
Assessment of Biofilm Formation by Candida albicans Strains Isolated from Hemocultures and Their Role in Pathogenesis in the Zebrafish Model
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Their In Vitro Virulence Study
2.2. Microinjection in Zebrafish Larvae
2.3. Survival Study
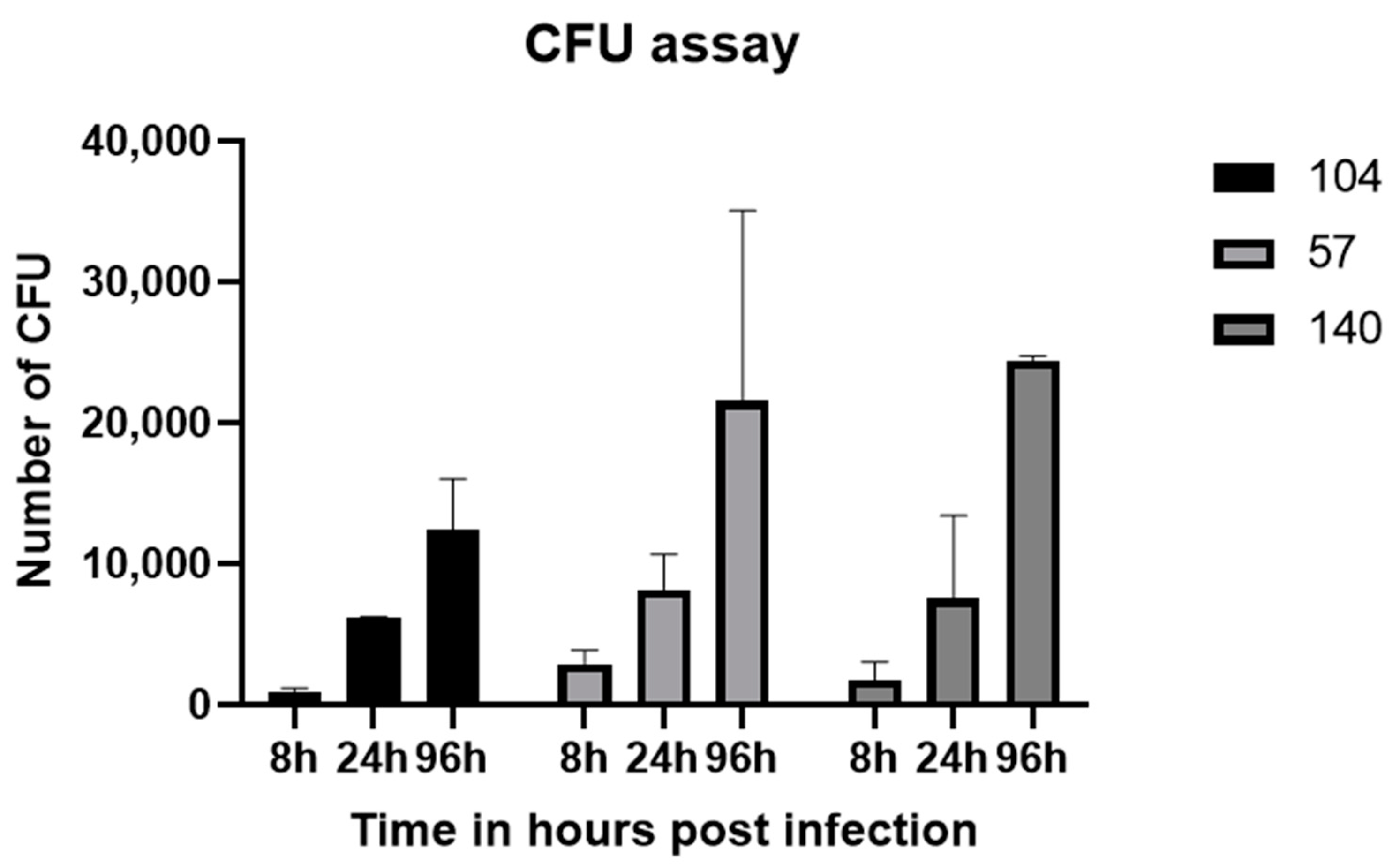
2.4. Fungal Tissue Burden/CFU Assay
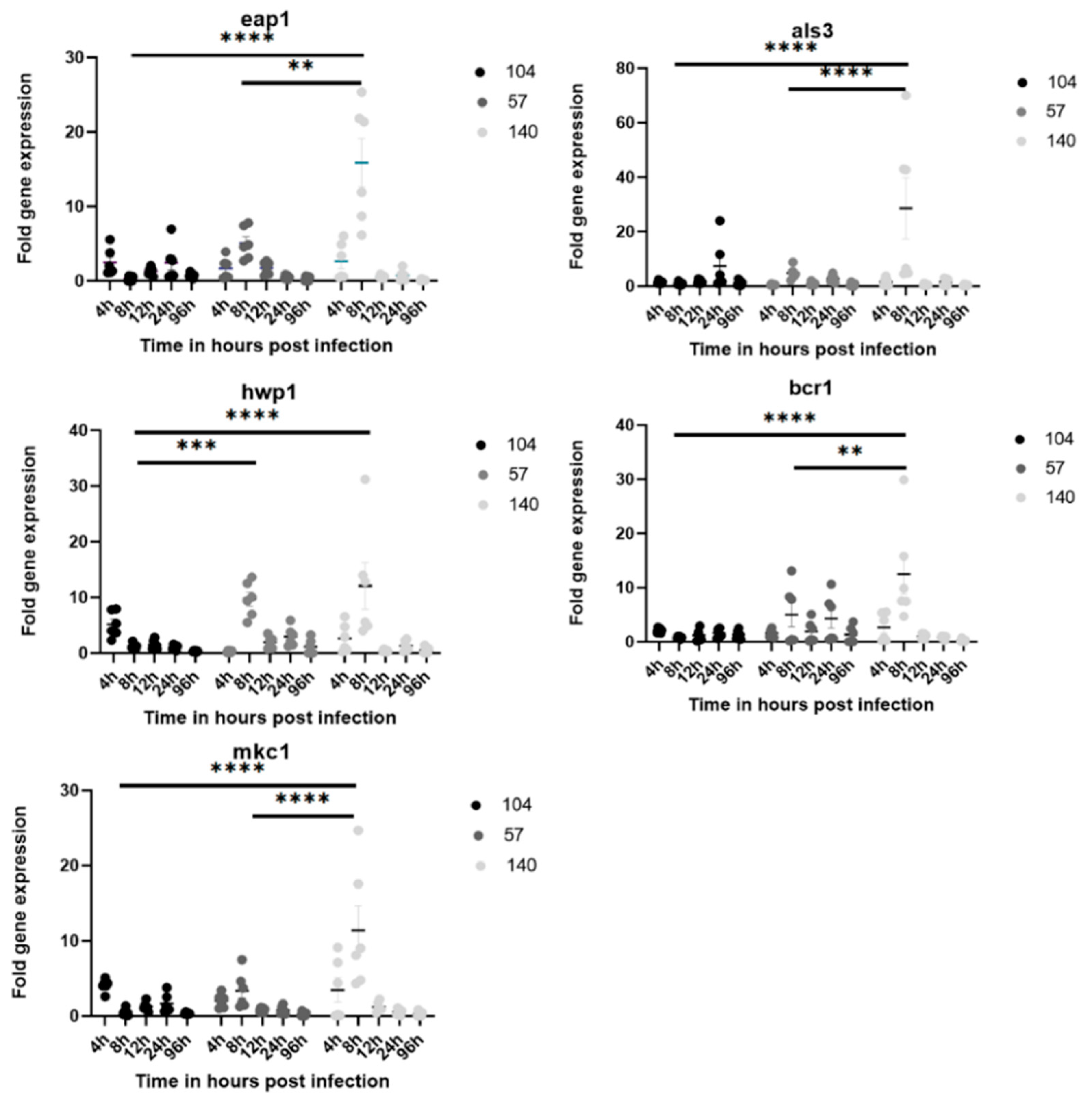
2.5. RNA Extraction and Gene Expression Analysis
2.6. Statistical Analysis
3. Results
3.1. In Vitro Virulence Assessment
3.2. Survival Assay
3.3. Tissue Burden Assessment
3.4. Gene Expression Related to Biofilm Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hall, R.A. Dressed to impress: Impact of environmental adaptation on the Candida albicans cell wall. Mol. Microbiol. 2015, 97, 7–17. [Google Scholar] [CrossRef]
- Almirante, B.; Rodríguez, D.; Park, B.J.; Cuenca-Estrella, M.; Planes, A.M.; Almela, M.; Mensa, J.; Sanchez, F.; Ayats, J.; Gimenez, M.; et al. Epidemiology and predictors of mortality in cases of Candida bloodstream infection: Results from population-based surveillance, Barcelona, Spain, from 2002 to 2003. J. Clin. Microbiol. 2005, 43, 1829–1835. [Google Scholar] [CrossRef]
- Klevay, M.J.; Ernst, E.J.; Hollanbaugh, J.L.; Miller, J.G.; Pfaller, M.A.; Diekema, D.J. Therapy and outcome of Candida glabrata versus Candida albicans bloodstream infection. Diagn. Microbiol. Infect. Dis. 2008, 60, 273–277. [Google Scholar] [CrossRef]
- Leroy, O.; Gangneux, J.P.; Montravers, P.; Mira, J.P.; Gouin, F.; Sollet, J.P.; Carlet, J.; Reynes, J.; Rosenheim, M.; Regnier, B.; et al. Epidemiology, management, and risk factors for death of invasive Candida infections in critical care: A multicenter, prospective, observational study in France (2005–2006). Crit. Care Med. 2009, 37, 1612–1618. [Google Scholar] [CrossRef] [PubMed]
- Talapko, J.; Juzbašić, M.; Matijević, T.; Pustijanac, E.; Bekić, S.; Kotris, I.; Škrlec, I. Candida albicans-The Virulence Factors and Clinical Manifestations of Infection. J. Fungi 2021, 7, 79. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Stoodley, P. Evolving concepts in biofilm infections. Cell Microbiol. 2009, 11, 1034–1043. [Google Scholar] [CrossRef]
- Fanning, S.; Mitchell, A.P. Fungal biofilms. PLoS Pathog. 2012, 8, e1002585. [Google Scholar] [CrossRef]
- Xu, Z.; Huang, T.; Min, D.; Soteyome, T.; Lan, H.; Hong, W.; Peng, F.; Fu, X.; Peng, G.; Liu, J.; et al. Regulatory network controls microbial biofilm development, with Candida albicans as a representative: From adhesion to dispersal. Bioengineered 2022, 13, 253–267. [Google Scholar] [CrossRef]
- Tsui, C.; Kong, E.F.; Jabra-Rizk, M.A. Pathogenesis of Candida albicans biofilm. Pathog. Dis. 2016, 74, ftw018. [Google Scholar] [CrossRef]
- Nett, J.E.; Andes, D.R. Contributions of the Biofilm Matrix to Candida Pathogenesis. J. Fungi 2020, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- Hirschfeld, J. Dynamic interactions of neutrophils and biofilms. J. Oral. Microbiol. 2014, 6, 26102. [Google Scholar] [CrossRef] [PubMed]
- Uppuluri, P.; Chaturvedi, A.K.; Srinivasan, A.; Banerjee, M.; Ramasubramaniam, A.K.; Köhler, J.R.; Kadosh, D.; Lopez-Ribot, J.L. Dispersion as an important step in the Candida albicans biofilm developmental cycle. PLoS Pathog. 2010, 6, e1000828. [Google Scholar] [CrossRef] [PubMed]
- Zarnowski, R.; Sanchez, H.; Covelli, A.S.; Dominguez, E.; Jaromin, A.; Bernhardt, J.; Mitchell, K.F.; Heiss, C.; Azadi, P.; Mitchell, A. Candida albicans biofilm–induced vesicles confer drug resistance through matrix biogenesis. PLoS Biol. 2018, 16, e2006872. [Google Scholar] [CrossRef] [PubMed]
- Sherry, L.; Rajendran, R.; Lappin, D.F.; Borghi, E.; Perdoni, F.; Falleni, M.; Tosi, D.; Smith, K.; Williams, C.; Jones, B.; et al. Biofilms formed by Candida albicans bloodstream isolates display phenotypic and transcriptional heterogeneity that are associated with resistance and pathogenicity. BMC Microbiol. 2014, 14, 182. [Google Scholar] [CrossRef]
- Vitális, E.; Nagy, F.; Tóth, Z.; Forgács, L.; Bozó, A.; Kardos, G.; Majoros, L.; Kovács, R. Candida biofilm production is associated with higher mortality in patients with candidaemia. Mycoses 2020, 63, 352–360. [Google Scholar] [CrossRef]
- Pham, L.T.T.; Pharkjaksu, S.; Chongtrakool, P.; Suwannakarn, K.; Ngamskulrungroj, P. A predominance of clade 17 Candida albicans isolated from hemocultures in a tertiary care hospital in Thailand. Front. Microbiol. 2019, 10, 1194. [Google Scholar] [CrossRef]
- Tulyaprawat, O.; Pharkjaksu, S.; Chongtrakool, P.; Ngamskulrungroj, P. An Association of an eBURST group with triazole resistance of Candida tropicalis blood isolates. Front. Microbiol. 2020, 11, 934. [Google Scholar] [CrossRef]
- Sachin, C.; Ruchi, K.; Santosh, S. In vitro evaluation of proteinase, phospholipase and haemolysin activities of Candida species isolated from clinical specimens. Int. J. Med. Biomed. Res. 2012, 1, 153–157. [Google Scholar] [CrossRef]
- Oz, Y.; Dag, I.; Kiraz, N. Efficacy of disinfectants on Candida biofilms at different concentrations and contact times. Br. Microbiol. Res. J. 2012, 2, 40–52. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Chao, C.C.; Liu, F.C.; Hsu, P.C.; Chen, H.F.; Peng, S.C.; Chuang, Y.J.; Lan, C.Y.; Hsieh, W.P.; Wong, D.S. Dynamic transcript profiling of Candida albicans infection in zebrafish: A pathogen-host interaction study. PLoS ONE 2013, 8, e72483. [Google Scholar] [CrossRef]
- Li, X.; Yan, Z.; Xu, J. Quantitative variation of biofilms among strains in natural populations of Candida albicans. Microbiology 2003, 149, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Kadkhoda, H.; Ghalavand, Z.; Nikmanesh, B.; Kodori, M.; Houri, H.; Taghizadeh Maleki, D.; Karimi Bavandpour, A.; Eslami, G. Characterization of biofilm formation and virulence factors of Staphylococcus aureus isolates from paediatric patients in Tehran, Iran. Iran J. Basic. Med. Sci. 2020, 23, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Li, X.F.; Shi, H.Q.; Liang, Y.; Li, J.; Jiang, B.; Song, G.B. Interaction of biofilm and efflux pump in clinical isolates of carbapenem resistant P. aeruginosa. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 1729–1737. [Google Scholar] [CrossRef] [PubMed]
- Pharkjaksu, S.; Boonmee, N.; Mitrpant, C.; Ngamskulrungroj, P. Immunopathogenesis of Emerging Candida auris and Candida haemulonii Strains. J. Fungi 2021, 7, 725. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, A.C.; Seman, B.G.; Hammond, J.H.; Archambault, L.S.; Hogan, D.A.; Wheeler, R.T. Candida albicans and Pseudomonas aeruginosa Interact To Enhance Virulence of Mucosal Infection in Transparent Zebrafish. Infect. Immun. 2017, 85, e00475-17. [Google Scholar] [CrossRef]
- Andes, D.; Nett, J.; Oschel, P.; Albrecht, R.; Marchillo, K.; Pitula, A. Development and characterization of an in vivo central venous catheter Candida albicans biofilm model. Infect. Immun. 2004, 72, 6023–6031. [Google Scholar] [CrossRef]
- Theberge, S.; Semlali, A.; Alamri, A.; Leung, K.P.; Rouabhia, M.C. albicansgrowth, transition, biofilm formation, and gene expression modulation by antimicrobial decapeptide KSL-W. BMC Microbiol. 2013, 13, 246. [Google Scholar] [CrossRef]
- Morici, P.; Fais, R.; Rizzato, C.; Tavanti, A.; Lupetti, A. Inhibition of Candida albicans Biofilm Formation by the Synthetic Lactoferricin Derived Peptide hLF1-11. PLoS ONE 2016, 11, e0167470. [Google Scholar] [CrossRef]
- Ribeiro, F.C.; de Barros, P.P.; Rossoni, R.D.; Junqueira, J.C.; Jorge, A.O. Lactobacillus rhamnosus inhibits Candida albicans virulence factors in vitro and modulates immune system in Galleria mellonella. J. Appl. Microbiol. 2017, 122, 201–211. [Google Scholar] [CrossRef]
- Kuhn, D.M.; Chandra, J.; Mukherjee, P.K.; Ghannoum, M.A. Comparison of biofilms formed by Candida albicans and Candida parapsilosis on bioprosthetic surfaces. Infect. Immun. 2002, 70, 878–888. [Google Scholar] [CrossRef] [Green Version]
- Hasan, F.; Xess, I.; Wang, X.; Jain, N.; Fries, B.C. Biofilm formation in clinical Candida isolates and its association with virulence. Microbes Infect. 2009, 11, 753–761. [Google Scholar] [CrossRef] [PubMed]
- Kernien, J.F.; Johnson, C.J.; Bayless, M.L.; Chovanec, J.F.; Nett, J.E. Neutrophils From Patients with Invasive Candidiasis Are Inhibited by Candida albicans Biofilms. Front. Immunol. 2020, 11, 587956. [Google Scholar] [CrossRef] [PubMed]
- Douglas, L.J. Candida biofilms and their role in infection. Trends Microbiol. 2003, 11, 30–36. [Google Scholar] [CrossRef]
- Rajendran, R.; May, A.; Sherry, L.; Kean, R.; Williams, C.; Jones, B.L.; Burgess, K.V.; Heringa, J.; Abeln, S.; Brandt, B.W.; et al. Integrating Candida albicans metabolism with biofilm heterogeneity by transcriptome mapping. Sci. Rep. 2016, 6, 35436. [Google Scholar] [CrossRef]
- Mohammadi, F.; Hemmat, N.; Bajalan, Z.; Javadi, A. Analysis of Biofilm-Related Genes and Antifungal Susceptibility Pattern of Vaginal Candida albicans and Non-Candida albicans Species. Biomed. Res. Int. 2021, 2021, 5598907. [Google Scholar] [CrossRef]
- Nailis, H.; Kucharíková, S.; Ricicová, M.; Van Dijck, P.; Deforce, D.; Nelis, H.; Coenye, T. Real-time PCR expression profiling of genes encoding potential virulence factors in Candida albicans biofilms: Identification of model-dependent and -independent gene expression. BMC Microbiol. 2010, 10, 114. [Google Scholar] [CrossRef]
- Desai, J.V.; Mitchell, A.P. Candida albicans Biofilm Development and Its Genetic Control. Microbiol. Spectr. 2015, 3, 99–114. [Google Scholar] [CrossRef]
- Aboualigalehdari, E.; Tahmasebi Birgani, M.; Fatahinia, M.; Hosseinzadeh, M. Transcription Factors of CAT1, EFG1, and BCR1 Are Effective in Persister Cells of Candida albicans-Associated HIV-Positive and Chemotherapy Patients. Front. Microbiol. 2021, 12, 651221. [Google Scholar] [CrossRef]
- Seman, B.G.; Moore, J.L.; Scherer, A.K.; Blair, B.A.; Manandhar, S.; Jones, J.M.; Wheeler, R.T. Yeast and Filaments Have Specialized, Independent Activities in a Zebrafish Model of Candida albicans Infection. Infect. Immun. 2018, 86, e00415-18. [Google Scholar] [CrossRef]
- Almeida, R.S.; Brunke, S.; Albrecht, A.; Thewes, S.; Laue, M.; Edwards, J.E.; Filler, S.G.; Hube, B. the hyphal-associated adhesin and invasin Als3 of Candida albicans mediates iron acquisition from host ferritin. PLoS Pathog. 2008, 4, e1000217. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sequence Name | Gene Function | Sequence (5′-3′) | Ta (°C) | References |
|---|---|---|---|---|
| Act1 | Housekeeping gene (Internal control) | F: GCTGGTAGAGACTTGACCAACCA R: GACAATTTCTCTTTCAGCACTAGTAGTGA | 54 | [27] |
| Eap1 | Cell–cell adhesion, mediates adhesion to biotic and abiotic surfaces | F: CTGCTCACTCAACTTCAATTGTCG R: GAACACATCCACCTTCGGGA | 54 | [27] |
| Als3 | Mediates attachment to host cells and matrix proteins | F: GGTTATCGTCCATTTGTTGA R: TTCTGTATCCAGTCCATCTT | 54 | [28] |
| Hwp1 | Cell adherence | F: ACAGGTAGACGGTCAAGG R: GGGTAATCATCACATGGTTC | 50 | [28] |
| Bcr1 | Transcription factor controls the als1, als3, hwp1, and hyr1 expressions | F: AATGCCTGCAGGTTATTTGG R: TTTTAGGTGGTGGTGGCAAT | 50 | [29] |
| Mkc1 | Induced invasive filamentation and cell growth under stressed condition | F: AATGGGTCCAAAAAGGTTCC R: TTATGGCCCCTGAAGAACTG | 50 | This study |
| C. albicans Strains Used | Phospholipase Activity | Proteinase Activity | Hemolytic Activity | Biofilm Activity | ||||
|---|---|---|---|---|---|---|---|---|
| Pz Value | Interpretation | Pr Value | Interpretation | Hz Value | Interpretation | OD | Interpretation | |
| No. 104 | 1.00 | - | 1.00 | - | 0.59 | ++ | 0.031 | No |
| No. 57 | 1.00 | - | 1.00 | - | 0.52 | ++ | 0.073 | Weak |
| No. 140 | 1.00 | - | 1.00 | - | 0.53 | ++ | 0.396 | Strong |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pokhrel, S.; Boonmee, N.; Tulyaprawat, O.; Pharkjaksu, S.; Thaipisutikul, I.; Chairatana, P.; Ngamskulrungroj, P.; Mitrpant, C. Assessment of Biofilm Formation by Candida albicans Strains Isolated from Hemocultures and Their Role in Pathogenesis in the Zebrafish Model. J. Fungi 2022, 8, 1014. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8101014
Pokhrel S, Boonmee N, Tulyaprawat O, Pharkjaksu S, Thaipisutikul I, Chairatana P, Ngamskulrungroj P, Mitrpant C. Assessment of Biofilm Formation by Candida albicans Strains Isolated from Hemocultures and Their Role in Pathogenesis in the Zebrafish Model. Journal of Fungi. 2022; 8(10):1014. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8101014
Chicago/Turabian StylePokhrel, Sabi, Nawarat Boonmee, Orawan Tulyaprawat, Sujiraphong Pharkjaksu, Iyarit Thaipisutikul, Phoom Chairatana, Popchai Ngamskulrungroj, and Chalermchai Mitrpant. 2022. "Assessment of Biofilm Formation by Candida albicans Strains Isolated from Hemocultures and Their Role in Pathogenesis in the Zebrafish Model" Journal of Fungi 8, no. 10: 1014. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8101014