
Influence of Interactions between Nitrogen, Phosphorus Supply and Epichloёbromicola on Growth of Wild Barley (Hordeum brevisubulatum)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Experimental Design
2.2.1. Preparation of Nutrient Solution
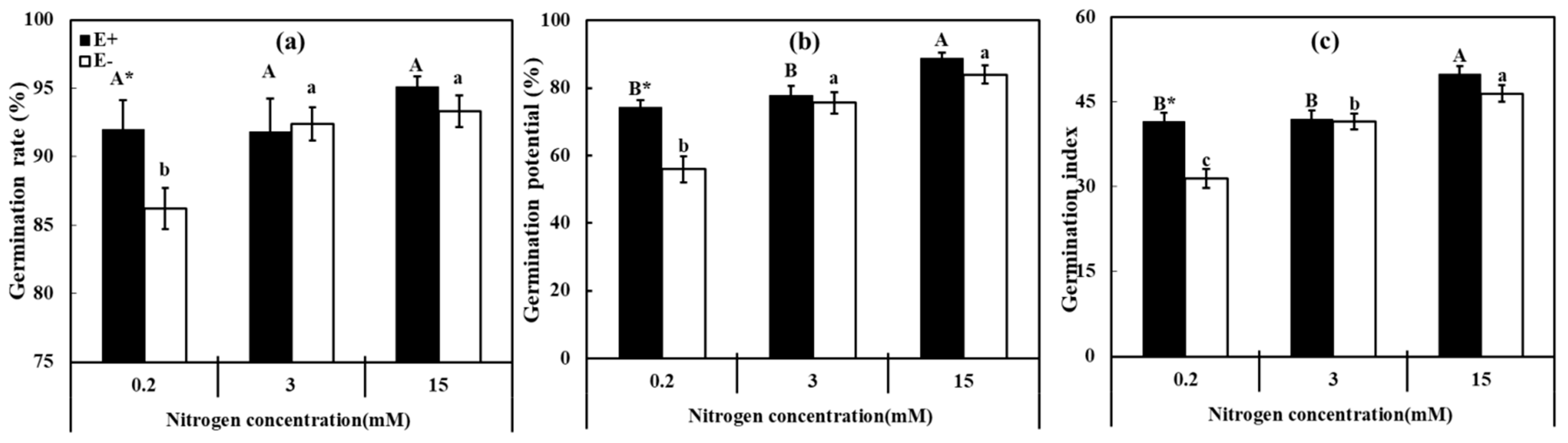
2.2.2. Seed Germination Experiment
2.2.3. Hydroponic Experiment
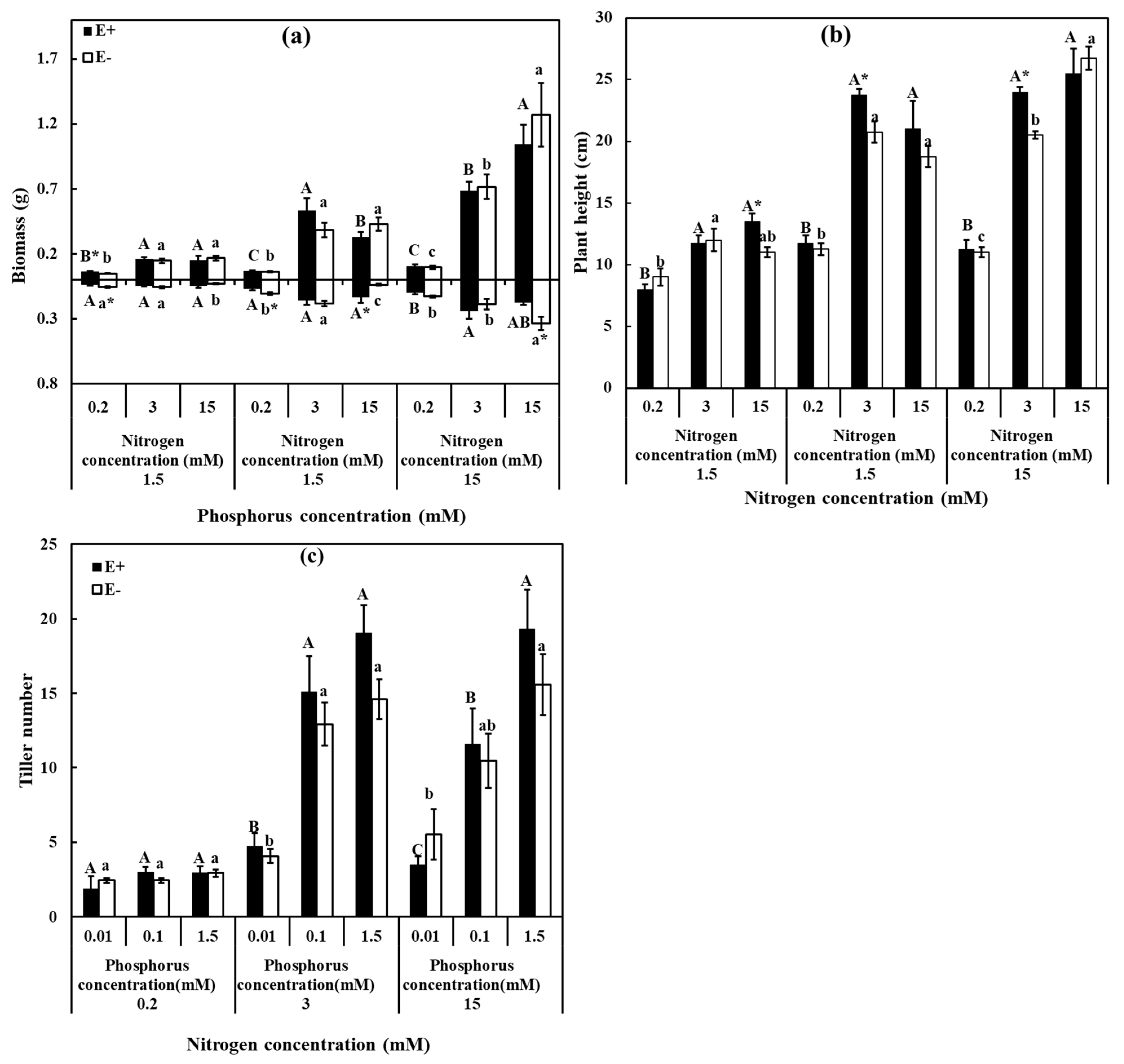
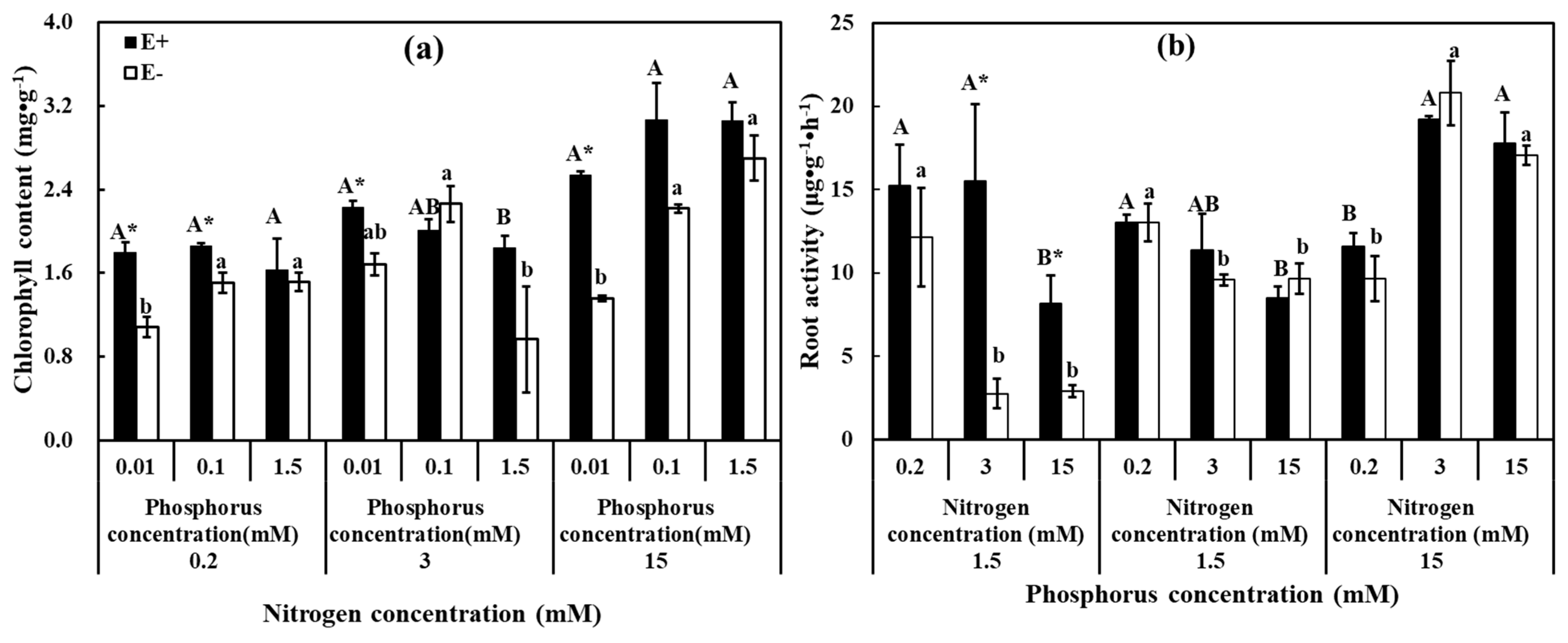
2.2.4. Plant Biomass and Physiological Indexes
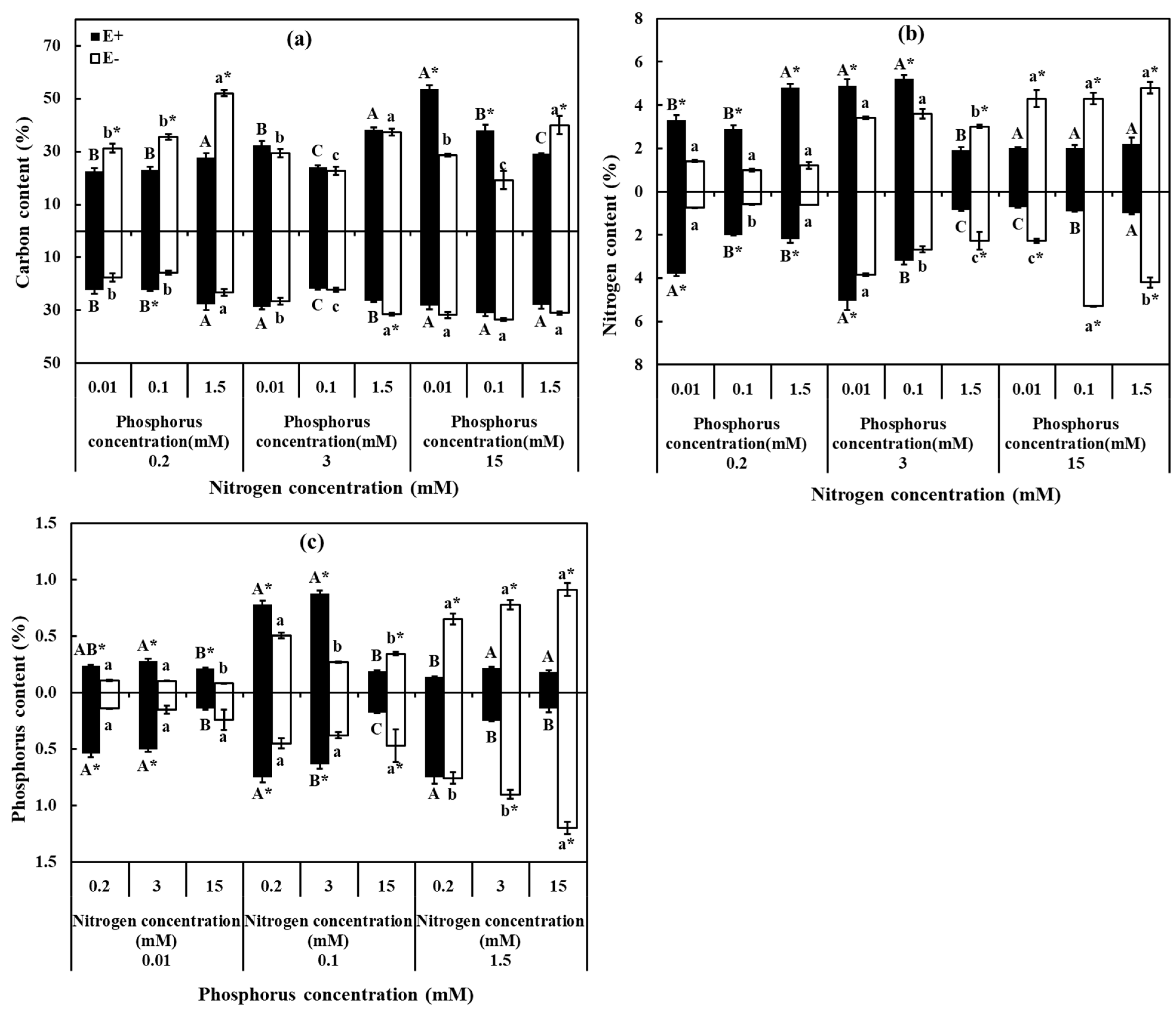
2.2.5. Chemical Analysis
2.3. Statistical Analysis
3. Results
3.1. Seed Germination
3.2. Growth and Physiological Parameters
3.3. C, N, and P Contents
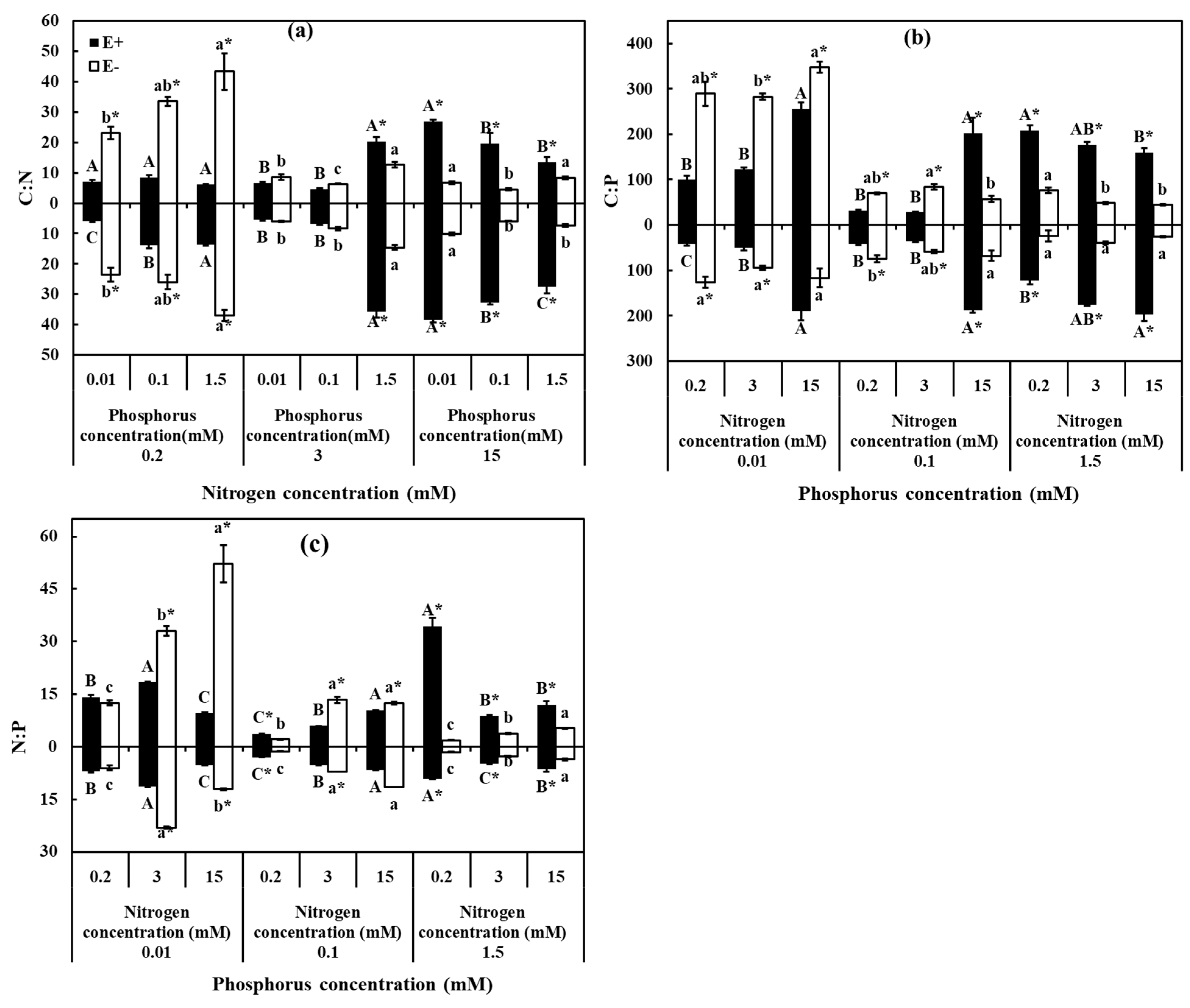
3.4. Stoichiometric Ratios of C, N, P
3.5. Na+, K+ Contents
4. Discussion
4.1. Effect of Epichloë Endophyte Infection on Seed Germination and Growth of Wild Barley
4.2. Effect of N and P Supplements on Seed Germination and Growth of Wild Barley
4.3. Effect of Interaction of N, P Supplements and Epichloë Endophyte Infection on Seed Germination and Growth of Wild Barley
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wei, N.; Song, L.R.; Gan, N.Q. Quantitative proteomic and microcystin production response of microcystis aeruginosa to phosphorus depletion. Microorganisms 2021, 9, 1183. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Peng, Q.; Li, K.; Liu, Y.; Gong, Y.; Han, W. Patterns of nitrogen and phosphorus stoichiometry among leaf, stem and root of desert plants and responses to climate and soil factors in Xinjiang, China. Catena 2021, 199, 105100. [Google Scholar] [CrossRef]
- Elser, J.J.; Sterner, R.; Gorokhova, E.; Fagan, W.; Markow, T.; Cotner, J.; Harrison, J.; Hobbie, S.; Odell, G.; Weider, L. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2000, 3, 540–550. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Chai, Q.; Li, X.; Yao, X.; Li, C.; Christensen, M.; Nan, Z. An asexual Epichloë endophyte modifies the nutrient stoichiometry of wild barley (Hordeum brevisubulatum) under salt stress. Plant Soil 2015, 387, 153–165. [Google Scholar] [CrossRef]
- Güsewell, S. N: P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2010, 164, 243–266. [Google Scholar] [CrossRef] [PubMed]
- Hessen, D.O.; Jensen, T.C.; Kyle, M.; Elser, J.J. RNA responses to N- and P-limitation; reciprocal regulation of stoichiometry and growth rate in Brachionus. Funct. Ecol. 2007, 21, 956–962. [Google Scholar] [CrossRef]
- Pomraning, K.R.; Bredeweg, E.L.; Baker, S.E.; Mitchell, A.P. Regulation of nitrogen metabolism by GATA zinc finger transcription factors in Yarrowia lipolytica. Msphere 2017, 2, e00038-17. [Google Scholar] [CrossRef] [Green Version]
- Malinowski, D.P.; Belesky, D.P.; Hill, N.S.; Baligar, V.C.; Fedders, J.M. Influence of phosphorus on the growth and ergot alkaloid content of Neotyphodium coenophialum-infected tall fescue (Festuca arundinacea Schreb.). Plant Soil 1998, 198, 53–61. [Google Scholar] [CrossRef]
- Hallasgo, A.M.; Spangl, B.; Steinkellner, S.; Hage-Ahmed, K. The fungal endophyte Serendipita williamsii does not affect phosphorus status but carbon and nitrogen dynamics in arbuscular mycorrhizal tomato plants. J. Fungi 2020, 6, 233. [Google Scholar] [CrossRef]
- Nan, Z.B.; Li, C.J. Roles of the grass-Neotyphodium association in pastoral agriculture systems. Acta Ecol. Sin. 2004, 24, 605–616. [Google Scholar]
- Xia, Z.W.; Yang, J.Y.; Sang, C.P.; Wang, X.; Sun, L.F.; Jiang, P.; Wang, C.; Bai, E. Phosphorus reduces negative effects of nitrogen addition on soil microbial communities and functions. Microorganisms 2020, 8, 1828. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Chang, Y.; Hussain, M.; Lu, B.; Zhang, J.; Song, X.; Lei, X.; Pei, D. Soil chemical and microbiological properties are changed by long-term chemical fertilizers that limit ecosystem functioning. Microorganisms 2020, 8, 694. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.J.; Jin, Y.Y.; Yao, X.; Chen, T.X.; Nan, Z.B. Fungal endophyte improves survival of Lolium perenne in low fertility soils by increasing root growth, metabolic activity and absorption of nutrients. Plant Soil 2020, 452, 185–206. [Google Scholar] [CrossRef]
- Chen, Z.J.; Jin, Y.Y.; Yao, X.; Wei, X.K.; Li, X.Z.; Li, C.J.; White, J.F.; Nan, Z.B. Gene analysis reveals that leaf litter from Epichloë endophyte-infected perennial ryegrass alters diversity and abundance of soil microbes involved in nitrification and denitrification. Soil Biol. Biochem. 2021, 154, 108123. [Google Scholar] [CrossRef]
- Schardl, C.L.; Leuchtmann, A.; Spiering, M.J. Symbioses of grasses with seedborne fungal endophytes. Annu. Rev. Plant Biol. 2004, 55, 315–340. [Google Scholar] [CrossRef]
- Chen, Z.J.; Li, C.J.; Nan, Z.B.; Wei, X.K.; White, J.F. Segregation of Lolium perenne into a subpopulation with high infection by endophyte Epichloë festucae var. lolii results in improved agronomic performance. Plant Soil 2020, 446, 595–612. [Google Scholar] [CrossRef]
- Wang, Z.F.; Li, C.J.; White, J.F. Effects of Epichloë endophyte infection on growth, physiological properties and seed germination of wild barley under saline conditions. J. Agron. Crop Sci. 2020, 206, 43–51. [Google Scholar] [CrossRef]
- White, J.F.; Kingsley, K.L.; Verma, S.K.; Kowalski, K.P. Rhizophagy cycle: An oxidative process in plants for nutrient extraction from symbiotic microbes. Microorganisms 2018, 6, 95. [Google Scholar] [CrossRef] [Green Version]
- Ren, A.Z.; Gao, Y.B.; Wang, W.; Wang, J.L.; Zhao, N.X. Influence of nitrogen fertilizer and endophyte infection on ecophysiological parameters and mineral element content of Perennial Ryegrass. J. Integr. Plant Biol. 2009, 51, 75–83. [Google Scholar] [CrossRef]
- Marks, S.; Clay, K. Low resource availability differentially affects the growth of host grasses infected by fungal endophytes. Int. J. Plant Sci. 2007, 168, 1269–1277. [Google Scholar] [CrossRef]
- Yin, C.Y.; Pu, X.Z.; Xiao, Q.Y.; Zhao, C.Z.; Liu, Q. Effects of night warming on spruce root around non-growing season vary with branch order and month. Plant Soil 2014, 380, 249–263. [Google Scholar] [CrossRef] [Green Version]
- Lambers, H.; Raven, J.A.; Shaver, G.R.; Smith, S.E. Plant nutrient-acquisition strategies change with soil age. Trends Ecol. Evol. 2008, 23, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-De-Aldana, B.R.; Antonia, G.C.; Balbino, G.C.; Santiago, V.T.; Igo, I.Z.; Raffaella, B. Fungal endophyte (Epichloë festucae) alters the nutrient content of Festuca rubra regardless of water availability. PLoS ONE 2013, 8, e84539. [Google Scholar]
- Bihari, B.; Kumari, R.; Padbhushan, R.; Shambhavi, S.; Kumar, R. Impact of inorganic and organic sources on bio-growth and nutrient accumulation in tomato crop cv. H-86 (Kashi Vishesh). J. Pharmacogn. Phytochem. 2018, 7, 756–760. [Google Scholar]
- Zhang, J.J.; Yan, X.B.; Su, F.L.; Li, Z.; Wang, Y.; Wei, Y.N.; Ji, Y.G.; Yang, Y.; Zhou, X.H.; Guo, H.; et al. Long-term N and P additions alter the scaling of plant nitrogen to phosphorus in a Tibetan alpine meadow. Sci. Total Environ. 2018, 625, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Sterner, R.W.; Elser, J.J. Ecological stoichiometry: The biology of elements from molecules to the biosphere. Nature 2002, 423, 225–226. [Google Scholar]
- Wang, J.F.; Nan, Z.B.; Christensen, M.J.; Li, C.J. Glucose-6-phosphate dehydrogenase plays a vital role in Achnatherum inebrians plants host to Epichloë gansuensis by improving growth under nitrogen deficiency. Plant Soil 2018, 430, 37–48. [Google Scholar] [CrossRef]
- Fujita, Y.; Robroek, B.; Ruiter, P.; Heil, G.W.; Wassen, M.J. Increased N affects P uptake of eight grassland species: The role of root surface phosphatase activity. Oikos 2010, 119, 1665–1673. [Google Scholar] [CrossRef]
- Vanderwoude, B.; Pegtel, D.; Bakker, J. Nutrient limitation after long-term nitrogen fertilizer application in cut grasslands. J. Appl. Ecol. 1994, 31, 405–412. [Google Scholar]
- Elser, J.J.; Bracken, M.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.X.; Li, C.; White, J.F.; Nan, Z. Effect of the fungal endophyte Epichloë bromicola on polyamines in wild barley (Hordeum brevisubulatum) under salt stress. Plant Soil 2019, 436, 29–48. [Google Scholar] [CrossRef]
- Chen, Z.J.; Wei, X.K.; Ying, C.; Pei, T.; Zhao, X.J.; Li, C.J. Research progress of methods on grass fungal endophyte detection. Pratacult. Sci. 2017, 34, 1419–1433. [Google Scholar]
- Lieffering, M.; Andrews, M.; Mckenzie, B.A. Nitrate stimulation of mobilization of seed reserves in temperate cereals: Importance of water uptake. Ann. Bot. 1996, 78, 695–701. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Choi, J.H.; Kim, W.S.; Han, T.H.; Park, Y.S.; Gemma, H. Effect of soil water stress on the development of stone cells in pear (Pyrus pyrifolia cv. ‘Niitaka’) flesh. Sci. Hortic. 2006, 110, 247–253. [Google Scholar] [CrossRef]
- Tanveer, S.K.; Zhang, J.; Lu, X.; Wen, X.; Liao, Y.C. Effect of corn residue mulch and N fertilizer application on nitrous oxide (N2O) emission and wheat crop productivity under rain-fed condition of loess plateau China. Int. J. Agric. Biol. 2014, 16, 505–512. [Google Scholar]
- Chen, N.; He, R.L.; Chai, Q.; Li, C.J.; Nan, Z.B. Transcriptomic analyses giving insights into molecular regulation mechanisms involved in cold tolerance by Epichloë endophyte in seed germination of Achnatherum inebrians. Plant Growth Regul. 2016, 80, 367–375. [Google Scholar] [CrossRef]
- Xu, Q.; Huang, B. Seasonal changes in root metabolic activity and nitrogen uptake for two cultivars of creeping bentgrass. Hortscience 2006, 41, 822–826. [Google Scholar] [CrossRef] [Green Version]
- Hikosaka, K. Interspecific difference in the photosynthesis-nitrogen relationship: Patterns, physiological causes and ecological importance. J. Plant Res. 2004, 117, 481–494. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Zhou, Y.F.; Zhang, X.X.; Duan, T.Y.; Nan, Z.B. Effects of Epichloë endophyte on antioxidant enzymes activities, photosynthesis and growth of three ecotypes of Elymus dahuricus. FASE 2018, 5, 148–158. [Google Scholar] [CrossRef]
- Chen, T.X.; Richard, J.; Chen, S.H.; Lv, H.; Zhou, J.L.; Li, C.J. Infection by the fungal endophyte Epichloë bromicola enhances the tolerance of wild barley (Hordeum brevisubulatum) to salt and alkali stresses. Plant Soil 2018, 428, 353–370. [Google Scholar] [CrossRef]
- Wang, J.F.; Hou, W.P.; Christensen, M.J.; Li, X.Z.; Nan, Z.B. Role of Epichloë endophytes in improving host grass resistance ability and soil properties. J. Agric. Food Chem. 2020, 68, 6944–6955. [Google Scholar] [CrossRef]
- Rudgers, J.A.; Mattingly, W.B.; Koslow, J.M. Mutualistic fungus promotes plant invasion into diverse communities. Oecologia 2005, 144, 463–471. [Google Scholar] [CrossRef]
- Song, M.L.; Li, X.Z.; Saikkonen, K.; Li, C.J.; Nan, Z.B. An asexual Epichloë endophyte enhances waterlogging tolerance of Hordeum brevisubulatum. Fungal Ecol. 2015, 13, 44–52. [Google Scholar] [CrossRef]
- Lewis, G.C. Effects of biotic and abiotic stress on the growth of three genotypes of Lolium perenne with and without infection by the fungal endophyte Neotyphodium lolii. Ann. Appl. Biol. 2004, 144, 53–63. [Google Scholar] [CrossRef]
- Berman-Frank, I.; Dubinsky, Z. Balanced growth in aquatic plants: Myth or reality? Bioscience 1999, 49, 29–37. [Google Scholar] [CrossRef]
- Finch Savage, W.E.; Cadman, C.; Toorop, P.E.; Lynn, J.R.; Hilhorst, H. Seed dormancy release in Arabidopsis Cvi by dry after-ripening, low temperature, nitrate and light shows common quantitative patterns of gene expression directed by environmentally specific sensing. Plant J. 2007, 51, 60–78. [Google Scholar] [CrossRef] [PubMed]
- Atia, A.; Debez, A.; Rabhi, M.; Smaoui, A. Interactive effects of salinity, nitrate, light and seed weight on the germination of the halophyte Crithmum maritimum. Acta Biol. Hung. 2009, 60, 433–439. [Google Scholar] [CrossRef]
- Defez, R.; Andreozzi, A.; Romano, S.; Pocsfalvi, G.; Fiume, I.; Esposito, R.; Angelini, C.; Bianco, C. Bacterial IAA-delivery into Medicago root nodules triggers a balanced stimulation of C and N metabolism leading to a biomass increase. Microorganisms 2019, 7, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X.M.; Yang, L.X.; Yang, X.G.; Guan, Q.X.; Ma, Z.S.; Pan, J.; Jiang, X.Y.; Hou, H.W. Effects of N and P fertilization on the biomass and ecological stoichiometric characteristics of Agropyron michnoi in sandy grasslands. Chem. Ecol. 2020, 36, 938–952. [Google Scholar] [CrossRef]
- Hautier, Y.; Niklaus, P.A.; Hector, A. Competition for light causes plant biodiversity loss after eutrophication. Science 2009, 324, 636–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.B.; Wang, H.Y.; Lv, X.T.; Wang, Z.W. Effects of nitrogen and phosphorus addition on C:N:P stoichiometry in roots and leaves of four dominant plant species in a meadow steppe of Hulunbuir. Chin. J. Ecol. 2016, 36, 80–88. [Google Scholar]
- Schleuss, P.M.; Widdig, M.; Heintz, B.A.; Kirkman, K.; Spohn, M. Interactions of nitrogen and phosphorus cycling promote P acquisition and explain synergistic plant-growth responses. Ecology 2020, 101, e03003. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.M.; Yin, H.B.; O’Connor, P.; Wang, Y.S.; Zhu, Y.G. C: N: P stoichiometry and specific growth rate of clover colonized by arbuscular mycorrhizal fungi. Plant Soil 2010, 326, 21–29. [Google Scholar] [CrossRef]
- Ren, A.Z.; Li, X.; Han, R.; Yin, L.J.; Wei, M.Y.; Gao, Y.B. Benefits of a symbiotic association with endophytic fungi are subject to water and nutrient availability in Achnatherum sibiricum. Plant Soil 2011, 346, 363–373. [Google Scholar] [CrossRef]
- Wardle, D.A.; Walker, L.R.; Bardgett, R.D. Ecosystem properties and forest decline in contrasting long-term chronosequences. Science 2004, 305, 509–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kannadan, S.; Rudgers, J.A. Endophyte symbiosis benefits a rare grass under low water availability. Funct. Ecol. 2010, 22, 706–713. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HNHP | HNMP | HNLP | MNHP | MNMP | MNLP | LNHP | LNMP | LNLP | |
|---|---|---|---|---|---|---|---|---|---|
| Nitrogen | 15.00 | 15.00 | 15.00 | 3.00 | 3.00 | 3.00 | 0.20 | 0.20 | 0.20 |
| Phosphorus | 1.50 | 0.10 | 0.01 | 1.50 | 0.10 | 0.01 | 1.50 | 0.10 | 0.01 |
| Treatments | dF | Carbon Content | Nitrogen Content | Phosphorus Content | ||||
|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |||
| Aboveground | N | 2 | 10.245 | <0.001 | 130.149 | <0.001 | 24.503 | <0.001 |
| P | 2 | 63.507 | <0.001 | 59.87 | <0.001 | 117.828 | <0.001 | |
| E | 1 | 1.04 | 0.315 | 22.591 | <0.001 | 31.596 | <0.001 | |
| N × P | 4 | 25.314 | <0.001 | 127.55 | <0.001 | 10.9 | <0.001 | |
| N × E | 2 | 108.468 | <0.001 | 494.878 | <0.001 | 111.535 | <0.001 | |
| P × E | 2 | 57.469 | <0.001 | 86.206 | <0.001 | 139.707 | <0.001 | |
| N × P × E | 4 | 18.014 | <0.001 | 17.51 | <0.001 | 9.807 | <0.001 | |
| Underground | N | 2 | 102.024 | <0.001 | 62.323 | <0.001 | 20.652 | <0.001 |
| P | 2 | 15.147 | <0.001 | 2.528 | 0.093 | 255.71 | <0.001 | |
| E | 1 | 0.647 | 0.426 | 6.856 | 0.013 | 30.452 | <0.001 | |
| N × P | 4 | 14.937 | <0.001 | 35.062 | <0.001 | 48.885 | <0.001 | |
| N × E | 2 | 22.853 | <0.001 | 225.816 | <0.001 | 50.025 | <0.001 | |
| P × E | 2 | 2.404 | 0.105 | 1.718 | 0.193 | 385.567 | <0.001 | |
| N × P × E | 4 | 1.845 | 0.142 | 18.922 | <0.001 | 24.548 | <0.001 | |
| Treatments | dF | C:N | C:P | N:P | ||||
|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |||
| Aboveground | N | 2 | 49.046 | <0.001 | 41.894 | <0.001 | 25.333 | <0.001 |
| P | 2 | 10.785 | <0.001 | 316.173 | <0.001 | 216.773 | <0.001 | |
| E | 1 | 18.402 | <0.001 | 0.125 | 0.726 | 11.111 | 0.002 | |
| N × P | 4 | 14.618 | <0.001 | 27.108 | <0.001 | 60.294 | <0.001 | |
| N × E | 2 | 170.645 | <0.001 | 29.508 | <0.001 | 133.427 | <0.001 | |
| P × E | 2 | 8.672 | <0.001 | 236.95 | <0.001 | 219.077 | <0.001 | |
| N × P × E | 4 | 10.371 | <0.001 | 13.654 | <0.001 | 39.311 | <0.001 | |
| Underground | N | 2 | 95.05 | <0.001 | 69.46 | <0.001 | 551.407 | <0.001 |
| P | 2 | 35.921 | <0.001 | 23.858 | <0.001 | 764.766 | <0.001 | |
| E | 1 | 49.63 | <0.001 | 43.483 | <0.001 | 42.373 | <0.001 | |
| N × P | 4 | 61.98 | <0.001 | 6.562 | <0.001 | 389.111 | <0.001 | |
| N × E | 2 | 484.522 | <0.001 | 40.37 | <0.001 | 131.882 | <0.001 | |
| P × E | 2 | 4.697 | 0.016 | 69.82 | <0.001 | 168.172 | <0.001 | |
| N × P × E | 4 | 19.702 | <0.001 | 2.754 | 0.044 | 119.217 | <0.001 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lang, M.; Zhou, J.; Chen, T.; Chen, Z.; Malik, K.; Li, C. Influence of Interactions between Nitrogen, Phosphorus Supply and Epichloёbromicola on Growth of Wild Barley (Hordeum brevisubulatum). J. Fungi 2021, 7, 615. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080615
Lang M, Zhou J, Chen T, Chen Z, Malik K, Li C. Influence of Interactions between Nitrogen, Phosphorus Supply and Epichloёbromicola on Growth of Wild Barley (Hordeum brevisubulatum). Journal of Fungi. 2021; 7(8):615. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080615
Chicago/Turabian StyleLang, Mingxiao, Jingle Zhou, Taixiang Chen, Zhenjiang Chen, Kamran Malik, and Chunjie Li. 2021. "Influence of Interactions between Nitrogen, Phosphorus Supply and Epichloёbromicola on Growth of Wild Barley (Hordeum brevisubulatum)" Journal of Fungi 7, no. 8: 615. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080615