
Towards a Natural Classification of Hyphodontia Sensu Lato and the Trait Evolution of Basidiocarps within Hymenochaetales (Basidiomycota)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
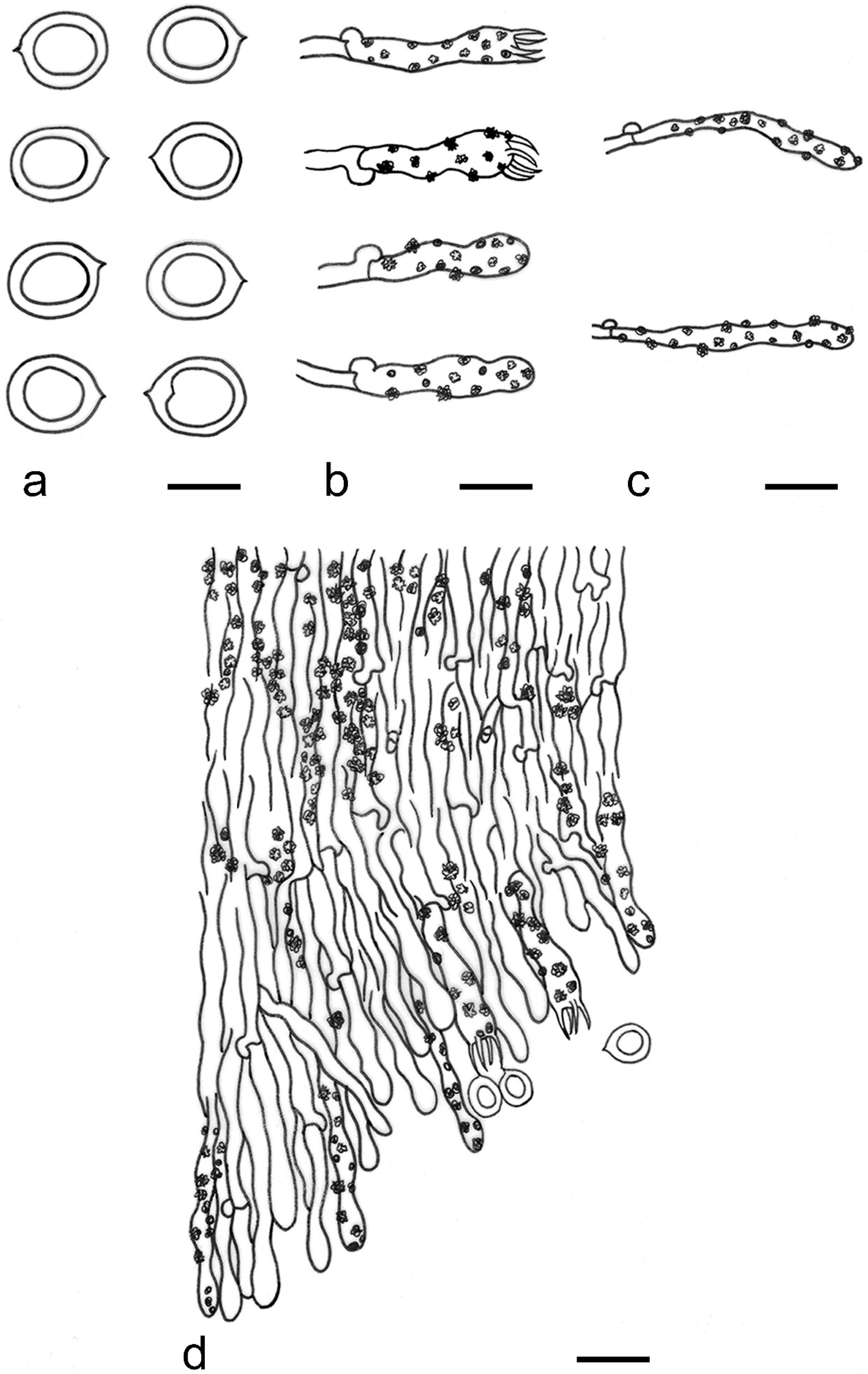{kind=link}
{kind=link}
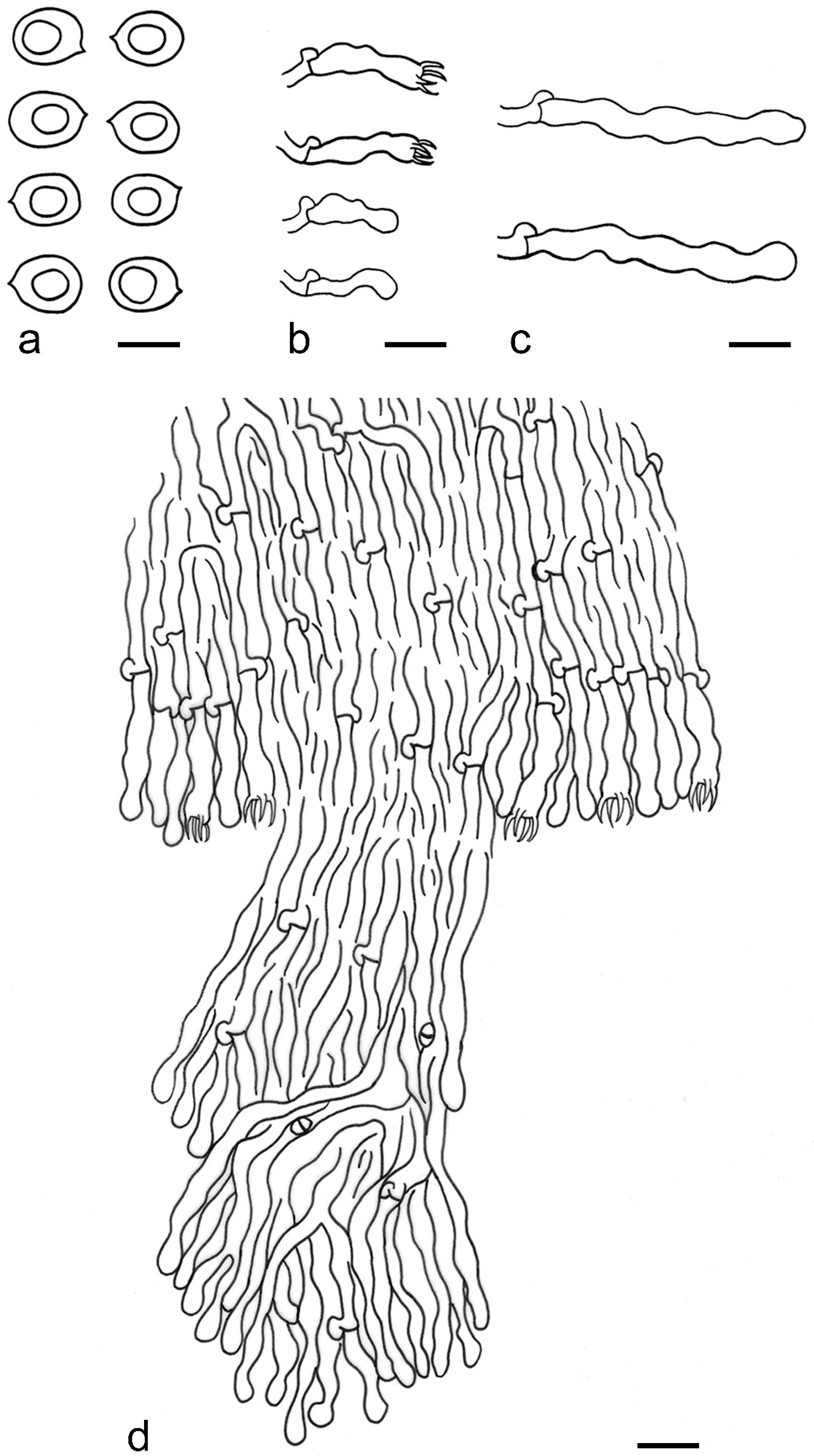{kind=link}
{kind=link}
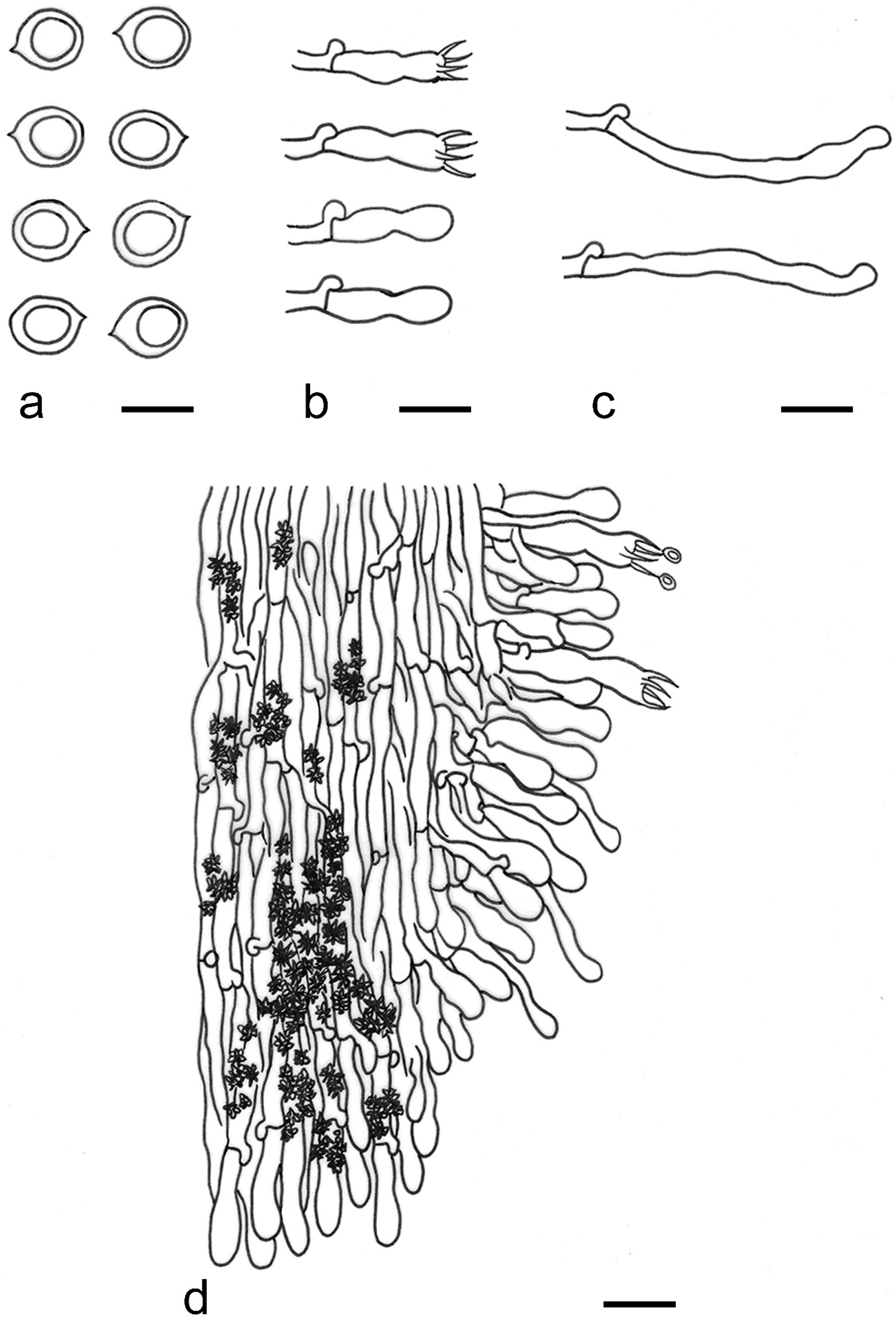{kind=link}
{kind=link}
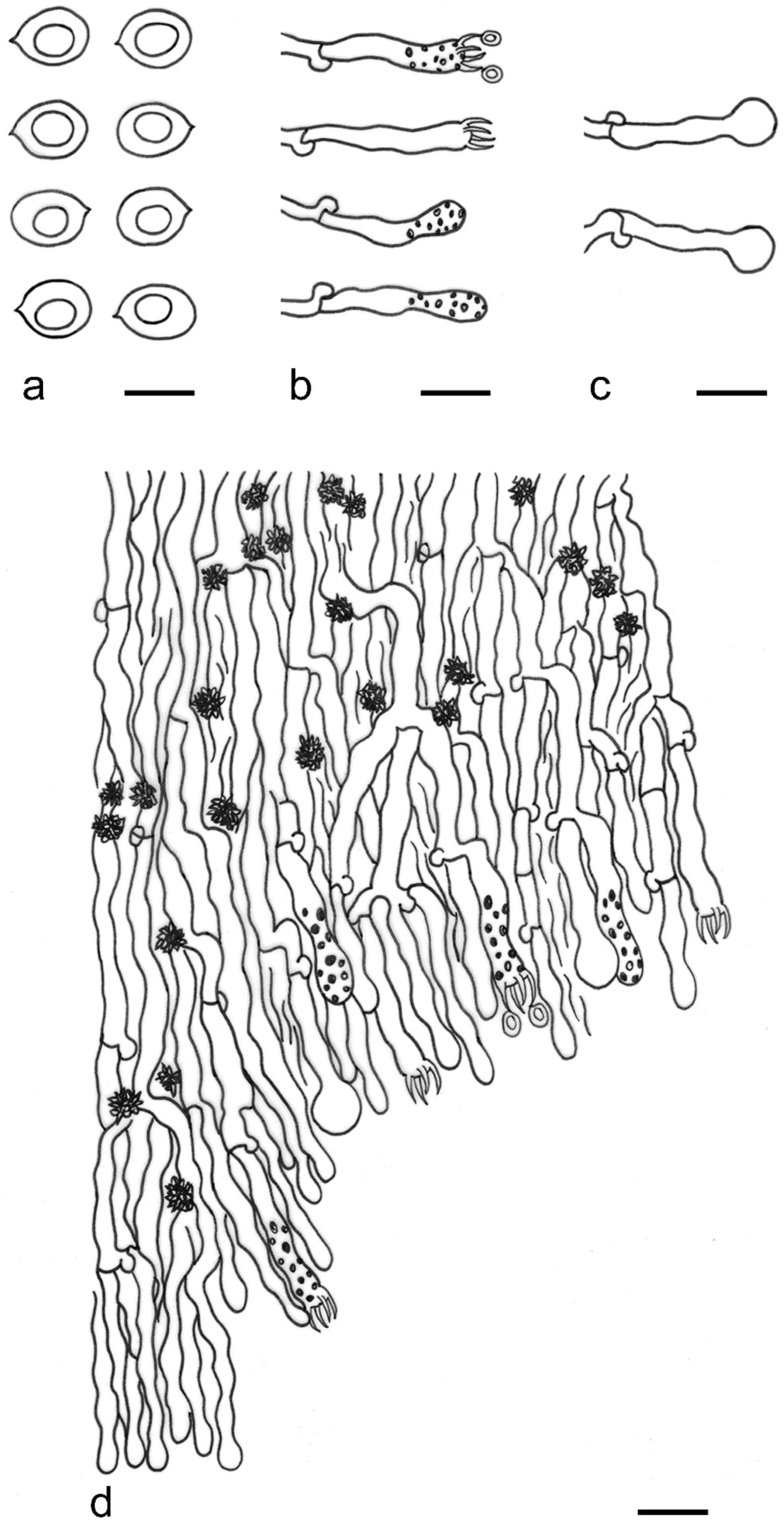{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Voucher Deposition
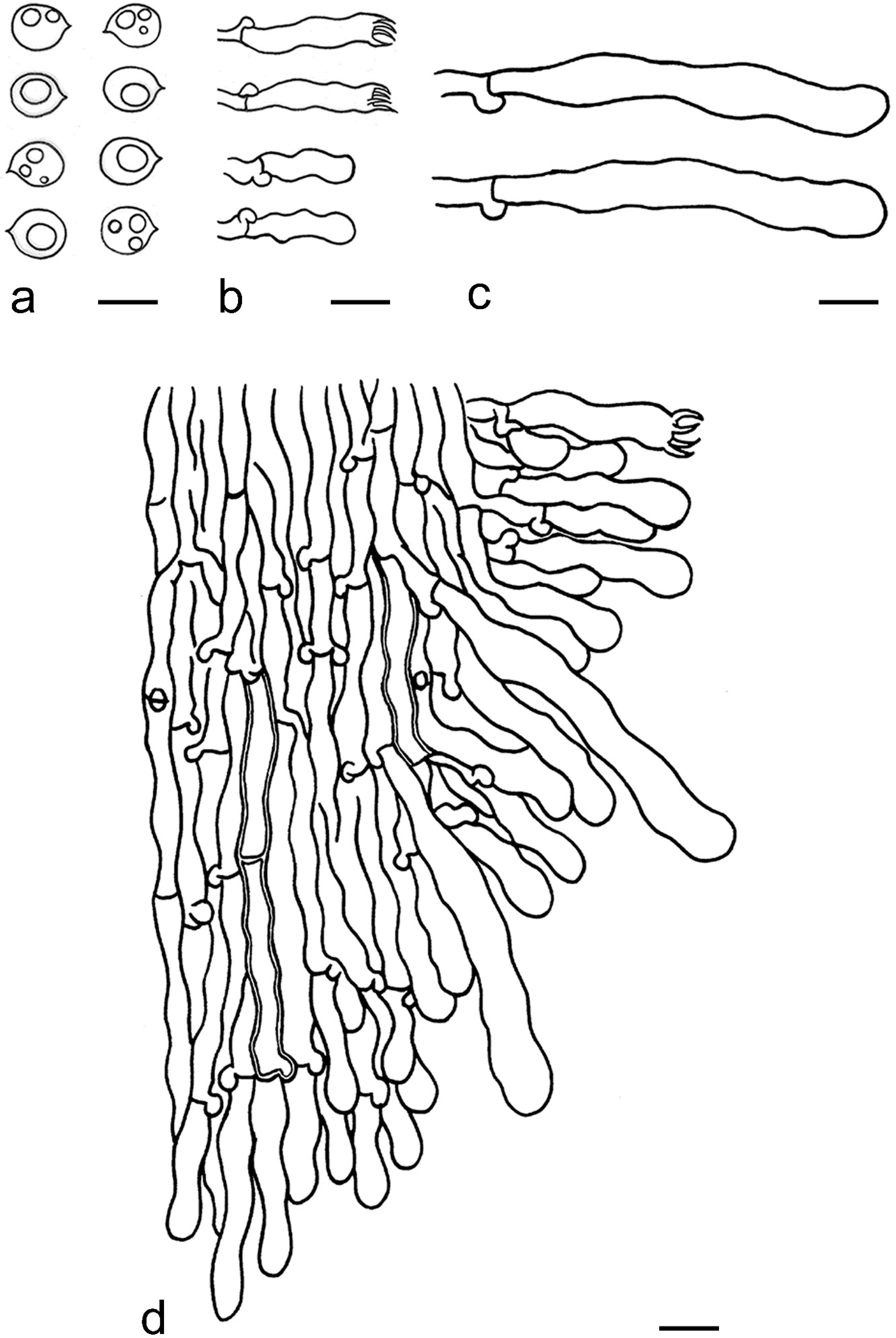
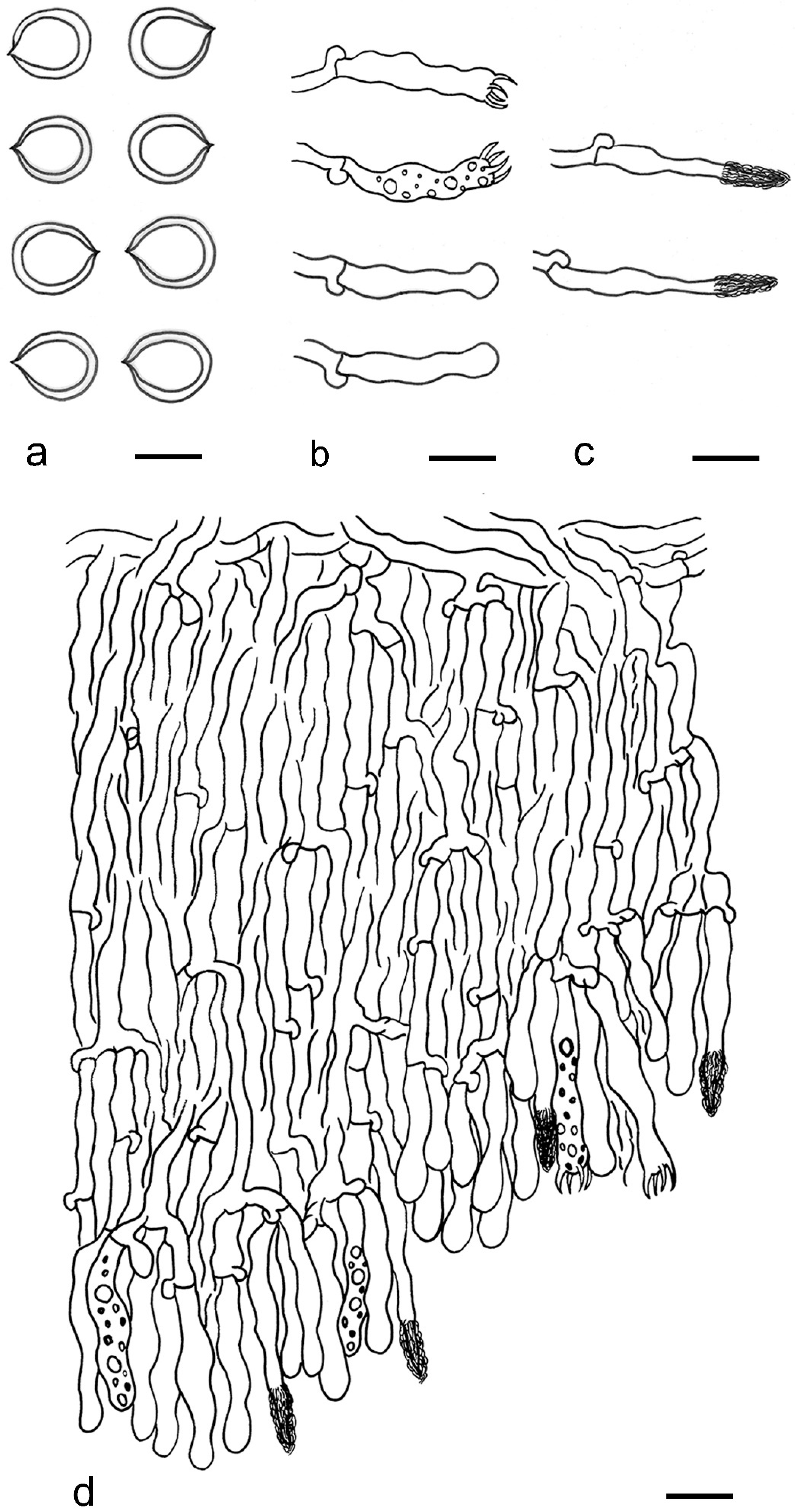
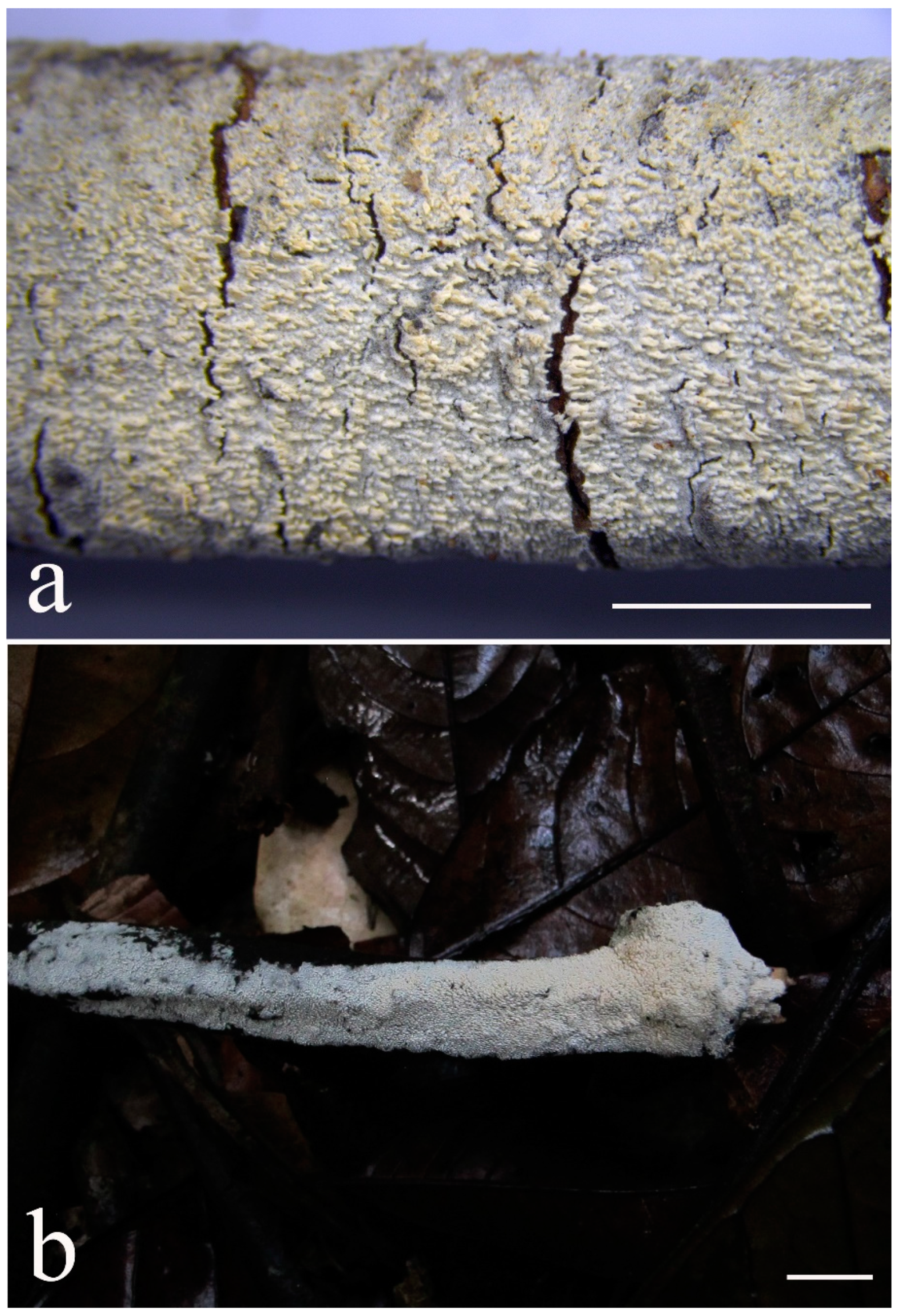
2.2. Morphological Examination
2.3. Molecular Sequencing
2.4. Phylogenetic Analyses
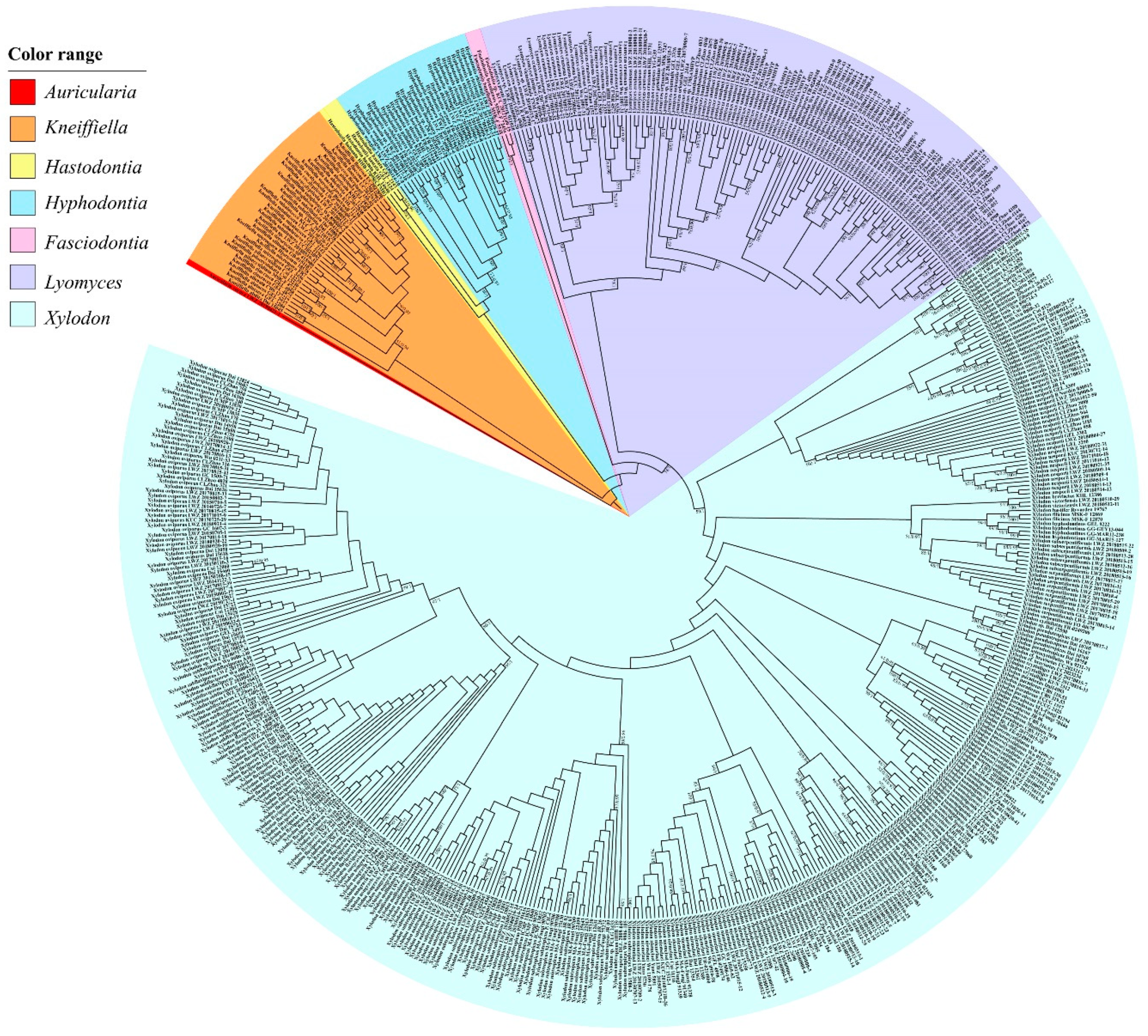
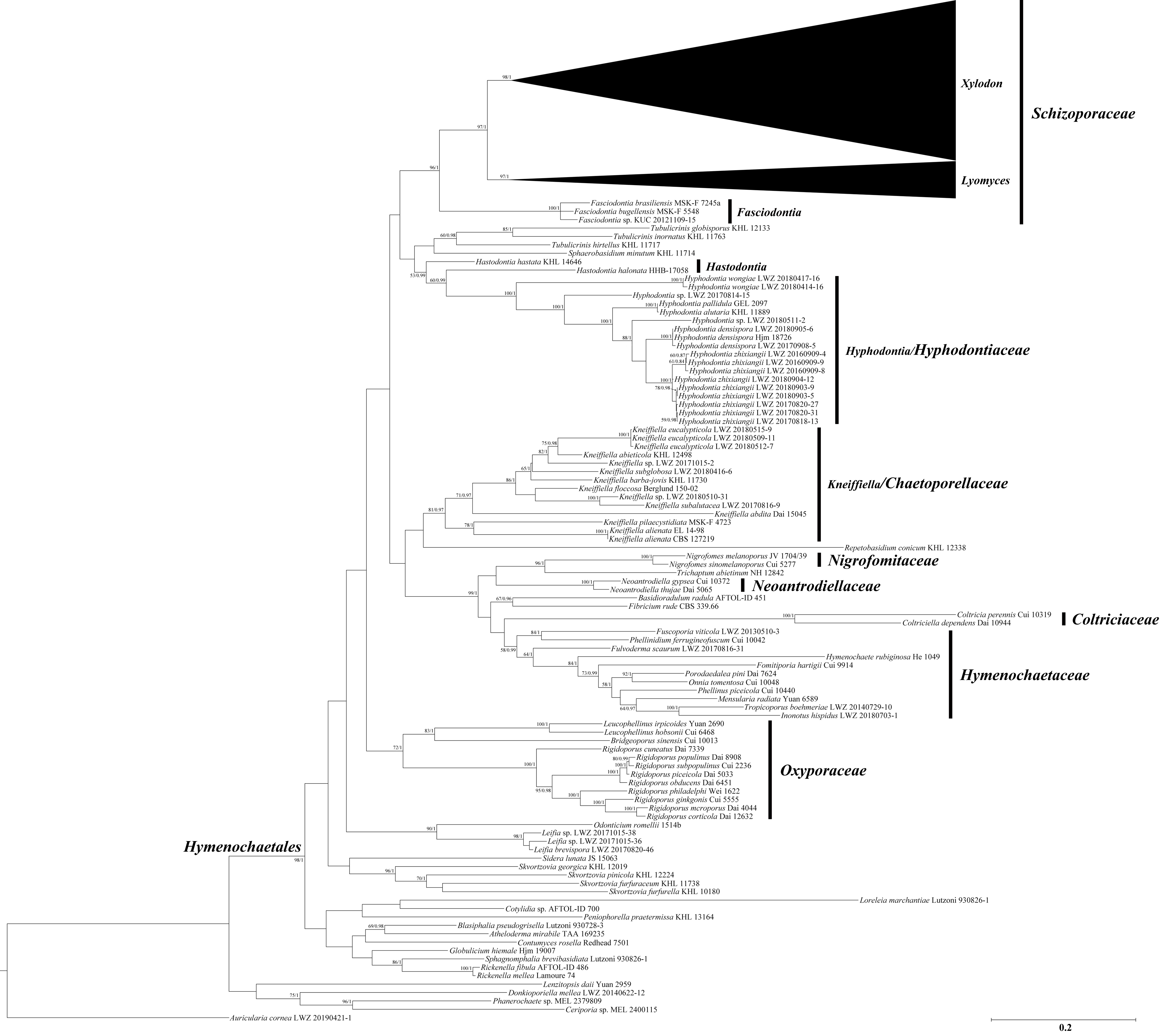
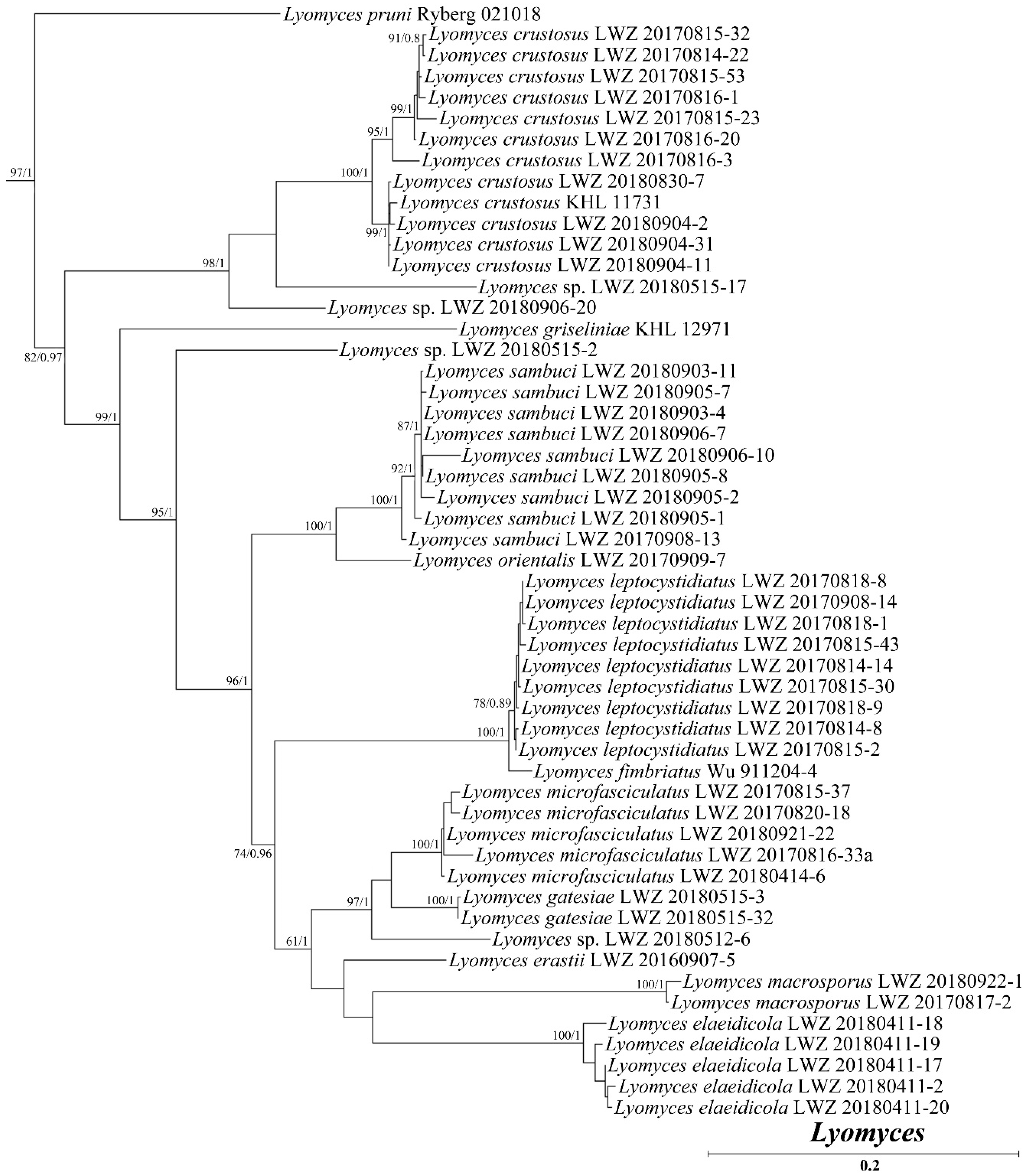
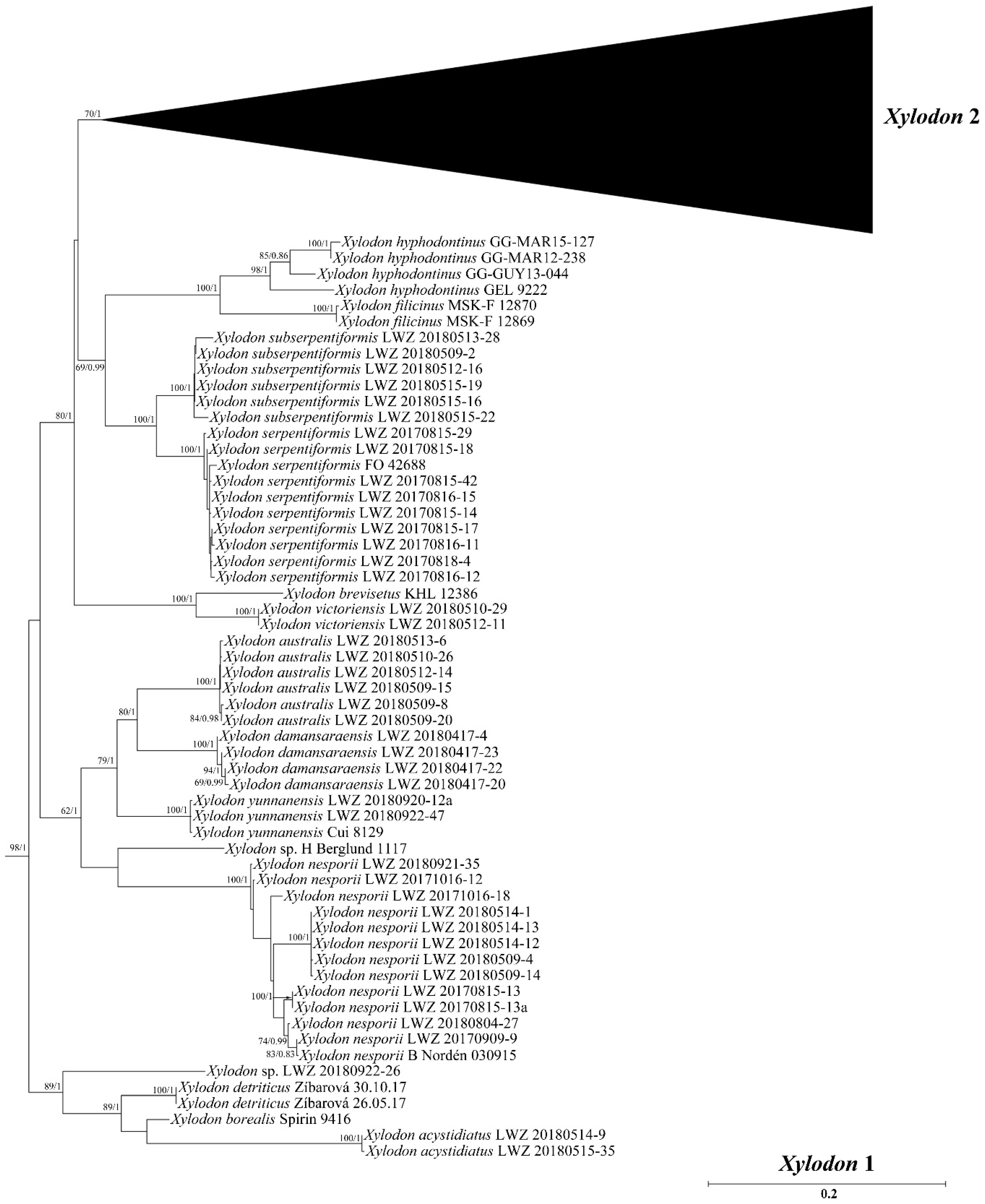
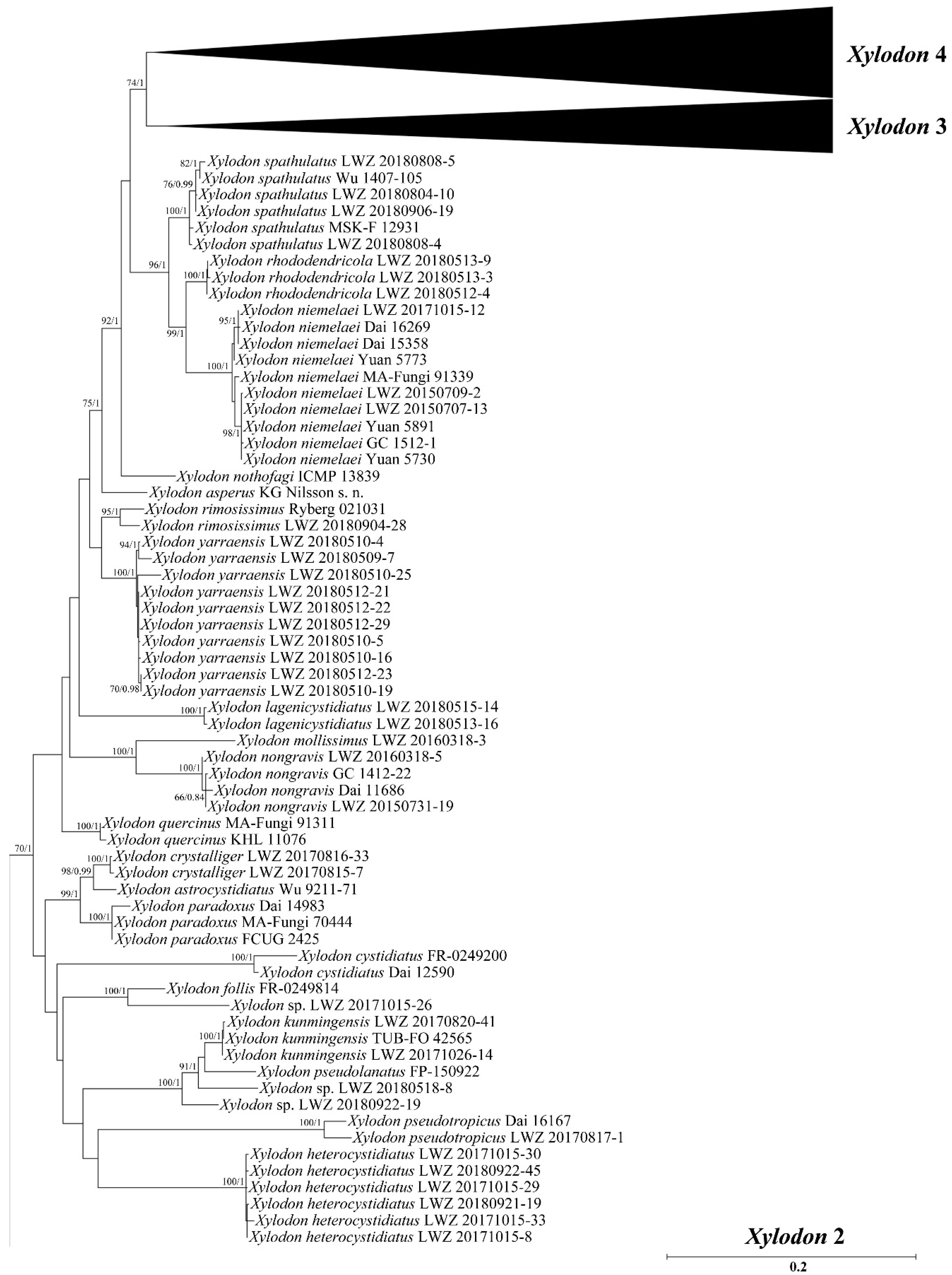
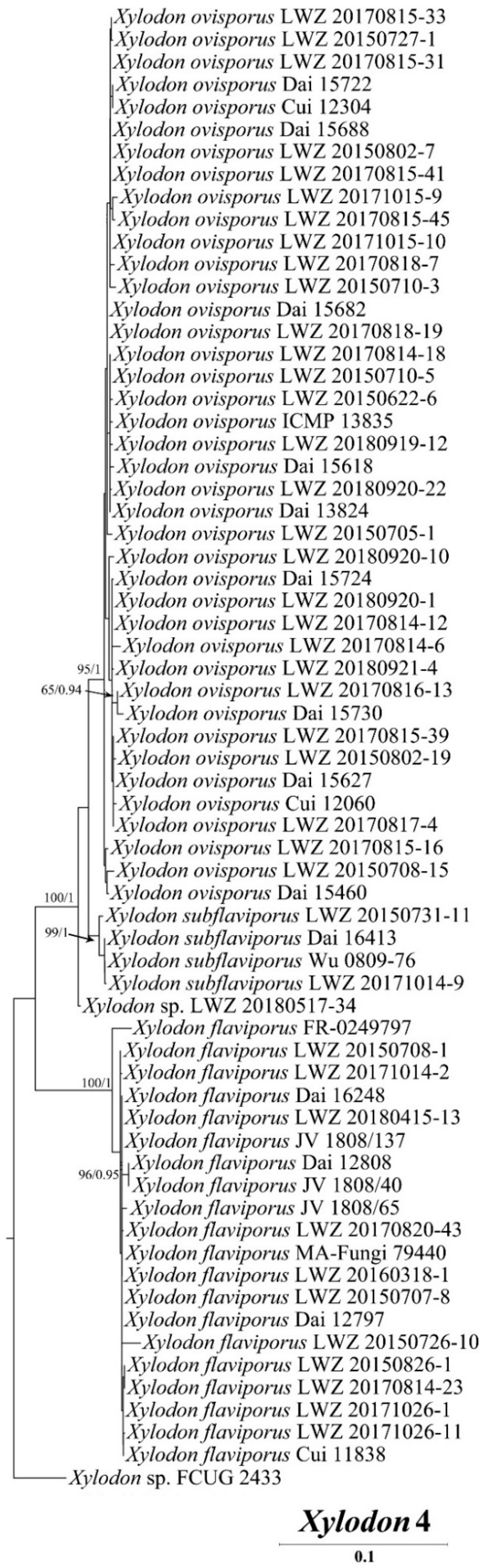
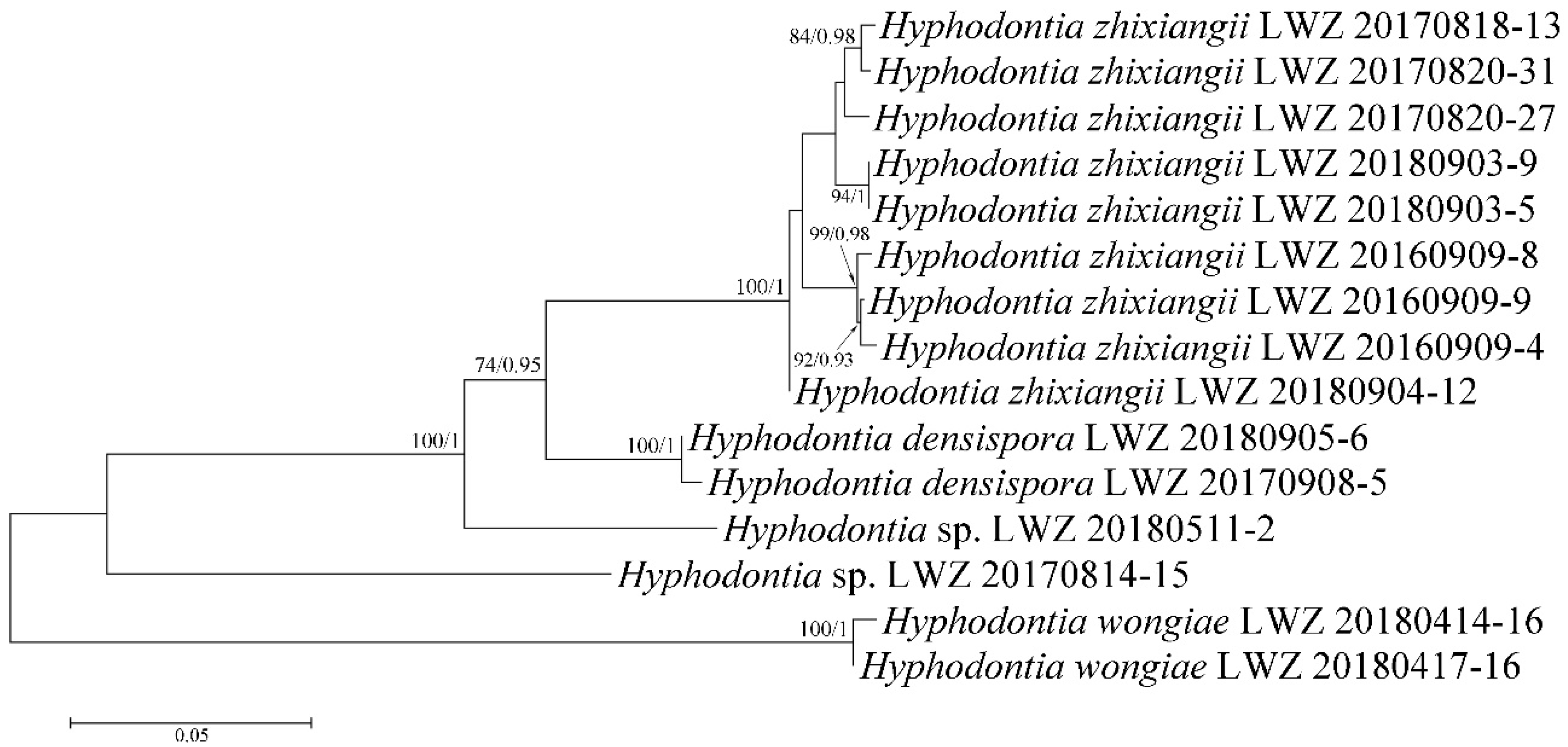
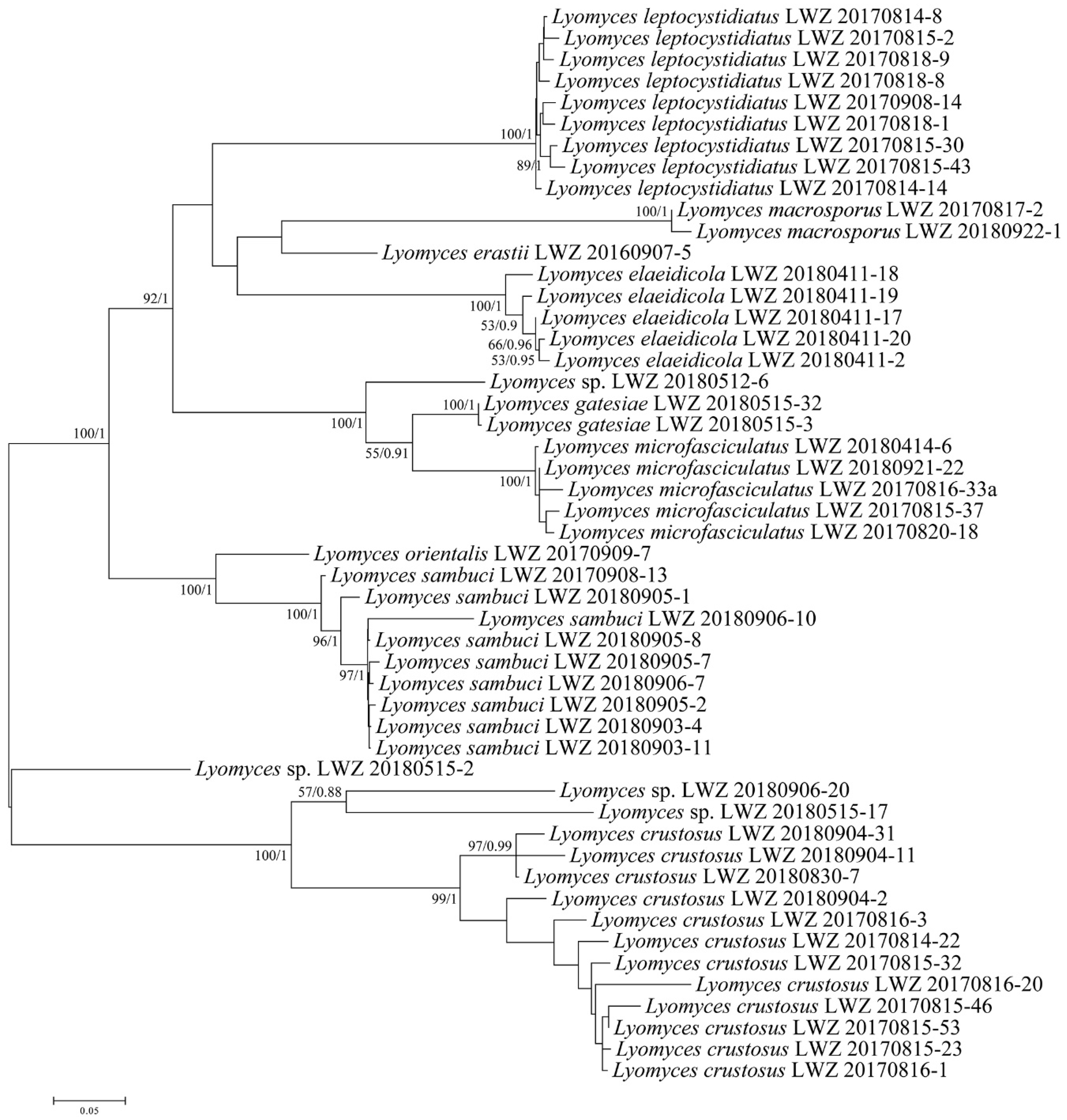
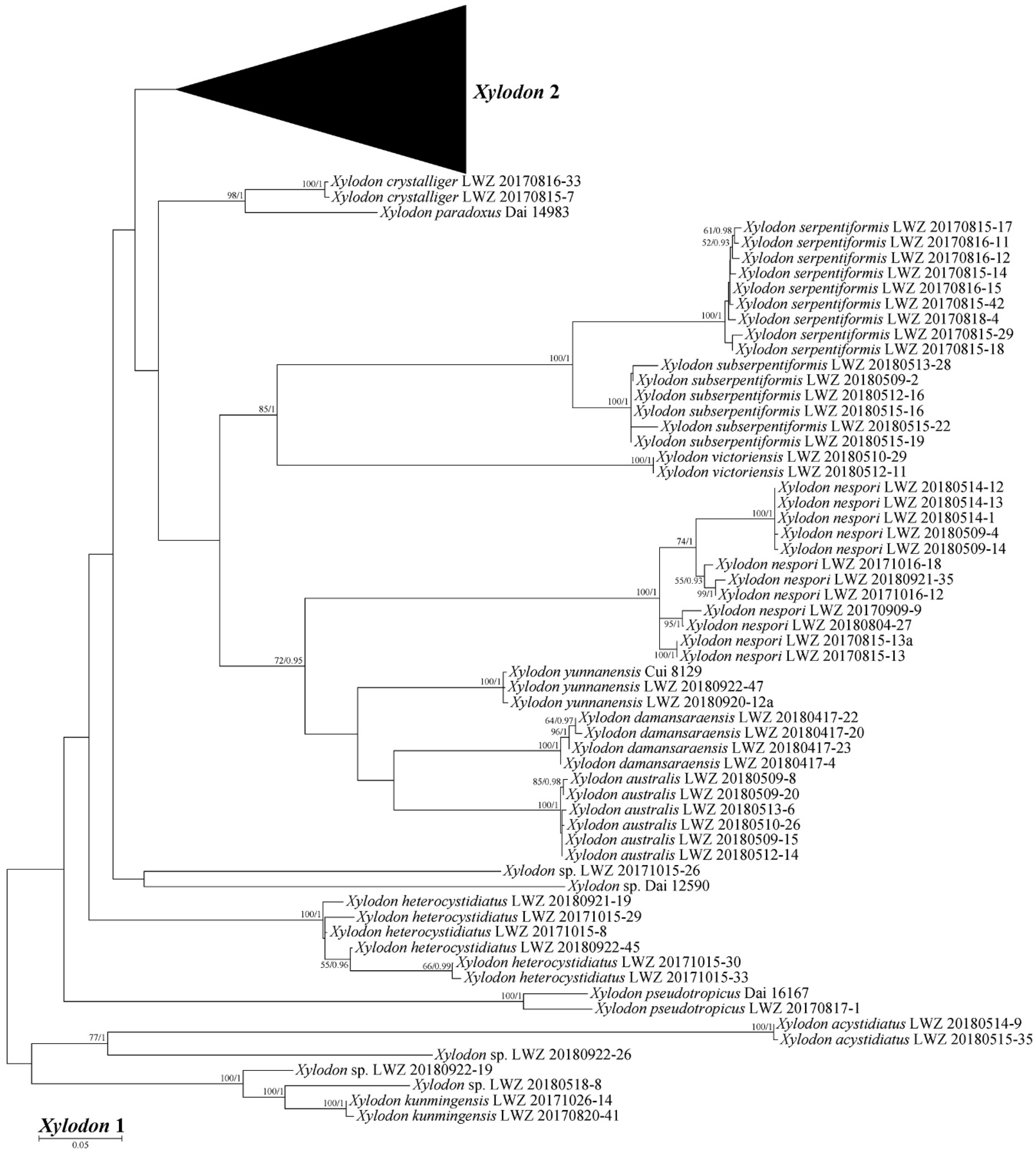
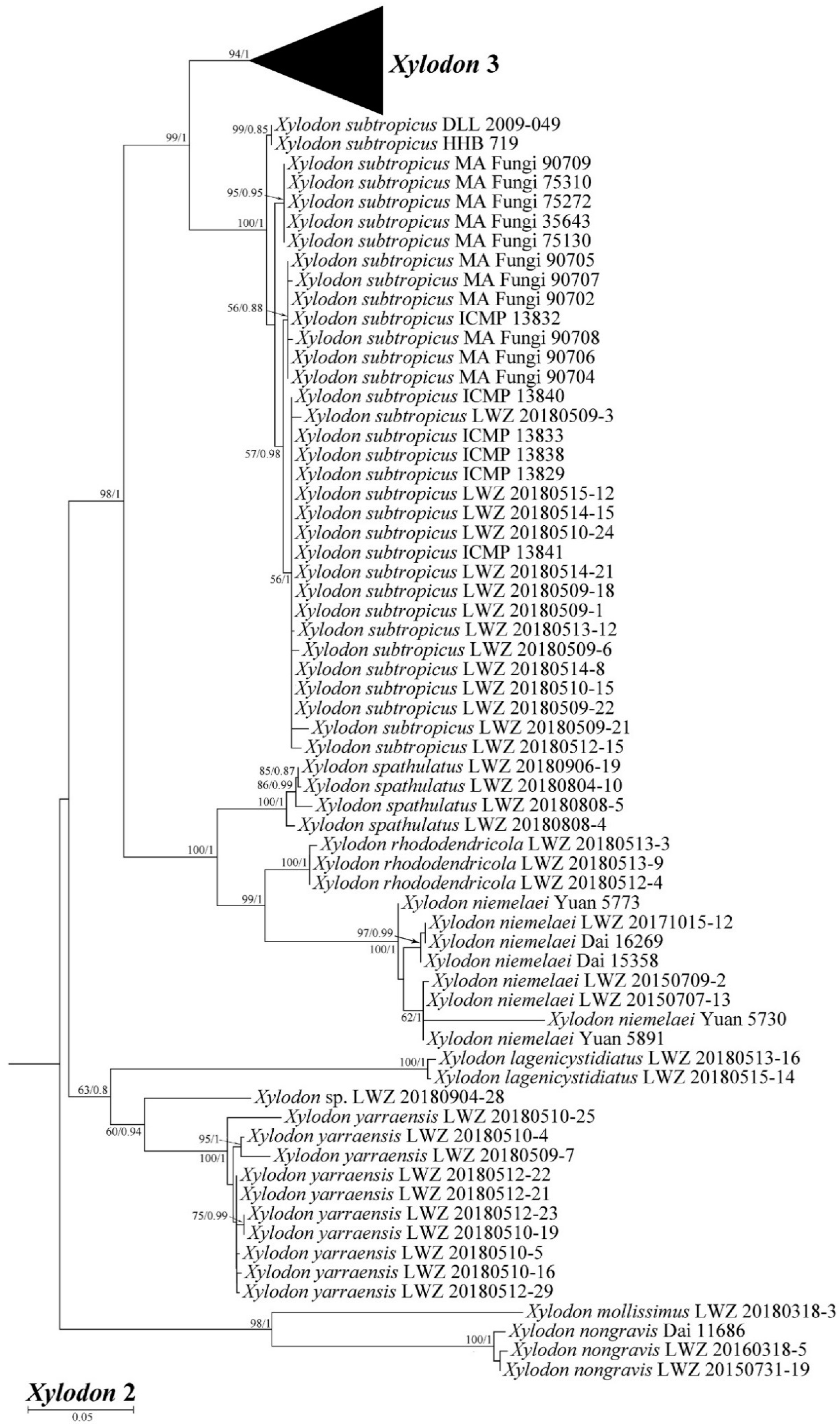
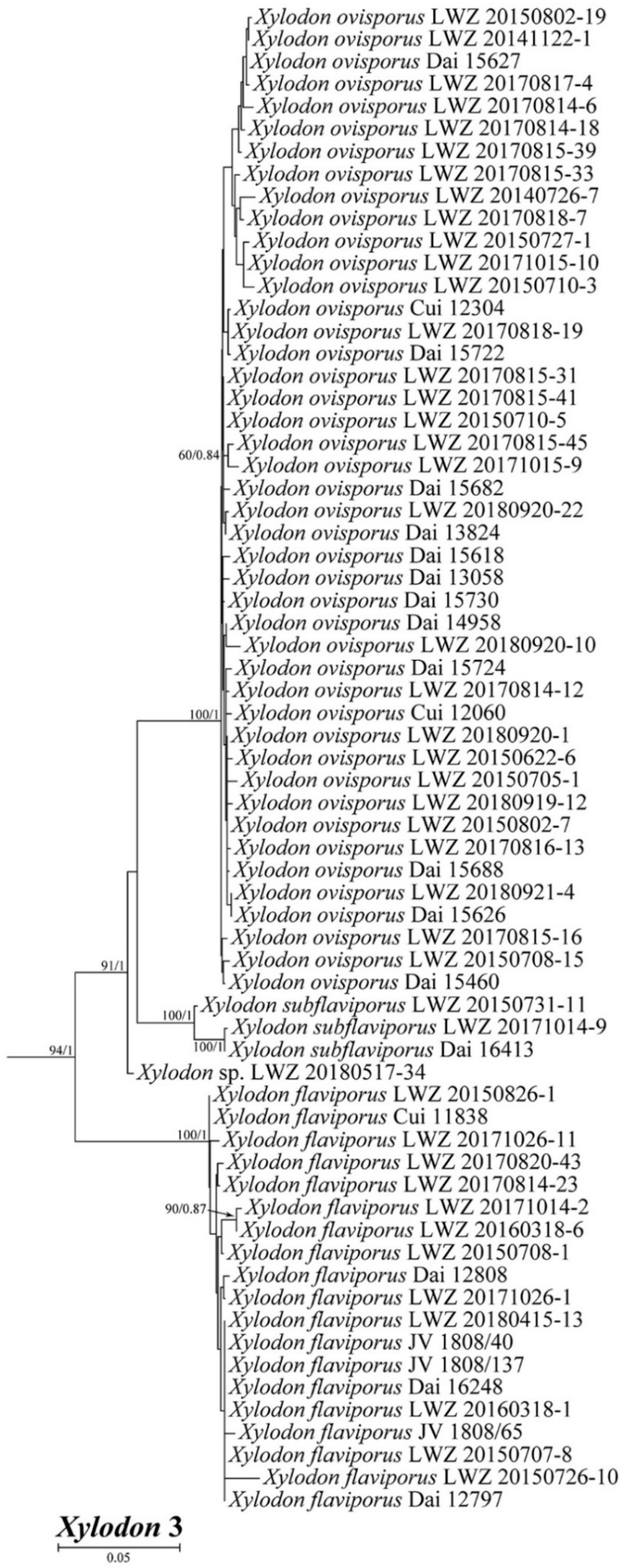
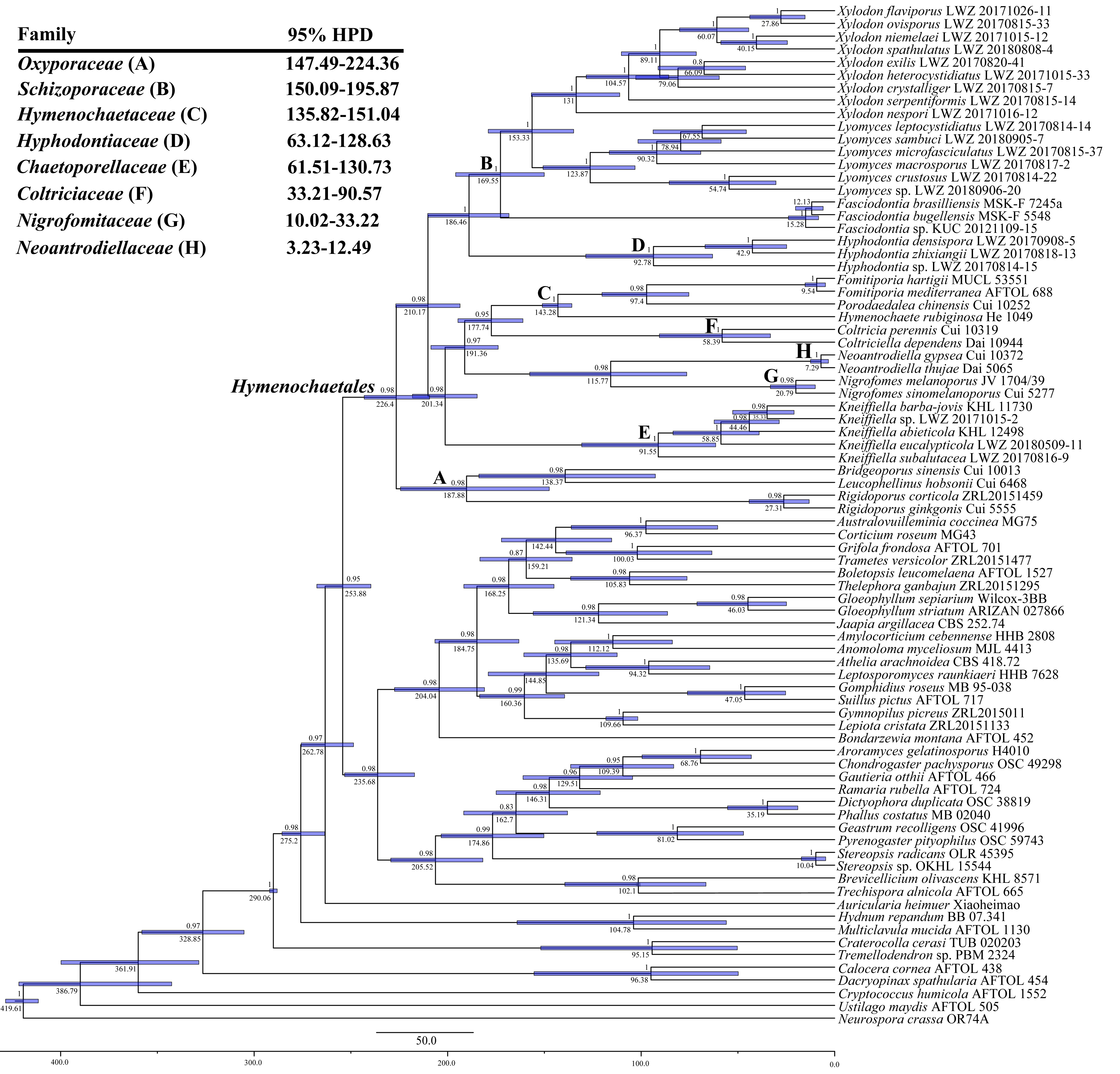
3. Results
4. Taxonomy
| A key to 23 accepted species of Kneiffiella | |
| 1a. Hymenophore smooth................................................................................................................................................................. | 2 |
| b. Hymenophore tuberculate, grandinioid, odontioid, coralloid, irpicoid or poroid.............................................................. | 10 |
| 2a. Basidia with two, rarely three sterigmata.................................................................................................................. | K. efibulata |
| b. Basidia with four sterigmata......................................................................................................................................................... | 3 |
| 3a. Without clamp connections......................................................................................................................................................... | 4 |
| b. With clamp connections................................................................................................................................................................. | 5 |
| 4a. Hymenophore cream; basidiospores less than 5 µm long, less than 2.5 µm wide.............................................. | K. tetraspora |
| b. Hymenophore whitish to ochraceous or slightly ochre-brown; basidiospores more than 5 µm long, more than 2.5 µm wide.......................................................................................................................................................................... | K. crassispora |
| 5a. Basidiospores less than 6 μm long.............................................................................................................................................. | 6 |
| b. Basidiospores more than 6 μm long............................................................................................................................................. | 8 |
| 6a. Basidiospores cylindrical to allantoid; tramacystidia without exudate.................................................................... | K. altaica |
| b. Basidiospores ellipsoid or cylindrical to suballantoid; tramacystidia sometimes apically with exudate........................ | 7 |
| 7a. Basidiospores less than 3 μm wide....................................................................................................................... | K. microspora |
| b. Basidiospores more than 3 μm wide............................................................................................................................ | K. alienata |
| 8a. Basidiospores less than 2.5 μm wide................................................................................................................... | K. subalutacea |
| b. Basidiospores more than 2.5 μm wide........................................................................................................................................ | 9 |
| 9a. Hymenophore white to slightly cream; tramacystidia without crystals............................................................ | K. cineracea |
| b. Hymenophore creamish white to slightly yellow, light ochre in spots; tramacystidia smooth or encrusted, sometimes with simple secondary septa............................................................................................................ | K. decorticans |
| 10a. Without clamp connections.................................................................................................................................................... | 11 |
| b. With clamp connections.............................................................................................................................................................. | 16 |
| 11a. Basidiospores less than 5 μm long......................................................................................................................................... | 12 |
| b. Basidiospores more than 5 μm long.......................................................................................................................................... | 15 |
| 12a. Hymenophore odontioid or irpicoid..................................................................................................................................... | 13 |
| b. Hymenophore slightly grandinioid or tuberculate to grandinioid...................................................................................... | 14 |
| 13a. Hymenophore odontioid, fresh cream-colored, dry buff or dark brown; basidiospores less than 4 μm long......................................................................................................................................................................... | K. subefibulata |
| b. Hymenophore odontioid to irpicoid, white; basidiospores more than 4 μm long.......................................... | K. subglobosa |
| 14a. Hymenophore tuberculate to grandinioid, yellowish brown to ochraceous; cystidia without exudate; basidiospores more than 2.5 μm wide.................................................................................................................. | K. byssoidea |
| b. Hymenophore slightly grandinioid, cream; cystidia with exudate; basidiospores less than 2.5 μm wide... | K. tetraspora |
| 15a. Basidia with two or rarely three sterigmata; basidiospores cylindrical to slightly allantoid......................... | K. efibulata |
| b. Basidia with four sterigmata; basidiospores ellipsoid......................................................................................... | K. crassispora |
| 16a. Capitate cystidia in subiculum present................................................................................................................................. | 17 |
| b. Capitate cystidia in subiculum absent...................................................................................................................................... | 18 |
| 17a. Basidiospores broadly ellipsoid (Q < 1.5), thin- to slightly thick-walled…....................................................... | K. sinensis |
| b. Basidiospores ellipsoid (Q > 1.5), thin-walled.................................................................................................. | K. pilaecystidiata |
| 18a. Tramacystidia thin-walled...................................................................................................................................................... | 19 |
| b. Tramacystidia thick-walled........................................................................................................................................................ | 22 |
| 19a. Hymenophore irpicoid to poroid, white to yellowish; spores narrowly allantoid, spores 3–4 × 0.5–1 μm..... | K. abdita |
| b. Hymenophore grandinioid to odontioid.................................................................................................................................. | 20 |
| 20a. Basidiospores narrowly allantoid or semicircle-like, no more than 2 µm wide............................................................. | 21 |
| b. Basidiospores broadly ellipsoid to ovoid, more than 2 µm wide................................................................... | K. eucalypticola |
| 21a. Basidiospores narrowly allantoid, more than 5 µm long; aculei sometimes grouped together then coralloid....................................................................................................................................................................... | K. alutacea |
| b. Basidiospores semicircle-like, no more than 5 µm long; aculei not grouped.................................................... | K. curvispora |
| 22a. Septa of basal hyphae and part of subiculum without clamp connections; short aculei (up to 50 μm) among long aculei (up to 800 μm)............................................................................................................................................... | K. stereicola |
| b. Clamp connections everywhere; aculei uniform..................................................................................................................... | 23 |
| 23a. Basidiospores less than 4 μm wide........................................................................................................................................ | 24 |
| b. Basidiospores more than 4 μm wide......................................................................................................................................... | 27 |
| 24a. Basidiospores less than 5.5 μm long................................................................................................................. | K. tubuliformis |
| b. Basidiospores more than 6 μm long.......................................................................................................................................... | 25 |
| 25a. Basidiospores cylindrical to suballantoid, more than 2.5 μm wide............................................................... | K. decorticans |
| b. Basidiospores long allantoid, less than 2.5 μm wide.............................................................................................................. | 26 |
| 26a. Hymenophore odontioid........................................................................................................................................... | K. floccosa |
| b. Hymenophore slightly grandinioid....................................................................................................................... | K. subalutacea |
| 27a. Basidiospores cylindrical; hymenophore cream to ochraceous, aculei up to 1 mm long.............................. | K. abieticola |
| b. Basidiospores ellipsoid; hymenophore cream, aculei up to 3 mm long............................................................ | K. barba-jovis |
| A key to 13 accepted species of Hyphodontia | |
| 1a. Hymenophore smooth to grandinioid....................................................................................................................................... | 2 |
| b. Hymenophore odontioid or poroid.............................................................................................................................................. | 7 |
| 2a. Lagenocystidia abundant............................................................................................................................................................ | 3 |
| b. Lagenocystidia rare or absent....................................................................................................................................................... | 5 |
| 3a. Capitate septocystidia absent; basidiospores thick-walled, more than 4 μm wide......................................... | H. pachyspora |
| b. Capitate septocystidia present; basidiospores thin-walled, less than 4 μm wide................................................................... | 4 |
| 4a. Hymenophore colliculose............................................................................................................................................ | H. alutaria |
| b. Hymenophore grandinioid........................................................................................................................................ | H. zhixiangii |
| 5a. Basidiospores less than 5 μm long, less than 3 µm wide......................................................................................... | H. pallidula |
| b. Basidiospores more than 5 μm long, more than 4 µm wide...................................................................................................... | 6 |
| 6a. Lagenocystidia generally rare............................................................................................................................... | H. subdetritica |
| b. Lagenocystidia absent............................................................................................................................................. | H. subpallidula |
| 7a. Hymenophore poroid................................................................................................................................................ | H. borbonica |
| b. Hymenophore odontioid............................................................................................................................................................... | 8 |
| 8a. Hymenophore brownish orange, cinnamon-buff, tawny olive or buckthorn brown....................................... | H. ochroflava |
| b. Hymenophore whitish to yellowish, cream or orange............................................................................................................... | 9 |
| 9a. Basidiospores globose, slightly thick-walled..................................................................................................... | H. sphaerospora |
| b. Basidiospores ellipsoid, cylindrical or ovoid, thin-walled...................................................................................................... | 10 |
| 10a. Capitate cystidia absent........................................................................................................................................................... | 11 |
| b. Capitate cystidia present.............................................................................................................................................................. | 12 |
| 11a. Basidiospores ellipsoid to cylindrical, more than 4.5 µm long, less than 3 µm wide.......................................... | H. wrightii |
| b. Basidiospores broadly ellipsoid to ovoid, less than 4.5 µm long, more than 3 µm wide....................................... | H. wongiae |
| 12a. Basidiospores more than 3 μm wide; aculei up to 2 mm long.................................................................................. | H. arguta |
| b. Basidiospores less than 3 μm wide; aculei up to 6 mm long..................................................................................... | H. dhingrae |
| A key to 28 accepted species of Lyomyces | |
| 1a. Hymenophore verrucose, grandinioid or odontioid................................................................................................................ | 2 |
| b. Hymenophore smooth or slightly grandinioid......................................................................................................................... | 13 |
| 2a. Subulate cystidia present, sometimes apically with a small capitate structure..................................................................... | 3 |
| b. Subulate cystidia absent................................................................................................................................................................. | 7 |
| 3a. Capitate cystidia present.......................................................................................................................................... | L. macrescens |
| b. Capitate cystidia absent, very few subcapitate cystidioles possible......................................................................................... | 4 |
| 4a. Subulate cystidioles only laterally in aculei.............................................................................................................. | L. stratosus |
| b. Subulate cystidia in hymenium.................................................................................................................................................... | 5 |
| 5a. Subulate cystidia apically not capitate........................................................................................................................ | L. juniperi |
| b. Subulate cystidia sometimes apically capitate............................................................................................................................ | 6 |
| 6a. Hymenophore young grandinioid, odontioid when older, white to yellowish or cream; basidiospores narrowly ellipsoid to cylindrical, sometimes slightly allantoid........................................................................................... | L. crustosus |
| b. Hymenophore partly fine aculeate, white to pale greyish; basidiospores cylindrical.................................... | L. vietnamensis |
| 7a. With septocystidia, capitate cystidia and transitional forms........................................................................................ | L. albus |
| b. Without septocystidia.................................................................................................................................................................... | 8 |
| 8a. Basidiospores globose, 4–5 μm in diam, verrucose, slightly thick-walled.......................................................... | L. griseliniae |
| b. Basidiospores ellipsoid, smooth, thin-walled............................................................................................................................. | 9 |
| 9a. Basidiospores less than 3.5 μm wide........................................................................................................................................ | 10 |
| b. Basidiospores more than 3.5 μm wide....................................................................................................................................... | 11 |
| 10a. Basidiospores less than 5.5 µm long; capitate cystidia present............................................................... | L. microfasciculatus |
| b. Basidiospores mostly more than 5.5 µm long; capitate cystidia absent................................................................... | L. stratosus |
| 11a. Hymenophore odontioid, margin abrupt.............................................................................................................................. | 12 |
| b. Hymenophore grandinioid, margin thinning out...................................................................................... | L. capitatocystidiatus |
| 12a. Hyphae with a lot of crystals................................................................................................................................... | L. fimbriatus |
| b. Hyphae without crystals..................................................................................................................................................... | L. pruni |
| 13a. Basidiospores mostly more than 5.5 μm wide.................................................................................................... | L. incrustatus |
| b. Basidiospores less than 5.5 μm wide.......................................................................................................................................... | 14 |
| 14a. Basidiospores suballantoid to reniform.................................................................................................................. | L. eburneus |
| b. Basidiospores not reniform......................................................................................................................................................... | 15 |
| 15a. Generally with two or rarely three sterigmata................................................................................................ | L. bisterigmatus |
| b. Generally with four sterigmata................................................................................................................................................... | 16 |
| 16a. Hymenophore margin abrupt................................................................................................................................................. | 17 |
| b. Hymenophore margin thinning out........................................................................................................................................... | 24 |
| 17a. Capitate or subcapitate cystidia present................................................................................................................................ | 18 |
| b. Capitate or subcapitate cystidia absent...................................................................................................................................... | 23 |
| 18a. Basidiospores cyanophilous.................................................................................................................................................... | 19 |
| b. Basidiospores acyanophilous...................................................................................................................................................... | 22 |
| 19a. Submoniliform cystidia present.......................................................................................................................... | L. mascarensis |
| b. Submoniliform cystidia absent, tapering cystidia present.................................................................................................. | 20 |
| 20a. Tapering cystidia more than 40 μm long............................................................................................................ | L. bambusinus |
| b. Tapering cystidia less than 40 μm long.................................................................................................................................. | 21 |
| 21a. Fusoid cystidioles present........................................................................................................................... | L. wuliangshanensis |
| b. Fusoid cystidioles absent............................................................................................................................................ | L. cremeus |
| 22a. Basidiospores mostly less than 5 μm long................................................................................................................. | L. gatesiae |
| b. Basidiospores more than 5 μm long............................................................................................................................ | L. orientalis |
| 23a. Fusiform or cylindrical hyphidia-like cystidia present....................................................................................... | L. organensis |
| b. Subulate cystidia present................................................................................................................................................ | L. juniperi |
| 24a. Basidiospores mostly more than 6.5 µm wide.................................................................................................. | L. macrosporus |
| b. Basidiospores less than 6.5 µm wide.......................................................................................................................................... | 25 |
| 25a. Capitate cystidia absent........................................................................................................................................................... | 26 |
| b. Capitate cystidia present.............................................................................................................................................................. | 28 |
| 26a. Basidiospores more than 7 μm long.................................................................................................................. | L. allantosporus |
| b. Basidiospores less than 7 μm long.............................................................................................................................................. | 27 |
| 27a. Basidiospores cyanophilous................................................................................................................................... | L. elaeidicola |
| b. Basidiospores acyanophilous...................................................................................................................................................... | 29 |
| 28a. Hymenophore smooth; capitate cystidia; hyphae usually with a lot of crystals.................................................. | L. sambuci |
| b. Hymenophore first for a long time porose-reticulate, later smooth; hyphae usually without crystals................... | L. erastii |
| 29a. Leptocystidia present............................................................................................................................................................... | 30 |
| b. Leptocystidia absent..................................................................................................................................................................... | 31 |
| 30a. Leptocystidia slightly fusiform, not encrusted, thin-walled, more than 5 µm wide; hyphae up to 4 µm in diam........................................................................................................................................................................... | L. boninensis |
| b. Leptocystidia not fusiform, encrusted with crystals, thin- to slightly thick-walled, mostly less than 5 µm wide; hyphae up to 5.5 µm in diam........................................................................................................................................ | L. leptocystidiatus |
| 31a. Hymenophore very finely aculeate................................................................................................................... | L. vietnamensis |
| b. Hymenophore fairly smooth................................................................................................................................... | L. tenuissimus |
| A key to 87 accepted species of Xylodon | |
| 1a. Hymenophore odontioid or coralloid........................................................................................................................................ | 2 |
| b. Hymenophore smooth, tuberculate, grandinioid, irpicoid or poroid.................................................................................... | 66 |
| 2a. Capitate or subcapitate cystidia or cystidioles present............................................................................................................ | 3 |
| b. Capitate or subcapitate cystidia or cystidioles absent.............................................................................................................. | 38 |
| 3a. Basidiospores ellipsoid................................................................................................................................................................ | 4 |
| b. Basidiospores globose to subglobose, ovoid, cylindrical or allantoid.................................................................................... | 24 |
| 4a. Moniliform or submoniliform cystidia present........................................................................................................................ | 5 |
| b. Moniliform or submoniliform cystidia absent.......................................................................................................................... | 11 |
| 5a. Basidiospores thin-walled........................................................................................................................................................... | 6 |
| b. Basidiospores thick-walled........................................................................................................................................................... | 9 |
| 6a. Basidiospores up to 7.5 μm long...................................................................................................................... | X. anmashanensis |
| b. Basidiospores up to 6 μm long...................................................................................................................................................... | 7 |
| 7a. Moniliform cystidia originating only in subhymenium..................................................................................... | X. spathulatus |
| b. Moniliform cystidia originating only in subiculum or in subiculum and subhymenium..................................................... | 8 |
| 8a. Aculei up to 1 mm long, hymenophoral margin thinning out............................................................................. | X. brevisetus |
| b. Aculei up to 0.3 mm long, hymenophoral margin abrupt or byssoid................................................................. | X. subclavatus |
| 9a. Astrocystidia present; basidiospores cyanophilous............................................................................................ | X. ussuriensis |
| b. Astrocystidia absent; basidiospores acyanophilous................................................................................................................ | 10 |
| 10a. Hyphoid or subulate cystidia absent................................................................................................................. | X. crassisporus |
| b. Hyphoid or subulate cystidia present............................................................................................................... | X. anmashanensis |
| 11a. Hymenophore coralloid............................................................................................................................................... | X. archeri |
| b. Hymenophore odontioid............................................................................................................................................................. | 12 |
| 12a. Basidia cylindrical to subcylindrical....................................................................................................................................... | 13 |
| b. Basidia utriform............................................................................................................................................................................ | 15 |
| 13a. Pin- or sting-shaped cystidia present........................................................................................................................ | X. borealis |
| b. Pin- or sting-shaped cystidia absent........................................................................................................................................... | 14 |
| 14a. Aculei 0.03–0.12 mm long, 8–14 per mm; basidiospores thin- or slightly thick-walled............................ | X. pseudolanatus |
| b. Aculei 0.13–0.35 mm long, 4 per mm; basidiospores thin-walled........................................................................ | X. vesiculosus |
| 15a. Astrocystidia present..................................................................................................................................... | X. astrocystidiatus |
| b. Astrocystidia absent..................................................................................................................................................................... | 16 |
| 16a. Only capitate cystidia and leptocystidia present...................................................................................... | X. heterocystidiatus |
| b. Different combinations of cystidia............................................................................................................................................. | 17 |
| 17a. Besides capitate to subcapitate cystidia, tramacystidia, subulate or apically encrusted hyphoid cystidia present........................................................................................................................................................................................ | 18 |
| b. Only capitate cystidia present..................................................................................................................................................... | 22 |
| 18a. Tramacystidia present.............................................................................................................................................................. | 19 |
| b. Tramacystidia absent................................................................................................................................................................... | 20 |
| 19a. Capitate cystidia slightly thick-walled; basidiospores thick-walled, more than 4 µm wide........................ | X. magnificus |
| b. Capitate cystidia thin-walled; basidiospores thin-walled, less than 4 µm wide...................................................... | X. lanatus |
| 20a. Subulate cystidia present....................................................................................................................................... | X. pruniaceus |
| b. Subulate cystidia absent............................................................................................................................................................... | 21 |
| 21a. Hymenophore cream; hyphoid cystidia sometimes with stellate crystalline cap.......................................... | X. attenuatus |
| b. Hymenophore white; hyphoid cystidia often heavily encrusted and rarely with stellate crystalline cap..... | X. crystalliger |
| 22a. Basidia subclavate..................................................................................................................................................... | X. capitatus |
| b. Basidia sinuous or utriform......................................................................................................................................................... | 23 |
| 23a. Basidiospores thin-walled, more than 3.5 µm wide................................................................................................. | X. asperus |
| b. Basidiospores slightly thick-walled, less than 3.5 µm wide............................................................................. | X. kunmingensis |
| 24a. Hymenophore coralloid or odontioid with fimbriate aculei............................................................................................... | 25 |
| b. Hymenophore odontioid without fimbriate aculei.................................................................................................................. | 28 |
| 25a. Aculei up to 3 mm long............................................................................................................................................ | X. quercinus |
| b. Aculei up to 1 mm long................................................................................................................................................................ | 26 |
| 26a. Basidiospores more than 3 μm wide........................................................................................................................... | X. archeri |
| b. Basidiospores less than 3 μm wide............................................................................................................................................. | 27 |
| 27a. Aculei up to 200 μm long; basidiospores less than 6 μm long................................................................................ | X. nesporii |
| b. Aculei up to 600 μm long; basidiospores more than 6 μm long................................................................................ | X. ramicida |
| 28a. Basidiospores more than 7 μm long........................................................................................................................................ | 29 |
| b. Basidiospores less than 7 μm long.............................................................................................................................................. | 30 |
| 29a. Basidiospores ovoid to allantoid, less than 5 µm wide.................................................................................. | X. adhaerisporus |
| b. Basidiospores globose to subglobose, more than 5 µm wide.......................................................................................... | X. follis |
| 30a. Basidiospores slightly thick-walled........................................................................................................................................ | 31 |
| b. Basidiospores thin-walled........................................................................................................................................................... | 32 |
| 31a. Pin- or sting-like cystidia abundant; basidiospores slightly cylindrical, ellipsoid or broadly ellipsoid........... | X. borealis |
| b. Pin- or sting-like cystidia absent; basidiospores globose or subglobose....................................................... | X. hyphodontinus |
| 32a. Basidiospores cylindrical or subcylindrical..................................................................................................... | X. rimosissimus |
| b. Basidiospores differently shaped............................................................................................................................................... | 33 |
| 33a. Moniliform cystidia scattered.............................................................................................................................. | X. subclavatus |
| b. Moniliform cystidia absent.......................................................................................................................................................... | 34 |
| 34a. Fusiform cystidia present in aculei............................................................................................................................ | X. hastifer |
| b. Fusiform cystidia absent.............................................................................................................................................................. | 35 |
| 35a. Basidiospores less than 5 μm long.......................................................................................................................................... | 36 |
| b. Basidiospores more than 5 μm long........................................................................................................................................... | 37 |
| 36a. Subcylindrical and hyphoid to narrowly ventricose cystidia absent.................................................................... | X. tenellus |
| b. Subcylindrical and hyphoid to narrowly ventricose cystidia present...................................................................... | X. filicinus |
| 37a. Capitate cystidia in hymenium and subhymenium, capitate hyphal endings in subiculum and aculei…...... | X. asperus |
| b. Capitate gloeocystidia or hyphal endings only in hymenium and/or aculei......................................................... | X. capitatus |
| 38a. Subulate cystidia present......................................................................................................................................................... | 39 |
| b. Subulate cystidia absent............................................................................................................................................................... | 48 |
| 39a. Basidiospores ellipsoid or narrowly ellipsoid....................................................................................................................... | 40 |
| b. Basidiospores cylindrical, allantoid, ovoid or subglobose...................................................................................................... | 45 |
| 40a. Leptocystidia present................................................................................................................................................. | X. australis |
| b. Leptocystidia absent.................................................................................................................................................................... | 41 |
| 41a. Hyphoid and moniliform to submoniliform cystidia present.................................................................... | X. anmashanensis |
| b. Hyphoid and moniliform to submoniliform cystidia absent.................................................................................................. | 42 |
| 42a. Basidiospores often glued together in packs of two to six basidiospores................................................... | X. candidissimus |
| b. Basidiospores separated.............................................................................................................................................................. | 43 |
| 43a. Subulate cystidia thin- or slightly thick-walled, less than 30 μm long...................................................... | X. submucronatus |
| b. Subulate cystidia thin-walled, more than 30 μm long.............................................................................................................. | 44 |
| 44a. Basidia utriform with one or two constrictions; subulate cystidia up to 75 μm long..................................... | X. knysnanus |
| b. Basidia utriform to galzinoid with several constrictions; subulate cystidia up to 50 μm long........................... | X. nudisetus |
| 45a. Basidiospores ovoid to allantoid, more than 7 μm long................................................................................ | X. adhaerisporus |
| b. Basidiospores cylindrical or subglobose, less than 7 μm long................................................................................................ | 46 |
| 46a. Hymenophore greyish-white to yellowish; subulate cystidia or hyphal endings................................ | X. crustosoglobosus |
| b. Hymenophore white to cream; subulate cystidia up to 50 or 75 μm long.............................................................................. | 47 |
| 47a. Basidia utriform with one or two constrictions; subulate cystidia up to 75 μm long..................................... | X. knysnanus |
| b. Basidia utriform to galzinoid with several constrictions; subulate cystidia up to 50 μm long........................... | X. nudisetus |
| 48a. Moniliform or submoniliform cystidia present.................................................................................................................... | 49 |
| b. Moniliform or submoniliform cystidia absent.......................................................................................................................... | 51 |
| 49a. Only moniliform cystidia present................................................................................................................................... | X. lenis |
| b. Other types of cystidia present.................................................................................................................................................... | 50 |
| 50a. Hyphoid cystidia and rare capitate cystidioles present.............................................................................. | X. anmashanensis |
| b. Cylindrical gloeocystidia and subclavate cystidia present............................................................................. | X. subscopinellus |
| 51a. Basidiospores suballantoid or subglobose............................................................................................................................. | 52 |
| b. Basidiospores ellipsoid or cylindrical........................................................................................................................................ | 54 |
| 52a. Basidiospores suballantoid, more than 6 µm long................................................................................................. | X. syringae |
| b. Basidiospores subglobose, less than 6 µm long........................................................................................................................ | 53 |
| 53a. Capitate hyphal endings present in aculei.................................................................................................................... | X. rudis |
| b. Capitate hyphal endings absent.............................................................................................................................. | X. subglobosus |
| 54a. Cystidia or cystidioles absent.................................................................................................................................................. | 55 |
| b. Cystidia or cystidioles present.................................................................................................................................................... | 56 |
| 55a. Basidiospores narrowly ellipsoid to cylindrical, more than 6 µm long, less than 3 µm wide......................... | X. nesporina |
| b. Basidiospores ellipsoid, less than 6 µm long, more than 3 µm wide............................................................... | X. mussooriensis |
| 56a. Cylindrical, clavate to subclavate, spathuliform cystidia or leptocystidia present.......................................................... | 57 |
| b. Tubular, ventricose, pyriform, hyphoid or sinuous cystidia present..................................................................................... | 60 |
| 57a. Clavate to spathuliform cystidia present............................................................................................................................... | 58 |
| b. Clavate to spathuliform cystidia absent..................................................................................................................................... | 59 |
| 58a. Hymenophore greyish-white or pale cream; basidiospores thick-walled, more than 4 µm wide................. | X. pruinosus |
| b. Hymenophore cinnamon to orange-brown; basidiospores thin-walled, less than 4 µm wide...................... | X. yunnanensis |
| 59a. Cylindrical gloeocystidia and subclavate cystidia....................................................................................... | X. subscopinellus |
| b. Leptocystidia present with encrustation......................................................................................................... | X. rhododendricola |
| 60a. Tubular cystidia present.......................................................................................................................................................... | 61 |
| b. Tubular cystidia absent................................................................................................................................................................ | 62 |
| 61a. Tubular cystidia thick-walled, slightly flexuous; basidia with two or rarely four sterigmata; basidiospores slightly thick-walled............................................................................................................................................................... | X. echinatus |
| b. Tubular tramacystidia thin-walled, snake-like sinuous; basidia with four sterigmata; basidiospores thin-walled........................................................................................................................................................... | X. subserpentiformis |
| 62a. Pyriform cystidia present......................................................................................................................................... | X. pelliculae |
| b. Pyriform cystidia absent.............................................................................................................................................................. | 63 |
| 63a. Ventricose cystidia present............................................................................................................................ | X. submucronatus |
| b. Ventricose cystidia absent........................................................................................................................................................... | 64 |
| 64a. Basidiospores more than 6 μm long........................................................................................................................ | X. lutescens |
| b. Basidiospores less than 6 μm long.............................................................................................................................................. | 65 |
| 65a. Basidiospores more than 3.5 µm wide........................................................................................................................... | X. rudis |
| b. Basidiospores less than 3.5 µm wide......................................................................................................................... | X. papillosus |
| 66a. Hymenophore irpicoid or poroid........................................................................................................................................... | 67 |
| b. Hymenophore smooth, tuberculate or grandinioid................................................................................................................. | 89 |
| 67a. Cystidia absent…..................................................................................................................................................... | X. nongravis |
| b. Cystidia present............................................................................................................................................................................ | 68 |
| 68a. Moniliform or submoniliform cystidia present.................................................................................................................... | 69 |
| b. Moniliform or submoniliform cystidia absent.......................................................................................................................... | 72 |
| 69a. Hymenophore irpicoid............................................................................................................................................................. | 70 |
| b. Hymenophore poroid.................................................................................................................................................................. | 71 |
| 70a. Hyphoid or subulate cystidia present mostly in aculei............................................................................... | X. anmashanensis |
| b. Hyphoid or subulate cystidia absent...................................................................................................................... | X. spathulatus |
| 71a. Basidiospores ellipsoid, less than 5.5 μm long..................................................................................................... | X. bresinskyi |
| b. Basidiospores suballantoid, more than 7.5 μm long.................................................................................................. | X. syringae |
| 72a. Basidiospores more than 6.5 μm long..................................................................................................................................... | 73 |
| b. Basidiospores less than 6.5 μm long........................................................................................................................................... | 74 |
| 73a. Pores less than 1 mm in diam; cystidia capitate; basidiospores less than 8 µm long........................................ | X. nothofagi |
| b. Pores more than 1 mm in diam; cystidia not capitate; basidiospores mostly more than 8 µm long..................... | X. syringae |
| 74a. Basidia with two sterigmata............................................................................................................................... | X. crassihyphus |
| b. Basidia with four sterigmata....................................................................................................................................................... | 75 |
| 75a. Hymenophore pileate; hyphal system trimitic.................................................................................................. | X. trametoides |
| b. Hymenophore resupinate or effused-reflected; hyphal system monomitic to dimitic........................................................ | 76 |
| 76a. Subulate or cylindrical cystidia present................................................................................................................................. | 77 |
| b. Subulate or cylindrical cystidia absent....................................................................................................................................... | 79 |
| 77a. Basidiospores broadly ellipsoid to subglobose, more than 4 μm wide........................................................ | X. apacheriensis |
| b. Basidiospores ellipsoid, less than 4 μm wide............................................................................................................................ | 78 |
| 78a. Basidiospores more than 5 µm long........................................................................................................................ | X. niemelaei |
| b. Basidiospores less than 5 µm long........................................................................................................................ | X. subflaviporus |
| 79a. Bottle-shaped cystidia present, apically with large rhomboid crystals........................................................... | X. cystidiatus |
| b. Bottle-shaped cystidia absent...................................................................................................................................................... | 80 |
| 80a. Hyphae without clamp connections......................................................................................................... | X. poroideoefibulatus |
| b. Hyphae with clamp connections................................................................................................................................................ | 81 |
| 81a. Capitate cystidia mostly bladder-like.................................................................................................................................... | 82 |
| b. Capitate cystidia not bladder-like............................................................................................................................................... | 83 |
| 82a. Basidia barrel-shaped to pyriform; basidiospores ellipsoid, less than 3.2 μm wide................................ | X. pseudotropicus |
| b. Basidia utriform to clavate; basidiospores broadly ellipsoid, more than 3.2 µm wide.................................... | X. mollissimus |
| 83a. Basidiospores subglobose or ovoid........................................................................................................................................ | 84 |
| b. Basidiospores ellipsoid................................................................................................................................................................ | 85 |
| 84a. Basidiospores subglobose, more than 4 µm wide............................................................................................. | X. hallenbergii |
| b. Basidiospores ovoid, less than 4 µm wide................................................................................................................ | X. flaviporus |
| 85a. Growth on palm or fern as substrate; capitate cystidia rare.................................................................................... | X. gracilis |
| b. Growth on angiosperms or gymnosperms; capitate cystidia not rare................................................................................... | 86 |
| 86a. Basidiospores more than 5.5 μm long................................................................................................................... | X. paradoxus |
| b. Basidiospores less than 5.5 μm long........................................................................................................................................... | 87 |
| 87a. Hymenophore cream........................................................................................................................................... | X. taiwanianus |
| b. Hymenophore orange, rose or buff............................................................................................................................................ | 88 |
| 88a. Hymenophore beige to slightly orange; capitate cystidia not encrusted....................................................... | X. subtropicus |
| b. Hymenophore cream, pinkish cream or buff; capitate cystidia partly encrusted with brownish yellow resinous material...................................................................................................................................................................... | X. ovisporus |
| 89a. Basidia with two sterigmata...................................................................................................................................... | X. bisporus |
| b. Basidia with four sterigmata....................................................................................................................................................... | 90 |
| 90a. Capitate or subcapitate cystidia present................................................................................................................................ | 91 |
| b. Capitate or subcapitate cystidia absent.................................................................................................................................... | 106 |
| 91a. Spores globose to subglobose.................................................................................................................................................. | 92 |
| b. Spores cylindrical, ellipsoid to broadly ellipsoid...................................................................................................................... | 94 |
| 92a. Hymenophore smooth.............................................................................................................................................. | X. pumilius |
| b. Hymenophore grandinioid......................................................................................................................................................... | 93 |
| 93a. Capitate cystidia rare; basidiospores less than 4.5 μm long................................................................................... | X. tenellus |
| b. Capitate cystidia not rare; basidiospores more than 4.5 μm long.................................................................. | X. hyphodontinus |
| 94a. Gloeocystidia present.......................................................................................................................................... | X. tuberculatus |
| b. Gloeocystidia absent.................................................................................................................................................................... | 95 |
| 95a. Tramacystidia present........................................................................................................................................... | X. verecundus |
| b. Tramacystidia absent................................................................................................................................................................... | 96 |
| 96a. Basidiospores thick-walled...................................................................................................................................................... | 97 |
| b. Basidiospores thin-walled......................................................................................................................................................... | 100 |
| 97a. Basidiospores less than 3.5 µm wide............................................................................................................................. | X. rickii |
| b. Basidiospores more than 3.5 µm wide....................................................................................................................................... | 98 |
| 98a. Hymenophore smooth.............................................................................................................................................. | X. pumilius |
| b. Hymenophore grandinioid......................................................................................................................................................... | 99 |
| 99a. Astrocystidia and submoniliform cystidia present, septocystidia absent...................................................... | X. ussuriensis |
| b. Astrocystidia and submoniliform cystidia absent, septocystidia present.................................................. | X. septocystidiatus |
| 100a. Only capitate cystidia present............................................................................................................................................. | 101 |
| b. Besides capitate cystidia, other types of cystidia present...................................................................................................... | 102 |
| 101a. Hyphae not encrusted; cystidia with exudation; basidiospores cylindrical to subcylindrical................ | X. rimosissimus |
| b. Hyphae sometimes encrusted; cystidia without exudation; basidiospores ellipsoid......................................... | X. yarraensis |
| 102a. Basidiospores less than 3.5 μm wide.................................................................................................................................. | 103 |
| b. Basidiospores more than 3.5 μm wide..................................................................................................................................... | 105 |
| 103a. Cystidia not encrusted................................................................................................................................ | X. heterocystidiatus |
| b. Cystidia strongly encrusted...................................................................................................................................................... | 104 |
| 104a. Basidiospores broadly ellipsoid, less than 5 µm long......................................................................................... | X. erikssonii |
| b. Basidiospores cylindrical to subcylindrical, more than 5 µm long...................................................................... | X. gamundiae |
| 105a. Hyphoid cystidia absent, lageniform, clavate and cylindrical cystidia present........................................... | X. hjortstamii |
| b. Hyphoid cystidia present, lageniform, clavate and cylindrical cystidia absent................................................. | X. attenuatus |
| 106a. Gloeocystidia present........................................................................................................................................ | X. tuberculatus |
| b. Gloeocystidia absent.................................................................................................................................................................. | 107 |
| 107a. Basidiospores globose to subglobose................................................................................................................................. | 108 |
| b. Basidiospores ellipsoid or cylindrical...................................................................................................................................... | 109 |
| 108a. Subulate cystidia or hyphal endings present, leptocystidia absent; basidiospores more than 5 µm long, more than 3.8 µm wide................................................................................................................................................... | X. crustosoglobosus |
| b. Subulate cystidia or hyphal endings absent, leptocystidia present; basidiospores more than 5 µm long, more than 3.8 µm wide.................................................................................................................................................................. | X. victoriensis |
| 109a. Cystidia absent...................................................................................................................................................................... | 110 |
| b. Cystidia present.......................................................................................................................................................................... | 112 |
| 110a. Cystidioles absent............................................................................................................................................... | X. acystidiatus |
| b. Cystidioles present..................................................................................................................................................................... | 111 |
| 111a. Basidiospores less than 5 µm long....................................................................................................................... | X. papillosus |
| b. Basidiospores more than 5 µm long.................................................................................................................... | X. tenuicystidius |
| 112a. Snake-like sinuous tubular tramacystidia present........................................................................................................... | 113 |
| b. Tubular tramacystidia absent................................................................................................................................................... | 114 |
| 113a. Hymenial hyphae encrusted.......................................................................................................................... | X. serpentiformis |
| b. Hymenial hyphae not encrusted................................................................................................................... | X. subserpentiformis |
| 114a. Subulate cystidia present........................................................................................................................................ | X. australis |
| b. Subulate cystidia absent............................................................................................................................................................. | 115 |
| 115a. Basidiospores more than 4 µm wide.................................................................................................................... | X. pruinosus |
| b. Basidiospores less than 4 µm wide........................................................................................................................................... | 116 |
| 116a. Clavate-sinuous to submoniliform cystidia present; basidiospores narrowly ellipsoid..................... | X. damansaraensis |
| b. Clavate-sinuous to submoniliform cystidia absent, basidiospores ellipsoid to broadly ellipsoid................................... | 117 |
| 117a. Leptocystidia and lagenocystidia absent, cylindrical to clavate cystidia and astrocystidia present............ | X. detriticus |
| b. Leptocystidia and lagenocystidia present, cylindrical to clavate cystidia and astrocystidia absent...... | X. lagenicystidiatus |
5. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eriksson, J. Studies in the Heterobasidiomycetes and Homobasidiomycetes–Aphyllophorales of Muddus National Park in North Sweden. Acta. Univ. Ups. Symb. Bot. Ups. 1958, 16, 1–172. [Google Scholar]
- Jülich, W. Higher taxa of Basidiomycetes. Bibl. Mycol. 1981, 85, 1–485. [Google Scholar]
- Eriksson, J.; Hjortstam, K.; Parmasto, E.; Ryvarden, L. (674) Proposal to conserve Hyphodontia Eriksson, 1958 (Fungi, Corticiaceae) over Kneiffiella Karsten, 1889. Taxon 1982, 31, 744–746. [Google Scholar]
- Gams, W. Report of the Committee for Fungi and Lichens: 3. Taxon 1993, 42, 112–118. [Google Scholar] [CrossRef]
- Langer, E.; Hallenberg, N.; Knudsen, H.; Kõljalg, U.; Langer, G.; Larsson, K.H.; Oberwinkler, F.; Parmasto, E.; Ryvarden, L.; Vesterhol, J. (1255) Proposal to reject the names Xylodon and Schizopora in favour of Hyphodontia, nom. cons. (Fungi, Corticiaceae). Taxon 1996, 45, 685–686. [Google Scholar] [CrossRef]
- Binder, M.; Hibbett, D.S.; Larsson, K.H.; Larsson, E.; Langer, E.; Langer, G. The phylogenetic distribution of resupinate forms across the major clades of mushroom-forming fungi (Homobasidiomycetes). Syst. Biodivers. 2005, 3, 113–157. [Google Scholar] [CrossRef]
- Larsson, K.H.; Parmasto, E.; Fischer, M.; Langer, E.; Nakasone, K.K.; Redhead, S.A. Hymenochaetales: A molecular phylogeny for the hymenochaetoid clade. Mycologia 2006, 98, 926–936. [Google Scholar] [CrossRef] [PubMed]
- Larsson, K.H. Re-thinking the classification of corticioid fungi. Mycol. Res. 2007, 111, 1040–1063. [Google Scholar] [CrossRef]
- Hjortstam, K.; Ryvarden, L. Checklist of corticioid fungi (Basidiomycotina) from the tropics, subtropics, and the southern hemisphere. Synop. Fungorum. 2007, 22, 27–146. [Google Scholar]
- Hjortstam, K.; Ryvarden, L. A checklist of names in Hyphodontia sensu stricto - sensu lato and Schizopora with new combinations in Lagarobasidium, Lyomyces, Kneiffiella, Schizopora, and Xylodon. Synop. Fungorum. 2009, 26, 33–55. [Google Scholar]
- Hjortstam, K.; Ryvarden, L. The genus Palifer (Basidiomycotina, Aphyllophorales). Synop. Fungorum. 2007, 22, 7–10. [Google Scholar]
- Ţura, D.A.; Zmitrovich, I.V.; Wasser, S.P.; Spirin, W.A.; Nevo, E. Biodiversity of the Heterobasidiomycetes and Non-Gilled Hymenomycetes (Former Aphyllophorales) of Israel; A.R.A. Gantner Verlag K.G.: Ruggell, Liechtenstein, 2010. [Google Scholar]
- Yurchenko, E.; Riebesehl, J.; Langer, E. Fasciodontia gen. nov. (Hymenochaetales, Basidiomycota) and the taxonomic status of Deviodontia. Mycol. Prog. 2020, 19, 171–184. [Google Scholar] [CrossRef]
- Yurchenko, E.; Wu, S.H. A key to the species of Hyphodontia sensu lato. MycoKeys 2016, 12, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.C.; Wu, S.H.; Chen, C.Y. Three new species of Hyphodontia s.l. (Basidiomycota) with poroid or raduloid hymenophore. Mycol. Prog. 2017, 16, 553–564. [Google Scholar] [CrossRef]
- Riebesehl, J.; Langer, E. Hyphodontia s.l. (Hymenochaetales, Basidiomycota): 35 new combinations and new keys to all 120 current species. Mycol. Prog. 2017, 16, 637–666. [Google Scholar] [CrossRef]
- Yurchenko, E.; Riebesehl, J.; Langer, E. Clarification of Lyomyces sambuci complex with the descriptions of four new species. Mycol. Prog. 2017, 16, 865–876. [Google Scholar] [CrossRef]
- Riebesehl, J.; Yurchenko, E.; Nakasone, K.K.; Langer, E. Phylogenetic and morphological studies in Xylodon (Hymenochaetales, Basidiomycota) with the addition of four new species. MycoKeys 2019, 47, 97–137. [Google Scholar] [CrossRef]
- Langer, E. Die Gattung Hyphodontia John Eriksson. Bibl. Mycol. 1994, 154, 1–298. [Google Scholar]
- Eriksson, J.; Ryvarden, L. The Corticiaceae of North Europe 4. Hyphodermella-Mycoacia; Fungiflora: Oslo, Norway, 1976. [Google Scholar]
- Eriksson, J.; Ryvarden, L. The Corticiaceae of North Europe 7. Schizopora-Suillosporium; Fungiflora: Oslo, Norway, 1984. [Google Scholar]
- Bernicchia, A.; Gorjón, S.P. Corticiaceae s. l. Fungi Europaei 12; Candusso Edizioni: Alassio, Italy, 2010. [Google Scholar]
- Ginns, J.; Lefebvre, M.N.L. Lignicolous corticioid fungi (Basidiomycota) of North America: Systematics, distribution, and ecology. Mycol. Memoir. 1993, 19, 1–247. [Google Scholar]
- Ginns, J. Genera of the North American Corticiaceae sensu lato. Mycologia 1998, 90, 1–35. [Google Scholar] [CrossRef]
- Greslebin, A.G.; Rajchenberg, M. The genus Hyphodontia in the Patagonian Andes forest of Argentina. Mycologia 2000, 92, 1155–1165. [Google Scholar] [CrossRef]
- Hjortstam, K.; Ryvarden, L.; Iturriaga, T. Studies in corticioid fungi from Venezuela II (Basidiomycotina, Aphyllophorales). Synop. Fungorum. 2005, 20, 42–78. [Google Scholar]
- Hjortstam, K.; Ryvarden, L. Studies in corticioid fungi from Venezuela III (Basidiomycotina, Aphyllophorales). Synop. Fungorum. 2007, 23, 56–107. [Google Scholar]
- Hjortstam, K.; Ryvarden, L. Studies in tropical corticioid fungi (Basidiomycotina, Aphyllophorales) Alutaceodontia, Botryodontia, Hyphodontia s.s. and Kneiffiella. Synop. Fungorum. 2002, 15, 7–17. [Google Scholar]
- Gafforov, Y.; Riebesehl, J.; Ordynets, A.; Langer, E.; Yarasheva, M.; Ghobad-Nejhad, M.; Zhou, L.W.; Wang, X.W.; de Mello Gugliotta, A. Hyphodontia (Hymenochaetales, Basidiomycota) and similar taxa from Central Asia. Botany 2017, 95, 1041–1056. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.H. The Corticiaceae (Basidiomycetes) subfamilies Phlebioideae, Phanerochaetoideae and Hyphodermoideae in Taiwan. Acta Bot. Fenn. 1990, 142, 1–123. [Google Scholar]
- Dai, Y.C.; Xiong, H.X. Flora fungorum sinicorum Vol 42. Cortiaceae s.l. (I); Science Press: Beijing, China, 2012. [Google Scholar]
- Douanla-Meli, C.; Langer, E. Fungi of Cameroon 1. New corticioid species (Basidiomycetes). Mycotaxon 2009, 107, 95–103. [Google Scholar] [CrossRef]
- Sanyal, S.K.; Dhingra, G.S. Hyphodontia dhingrae sp. nov. from India. Mycotaxon 2014, 129, 209–211. [Google Scholar]
- Riebesehl, J.; Langer, E.; Ordynets, A.; Striegel, M.M.; Witzany, C. Hyphodontia borbonica, a new species from La Réunion. Mycol. Prog. 2015, 14, 104. [Google Scholar] [CrossRef]
- Chen, C.C.; Wu, S.H.; Chen, C.Y. Xylodon subflaviporus sp. nov. (Hymenochaetales, Basidiomycota) from East Asia. Mycoscience 2018, 59, 343–353. [Google Scholar] [CrossRef]
- Kan, Y.H.; Gafforov, Y.; Li, T.; Zhou, L.W. Hyphodontia zhixiangii sp. nov. (Schizoporaceae, Basidiomycota) from Uzbekistan. Phytotaxa 2017, 299, 273–279. [Google Scholar] [CrossRef]
- Kan, Y.H.; Qin, W.M.; Zhou, L.W. Hyphodontia mollissima sp. nov. (Schizoporaceae, Hymenochaetales) from Hainan, southern China. Mycoscience 2017, 58, 297–301. [Google Scholar] [CrossRef]
- Wang, M.; Chen, Y.Y. Phylogeny and taxonomy of the genus Hyphodontia (Hymenochaetales, Basidiomycota) in China. Phytotaxa 2017, 309, 45–54. [Google Scholar] [CrossRef]
- Crous, P.W.; Luangsa-ard, J.J.; Wingfield, M.J.; Carnegie, A.J.; Hernández-Restrepo, M.; Lombard, L.; Roux, J.; Barreto, R.W.; Baseia, I.G.; Cano-Lira, J.F.; et al. Fungal Planet description sheets: 785–867. Persoonia 2018, 41, 238–417. [Google Scholar] [CrossRef] [PubMed]
- Viner, I.; Spirin, V.; Zíbarová, L.; Larsson, K.H. Additions to the taxonomy of Lagarobasidium and Xylodon (Hymenochaetales, Basidiomycota). MycoKeys 2018, 41, 65–90. [Google Scholar] [CrossRef] [PubMed]
- Fernández-López, J.; Telleria, M.T.; Dueñas, M.; Wilson, A.W.; Padamsee, M.; Buchanan, P.K.; Mueller, G.M.; Martin, M.P. Addressing the diversity of Xylodon raduloides complex through integrative taxonomy. IMA Fungus 2019, 10, 2. [Google Scholar] [CrossRef]
- Shi, Z.W.; Wang, X.W.; Zhou, L.W.; Zhao, C.L. Xylodon kunmingensis sp. nov. (Hymenochaetales, Basidiomycota) from southern China. Mycoscience 2019, 60, 184–188. [Google Scholar] [CrossRef]
- Chen, J.Z.; Zhao, C.L. Morphological and molecular identification of four new resupinate species of Lyomyces (Hymenochaetales) from southern China. MycoKeys 2020, 65, 101–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.W.; Jiang, J.H.; Zhou, L.W. Basidioradulum mayi and B. tasmanicum spp. nov. (Hymenochaetales, Basidiomycota) from both sides of Bass Strait, Australia. Sci. Rep. 2020, 10, 102. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth & Bisby’s Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008. [Google Scholar]
- He, M.Q.; Zhao, R.L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.C.; Raspé, O.; Kakishima, M.; Sánchez-Ramírez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar]
- Petersen, J.H. Farvekort. The Danish Mycological Society’s Colour Chart; Foreningen til Svampekundskabens Fremme: Greve, Denmark, 1996. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specifity for Basidiomycetes: Application to identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Matheny, P.B.; Wang, Z.; Binder, M.; Curtis, J.M.; Lim, Y.M.; Nilsson, R.H.; Hughes, K.W.; Hofstetter, V.; Ammirati, J.F.; Schoch, C.L.; et al. Contributions of rpb2 and tef1 to the phylogeny of mushrooms and allies (Basidiomycota, Fungi). Mol. Phylogenet. Evol. 2007, 43, 430–451. [Google Scholar] [CrossRef]
- Matheny, P.B.; Liu, Y.J.; Ammirati, J.F.; Hall, B.D. Using RPB1 sequences to improve phylogenetic inference among mushrooms (Inocybe, Agaricales). Am. J. Bot. 2002, 89, 688–698. [Google Scholar] [CrossRef]
- Liu, Y.L.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Matheny, P.B. Improving phylogenetic inference of mushrooms with RPB1 and RPB2 nucleotide sequences (Inocybe, Agaricales). Mol. Phylogenet. Evol. 2005, 35, 1–20. [Google Scholar] [CrossRef]
- Kretzer, A.M.; Bruns, T.D. Use of atp6 in fungal phylogenetics: An example from the Boletales. Mol. Phylogenet. Evol. 1999, 13, 483–492. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Kuma, K.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A simple, fast and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Posada, D. jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Pattengale, N.D.; Alipour, M.; Bininda-Emonds, O.R.P.; Moret, B.M.E.; Stamatakis, A. How many bootstrap replicates are necessary? J. Comput. Biol. 2010, 17, 337–354. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; Maio, N.D.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [Green Version]
- Hibbett, D.S.; Grimaldi, D.; Donoghue, M.J. Cretaceous mushrooms in amber. Nature 1995, 377, 487. [Google Scholar] [CrossRef]
- Hibbett, D.S.; Grimaldi, D.; Donoghue, M.J. Fossil mushrooms from Miocene and Cretaceous ambers and the evolution of Homobasidiomycetes. Am. J. Bot. 1997, 84, 981–991. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.Y.; Currah, R.S.; Stockey, R.A. Cretaceous and Eocene poroid hymenophores from Vancouver Island, British Columbia. Mycologia 2004, 96, 180–186. [Google Scholar] [CrossRef]
- Berbee, M.L.; Taylor, J.W. Dating the molecular clock in fungi – how close are we? Fungal Biol. Rev. 2010, 24, 1–16. [Google Scholar] [CrossRef]
- Taylor, T.N.; Hass, H.; Kerp, H. The oldest fossil ascomycetes. Nature 1999, 399, 648. [Google Scholar] [CrossRef] [PubMed]
- Taylor, T.N.; Hass, H.; Kerp, H.; Krings, M.; Hanlin, R.T. Perithecial ascomycetes from the 400 million year old Rhynie chert: An example of ancestral polymorphism. Mycologia 2005, 97, 269–285. [Google Scholar] [CrossRef] [PubMed]
- Floudas, D.; Binder, M.; Riley, R.; Barry, K.; Blanchette, R.A.; Henrissat, B.; Martínez, A.T.; Otillar, R.; Spatafora, J.W.; Yadav, J.S.; et al. The Paleozoic origin of enzymatic lignin decomposition reconstructed from 31 fungal genomes. Science 2012, 336, 1715–1719. [Google Scholar] [CrossRef] [Green Version]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evolution. 2018, 4, vey016. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Harris, A.J.; Blair, C.; He, X. RASP (Reconstruct Ancestral State in Phylogenies): A tool for historical biogeography. Mol. Phylogenet. Evol. 2015, 87, 46–49. [Google Scholar] [CrossRef]
- Yu, Y.; Blair, C.; He, X.J. RASP 4: Ancestral state reconstruction tool for multiple genes and characters. Mol. Biol. Evol. 2020, 37, 604–606. [Google Scholar] [CrossRef]
- Wu, S.H. Hyphodontia tubuliformis, a new species from Taiwan. Mycotaxon 2006, 95, 185–188. [Google Scholar]
- Rick, J.E. Basidiomycetes Eubasidii in Rio Grande do Sul - Brasilia. 3. Hypochnaceae, Clavariaceae, Craterellaceae, Hydnaceae. Iheringia 1959, 5, 125–192. [Google Scholar]
- Chen, J.J.; Zhou, L.W.; Ji, X.H.; Zhao, C.L. Hyphodontia dimitica and H. subefibulata spp. nov. (Schizoporaceae, Hymenochaetales) from southern China based on morphological and molecular characters. Phytotaxa 2016, 269, 1–13. [Google Scholar] [CrossRef]
- Giraud, T.; Refrégier, G.; Gac, M.L.; de Viennea, D.M.; Hood, M.E. Speciation in fungi. Fungal Genet. Biol. 2008, 45, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Yurchenko, E.; Xiong, H.X.; Wu, S.H. Hyphodontia juniperi (Basidiomycota) newly recorded from China. Fungal Sci. 2013, 28, 45–53. [Google Scholar]
- Hjortstam, K. A check-list to genera and species of corticioid fungi (Hymenomycetes). Windahlia 1987, 17, 55–85. [Google Scholar]
- Dhingra, G.S. Diversity of resupinate, non-poroid agaricomycetous fungi in the Himalaya and adjoining areas. In Proceedings of the 8th International Conference on Mushroom Biology and Mushroom Products (ICMBMP8); Singh, M., Ed.; Yugantar Prakashan Pvt. Ltd.: New Delhi, India, 2014; pp. 24–41. [Google Scholar]
- Hjortstam, K. Two new genera and some new combinations of corticioid fungi (Basidiomycotina, Aphyllophorales) from tropical and subtropical areas. Mycotaxon 1995, 54, 183–193. [Google Scholar]
- Greslebin, A.G.; Rajchenberg, M.; Bianchinotti, M.V. On Hyphotontia australis (Corticiaceae, Basidiomycota). Mycotaxon 2000, 74, 37–43. [Google Scholar]
- Yurchenko, E.; Xiong, H.X.; Wu, S.H. Four new species of Hyphodontia (Xylodon s.s. Hjortstam & Ryvarden, Basidiomycota) from Taiwan. Nova Hedwigia 2013, 96, 545–559. [Google Scholar]
- Zhao, C.L.; Cui, B.K.; Dai, Y.C. Morphological and molecular identification of two new species of Hyphodontia (Schizoporaceae, Hymenochaetales) from southern China. Cryptogam. Mycol. 2014, 35, 87–97. [Google Scholar] [CrossRef]
- Paulus, B.; Hallenberg, N.; Buchanan, P.K.; Chambers, G.K. A phylogenetic study of the genus Schizopora (Basidiomycota) based on ITS DNA sequences. Mycol. Res. 2000, 104, 1155–1163. [Google Scholar] [CrossRef]
- Grewe, F.; Huang, J.P.; Leavitt, S.D.; Lumbsch, H.T. Reference-based RADseq resolves robust relationships among closely related species of lichen-forming fungi using metagenomic DNA. Sci. Rep. 2017, 7, 9884. [Google Scholar] [CrossRef] [Green Version]
- Hallenberg, N.; Hjortstam, K. Four new species of corticioid fungi (Basidiomycotina, Aphyllophorales) from Argentina. Mycotaxon 1996, 57, 117–123. [Google Scholar]
- Frey, W.; Hurka, K.; Oberwinkler, F. Beiträge zur Biologie der Niederen Pflanzen; Systematik, Stammesgeschichte, Ökologie; Gustav Fischer Verlag: Stuttgart, Germany; New York, NY, USA, 1977. [Google Scholar]
- Zmitrovich, I.V.; Malysheva, V.F. Studies on Oxyporus. I. Segregation of Emmia and general topology of phylogenetic tree. Mikol. Fitopatol. 2014, 48, 161–171. [Google Scholar]
- Ariyawansa, H.A.; Hyde, K.D.; Jayasiri, S.C.; Buyck, B.; Chethana, K.W.T.; Dai, D.Q.; Dai, Y.C.; Daranagama, D.A.; Jayawardena, R.S.; Lücking, R.; et al. Fungal diversity notes 111–252. Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2015, 75, 27–274. [Google Scholar] [CrossRef]
- Zhou, L.W.; Wang, X.W.; Vlasák, J.; Ren, G.J. Resolution of phylogenetic position of Nigrofomitaceae within Hymenochaetales (Basidiomycota) and Nigrofomes sinomelanoporus sp. nov. (Nigrofomitaceae) from China. MycoKeys 2018, 29, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.C. Hymenochaetaceae (Basidiomycota) in China. Fungal Divers. 2010, 45, 131–343. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. International Code of Nomenclature for algae, fungi, and plants (Shenzhen Code) adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017; Koeltz Botanical Books: Glashütten, Germany, 2018. [Google Scholar]
- Korotkin, H.B.; Swenie, R.A.; Miettinen, O.; Budke, J.M.; Chen, K.-H.; Lutzoni, F.; Smith, M.E.; Matheny, P.B. Stable isotope analyses reveal previously unknown trophic mode diversity in the Hymenochaetales. Am. J. Bot. 2018, 105, 1869–1887. [Google Scholar] [CrossRef]
- Miettinen, O.; Larsson, K.H. Sidera, a new genus in Hymenochaetales with poroid and hydnoid species. Mycol. Prog. 2011, 10, 131–141. [Google Scholar] [CrossRef]
- Zhao, R.L.; Li, G.J.; Sánchez-Ramírez, S.; Stata, M.; Yang, Z.L.; Wu, G.; Dai, Y.C.; He, S.H.; Cui, B.K.; Zhou, J.L.; et al. A six-gene phylogenetic overview of Basidiomycota and allied phyla with estimated divergence times of higher taxa and a phyloproteomics perspective. Fungal Divers. 2017, 84, 43–74. [Google Scholar] [CrossRef]
- Lücking, R. Stop the abuse of time! Strict temporal banding is not the future of rank-based classifications in fungi (including lichens) and other organisms. CRC. Crit. Rev. Plant. Sci. 2019, 38, 199–253. [Google Scholar] [CrossRef]
- Budd, G.E.; Mann, R.P. The dynamics of stem and crown groups. Sci. Adv. 2020, 6, eaaz1626. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.K.; Hyde, K.D.; Jeewon, R.; Phillips, A.J.L.; Maharachchikumbura, S.S.N.; Ryberg, M.; Liu, Z.Y.; Zhao, Q. Ranking higher taxa using divergence times: A case study in Dothideomycetes. Fungal Divers. 2017, 84, 75–99. [Google Scholar] [CrossRef]
- Wu, F.; Chen, J.J.; Ji, X.H.; Vlasák, J.; Dai, Y.C. Phylogeny and diversity of the morphologically similar polypore genera Rigidoporus, Physisporinus, Oxyporus, and Leucophellinus. Mycologia 2017, 109, 749–765. [Google Scholar] [CrossRef] [PubMed]
- Nagy, L.G.; Tóth, R.; Kiss, E.; Slot, J.; Gácser, A.; Kovács, G.M. Six key traits of fungi: Their evolutionary origins and genetic bases. Microbiol. Spectr. 2017, 5, FUNK-0036-2016. [Google Scholar] [CrossRef] [Green Version]
- Hibbett, D.S.; Binder, M. Evolution of complex fruiting-body morphologies in homobasidiomycetes. Proc. R. Soc. Lond. B Biol. Sci. 2002, 269, 1963–1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hibbett, D.S. Trends in morphological evolution in homobasidiomycetes inferred using maximum likelihood: A comparison of binary and multistate approaches. Syst. Biol. 2004, 53, 889–903. [Google Scholar] [CrossRef] [PubMed]
- Varga, T.; Krizsán, K.; Földi, C.; Dima, B.; Sánchez-García, M.; Sánchez-Ramírez, S.; Szöllősi, G.J.; Szarkándi, G.J.; Papp, V.; Albert, L.; et al. Megaphylogeny resolves global patterns of mushroom evolution. Nat. Ecol. Evol. 2019, 3, 668–678. [Google Scholar] [CrossRef] [Green Version]












































Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.-W.; May, T.W.; Liu, S.-L.; Zhou, L.-W. Towards a Natural Classification of Hyphodontia Sensu Lato and the Trait Evolution of Basidiocarps within Hymenochaetales (Basidiomycota). J. Fungi 2021, 7, 478. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7060478
Wang X-W, May TW, Liu S-L, Zhou L-W. Towards a Natural Classification of Hyphodontia Sensu Lato and the Trait Evolution of Basidiocarps within Hymenochaetales (Basidiomycota). Journal of Fungi. 2021; 7(6):478. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7060478
Chicago/Turabian StyleWang, Xue-Wei, Tom W. May, Shi-Liang Liu, and Li-Wei Zhou. 2021. "Towards a Natural Classification of Hyphodontia Sensu Lato and the Trait Evolution of Basidiocarps within Hymenochaetales (Basidiomycota)" Journal of Fungi 7, no. 6: 478. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7060478