
Prediction of Genetic Groups within Brettanomyces bruxellensis through Cell Morphology Using a Deep Learning Tool

Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Isolates and Strains
2.2. Genetic Analysis
2.2.1. DNA Extraction
2.2.2. RAPD-PCR
2.3. Cell Polymorphism Analysis
2.3.1. Cultures
2.3.2. Microscopic Observations
2.3.3. Cell Shape Determination
2.3.4. Deep Learning
3. Results and Discussion
3.1. Intraspecific Discrimination of the B. bruxellensis Isolates into Genetic Groups
3.2. From Cell Polymorphism to Genetic Groups
3.2.1. Qualitative and Quantitative Description of Cell Morphology
3.2.2. Use of Deep Leaning to Predict the Genetic Group of B. bruxellensis Isolates
3.2.3. Does the Link between Genetic Groups and Cell Morphologies Predict any Specific Functions?
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chatonnet, P.; Dubourdie, D.; Boidron, J.; Pons, M. The Origin of Ethylphenols in Wines. J. Sci. Food Agric. 1992, 60, 165–178. [Google Scholar] [CrossRef]
- Wedral, D.; Shewfelt, R.; Frank, J. The Challenge of Brettanomyces in Wine. LWT Food Sci. Technol. 2010, 43, 1474–1479. [Google Scholar] [CrossRef]
- Fugelsang, K.C.; Zoecklein, B.W. Population Dynamics and Effects of Brettanomyces bruxellensis Strains on Pinot Noir (Vitis vinifera L.) Wines. Am. J. Enol. Vitic. 2003, 54, 294–300. [Google Scholar]
- Smith, B.D.; Divol, B. Brettanomyces bruxellensis, A Survivalist Prepared for the Wine Apocalypse and Other Beverages. Food Microbiol. 2016, 59, 161–175. [Google Scholar] [CrossRef]
- Avramova, M.; Vallet-Courbin, A.; Maupeu, J.; Masneuf-Pomarède, I.; Albertin, W. Molecular Diagnosis of Brettanomyces bruxellensis’ Sulfur Dioxide Sensitivity through Genotype Specific Method. Front. Microbiol. 2018, 9, 1260. [Google Scholar] [CrossRef]
- Conterno, L.; Joseph, C.M.L.; Arvik, T.J.; Henick-kling, T.; Bisson, L.F. Genetic and Physiological Characterization of Brettanomyces bruxellensis Strains Isolated from Wines. Am. J. Enol. Vitic. 2006, 57, 139–147. [Google Scholar]
- Longin, C.; Degueurce, C.; Julliat, F.; Guilloux-Benatier, M.; Rousseaux, S.; Alexandre, H. Efficiency of Population-Dependent Sulfite against Brettanomyces bruxellensis in Red Wine. Food Res. Int. 2016, 89, 620–630. [Google Scholar] [CrossRef]
- Serpaggi, V.; Remize, F.; Recorbet, G.; Gaudot-Dumas, E.; Sequeira-Le Grand, A.; Alexandre, H. Characterization of the “Viable but Nonculturable” (VBNC) State in the Wine Spoilage Yeast Brettanomyces. Food Microbiol. 2012, 30, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Rubio, P.; Garijo, P.; Santamaría, P.; López, R.; Martínez, J.; Gutierrez, A.R. Influence of Oak Origin and Ageing Conditions on Wine Spoilage by Brettanomyces Yeasts. Food Control 2015, 54, 176–180. [Google Scholar] [CrossRef]
- Suárez, R.; Suárez-Lepe, J.A.; Morata, A.; Calderón, F. The Production of Ethylphenols in Wine by Yeasts of the Genera Brettanomyces and Dekkera: A Review. Food Chem. 2007, 102, 10–21. [Google Scholar] [CrossRef]
- Lebleux, M.; Abdo, H.; Coelho, C.; Basmaciyan, L.; Albertin, W.; Maupeu, J.; Laurent, J.; Roullier-Gall, C.; Alexandre, H.; Guilloux-Benatier, M.; et al. New Advances on the Brettanomyces bruxellensis Biofilm Mode of Life. Int. J. Food Microbiol. 2020, 318, 108464. [Google Scholar] [CrossRef]
- Cibrario, A.; Avramova, M.; Dimopoulou, M.; Magani, M.; Miot-Sertier, C.; Mas, A.; Portillo, M.C.; Ballestra, P.; Albertin, W.; Masneuf-Pomarede, I.; et al. Brettanomyces bruxellensis Wine Isolates Show High Geographical Dispersal and Long Persistence in Cellars. PLoS ONE 2019, 14, e0222749. [Google Scholar] [CrossRef] [Green Version]
- Cartwright, Z.M.; Glawe, D.A.; Edwards, C.G. Reduction of Brettanomyces bruxellensis Populations from Oak Barrel Staves Using Steam. Am. J. Enol. Vitic. 2018, 69, 400–409. [Google Scholar] [CrossRef]
- Escudier, J.L.; Bes, M.; Salmon, J.M.; Caillé, D.; Samson, A. Stress Hydrique Prolongé Des Vignes: Comment Adapter Les Pratiques Œnologiques en Conséquence? Innov. Agron. 2014, 38, 67–86. [Google Scholar]
- Van Leeuwen, C. L’augmentation du pH des Moûts et des Vins, un Facteur Important de Développement de Brettanomyces. In Proceedings of the 13e Matinée des OEnologues de Bordeaux, Université de Bordeaux, Bordeaux, France, 20 March 2015. [Google Scholar]
- Lattey, K.A.; Bramley, B.R.; Francis, I.L. Consumer Acceptability, Sensory Properties and Expert Quality Judgements of Australian Cabernet Sauvignon and Shiraz Wines. Aust. J. Grape Wine Res. 2010, 16, 189–202. [Google Scholar] [CrossRef]
- Oelofse, A.; Malherbe, S.; Pretorius, I.S.; Du Toit, M. Preliminary Evaluation of Infrared Spectroscopy for the Differentiation of Brettanomyces bruxellensis Strains Isolated from Red Wines. Int. J. Food Microbiol. 2010, 143, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Vigentini, I.; De Lorenzis, G.; Picozzi, C.; Imazio, S.; Merico, A.; Galafassi, S.; Piškur, J.; Foschino, R. Intraspecific Variations of Dekkera/Brettanomyces bruxellensis Genome Studied by Capillary Electrophoresis Separation of the Intron Splice Site Profiles. Int. J. Food Microbiol. 2012, 157, 6–15. [Google Scholar] [CrossRef]
- Martorell, P.; Barata, A.; Malfeito-Ferreira, M.; Fernández-Espinar, M.T.; Loureiro, V.; Querol, A. Molecular Typing of the Yeast Species Dekkera bruxellensis and Pichia guilliermondii Recovered from Wine Related Sources. Int. J. Food Microbiol. 2006, 106, 79–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibeas, J.I.; Lozano, I.; Perdigones, F.; Jimenez, J. Detection of Dekkera-Brettanomyces Strains in Sherry by A Nested PCR Method. Appl. Environ. Microbiol. 1996, 62, 998–1003. [Google Scholar] [CrossRef] [Green Version]
- Miot-Sertier, C.; Lonvaud-Funel, A. Development of a Molecular Method for the Typing of Brettanomyces bruxellensis (Dekkera bruxellensis) at the Strain Level. J. Appl. Microbiol. 2007, 102, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Curtin, C.D.; Bellon, J.R.; Henschke, P.A.; Godden, P.W.; de Barros Lopes, M.A. Genetic Diversity of Dekkera bruxellensis Yeasts Isolated from Australian Wineries. FEMS Yeast Res. 2007, 7, 471–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Barros Lopes, M.; Rainieri, S.; Henschke, P.A.; Langridge, P. AFLP Fingerprinting for Analysis of Yeast Genetic Variation. Int. J. Syst. Bacteriol. 1999, 49, 915–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campolongo, S.; Rantsiou, K.; Giordano, M.; Gerbi, V.; Cocolin, L. Prevalence and Biodiversity of Brettanomyces bruxellensis in Wine from Northwestern Italy. Am. J. Enol. Vitic. 2010, 61, 486–491. [Google Scholar] [CrossRef]
- Corich, V.; Mattiazzi, A.; Soldati, E.; Carraro, A.; Giacomini, A. Sau-PCR, A Novel Amplification Technique for Genetic Fingerprinting of Microorganisms. Appl. Environ. Microbiol. 2005, 71, 6401–6406. [Google Scholar] [CrossRef] [Green Version]
- Vigentini, I.; Picozzi, C.; Foschino, R. Intron Splice Site PCR Analysis as a Tool to Discriminate Dekkera bruxellensis Strains. Ann. Microbiol. 2011, 61, 153–157. [Google Scholar] [CrossRef]
- Oelofse, A.; Lonvaud-Funel, A.; du Toit, M. Molecular Identification of Brettanomyces bruxellensis Strains Isolated from Red Wines and Volatile Phenol Production. Food Microbiol. 2009, 26, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Albertin, W.; Panfili, A.; Miot-Sertier, C.; Goulielmakis, A.; Delcamp, A.; Salin, F.; Lonvaud-Funel, A.; Curtin, C.; Masneuf-Pomarede, I. Development of Microsatellite Markers for the Rapid and Reliable Genotyping of Brettanomyces bruxellensis at Strain Level. Food Microbiol. 2014, 42, 188–195. [Google Scholar] [CrossRef]
- Avramova, M.; Cibrario, A.; Peltier, E.; Coton, M.; Coton, E.; Schacherer, J.; Spano, G.; Capozzi, V.; Blaiotta, G.; Salin, F.; et al. Brettanomyces bruxellensis Population Survey Reveals a Diploid-Triploid Complex Structured according to Substrate of Isolation and Geographical Distribution. Sci. Rep. 2018, 8, 4136. [Google Scholar] [CrossRef]
- Dimopoulou, M.; Hatzikamari, M.; Masneuf-Pomarede, I.; Albertin, W. Sulfur Dioxide Response of Brettanomyces bruxellensis Strains Isolated from Greek Wine. Food Microbiol. 2019, 78, 155–163. [Google Scholar] [CrossRef]
- Mitrakul, C.M.; Henick-Kling, T.; Egli, C.M. Discrimination of Brettanomyces/Dekkera Yeast Isolates from Wine by Using Various DNA Finger-Printing Methods. Food Microbiol. 1999, 16, 3–14. [Google Scholar] [CrossRef]
- Crauwels, S.; Zhu, B.; Steensels, J.; Busschaert, P.; De Samblanx, G.; Marchal, K.; Willems, K.A.; Verstrepen, K.J.; Lievens, B. Assessing Genetic Diversity among Brettanomyces Yeasts by DNA Fingerprinting and Whole-Genome Sequencing. Appl. Environ. Microbiol. 2014, 80, 4398–4413. [Google Scholar] [CrossRef] [Green Version]
- Agnolucci, M.; Vigentini, I.; Capurso, G.; Merico, A.; Tirelli, A.; Compagno, C.; Foschino, R.; Nuti, M. Genetic Diversity and Physiological Traits of Brettanomyces bruxellensis Strains Isolated from Tuscan Sangiovese Wines. Int. J. Food Microbiol. 2009, 130, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Godoy, L.; Garrido, D.; Martínez, C.; Saavedra, J.; Combina, M.; Ganga, M.A. Study of the Coumarate Decarboxylase and Vinylphenol Reductase Activities of Dekkera bruxellensis (anamorph Brettanomyces bruxellensis) Isolates. Lett. Appl. Microbiol. 2009, 48, 452–457. [Google Scholar] [CrossRef] [PubMed]
- G-Poblete, C.; Peña-Moreno, I.C.; de Morais, M.A.; Moreira, S.; Ganga, M.A. Biodiversity among Brettanomyces bruxellensis Strains Isolated from Different Wine Regions of Chile: Key Factors Revealed about Its Tolerance to Sulphite. Microorganisms 2020, 8, 557. [Google Scholar] [CrossRef]
- Oro, L.; Canonico, L.; Marinelli, V.; Ciani, M.; Comitini, F. Occurrence of Brettanomyces bruxellensis on Grape Berries and in Related Winemaking Cellar. Front. Microbiol. 2019, 10, 415. [Google Scholar] [CrossRef] [Green Version]
- Sangorrín, M.P.; García, V.; Lopes, C.A.; Sáez, J.S.; Martínez, C.; Ganga, M.A. Molecular and Physiological Comparison of Spoilage Wine Yeasts. J. Appl. Microbiol. 2013, 114, 1066–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quesada, M.P.; Cenis, J.L. Use of Random Amplified Polymorphic DNA (RAPD-PCR) in the Characterization of Wine Yeasts. Am. J. Enol. Vitic. 1995, 46, 204–208. [Google Scholar]
- Giraffa, G.; Carminati, D. Molecular Techniques in Food Fermentation: Principles and Applications. In Molecular Techniques in the Microbial Ecology of Fermented Foods; Cocolin, L., Ercolini, D., Eds.; Springer: New York, NY, USA, 2008; pp. 1–30. ISBN 978-0-387-74519-0. [Google Scholar]
- Moen, E.; Bannon, D.; Kudo, T.; Graf, W.; Covert, M.; Van Valen, D. Deep Learning for Cellular Image Analysis. Nat. Methods 2019, 16, 1233–1246. [Google Scholar] [CrossRef]
- Jones, W.; Alasoo, K.; Fishman, D.; Parts, L. Computational biology: Deep learning. Emerg. Top. Life Sci. 2017, 1, 133–150. [Google Scholar] [CrossRef] [Green Version]
- Kang, R.; Park, B.; Eady, M.; Ouyang, Q.; Chen, K. Single-Cell Classification of Foodborne Pathogens Using Hyperspectral Microscope Imaging Coupled with Deep Learning Frameworks. Sens. Actuators B Chem. 2020, 309, 127789. [Google Scholar] [CrossRef]
- Alzubaidi, L.; Fadhel, M.A.; Al-Shamma, O.; Zhang, J.; Duan, Y. Deep Learning Models for Classification of Red Blood Cells in Microscopy Images to Aid in Sickle Cell Anemia Diagnosis. Electronics 2020, 9, 427. [Google Scholar] [CrossRef] [Green Version]
- Longden, J.; Robin, X.; Engel, M.; Ferkinghoff-Borg, J.; Kjær, I.; Horak, I.D.; Pedersen, M.W.; Linding, R. Deep Neural Networks Identify Signaling Mechanisms of ErbB-Family Drug Resistance from a Continuous Cell Morphology Space. Cell Rep. 2021, 34, 108657. [Google Scholar] [CrossRef]
- Dickinson, J.R. Filament formation in Saccharomyces cerevisiae—A review. Folia Microbiol. 2008, 53, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Louw, M.; du Toit, M.; Alexandre, H.; Divol, B. Comparative Morphological Characteristics of Three Brettanomyces bruxellensis Wine Strains in the Presence/Absence of Sulfur Dioxide. Int. J. Food Microbiol. 2016, 238, 79–88. [Google Scholar] [CrossRef]
- Echeverrigaray, S.; Randon, M.; da Silva, K.; Zacaria, J.; Delamare, A.P.L. Identification and Characterization of Non-Saccharomyces Spoilage Yeasts Isolated from Brazilian Wines. World J. Microbiol. Biotechnol. 2013, 29, 1019–1027. [Google Scholar] [CrossRef]
- Aguilar-Uscanga, M.G.; Delia, M.-L.; Strehaiano, P. Nutritional Requirements of Brettanomyces bruxellensis: Growth and Physiology in Batch and Chemostat Cultures. Can. J. Microbiol. 2000, 46, 1046–1050. [Google Scholar] [CrossRef]
- Vigentini, I.; Lucy Joseph, C.M.; Picozzi, C.; Foschino, R.; Bisson, L.F. Assessment of the Brettanomyces bruxellensis Metabolome during Sulphur Dioxide Exposure. FEMS Yeast Res. 2013, 13, 597–608. [Google Scholar] [CrossRef] [Green Version]
- Oelofse, A. Investigating the Role of Brettanomyces and Dekkera during Winemaking. Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, December 2008. [Google Scholar]
- Lentz, M.; Putzke, T.; Hessler, R.; Luman, E. Genetic and Physiological Characterization of Yeast Isolated from Ripe Fruit and Analysis of Fermentation and Brewing Potential. J. Inst. Brew. 2014, 120, 559–564. [Google Scholar] [CrossRef]
- Giaever, G.; Chu, A.M.; Ni, L.; Connelly, C.; Riles, L.; Véronneau, S.; Dow, S.; Lucau-Danila, A.; Anderson, K.; André, B.; et al. Functional Profiling of the Saccharomyces cerevisiae Genome. Nature 2002, 418, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Kumar, A. An Overview of Autophagy and Yeast Pseudohyphal Growth: Integration of Signaling Pathways during Nitrogen Stress. Cells 2012, 1, 263–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullen, P.J.; Sprague, G.F. The Regulation of Filamentous Growth in Yeast. Genetics 2012, 190, 23–49. [Google Scholar] [CrossRef] [PubMed]
- Cullen, P.J.; Sprague, G.F. Glucose Depletion Causes Haploid Invasive Growth in Yeast. Proc. Natl. Acad. Sci. USA 2000, 97, 13619–13624. [Google Scholar] [CrossRef] [Green Version]
- Kuzdzal-Fick, J.J.; Chen, L.; Balázsi, G. Disadvantages and Benefits of Evolved Unicellularity Versus Multicellularity in Budding Yeast. Ecol. Evol. 2019, 9, 8509–8523. [Google Scholar] [CrossRef]
- Smukalla, S.; Caldara, M.; Pochet, N.; Beauvais, A.; Guadagnini, S.; Yan, C.; Vinces, M.D.; Jansen, A.; Prevost, M.C.; Latgé, J.-P.; et al. FLO1 Is a Variable Green Beard Gene That Drives Biofilm-Like Cooperation in Budding Yeast. Cell 2008, 135, 726–737. [Google Scholar] [CrossRef] [Green Version]
- Opalek, M.; Wloch-Salamon, D. Aspects of Multicellularity in Saccharomyces cerevisiae Yeast: A Review of Evolutionary and Physiological Mechanisms. Genes 2020, 11, 690. [Google Scholar] [CrossRef] [PubMed]
- Goossens, K.V.Y.; Ielasi, F.S.; Nookaew, I.; Stals, I.; Alonso-Sarduy, L.; Daenen, L.; Van Mulders, S.E.; Stassen, C.; van Eijsden, R.G.E.; Siewers, V.; et al. Molecular Mechanism of Flocculation Self-Recognition in Yeast and Its Role in Mating and Survival. mBio 2015, 6, e00427-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
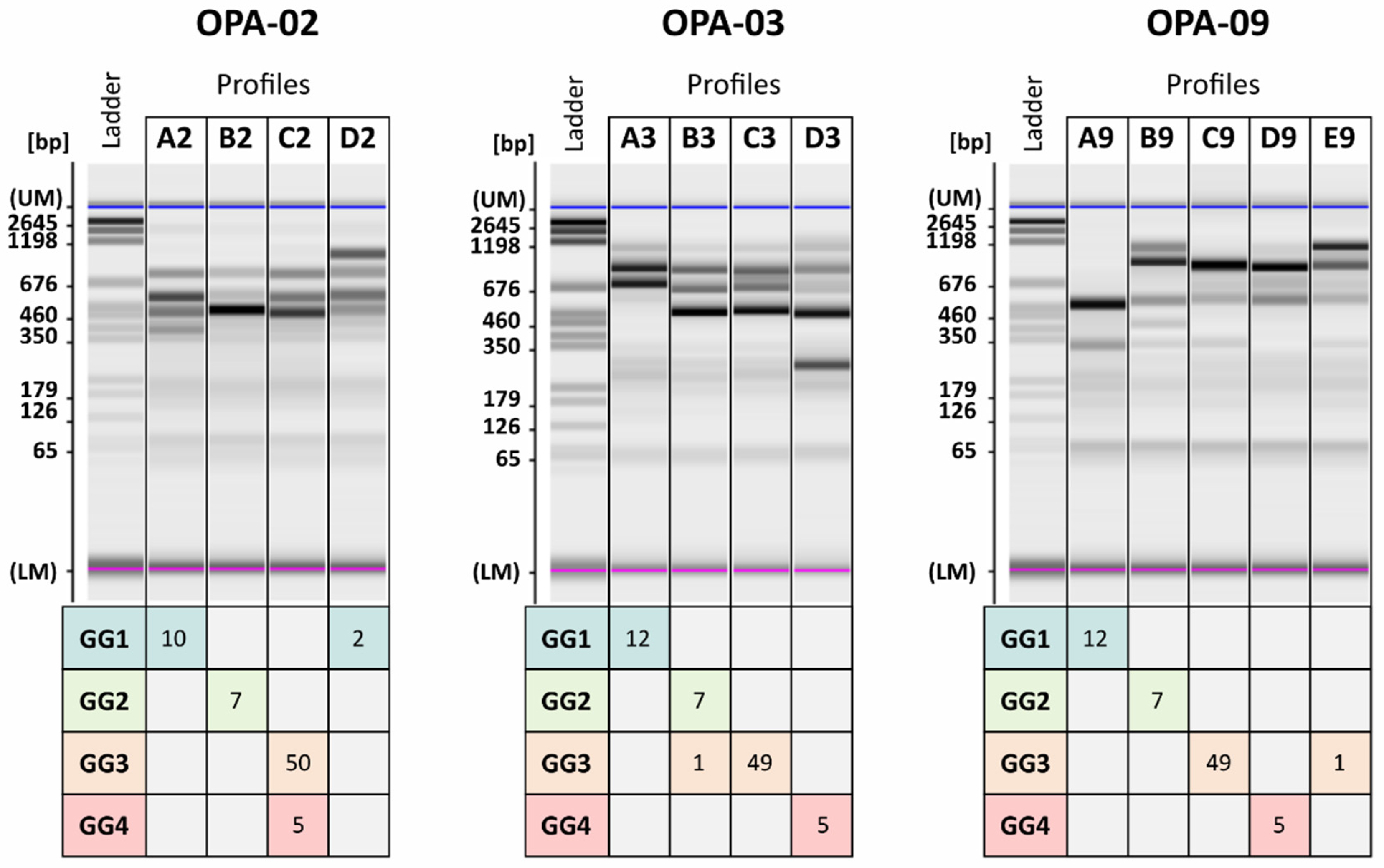
| Isolate/Strain of Reference | Genetic Group (GG) |
|---|---|
| 1, 14, 25, 26, 27, 30, 49, 61, 62, LO2E2, CDR3, CDR12 | GG1 |
| 2, 4, 6, 11, 17, 19, 20 | GG2 |
| 3, 5, 7, 8, 9, 10, 12, 13, 15, 16, 18, 21, 22, 23, 24, 28, 29, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 64, LO6/036, CDR9, CDR11 | GG3 |
| 63, CDR217, CDR219, LO417, AWRI1499 | GG4 |
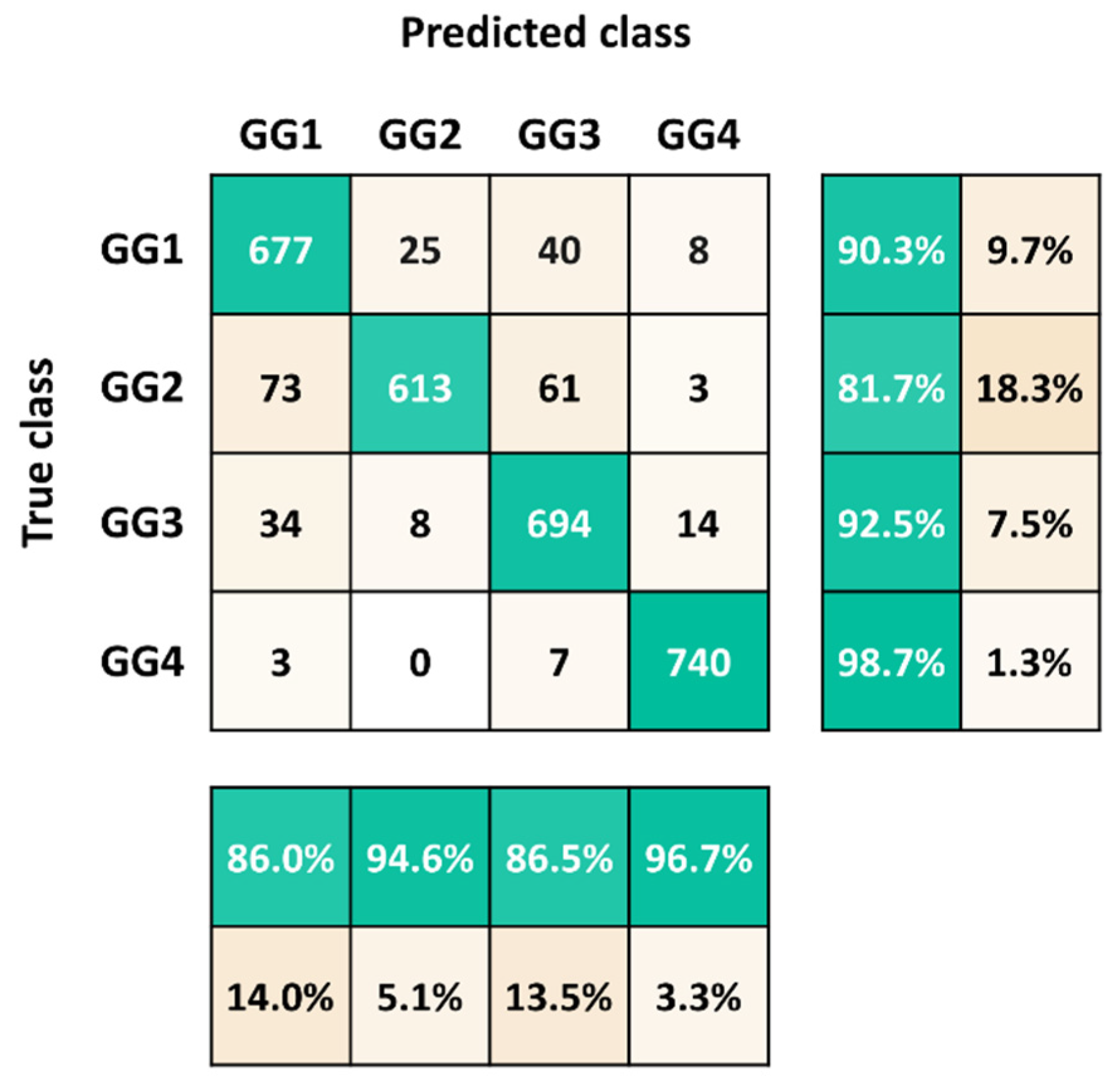
| Accuracy | Error | Sensitivity | Specificity | Precision | FalsePositive Rate | F1-Score |
|---|---|---|---|---|---|---|
| 0.9080 | 0.0920 | 0.9080 | 0.9693 | 0.9105 | 0.0307 | 0.9076 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lebleux, M.; Denimal, E.; De Oliveira, D.; Marin, A.; Desroche, N.; Alexandre, H.; Weidmann, S.; Rousseaux, S. Prediction of Genetic Groups within Brettanomyces bruxellensis through Cell Morphology Using a Deep Learning Tool. J. Fungi 2021, 7, 581. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080581
Lebleux M, Denimal E, De Oliveira D, Marin A, Desroche N, Alexandre H, Weidmann S, Rousseaux S. Prediction of Genetic Groups within Brettanomyces bruxellensis through Cell Morphology Using a Deep Learning Tool. Journal of Fungi. 2021; 7(8):581. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080581
Chicago/Turabian StyleLebleux, Manon, Emmanuel Denimal, Déborah De Oliveira, Ambroise Marin, Nicolas Desroche, Hervé Alexandre, Stéphanie Weidmann, and Sandrine Rousseaux. 2021. "Prediction of Genetic Groups within Brettanomyces bruxellensis through Cell Morphology Using a Deep Learning Tool" Journal of Fungi 7, no. 8: 581. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080581