
Investigations into Chemical Components from Monascus purpureus with Photoprotective and Anti-Melanogenic Activities

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.2. Cultivation and Preparation of Red Yeast Rice
2.3. Extraction and Isolation
2.4. Spectral Data
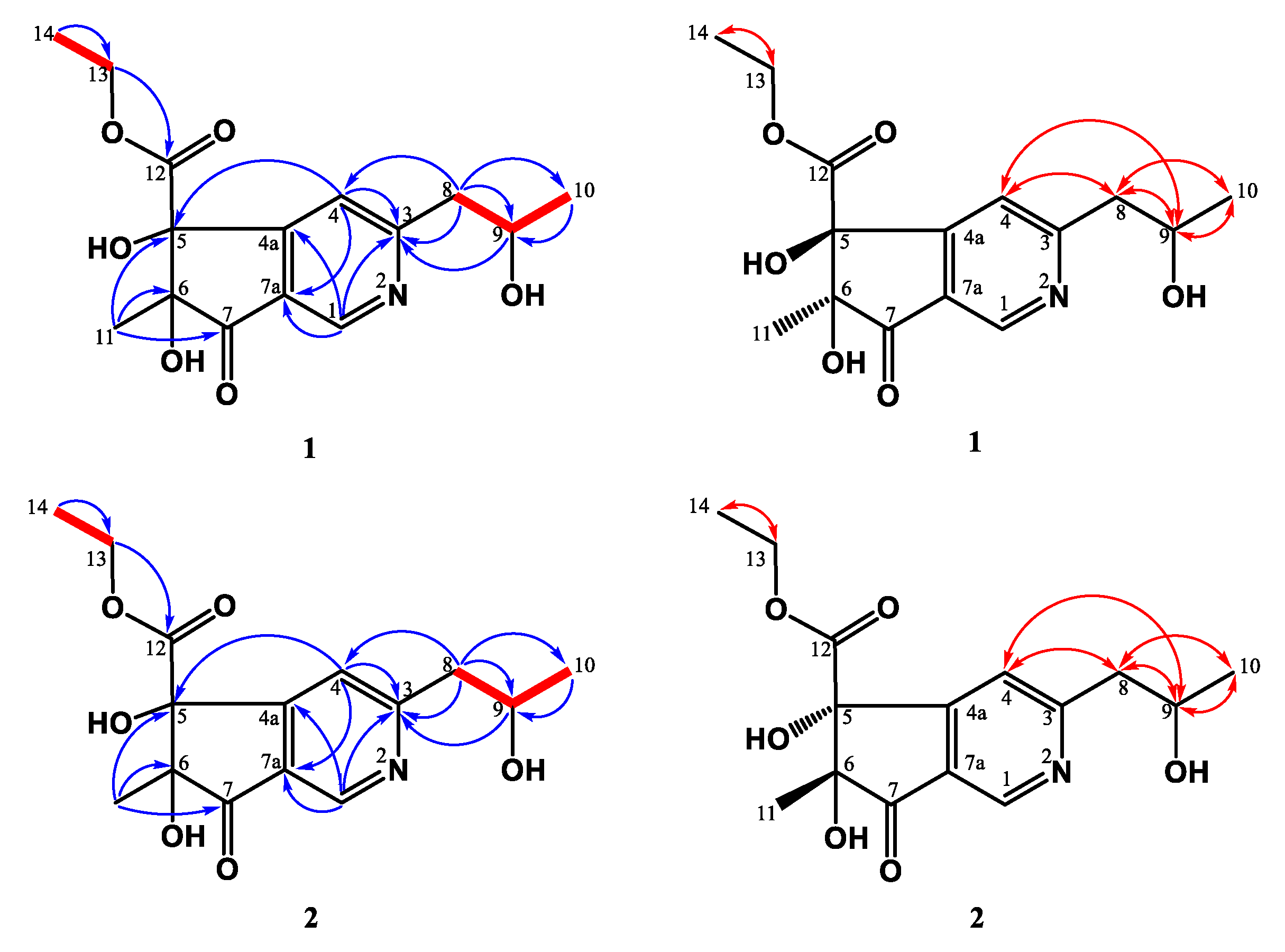
2.4.1. 5S,6S-Monaspurpyridine A (1)
2.4.2. 5R,6R-Monaspurpyridine A (2)
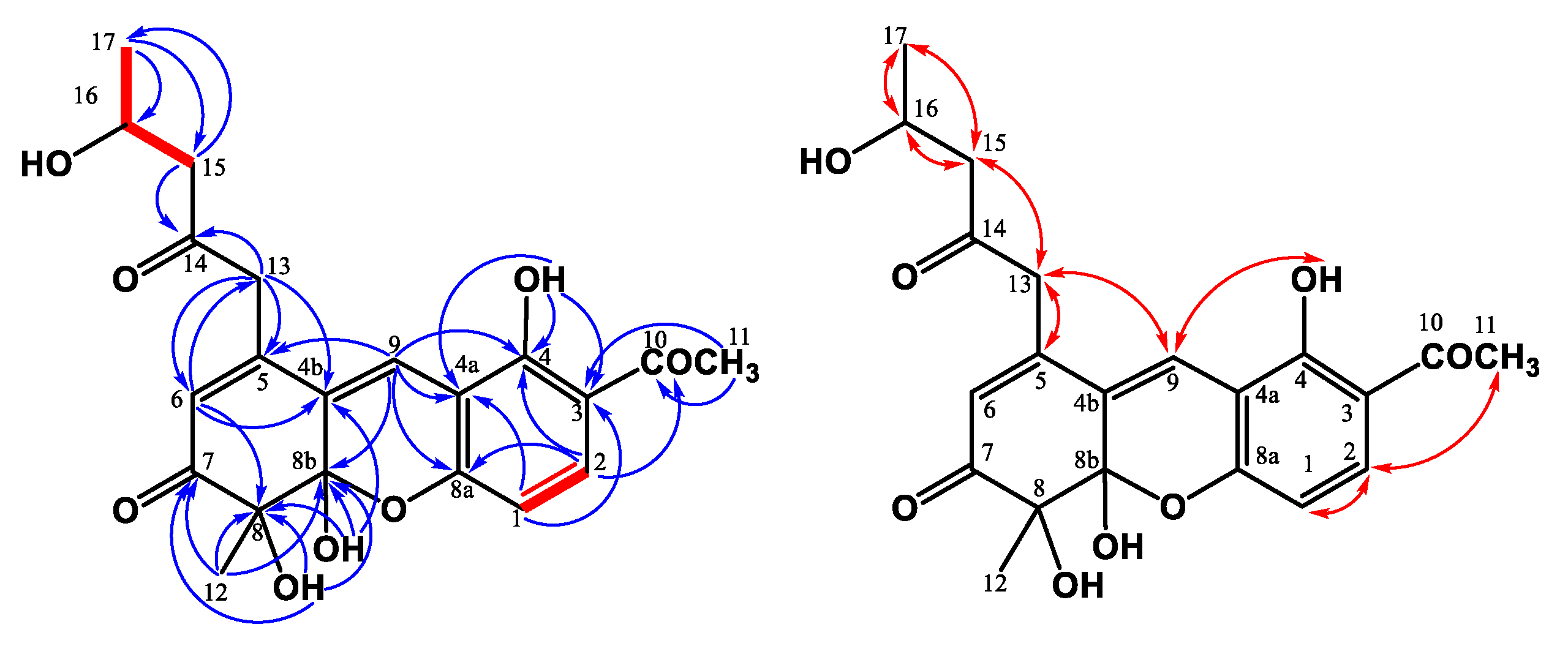
2.4.3. Monasxanthone A (3)
2.4.4. Monasxanthone B (4)
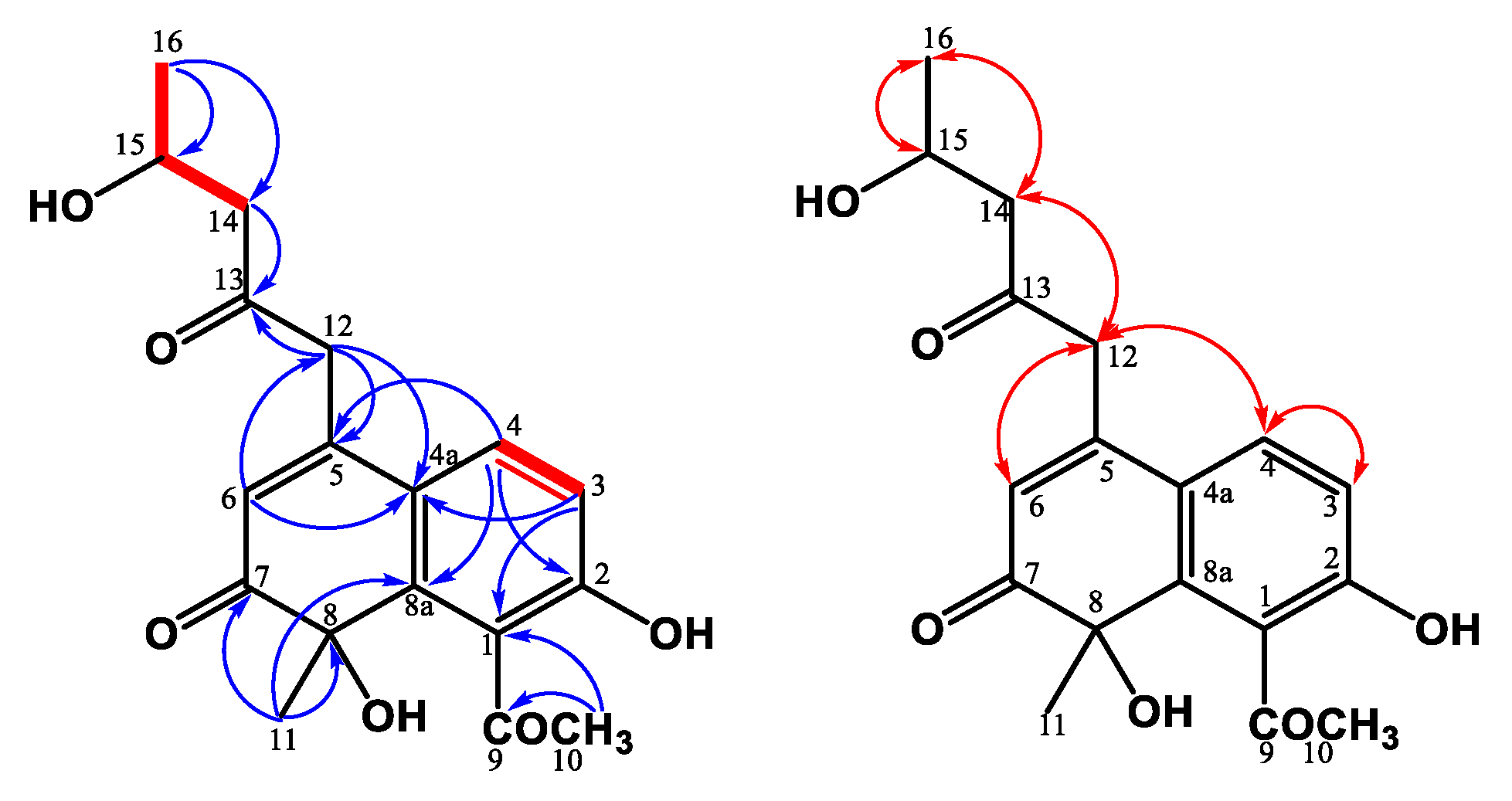
2.4.5. Monasnaphthalenone (5)
2.4.6. Monapurpurin (6)
2.5. Bioactivity Assays
2.5.1. Cell Culture
2.5.2. Cell Viability Assay
2.5.3. Determination of Anti-Melanogenic Potentials in Mouse Melanoma B16-F10 Cells
2.5.4. UV Irradiation in Human Keratinocyte HaCaT Cells
2.5.5. Measurement of Intracellular Reactive Oxygen Species (ROS) in HaCaT Cells
2.5.6. Statistical Analyses
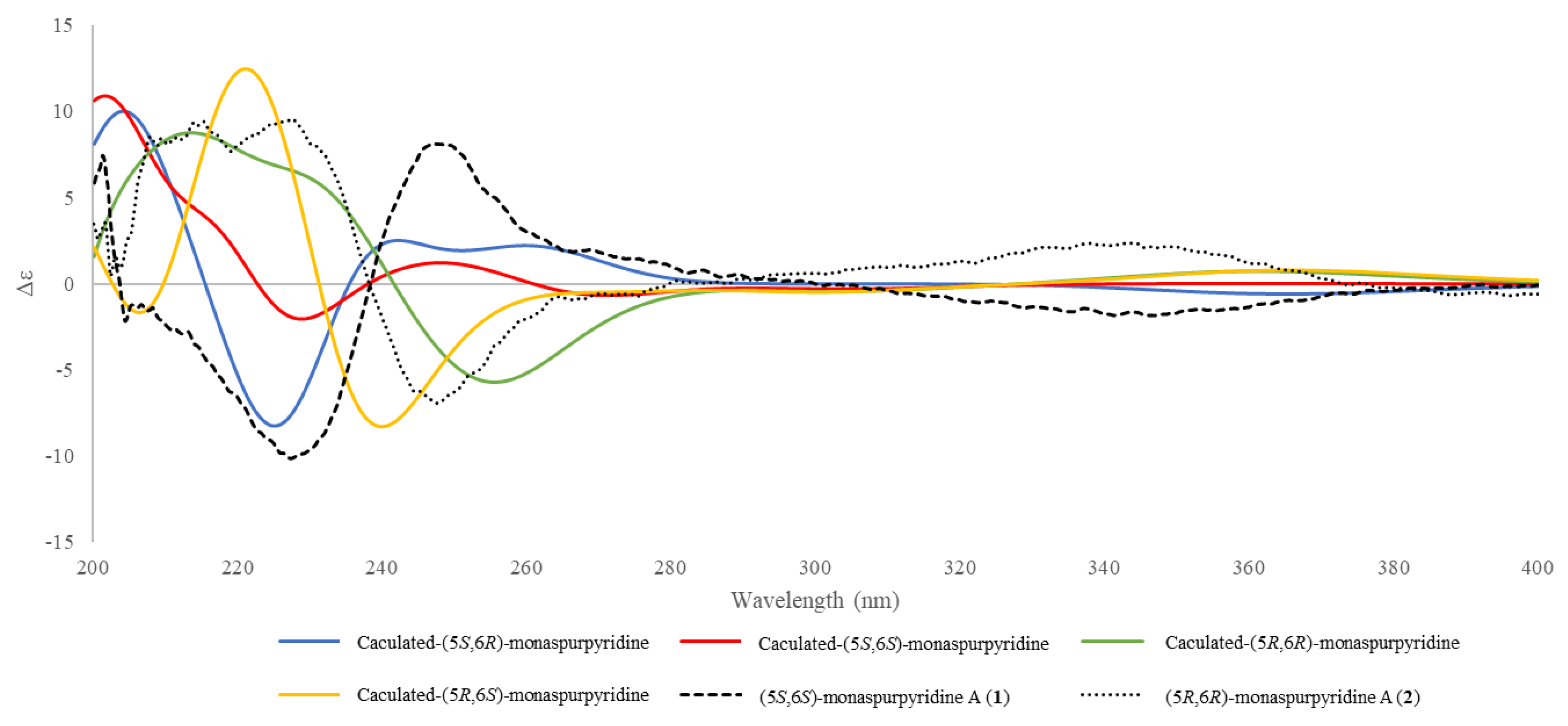
2.6. ECD Calculations
3. Results and Discussion
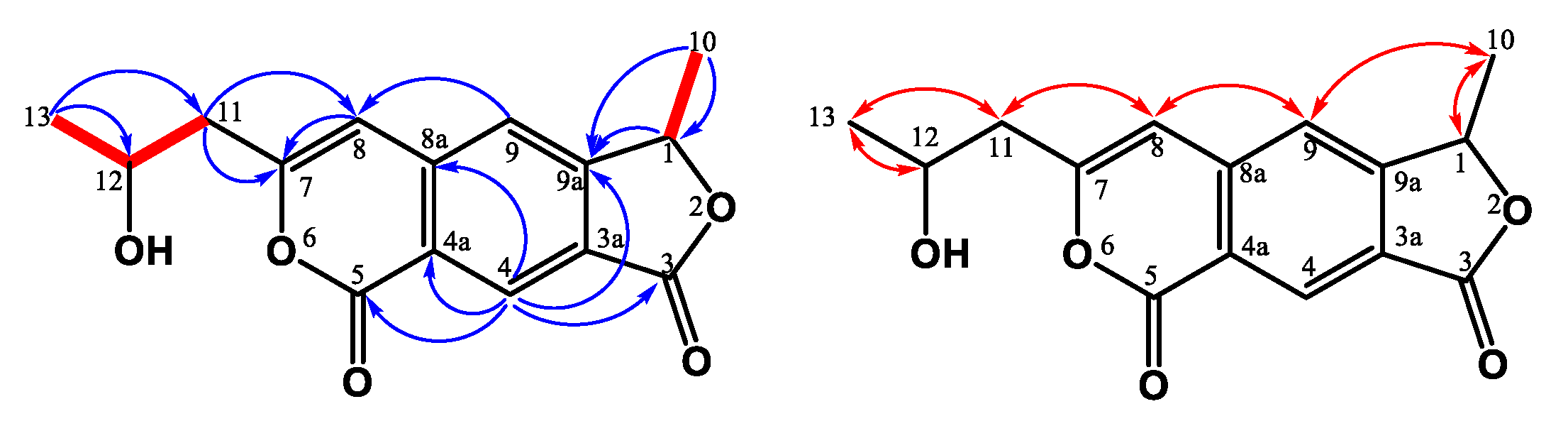
3.1. Structure Elucidation of the New Compounds
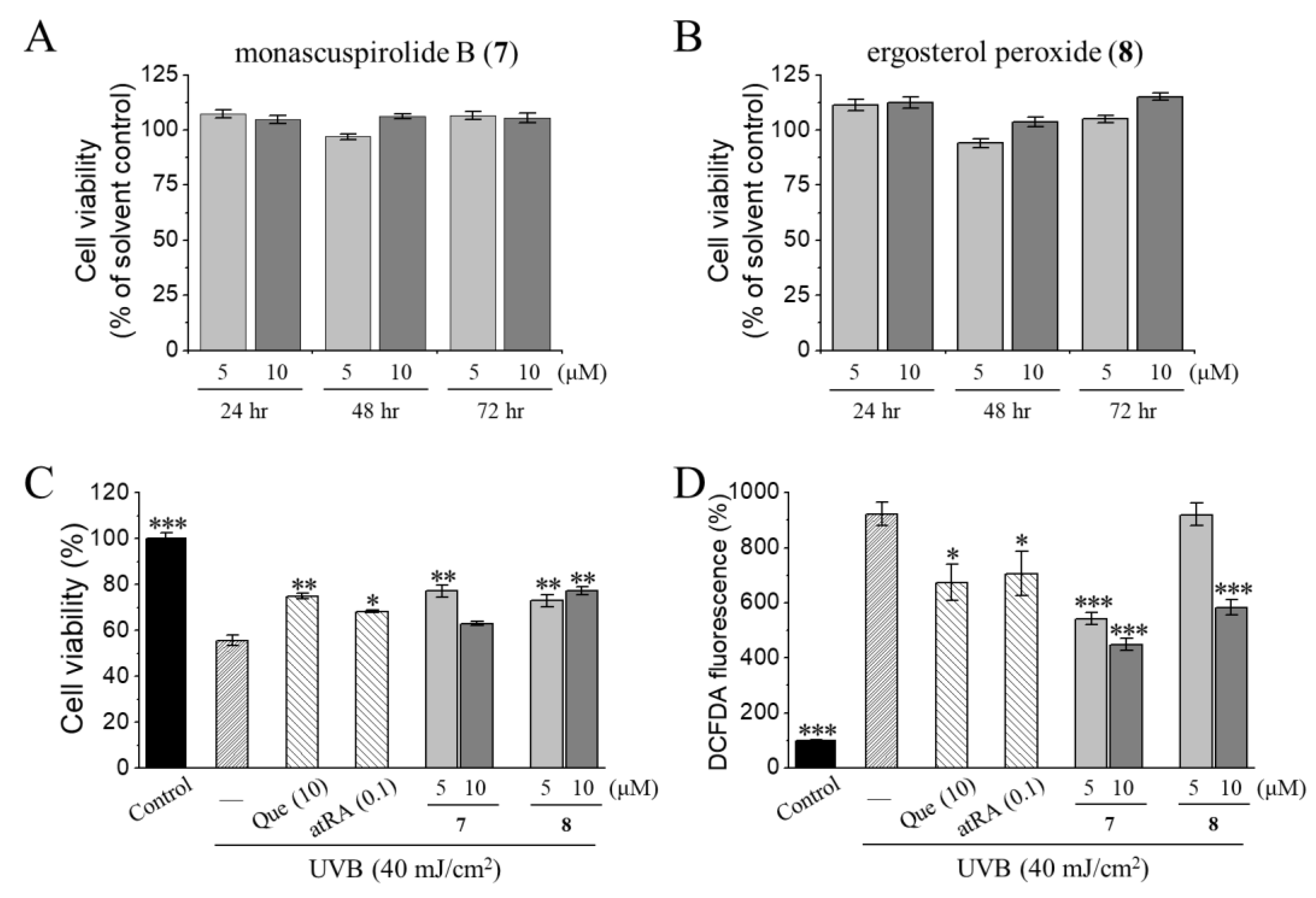
3.2. Photoprotective Activities of Monascuspirolide B (7) and Ergosterol Peroxide (8) in Human Keratinocytes HaCaT Cells
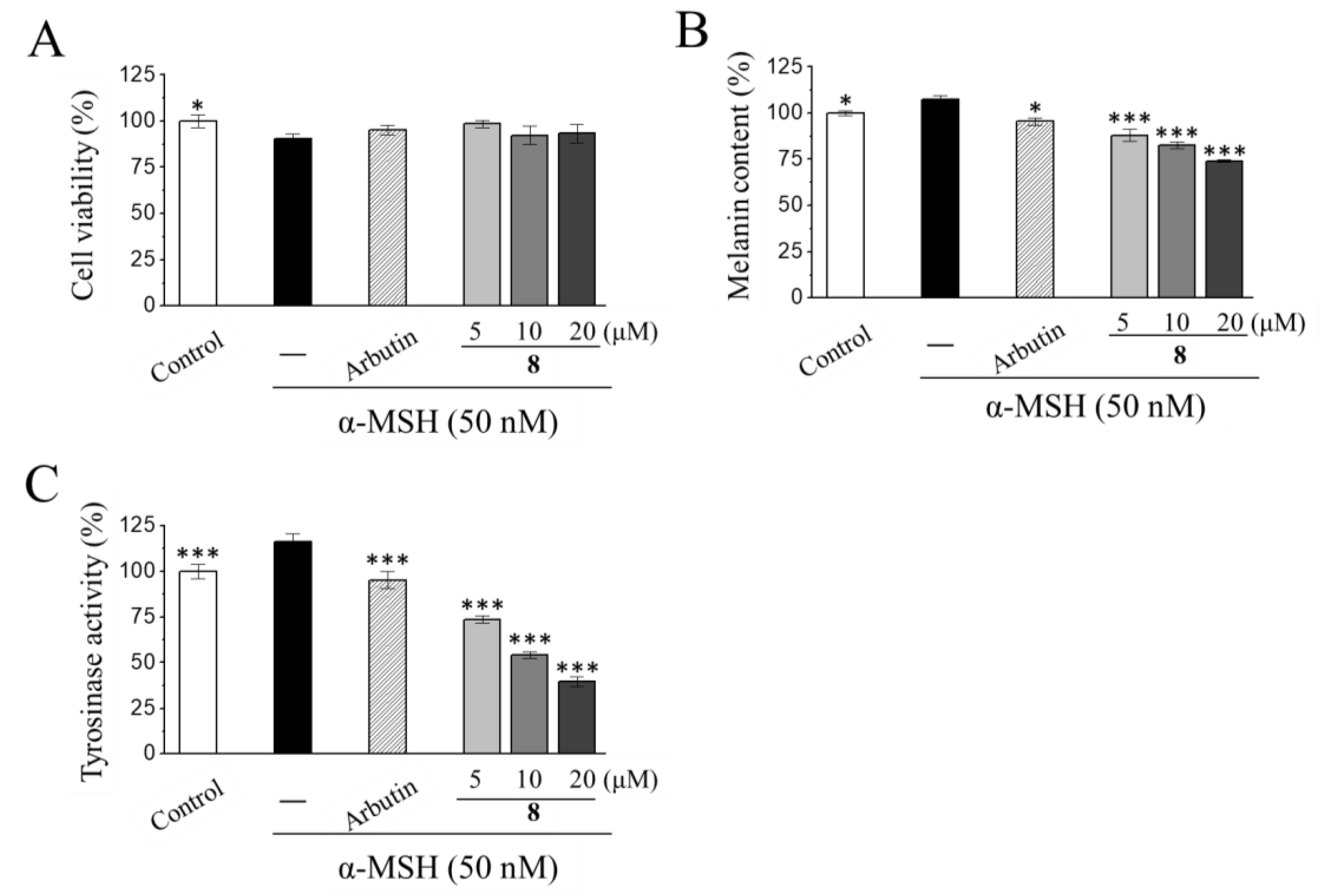
3.3. Anti-Melanogenic Activities of Ergosterol Peroxide (8) in Mouse Melanoma B16-F10 Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Li, C.L.; Zhu, Y.; Wang, Y.; Zhu, J.S.; Chang, J.; Kritchevsky, D. Monascus purpureus (Red Yeast): A natural product that lowers blood cholesterol in animal models of hyperlipidemia. Nutr. Res. 1998, 18, 71–81. [Google Scholar] [CrossRef]
- Kim, D.; Ku, S. Beneficial effects of Monascus sp. KCCM 10093 pigments and derivatives: A mini review. Molecules 2018, 23, 98. [Google Scholar] [CrossRef] [Green Version]
- Manan, M.A.; Mohamad, R.; Ariff, A. The morphology and structure of red pigment producing fungus: Monascus purpureus. J. Microbiol. Exp. 2017, 5. [Google Scholar] [CrossRef]
- Dogra, P.; Kumar, D. Characterization of Monascus purpureus isolated from red yeast rice and its evaluation for the production of cholesterol lowering lovastatin. Biol. Forum Int. J. 2017, 9, 70–76. [Google Scholar]
- Vendruscolo, F.; Bühler, R.M.M.; de Carvalho, J.C.; de Oliveira, D.; Moritz, D.E.; Schmidell, W.; Ninow, J.L. Monascus: A reality on the production and application of microbial pigments. Appl. Biochem. Biotechnol. 2016, 178, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Shang, X.Y.; Li, L.L.; Liu, M.T.; Zheng, J.Q.; Jin, Z.L. New cytotoxic azaphilones from Monascus purpureus-fermented rice (red yeast rice). Molecules 2010, 15, 1958–1966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, M.J.; Yang, P.H.; Wu, M.D.; Chen, I.S.; Hsieh, M.T.; Chen, Y.L.; Yuan, G.F. Secondary metabolites from the fungus Monascus purpureus and evaluation of their cytotoxic activity. Helv. Chim. Acta 2011, 94, 1638–1650. [Google Scholar] [CrossRef]
- Cheng, M.J.; Wu, M.D.; Chen, I.S.; Tseng, M.; Yuan, G.F. Chemical constituents from the fungus Monascus purpureus and their antifungal activity. Phytochem. Lett. 2011, 4, 372–376. [Google Scholar] [CrossRef]
- Wu, M.D.; Cheng, M.J.; Yech, Y.J.; Chen, Y.L.; Chen, K.P.; Yang, P.H.; Chen, I.S.; Yuan, G.F. Monascusazaphilones A–C, three new azaphilone analogues isolated from the fungus Monascus purpureus BCRC 38108. Nat. Prod. Res. 2013, 27, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.J.; Wu, M.D.; Chen, I.S.; Chen, C.Y.; Lo, W.L.; Yuan, G.F. Secondary metabolites from the red mould rice of Monascus purpureus BCRC 38113. Nat. Prod. Res. 2010, 24, 1719–1725. [Google Scholar] [CrossRef]
- Li, X.; Liu, C.; Duan, Z.; Guo, S. HMG-CoA reductase inhibitors from Monascus-fermented rice. J. Chem. 2013, 2013. [Google Scholar] [CrossRef] [Green Version]
- Knecht, A.; Cramer, B.; Humpf, H.U. New Monascus metabolites: Structure elucidation and toxicological properties studied with immortalized human kidney epithelial cells. Mol. Nutr. Food Res. 2006, 50, 314–321. [Google Scholar] [CrossRef]
- Campoy, S.; Rumbero, A.; Martín, J.F.; Liras, P. Characterization of an hyperpigmenting mutant of Monascus purpureus IB1: Identification of two novel pigment chemical structures. Appl. Microbiol. Biotechnol. 2006, 70, 488–496. [Google Scholar] [CrossRef]
- Mapari, S.A.; Hansen, M.E.; Meyer, A.S.; Thrane, U. Computerized screening for novel producers of Monascus-like food pigments in Penicillium species. J. Agric. Food Chem. 2008, 56, 9981–9989. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, G.; Singh, S.K. Purification and characterization of a new red pigment from Monascus purpureus in submerged fermentation. Process Biochem. 2011, 46, 188–192. [Google Scholar] [CrossRef]
- Hsu, Y.W.; Hsu, L.C.; Liang, Y.H.; Kuo, Y.H.; Pan, T.M. Monaphilones A–C, three new antiproliferative azaphilone derivatives from Monascus purpureus NTU 568. J. Agric. Food Chem. 2010, 58, 8211–8216. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.W.; Hsu, L.C.; Liang, Y.H.; Kuo, Y.H.; Pan, T.M. New bioactive orange pigments with yellow fluorescence from Monascus-fermented Dioscorea. J. Agric. Food Chem. 2011, 59, 4512–4518. [Google Scholar] [CrossRef] [PubMed]
- Hsu, L.C.; Hsu, Y.W.; Liang, Y.H.; Liaw, C.C.; Kuo, Y.H.; Pan, T.M. Induction of apoptosis in human breast adenocarcinoma cells MCF-7 by monapurpyridine A, a new azaphilone derivative from Monascus purpureus NTU 568. Molecules 2012, 17, 664–673. [Google Scholar] [CrossRef]
- Hsu, Y.W.; Hsu, L.C.; Chang, C.L.; Liang, Y.H.; Kuo, Y.H.; Pan, T.M. New anti-inflammatory and anti-proliferative constituents from fermented red mold rice Monascus purpureus NTU 568. Molecules 2010, 15, 7815–7824. [Google Scholar] [CrossRef] [PubMed]
- Salomon, H.; Karrer, P. Pflanzenfarbstoffe XXXVIII. Ein Farbstoff aus “rotem” Reis, Monascin. Helv. Chim. Acta 1932, 15, 18–22. [Google Scholar] [CrossRef]
- Fielding, B.C.; Holker, J.S.E.; Jones, D.F.; Powell, A.D.G.; Richmond, K.W.; Robertson, A.; Whalley, W.B. 898. The chemistry of fungi. Part XXXIX. The structure of monascin. J. Chem. Soc. 1961, 4579–4589. [Google Scholar] [CrossRef]
- Musso, L.; Dallavalle, S.; Merlini, L.; Bava, A.; Nasini, G.; Penco, S.; Giannini, G.; Giommarelli, C.; De Cesare, A.; Zuco, V.; et al. Natural and semisynthetic azaphilones as a new scaffold for Hsp90 inhibitors. Bioorg. Med. Chem. 2010, 18, 6031–6043. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wei, W.; Luan, N.; Li, J.; Lao, W.; Zhang, W.; Shang, X. Structure elucidation and NMR assignments of two unusual isomeric aromatic monacolin analogs from Monascus purpureus. Magn. Reson. Chem. 2015, 53, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.T.; Luan, N.; Li, J.J.; Huang, X.; Wang, Y.F.; Wang, A.L.; Shang, X.Y. Structure elucidation and NMR assignments of an unusual aromatic monacolin analog from Monascus purpureus-fermented rice. Magn. Reson. Chem. 2012, 50, 709–712. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, T.X.; Wang, A.L.; Li, J.J.; Wang, X.; Luan, N.; Ji, L.L.; Shang, X.Y. Four new monacolin analogs from Monascus purpureus-fermented rice. J. Asian Nat. Prod. Res. 2018, 20, 209–216. [Google Scholar] [CrossRef]
- Dhale, M.A.; Divakar, S.; Umesh-Kumar, S.; Vijayalakshmi, G. Characterization of dehydromonacolin-MV2 from Monascus purpureus mutant. J. Appl. Microbiol. 2007, 103, 2168–2173. [Google Scholar] [CrossRef]
- Wild, D.; Tóth, G.; Humpf, H.U. New Monascus metabolite isolated from red yeast rice (angkak, red koji). J. Agric. Food Chem. 2002, 50, 3999–4002. [Google Scholar] [CrossRef]
- Zhu, L.; Yau, L.F.; Lu, J.G.; Zhu, G.Y.; Wang, J.R.; Han, Q.B.; Hsiao, W.L.; Jiang, Z.H. Cytotoxic dehydromonacolins from red yeast rice. J. Agric. Food Chem. 2012, 60, 934–939. [Google Scholar] [CrossRef]
- Liu, M.T.; Li, J.J.; Shang, X.Y.; Li, S.; Li, L.L.; Luan, N.; Jin, Z.L. Structure elucidation and complete NMR spectral assignment of an unusual aromatic monacolin analog from Monascus purpureus-fermented rice. Magn. Reson. Chem. 2011, 49, 129–131. [Google Scholar] [CrossRef]
- Ma, J.; Li, Y.; Ye, Q.; Li, J.; Hua, Y.; Ju, D.; Zhang, D.; Cooper, R.; Chang, M. Constituents of red yeast rice, a traditional Chinese food and medicine. J. Agric. Food Chem. 2000, 48, 5220–5225. [Google Scholar] [CrossRef]
- Zhang, Y.T.; Wang, Y.; Zhang, X.T.; Wu, D.L.; Zhang, X.Q.; Ye, W.C. A new decalin derivative from red yeast rice. J. Asian Nat. Prod. Res. 2009, 11, 792–795. [Google Scholar] [CrossRef]
- Cheng, M.J.; Chen, J.J.; Wu, M.D.; Yang, P.S.; Yuan, G.F. Isolation and structure determination of one new metabolite isolated from the red fermented rice of Monascus purpureus. Nat. Prod. Res. 2010, 24, 979–988. [Google Scholar] [CrossRef]
- Jůzlová, P.; Řezanka, T.; Martínková, L.; Křen, V. Long-chain fatty acids from Monascus purpureus. Phytochemistry 1996, 43, 151–153. [Google Scholar] [CrossRef]
- Cheng, M.J.; Wu, M.D.; Cheng, Y.C.; Chen, J.J.; Chan, H.Y.; Chen, Y.L.; Chen, I.S.; Wu, P.H.; Yuan, G.F. One new compound from the extract of the fungus Monascus purpureus BCRC 31499. Chem. Nat. Compd. 2016, 52, 634–636. [Google Scholar] [CrossRef]
- Wei, W.; Lin, S.; Chen, M.; Liu, T.; Wang, A.; Li, J.; Guo, Q.; Shang, X. Monascustin, an unusual γ-lactam from red yeast rice. J. Nat. Prod. 2017, 80, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.Y.; Li, J.J.; Liu, M.T.; Li, S.; Liu, Y.; Wang, Y.F.; Huang, X.; Jin, Z.L. Cytotoxic steroids from Monascus purpureus-fermented rice. Steroids 2011, 76, 1185–1189. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.-M.; Li, Z.; Li, Y.; Duan, Z.-W.; Guo, S.-R. A new ergostane from Monascus purpureus. Chem. Nat. Compd. 2013, 49, 579–580. [Google Scholar] [CrossRef]
- Hsu, L.C.; Hsu, Y.W.; Liang, Y.H.; Lin, Z.H.; Kuo, Y.H.; Pan, T.M. Protective effect of deferricoprogen isolated from Monascus purpureus NTU 568 on citrinin-induced apoptosis in HEK-293 Cells. J. Agric. Food Chem. 2012, 60, 7880–7885. [Google Scholar] [CrossRef] [PubMed]
- Jůzlová, P.; Martínková, L.; Křen, V. Secondary metabolites of the fungus Monascus: A review. J. Ind. Microbiol. 1996, 65, 2569–2575. [Google Scholar] [CrossRef]
- Akihisa, T.; Tokuda, H.; Yasukawa, K.; Ukiya, M.; Kiyota, A.; Sakamoto, N.; Suzuki, T.; Tanabe, N.; Nishino, H. Azaphilones, furanoisophthalides, and amino acids from the extracts of Monascus pilosus-fermented rice (red-mold rice) and their chemopreventive effects. J. Agric. Food Chem. 2005, 53, 562–565. [Google Scholar] [CrossRef] [PubMed]
- Hsu, L.C.; Liang, Y.H.; Hsu, Y.W.; Kuo, Y.H.; Pan, T.M. Anti-inflammatory properties of yellow and orange pigments from Monascus purpureus NTU 568. J. Agric. Food Chem. 2013, 61, 2796–2802. [Google Scholar] [CrossRef]
- Zheng, G.; Zhan, Y.; Tang, Q.; Chen, T.; Zheng, F.; Wang, H.; Wang, J.; Wu, D.; Li, X.; Zhou, Y.; et al. Monascin inhibits IL-1β induced catabolism in mouse chondrocytes and ameliorates murine osteoarthritis. Food Funct. 2018, 9, 1454–1464. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Hsu, W.H.; Huang, T.; Chang, Y.Y.; Hsu, Y.W.; Pan, T.M. Effects of monascin on anti-inflammation mediated by Nrf2 activation in advanced glycation end product-treated THP-1 monocytes and methylglyoxal-treated Wistar rats. J. Agric. Food Chem. 2013, 61, 1288–1298. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.C.; Pan, T.M. Beneficial effects of Monascus purpureus NTU 568-fermented products: A review. Appl. Microbiol. Biotechnol. 2011, 90, 1207–1217. [Google Scholar] [CrossRef] [PubMed]
- Martínková, L.; J˚zlová, P.; Veselý, D. Biological activity of polyketide pigments produced by the fungus Monascus. J. Appl. Microbiol. 1995, 79, 609–616. [Google Scholar] [CrossRef]
- Kim, C.; Jung, H.; Kim, Y.O.; Shin, C.S. Antimicrobial activities of amino acid derivatives of Monascus pigments. FEMS Microbiol. Lett. 2006, 264, 117–124. [Google Scholar] [CrossRef]
- Feng, Y.; Shao, Y.; Chen, F. Monascus pigments. Appl. Microbiol. Biotechnol. 2012, 96, 1421–1440. [Google Scholar] [CrossRef]
- Su, N.W.; Lin, Y.L.; Lee, M.H.; Ho, C.Y. Ankaflavin from Monascus-fermented red rice exhibits selective cytotoxic effect and induces cell death on Hep G2 cells. J. Agric. Food Chem. 2005, 53, 1949–1954. [Google Scholar] [CrossRef]
- Chiu, H.W.; Chen, M.H.; Fang, W.H.; Hung, C.M.; Chen, Y.L.; Wu, M.D.; Yuan, G.F.; Wu, M.J.; Wang, Y.J. Preventive effects of Monascus on androgen-related diseases: Androgenetic alopecia, benign prostatic hyperplasia, and prostate cancer. J. Agric. Food Chem. 2013, 61, 4379–4386. [Google Scholar] [CrossRef]
- Chang, W.T.; Chuang, C.H.; Lee, W.J.; Huang, C.S. Extract of Monascus purpureus CWT715 fermented from Sorghum liquor biowaste inhibits migration and invasion of SK-Hep-1 human hepatocarcinoma cells. Molecules 2016, 21, 1691. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.I.; Lee, C.L.; Hwang, J.F.; Lee, Y.H.; Wang, J.J. Monascus-fermented red mold rice exhibits cytotoxic effect and induces apoptosis on human breast cancer cells. Appl. Microbiol. Biotechnol. 2013, 97, 1269–1278. [Google Scholar] [CrossRef]
- Yasukawa, K.; Takahashi, M.; Natori, S.; Kawai, K.i.; Yamazaki, M.; Takeuchi, M.; Takido, M. Azaphilones inhibit tumor promotion by 12-O-tetradecanoylphorbol-13-acetate in two-stage carcinogenesis in mice. Oncology 1994, 51, 108–112. [Google Scholar] [CrossRef]
- Akihisa, T.; Tokuda, H.; Ukiya, M.; Kiyota, A.; Yasukawa, K.; Sakamoto, N.; Kimura, Y.; Suzuki, T.; Takayasu, J.; Nishino, H. Anti-tumor-initiating effects of monascin, an azaphilonoid pigment from the extract of Monascus pilosus fermented rice (red-mold rice). Chem. Biodivers. 2005, 2, 1305–1309. [Google Scholar] [CrossRef]
- Wu, H.C.; Cheng, M.J.; Wu, M.D.; Chen, J.J.; Chen, Y.L.; Chang, H.S.; Chen, K.P. Secondary metabolites from the fermented rice of the fungus Monascus purpureus and their bioactivities. Nat. Prod. Res. 2019, 33, 3541–3550. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.C.; Cheng, M.J.; Wu, M.D.; Chen, J.J.; Chen, Y.L.; Chang, H.S. Three new constituents from the fungus of Monascus purpureus and their anti-inflammatory activity. Phytochem. Lett. 2019, 31, 242–248. [Google Scholar] [CrossRef]
- Sesto, A.; Navarro, M.; Burslem, F.; Jorcano, J.L. Analysis of the ultraviolet B response in primary human keratinocytes using oligonucleotide microarrays. Proc. Natl. Acad. Sci. USA 2002, 99, 2965–2970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, M.C.; Lee, T.H.; Chu, Y.T.; Syu, L.L.; Hsu, S.J.; Cheng, C.H.; Wu, J.; Lee, C.K. Melanogenesis inhibitors from the rhizoma of Ligusticum Sinense in B16-F10 melanoma cells in vitro and zebrafish in vivo. Int. J. Mol. Sci. 2018, 19, 3994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, C.M.; Huang, C.H.; Li, H.J.; Hsiao, C.Y.; Su, C.C.; Lee, P.L.; Hung, C.F. Protective effects of resveratrol against UVA-induced damage in ARPE19 cells. Int. J. Mol. Sci. 2015, 16, 5789–5802. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.H.; Jin, Z.H. Paracrine regulation of melanogenesis. Br. J. Dermatol. 2018, 178, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Promden, W.; Viriyabancha, W.; Monthakantirat, O.; Umehara, K.; Noguchi, H.; De-Eknamkul, W. Correlation between the potency of flavonoids on mushroom tyrosinase inhibitory activity and melanin synthesis in melanocytes. Molecules 2018, 23, 1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillaiyar, T.; Namasivayam, V.; Manickam, M.; Jung, S.H. Inhibitors of melanogenesis: An updated review. J. Med. Chem. 2018, 61, 7395–7418. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.N. Methodology for evaluation of melanin content and production of pigment cells in vitro. Photochem. Photobiol. 2008, 84, 645–649. [Google Scholar] [CrossRef]
- Boukamp, P.; Petrussevska, R.T.; Breitkreutz, D.; Hornung, J.; Markham, A.; Fusenig, N.E. Normal keratinization in a spontaneously immortalized aneuploid human keratinocyte cell line. J. Cell Biol. 1988, 106, 761–771. [Google Scholar] [CrossRef] [Green Version]
- Cheng, T.L.; Lai, C.H.; Chen, P.K.; Cho, C.F.; Hsu, Y.Y.; Wang, K.C.; Lin, W.L.; Chang, B.I.; Liu, S.K.; Wu, Y.T.; et al. Thrombomodulin promotes diabetic wound healing by regulating toll-like receptor 4 expression. J. Investig. Dermatol. 2015, 135, 1668–1675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, H.H.; Chang, Y.T.; Kuo, Y.H.; Lin, C.H.; Chen, Y.F. Oenothera laciniata Hill extracts exhibits antioxidant effects and attenuates melanogenesis in B16-F10 cells via downregulating CREB/MITF/tyrosinase and upregulating p-ERK and p-JNK. Plants 2021, 10, 727. [Google Scholar] [CrossRef]
- Jin, Y.J.; Lin, C.C.; Lu, T.M.; Li, J.H.; Chen, I.S.; Kuo, Y.H.; Ko, H.H. Chemical constituents derived from Artocarpus xanthocarpus as inhibitors of melanin biosynthesis. Phytochemistry 2015, 117, 424–435. [Google Scholar] [CrossRef]
- Mathes, S.H.; Ruffner, H.; Graf-Hausner, U. The use of skin models in drug development. Adv. Drug Deliv. Rev. 2014, 69–70, 81–102. [Google Scholar] [CrossRef]
- Colombo, I.; Sangiovanni, E.; Maggio, R.; Mattozzi, C.; Zava, S.; Corbett, Y.; Fumagalli, M.; Carlino, C.; Corsetto, P.A.; Scaccabarozzi, D.; et al. HaCaT cells as a reliable in vitro differentiation model to dissect the inflammatory/repair response of human keratinocytes. Mediat. Inflamm. 2017, 2017, 7435621. [Google Scholar] [CrossRef] [PubMed]
- O’Boyle, N.; Tenderholt, A.; Langner, K. cclib: A library for package-independent computational chemistry algorithms. J. Comput. Chem. 2008, 29, 839–845. [Google Scholar] [CrossRef]
- Zhu, L.J.; Hou, Y.L.; Shen, X.Y.; Pan, X.D.; Zhang, X.; Yao, X.S. Monoterpene pyridine alkaloids and phenolics from Scrophularia ningpoensis and their cardioprotective effect. Fitoterapia 2013, 88, 44–49. [Google Scholar] [CrossRef]
- Demole, E.; Demole, C. A chemical study of Burley tobacco flavour (Nicotiana tabacum L.) V. Identification and synthesis of the novel terpenoid alkaloids 1,3,6,6-tetramethyl-5,6,7,8-tetrahydro-isoquinolin-8-one and 3,6,6-trimethyl-5,6-dihydro-7H-2-pyrindin-7-one. Helv. Chim. Acta 1975, 58, 523–531. [Google Scholar] [CrossRef]
- Lee, M.S.; Wang, S.W.; Wang, G.J.; Pang, K.L.; Lee, C.K.; Kuo, Y.H.; Cha, H.J.; Lin, R.K.; Lee, T.H. Angiogenesis inhibitors and anti-inflammatory agents from Phoma sp. NTOU4195. J. Nat. Prod. 2016, 79, 2983–2990. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.R.; Wang, S.W.; Su, C.J.; Hu, H.C.; Yang, Y.L.; Hsieh, C.T.; Peng, C.C.; Chang, F.R.; Cheng, Y.B. Anti-lymphangiogenesis components from Zoanthid Palythoa tuberculosa. Mar. Drugs 2018, 16, 47. [Google Scholar] [CrossRef] [Green Version]
- Fangkrathok, N.; Sripanidkulchai, B.; Umehara, K.; Noguchi, H. Bioactive ergostanoids and a new polyhydroxyoctane from Lentinus polychrous mycelia and their inhibitory effects on E2-enhanced cell proliferation of T47D cells. Nat. Prod. Res. 2013, 27, 1611–1619. [Google Scholar] [CrossRef]
- Xu, D.; Hu, M.J.; Wang, Y.Q.; Cui, Y.L. Antioxidant activities of quercetin and its complexes for medicinal application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Park, S.S.; Min, T.J.; Yu, K.H. Antioxidant activity of ergosterol peroxide (5,8-epidioxy-5α,8α-ergosta-6,22E-dien-3β-ol) in Armillariella mellea. Bull. Korean Chem. Soc. 1999, 20, 819–823. [Google Scholar] [CrossRef]
- Bayer, M.; Proksch, P.; Felsner, I.; Brenden, H.; Kohne, Z.; Walli, R.; Duong, T.N.; Götz, C.; Krutmann, J.; Grether-Beck, S. Photoprotection against UVAR: Effective triterpenoids require a lipid raft stabilizing chemical structure. Exp. Dermatol. 2011, 20, 955–958. [Google Scholar] [CrossRef] [PubMed]
- Mukaiyama, T.; Tsujimura, N.; Otaka, S.; Kosaka, Y.; Hata, K.; Hori, K.; Sakamoto, K. Anti-melanogenic activity of ergosterol peroxide from Ganoderma lucidum on a mouse melanoma cell line. In Animal Cell Technology: Basic & Applied Aspects; Shirahata, S., Ikura, K., Nagao, M., Ichikawa, A., Teruya, K., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 273–277. [Google Scholar]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Mello, S.A.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling pathways in melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alijevic, O.; McHugh, D.; Rufener, L.; Mazurov, A.; Hoeng, J.; Peitsch, M. An electrophysiological characterization of naturally occurring tobacco alkaloids and their action on human α4β2 and α7 nicotinic acetylcholine receptors. Phytochemistry 2020, 170, 112187. [Google Scholar] [CrossRef]
- Velíšek, J.; Davídek, J.; Cejpek, K. Biosynthesis of food constituents: Natural pigments. Part 2—A review. Czech J. Food Sci. 2008, 26, 73–98. [Google Scholar] [CrossRef] [Green Version]
- Negi, J.S.; Bisht, V.K.; Singh, P.; Rawat, M.S.M.; Joshi, G.P. Naturally occurring xanthones: Chemistry and biology. J. Appl. Chem. 2013, 2013. [Google Scholar] [CrossRef] [Green Version]
- Agboyibor, C.; Kong, W.B.; Chen, D.; Zhang, A.M.; Niu, S.Q. Monascus pigments production, composition, bioactivity and its application: A review. Biocatal. Agric. Biotechnol. 2018, 16, 433–447. [Google Scholar] [CrossRef]
- El-Seedi, H.R.; El-Barbary, M.A.; El-Ghorab, D.M.; Bohlin, L.; Borg-Karlson, A.K.; Göransson, U.; Verpoorte, R. Recent insights into the biosynthesis and biological activities of natural xanthones. Curr. Med. Chem. 2010, 17, 854–901. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A. Naturally occurring naphthalenes: Chemistry, biosynthesis, structural elucidation, and biological activities. Phytochem. Rev. 2016, 15, 279–295. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | Monasxanthone A (3) | Monasxanthone B (4) | ||
|---|---|---|---|---|
| δH (m, J in Hz) | δC | δH (m, J in Hz) | δC | |
| 1 | 6.65 (dd, 9.0, 0.6) | 108.7 | 6.66 (dd, 9.0, 0.6) | 108.8 |
| 2 | 7.73 (d, 9.0) | 133.9 | 7.73 (d, 9.0) | 134.0 |
| 3 | 114.6 | 114.6 | ||
| 4 | 161.3 | 161.2 | ||
| 4a | 109.4 | 109.4 | ||
| 4b | 125.5 | 125.3 | ||
| 5 | 149.5 | 149.2 | ||
| 6 | 6.03 (s) | 123.1 | 6.03 (d, 0.5) | 123.3 |
| 7 | 198.2 | 198.2 | ||
| 8 | 79.1 | 79.1 | ||
| 8a | 157.2 | 157.2 | ||
| 8b | 97.0 | 97.0 | ||
| 9 | 7.42 (s) | 123.5 | 7.52 (s) | 123.8 |
| 10 | 203.2 | 203.2 | ||
| 11 | 2.59 (s) | 26.3 | 2.59 (s) | 26.3 |
| 12 | 1.52 (s) | 23.0 | 1.53 (s) | 23.0 |
| 13 | 3.62 (d, 16.5) | 46.9 | 3.67 (d, 17.1) | 48.0 |
| 3.79 (d, 16.5) | 3.90 (d, 17.1) | |||
| 14 | 205.1 | 205.9 | ||
| 15 | 2.57 (t, 7.5) | 44.6 | 2.66 (dd, 16.4, 3.3) | 50.5 |
| 2.75 (dd, 16.4, 9.0) | ||||
| 16 | 1.67 (sextet, 7.2) | 17.2 | 4.29 (m) | 64.6 |
| 17 | 0.94 (t, 7.2) | 13.6 | 1.25 (d, 6.6) | 22.97 |
| OCH3-16 | ||||
| OH-4 | 13.3 (s) | 13.4 (s) | ||
| OH-8 | 4.26 (s) | 4.25 (s) | ||
| OH-8b | 4.27 (s) | 4.28 (s) | ||
| OH-16 | 2.60 (br s) | |||
| Treatments | Cell Viability a (% of Basal Group) | Melanin Content a (% of Basal Group) | Tyrosinase Activity a (% of Basal Group) |
|---|---|---|---|
| α-MSH induction | 99.3 ± 3.6 | 106.8 ± 3.7 | 135.5 ± 1.8 |
| Monascuspirolide B (7) | 106.9 ± 2.9 | 96.2 ± 6.0 | 137.0 ± 1.5 |
| Ergosterol peroxide (8) | 98.6 ± 0.9 | 55.0 ± 0.7 * | 73.7 ± 1.9 * |
| Kojic acid b | 97.4 ± 0.7 | 88.0 ± 2.0 * | 109.7 ± 1.9 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.-C.; Chen, Y.-F.; Cheng, M.-J.; Wu, M.-D.; Chen, Y.-L.; Chang, H.-S. Investigations into Chemical Components from Monascus purpureus with Photoprotective and Anti-Melanogenic Activities. J. Fungi 2021, 7, 619. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080619
Wu H-C, Chen Y-F, Cheng M-J, Wu M-D, Chen Y-L, Chang H-S. Investigations into Chemical Components from Monascus purpureus with Photoprotective and Anti-Melanogenic Activities. Journal of Fungi. 2021; 7(8):619. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080619
Chicago/Turabian StyleWu, Ho-Cheng, Yih-Fung Chen, Ming-Jen Cheng, Ming-Der Wu, Yen-Lin Chen, and Hsun-Shuo Chang. 2021. "Investigations into Chemical Components from Monascus purpureus with Photoprotective and Anti-Melanogenic Activities" Journal of Fungi 7, no. 8: 619. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080619