and concentration-dependent mortality response experimentation. This study clearly showed that the addition of (+)-α-Pinene found to be compatible with the B. bassiana ARSEF 8465, and their dual application found to be an attractive alternative approach to circumvent slow mortality criticism by producing strong synergistic interaction, which tremendously induced the target host defense-related enzymatic regulation, and ultimately increase the mortality of O. afrasiaticus.
Naturally-occurring entomopathogenic fungi are well-known to infect the populations of pests [
43,
45,
46,
47]. The infection of the conidial suspensions of
B. bassiana ARSEF 8465 evaluated here against date palm dust mites exhibited the highest LC
50 value (LC
50 = 19.16 mg/mL). Furthermore, none of the tested conidial concentration able to impart 100% mortality, and their recorded cumulative mortality even at the highest concentration found to be <75%. The slow mortality response revealed here in the current study by the fungal conidial suspensions is in agreement with previous studies aiming to screen the most virulent isolate against various pest species [
5,
29,
45]. These findings enabled us to suggest that slow fungal conidial mortality drawback could be overcome on one hand by reprogramming fungal virulence controlling metalloproteinases [
48], or by incorporating compatible plant-based secondary compounds [
6], for effective pest management.
Plant secondary metabolites are untapped chemicals reservoir currently emerged as an attractive management option for agricultural pests due to the advancement in analytical chemistry and least risk to the environment. The studied (+)-α-Pinene is a polyphenolic terpene hydrocarbon with known toxicity against various pests including head lice [
31],
Sitophilus zeamais Motschulsky [
32],
Spodoptera litura and
Achaea janata [
33]. The concentration-dependent time mortality response of
O. afrasiaticus exhibited enhanced mortality response over time and concentrations in the current study. These results coincide with previous research findings [
32] that strengthened our findings by revealing that α-Pinene is a promising compound for developing a novel effective formulation against
S. zeamais Motschulsky. However, the low toxicity response (LC
50 = 3.41 mg/mL) recorded in this study is in compliance with previous studies against date palm dust mites exposed with some other plant secondary metabolites including 1-chlorooctadecane [
6], and phytol [
37], and these findings suggested the necessity to develop compatible toxin-pathogen synergistic interaction. Although the (+)-α-Pinene differed significantly from the control, vegetative growth and percent conidial viability of
B. bassiana ARSEF 8465 against all tested concentration was high, ranging from 95.00 to 98.20% (conidial germination), and 74.20 to 85.40 mm (vegetative growth). The biological index score (BI ranged from 79–95) of
B. bassiana ARSEF 8465 calculated against each tested concentration of (+)-α-Pinene demonstrated that their joint application is suitable to develop synergistic interaction against
O. afrasiaticus populations.
For successful
O. afrasiaticus populations control in the date palm plantations, the importance of compatible toxin-pathogen interaction is highly applicable to facilitate the development of eco-friendly products. Our results revealed that pairing of
B. bassiana ARSEF 8465 with (+)-α-Pinene in different proportions exhibited different responses among exposed date palm dust mites. However, the bioassay schemes with a higher proportion of (+)-α-Pinene contribute to enhanced mortality response, which ultimately lead to synergistic interaction. The enhanced mortality response among the treatments with a higher proportion of compounds recorded in the current study coincides with previous studies reporting the incorporation of phytol within the conidial suspensions tremendously enhance the combined treatment effect by revealing a very high score of joint toxicity index (691) [
37]. The enhancement of treatment efficacy through the joint application of
B. bassiana and commercial biopesticide against Colorado potato beetle larvae further strengthened the current findings [
49], and allowed us to demonstrate that combined treatment combination added advantage by reducing mortality time and enhancing percent mortality. Similarly, the Scheme IV bioassays (80% (+)-α-Pinene: 20%
B. bassiana ARSEF 8465 Conidia) depicted strong synergistic interaction (joint toxicity index = 755). These results are further supported by a previous study [
50], which showed that the degree of synergy in terms of joint toxicity index increased, as the share of plant secondary metabolites increased in the joint application treatment comprising of conidial suspensions and plant secondary metabolites.
The joint application of conidial suspension and chemical compounds, which mainly aimed at improving the treatment effect might also lead to antagonistic interaction that limits the combination of the applied mixture [
51]. In our study, the joint application under the Scheme I (20% (+)-α-Pinene: 80%
B. bassiana ARSEF 8465 Conidia) showed enhanced mortality response compared with
O. afrasiaticus exposed to (+)-α-Pinene or
B. bassiana ARSEF 8465 sole application, but without synergistic interaction. However, both the tested fractions interacted antagonistically (joint toxicity index = 58) that might because of low proportion of (+)-α-Pinene. The current interaction data agree with Al-mazra’awi at al. [
52], who showed that joint application of
B. bassiana and neem tree extracts although showed high mortality, but revealed antagonistic interaction. Taken together, these findings suggested that careful synthesis of joint application is critically important to shift antagonistic interaction towards strong synergistic interaction to develop strong toxin pathogen synergy for effective and efficient management of agricultural pests.
Antioxidant defense system is important to scavenge the reactive oxygen species (ROS) generated within the target host as a result of the exposure to toxin/pathogen. The enzymes including SOD and CAT that prevent the accumulation of ROS explored in the current study depicted significantly high activities compared to control treatment after 96 h post-exposure. The increase in CAT and SOD activities is consistent with previous studies [
37,
43], revealing that the most potent treatment led to strong activities of antioxidant enzymes, which ultimately tremendously declined at the lateral stage of infection. These results are further strengthened by previous research investigations, revealing that increase in stress whether biotic or abiotic tremendously induced the activities of enzymes regulating antioxidant defense mechanism [
43,
53]. Furthermore, the degree of response of antioxidant enzymes modulated with the intensity of the stress to prevent ROS accumulation and maintain balance.
Host defense mechanisms in the context of toxin-pathogen interaction studies are very much pertinent because these interactions act as drivers to impart physiological stress among target hosts. Obtained results indicated that synergistic effect greatly induced the relative GST and AchE activities compared to control treatment. The enhanced defense-related enzymatic response revealed especially from the interaction treatments (synergistic interactions) with a higher proportion of % (+)-α-Pinene represents date palm dust mites response, which ultimately leads to strong physiological stress also confirmed by previous studies conducted on the host defense mechanism [
54,
55,
56,
57]. Furthermore, our results on the physiological impacts of (+)-α-Pinene and
B. bassiana ARSEF 8465 conidia against date palm dust mites are consistent with those of Zibaee and Bandani [
58], who reported significantly enhanced GST response among
Eurygaster integriceps Puton exposed with
Artemisia annua. These findings allowed us to suggest that the enzymes involved in defense mechanism play a pivotal role as these mechanisms utilize most of the host energy to defend the toxin or pathogen. The host could withstand these circumstances to certain limits and these conditions ultimately led the target host towards death.





{kind=link}