
Fungal Proteases as Emerging Biocatalysts to Meet the Current Challenges and Recent Developments in Biomedical Therapies: An Updated Review

 ,
, 
 and
and 
Abstract
:1. Introduction
2. Fungal Enzymes
Coproduction of Fungal Enzymes
3. Fungal Proteases
3.1. Origin of Fungal Proteases
3.2. Classification of Fungal Proteases
3.2.1. Protease Classes Based on Amino Acids
Serine Protease
Threonine Protease
Cysteine Protease
Aspartate Protease
Glutamic Acid Protease
Metalloprotease

4. Market Value of Fungal Proteases
Global Proteases Market Segmentation
5. Molecular Approaches, Cloning and Expression of Fungal Proteases
6. Role of CRISPR Technology in Fungal Enzymes
7. Recent Developments
8. Role of Fungal Proteases in Living Organisms
8.1. Mitoproteases Function in Protein Processing and Activation
8.2. Cysteine Proteases in Atherosclerosis
8.3. Role of Fungal Proteases in the Pathogenesis of Chronic Rhinosinusitis with Nasal Polyps
8.4. Fungal Protease as a New Therapeutic Strategy for Colorectal Cancer
8.5. Protease as a New Therapeutic Strategy for Coeliac Disease
8.6. Protease as a New Therapeutic Strategy for Neurological Disorders
9. Industrial Applications
9.1. Food Industry
9.2. Waste Management and Bioremediation
9.3. Medical and Pharmaceutical Industry
9.4. Proteases in Silver Recovery
9.5. Proteases in Silk-Degumming
10. Novel Protease Inhibitors
10.1. Protease Inhibitors in Clinical Trials
10.1.1. HIV
10.1.2. HCV
10.1.3. Cancer
10.2. Mechanisms of Action for Protease Inhibitors
10.3. Discovery of Protease Inhibitors against COVID-19
10.4. NP-Delivery Systems for Discovery of Protease Inhibitors
11. Current Challenges and Future Perspective
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Monod, M.; Capoccia, S.; Léchenne, B.; Zaugg, C.; Holdom, M.; Jousson, O. Secreted proteases from pathogenic fungi. Int. J. Med. Microbiol. 2002, 292, 405–419. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Mittal, A.; Kumar, M.; Mehta, P.K. Microbial proteases in commercial applications. J. Pharm. Chem. Biol. Sci. 2016, 4, 365–374. [Google Scholar]
- Rao, M.B.; Tanksale, A.M.; Ghatge, M.S.; Deshpande, V.V. Molecular and biotechnological aspects of microbial proteases. Microbiol. Mol. Biol. Rev. 1998, 62, 597–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.; Gaur, S. GRAS Fungi: A New Horizon in Safer Food Product. In Fungi in Sustainable Food Production; Springer: Cham, Switzerland, 2021; pp. 27–37. [Google Scholar]
- Novelli, P.K.; Barros, M.M.; Fleuri, L.F. Novel inexpensive fungi proteases: Production by solid state fermentation and characterization. Food Chem. 2016, 198, 119–124. [Google Scholar] [CrossRef]
- Souza, P.M.d.; Bittencourt, M.L.d.A.; Caprara, C.C.; Freitas, M.d.; Almeida, R.P.C.d.; Silveira, D.; Fonseca, Y.M.; Ferreira, E.X.; Pessoa, A.; Magalhães, P.O. A biotechnology perspective of fungal proteases. Braz. J. Microbiol. 2015, 46, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Gautam, A.; Dutt, D. Screening of fungal resources for the production of cellulases and xylanases. Biotechnol. J. Int. 2015, 9, 1–13. [Google Scholar] [CrossRef]
- Lange, L. Fungal enzymes and yeasts for conversion of plant biomass to bioenergy and high-value products. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Damare, S.; Raghukumar, C.; Muraleedharan, U.D.; Raghukumar, S. Deep-sea fungi as a source of alkaline and cold-tolerant proteases. Enzym. Microb. Technol. 2006, 39, 172–181. [Google Scholar] [CrossRef]
- Kumar, A. Aspergillus nidulans: A Potential Resource of the Production of the Native and Heterologous Enzymes for Industrial Applications. Int. J. Microbiol. 2020, 2020, 8894215. [Google Scholar] [CrossRef]
- Gnanadoss, J.J.; Devi, S.K. Optimization of nutritional and culture conditions for improved protease production by Aspergillus nidulans and Aspergillus flavus. J. Microbiol. Biotechnol. Food Sci. 2021, 2021, 518–523. [Google Scholar] [CrossRef] [Green Version]
- Hoffmeister, D.; Keller, N.P. Natural products of filamentous fungi: Enzymes, genes, and their regulation. Nat. Prod. Rep. 2007, 24, 393–416. [Google Scholar] [CrossRef]
- Savitha, S.; Sadhasivam, S.; Swaminathan, K.; Lin, F.H. Fungal protease: Production, purification and compatibility with laundry detergents and their wash performance. J. Taiwan Inst. Chem. Eng. 2011, 42, 298–304. [Google Scholar] [CrossRef]
- Liu, E.; Li, M.; Abdella, A.; Wilkins, M.R. Development of a cost-effective medium for submerged production of fungal aryl alcohol oxidase using a genetically modified Aspergillus nidulans strain. Bioresour. Technol. 2020, 305, 123038. [Google Scholar] [CrossRef]
- Sharma, M.; Gat, Y.; Arya, S.; Kumar, V.; Panghal, A.; Kumar, A. A review on microbial alkaline protease: An essential tool for various industrial approaches. Ind. Biotechnol. 2019, 15, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Sabotič, J.; Kos, J. Microbial and fungal protease inhibitors—current and potential applications. Appl. Microbiol. Biotechnol. 2012, 93, 1351–1375. [Google Scholar] [CrossRef]
- Agbowuro, A.A.; Huston, W.M.; Gamble, A.B.; Tyndall, J.D. Proteases and protease inhibitors in infectious diseases. Med. Res. Rev. 2018, 38, 1295–1331. [Google Scholar] [CrossRef]
- Bezerra, V.H.S.; Cardoso, S.L.; Fonseca-Bazzo, Y.; Silveira, D.; Magalhães, P.O.; Souza, P.M. Protease Produced by Endophytic Fungi: A Systematic Review. Molecules 2021, 26, 7062. [Google Scholar] [CrossRef]
- Gupta, V.K.; Kubicek, C.P.; Berrin, J.-G.; Wilson, D.W.; Couturier, M.; Berlin, A.; Edivaldo Filho, X.; Ezeji, T. Fungal enzymes for bio-products from sustainable and waste biomass. Trends Biochem. Sci. 2016, 41, 633–645. [Google Scholar] [CrossRef] [Green Version]
- Yike, I. Fungal proteases and their pathophysiological effects. Mycopathologia 2011, 171, 299–323. [Google Scholar] [CrossRef] [PubMed]
- Manganyi, M.C.; Ateba, C.N. Untapped potentials of endophytic fungi: A review of novel bioactive compounds with biological applications. Microorganisms 2020, 8, 1934. [Google Scholar] [CrossRef]
- Srilakshmi, J.; Madhavi, J.; Lavanya, S.; Ammani, K. Commercial potential of fungal protease: Past, present and future prospects. J. Pharm. Chem. Biol. Sci. 2015, 2, 218–234. [Google Scholar]
- Adrio, J.L.; Demain, A.L. Fungal biotechnology. Int. Microbiol. 2003, 6, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Guillemette, T.; van Peij, N.N.; Goosen, T.; Lanthaler, K.; Robson, G.D.; van den Hondel, C.A.; Stam, H.; Archer, D.B. Genomic analysis of the secretion stress response in the enzyme-producing cell factory Aspergillus niger. BMC Genom. 2007, 8, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nevalainen, K.H.; Te’o, V.S.; Bergquist, P.L. Heterologous protein expression in filamentous fungi. Trends Biotechnol. 2005, 23, 468–474. [Google Scholar] [CrossRef]
- Paloheimo, M.; Haarmann, T.; Mäkinen, S.; Vehmaanperä, J. Production of industrial enzymes in Trichoderma reesei. In Gene Expression Systems in Fungi: Advancements and Applications; Springer: Berlin/Heidelberg, Germany, 2016; pp. 23–57. [Google Scholar]
- Benito, M.J.; Connerton, I.F.; Córdoba, J.J. Genetic characterization and expression of the novel fungal protease, EPg222 active in dry-cured meat products. Appl. Microbiol. Biotechnol. 2006, 73, 356–365. [Google Scholar] [CrossRef]
- Li, C.; Xu, D.; Zhao, M.; Sun, L.; Wang, Y. Production optimization, purification, and characterization of a novel acid protease from a fusant by Aspergillus oryzae and Aspergillus niger. Eur. Food Res. Technol. 2014, 238, 905–917. [Google Scholar] [CrossRef]
- Salazar-Cerezo, S.; Kun, R.S.; de Vries, R.P.; Garrigues, S. CRISPR/Cas9 technology enables the development of the filamentous ascomycete fungus Penicillium subrubescens as a new industrial enzyme producer. Enzym. Microb. Technol. 2020, 133, 109463. [Google Scholar] [CrossRef]
- Satish, L.; Shamili, S.; Muthubharathi, B.C.; Ceasar, S.A.; Kushmaro, A.; Singh, V.; Sitrit, Y. CRISPR-Cas9 System for Fungi Genome Engineering Toward Industrial Applications. In Genome Engineering via CRISPR-Cas9 System; Elsevier: Amsterdam, The Netherlands, 2020; pp. 69–81. [Google Scholar]
- Liu, R.; Chen, L.; Jiang, Y.; Zhou, Z.; Zou, G. Efficient genome editing in filamentous fungus Trichoderma reesei using the CRISPR/Cas9 system. Cell Discov. 2015, 1, 15007. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.A.; Ali, S.; Hassan, A.; Tahir, H.M.; Mumtaz, S.; Mumtaz, S. Biosynthesis and industrial applications of α-amylase: A review. Arch. Microbiol. 2021, 203, 1281–1292. [Google Scholar] [CrossRef]
- Singh, A.; Bajar, S.; Devi, A.; Pant, D. An overview on the recent developments in fungal cellulase production and their industrial applications. Bioresour. Technol. Rep. 2021, 14, 100652. [Google Scholar] [CrossRef]
- Mehta, A.; Guleria, S.; Sharma, R.; Gupta, R. The Lipases and Their Applications with Emphasis on Food Industry. In Microbial Biotechnology in Food and Health; Elsevier: Amsterdam, The Netherlands, 2021; pp. 143–164. [Google Scholar]
- Anitha, T.; Palanivelu, P. Purification and characterization of an extracellular keratinolytic protease from a new isolate of Aspergillus parasiticus. Protein Expr. Purif. 2013, 88, 214–220. [Google Scholar] [CrossRef]
- Zdarta, J.; Jędrzak, A.; Klapiszewski, Ł.; Jesionowski, T. Immobilization of cellulase on a functional inorganic–organic hybrid support: Stability and kinetic study. Catalysts 2017, 7, 374. [Google Scholar] [CrossRef] [Green Version]
- Khan, F.I.; Bisetty, K.; Singh, S.; Permaul, K.; Hassan, M.I. Chitinase from Thermomyces lanuginosus SSBP and its biotechnological applications. Extremophiles 2015, 19, 1055–1066. [Google Scholar] [CrossRef]
- Sikandar, S.; Ujor, V.C.; Ezeji, T.C.; Rossington, J.L.; Michel, F.C., Jr.; McMahan, C.M.; Ali, N.; Cornish, K. Thermomyces lanuginosus STm: A source of thermostable hydrolytic enzymes for novel application in extraction of high-quality natural rubber from Taraxacum kok-saghyz (Rubber dandelion). Ind. Crops Prod. 2017, 103, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Dagenais, T.R.; Keller, N.P. Pathogenesis of Aspergillus fumigatus in invasive aspergillosis. Clin. Microbiol. Rev. 2009, 22, 447–465. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Singh, S.; Bali, V.; Sharma, L.; Mangla, J. Production of fungal amylases using cheap, readily available agriresidues, for potential application in textile industry. BioMed Res. Int. 2014, 2014, 215748. [Google Scholar] [CrossRef] [Green Version]
- Wanderley, K.J.; Torres, F.A.; Moraes, L.M.; Ulhoa, C.J. Biochemical characterization of α-amylase from the yeast Cryptococcus flavus. FEMS Microbiol. Lett. 2004, 231, 165–169. [Google Scholar] [CrossRef] [Green Version]
- Stojanović, J.; Jakovljević, V.; Matović, I.; Gajović, O.; Mijušković, Z.; Nedeljković, T. Influence of detergent on metabolic activity of fungi Aspergillus niger. Nat. Sci. 2011, 3, 466–470. [Google Scholar] [CrossRef] [Green Version]
- Saleem, A.; Ebrahim, M.K. Production of amylase by fungi isolated from legume seeds collected in Almadinah Almunawwarah, Saudi Arabia. J. Taibah Univ. Sci. 2014, 8, 90–97. [Google Scholar] [CrossRef]
- Souza, P.M.d.; Magalhães, P.d.O. Application of microbial α-amylase in industry-A review. Braz. J. Microbiol. 2010, 41, 850–861. [Google Scholar] [CrossRef]
- Alves, M.H.; Campos-Takaki, G.M.; Porto, A.L.F.; Milanez, A.I. Screening of Mucor spp. for the production of amylase, lipase, polygalacturonase and protease. Braz. J. Microbiol. 2002, 33, 325–330. [Google Scholar] [CrossRef] [Green Version]
- Rahardjo, Y.S.; Weber, F.J.; Le Comte, E.P.; Tramper, J.; Rinzema, A. Contribution of aerial hyphae of Aspergillus oryzae to respiration in a model solid-state fermentation system. Biotechnol. Bioeng. 2002, 78, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, M.; Jeevarathinam, G.; Kumar, S.K.S.; Muniraj, I.; Uthandi, S. Optimization and scale-up of α-amylase production by Aspergillus oryzae using solid-state fermentation of edible oil cakes. BMC Biotechnol. 2021, 21, 33. [Google Scholar] [CrossRef] [PubMed]
- Saranraj, P.; Stella, D. Fungal amylase—a review. Int. J. Microbiol. Res. 2013, 4, 203–211. [Google Scholar]
- Sethi, B.K.; Jana, A.; Nanda, P.K.; DasMohapatra, P.K.; Sahoo, S.L.; Patra, J.K. Production of α-amylase by Aspergillus terreus NCFT 4269.10 using pearl millet and its structural characterization. Front. Plant Sci. 2016, 7, 639. [Google Scholar] [CrossRef] [Green Version]
- Khajuria, R.; Singh, S. Fungal Amylases for the Detergent Industry. In Microbes in Agriculture and Environmental Development; CRC Press: Boca Raton, FL, USA, 2020; pp. 153–164. [Google Scholar]
- Niyonzima, F.N.; More, S. Purification and properties of detergent-compatible extracellular alkaline protease from Scopulariopsis spp. Prep. Biochem. Biotechnol. 2014, 44, 738–759. [Google Scholar] [CrossRef]
- Zahra, T.; Irfan, M.; Nadeem, M.; Ghazanfar, M.; Ahmad, Q.; Ali, S.; Siddique, F.; Yasmeen, Z.; Franco, M. Cellulase Production by Trichoderma viride in Submerged Fermentation using Response Surface Methodology. Punjab Univ. J. Zool. 2020, 35, 223–228. [Google Scholar] [CrossRef]
- Niyonzima, F.N. Detergent-compatible fungal cellulases. Folia Microbiol. 2020, 66, 25–40. [Google Scholar] [CrossRef]
- Imran, M.; Anwar, Z.; Zafar, M.; Ali, A.; Arif, M. Production and characterization of commercial cellulase produced through Aspergillus niger IMMIS1 after screening fungal species. Pak. J. Bot. 2018, 50, 1563–1570. [Google Scholar]
- Imran, M.; Anwar, Z.; Irshad, M.; Javid, A.; Hussain, A.; Ali, S. Optimization of cellulase production from a novel strain of Aspergillus tubingensis IMMIS2 through response surface methodology. Biocatal. Agric. Biotechnol. 2017, 12, 191–198. [Google Scholar] [CrossRef]
- da Silva Delabona, P.; Lima, D.J.; Robl, D.; Rabelo, S.C.; Farinas, C.S.; da Cruz Pradella, J.G. Enhanced cellulase production by Trichoderma harzianum by cultivation on glycerol followed by induction on cellulosic substrates. J. Ind. Microbiol. Biotechnol. 2016, 43, 617–626. [Google Scholar] [CrossRef]
- Pham, T.H.; Quyen, D.T.; Nghiem, N.M. Purification and properties of an endoglucanase from Aspergillus niger VTCC-F021. Turk. J. Biol. 2012, 36, 694–701. [Google Scholar]
- Pham, T.H.; Quyen, D.T.; Nghiem, N.M. Optimization of endoglucanase production by Aspergillus niger VTCC-F021. Aust. J. Basic Appl. Sci. 2010, 6, 4151–5157. [Google Scholar]
- Niyonzima, F.N.; More, S.S. Purification and characterization of detergent-compatible protease from Aspergillus terreus gr. 3 Biotech 2015, 5, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Hernández, M.S.; Rodríguez, M.R.; Guerra, N.P.; Rosés, R.P. Amylase production by Aspergillus niger in submerged cultivation on two wastes from food industries. J. Food Eng. 2006, 73, 93–100. [Google Scholar] [CrossRef]
- Sethi, B.K.; Rout, J.R.; Das, R.; Nanda, P.K.; Sahoo, S.L. Lipase production by Aspergillus terreus using mustard seed oil cake as a carbon source. Ann. Microbiol. 2013, 63, 241–252. [Google Scholar] [CrossRef]
- Choudhary, V. Production, isolation and characterization of alkaline protease from Aspergillus versicolor PF/F/107. J. Acad. Indus. Res. 2012, 1, 272–277. [Google Scholar]
- Gopinath, S.C.; Hilda, A.; Anbu, P. Extracellular enzymatic activity profiles in fungi isolated from oil-rich environments. Mycoscience 2005, 46, 119–126. [Google Scholar] [CrossRef]
- da Silva, O.S.; de Almeida, E.M.; de Melo, A.H.F.; Porto, T.S. Purification and characterization of a novel extracellular serine-protease with collagenolytic activity from Aspergillus tamarii URM4634. Int. J. Biol. Macromol. 2018, 117, 1081–1088. [Google Scholar] [CrossRef]
- Dayanandan, A.; Rani, S.; Shanmugavel, M.; Gnanamani, A.; Rajakumar, G.S. Enhanced production of Aspergillus tamarii lipase for recovery of fat from tannery fleshings. Braz. J. Microbiol. 2013, 44, 1089–1095. [Google Scholar] [CrossRef] [Green Version]
- Pasin, T.; Benassi, V.; Moreira, E.; Jorge, J.; Polizeli, M. Prospecting filamentous fungi for amylase production: Standardization of Aspergillus japonicus culture conditions. Biotechnol. J. Int. 2014, 482–498. [Google Scholar] [CrossRef]
- Souza, L.T.A.; Oliveira, J.S.; dos Santos, V.L.; Regis, W.C.; Santoro, M.M.; Resende, R.R. Lipolytic potential of Aspergillus japonicus LAB01: Production, partial purification, and characterisation of an extracellular lipase. BioMed Res. Int. 2014, 2014, 108913. [Google Scholar] [CrossRef] [Green Version]
- Karanam, S.K.; Medicherla, N.R. Enhanced lipase production by mutation induced Aspergillus japonicus. Afr. J. Biotechnol. 2008, 7. [Google Scholar] [CrossRef]
- Niyonzima, F.N.; Veena, S.; More, S.S. Industrial Production and Optimization of Microbial Enzymes. In Microbial Enzymes: Roles and Applications in Industries; Springer: Singapore, 2020; pp. 115–135. [Google Scholar]
- Tamminen, A.; Kramer, A.; Labes, A.; Wiebe, M.G. Production of scopularide A in submerged culture with Scopulariopsis brevicaulis. Microb. Cell Factories 2014, 13, 89. [Google Scholar] [CrossRef] [Green Version]
- Macchione, M.M.; Merheb, C.W.; Gomes, E.; Da Silva, R. Protease production by different thermophilic fungi. Appl. Biochem. Biotechnol. 2008, 146, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Larcher, G.; Bouchara, J.-P.; Annaix, V.; Symoens, F.; Chabasse, D.; Tronchin, G. Purification and characterization of a fibrinogenolytic serine proteinase from Aspergillus fumigatus culture filtrate. FEBS Lett. 1992, 308, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Farnell, E.; Rousseau, K.; Thornton, D.J.; Bowyer, P.; Herrick, S.E. Expression and secretion of Aspergillus fumigatus proteases are regulated in response to different protein substrates. Fungal Biol. 2012, 116, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Sharma, N.S.; Saharan, M.R.; Singh, R. Extracellular acid protease from Rhizopus oryzae: Purification and characterization. Process Biochem. 2005, 40, 1701–1705. [Google Scholar] [CrossRef]
- Benabda, O.; M’hir, S.; Kasmi, M.; Mnif, W.; Hamdi, M. Optimization of protease and amylase production by Rhizopus oryzae cultivated on bread waste using solid-state fermentation. J. Chem. 2019, 2019, 3738181. [Google Scholar] [CrossRef] [Green Version]
- Abraham, L.D.; Breuil, C. Isolation and characterization of a subtilisin-like serine proteinase secreted by the sap-staining fungus Ophiostoma piceae. Enzym. Microb. Technol. 1996, 18, 133–140. [Google Scholar] [CrossRef]
- de Oliveira, T.B.; Gomes, E.; Rodrigues, A. Thermophilic fungi in the new age of fungal taxonomy. Extremophiles 2015, 19, 31–37. [Google Scholar] [CrossRef]
- Zucconi, L.; Pagano, S.; Fenice, M.; Selbmann, L.; Tosi, S.; Onofri, S. Growth temperature preferences of fungal strains from Victoria Land, Antarctica. Polar Biol. 1996, 16, 53–61. [Google Scholar] [CrossRef]
- Lasa, I.; Berenguer, J. Thermophilic enzymes and their biotechnological potential. Microbiologia 1993, 9, 77–89. [Google Scholar]
- Mikhailova, A.G.; Khairullin, R.F.; Demidyuk, I.V.; Kostrov, S.V.; Grinberg, N.V.; Burova, T.V.; Grinberg, V.Y.; Rumsh, L.D. Cloning, sequencing, expression, and characterization of thermostability of oligopeptidase B from Serratia proteamaculans, a novel psychrophilic protease. Protein Expr. Purif. 2014, 93, 63–76. [Google Scholar] [CrossRef]
- Wancura, J.H.; Rosset, D.V.; Tres, M.V.; Oliveira, J.V.; Mazutti, M.A.; Jahn, S.L. Production of biodiesel catalyzed by lipase from Thermomyces lanuginosus in its soluble form. Can. J. Chem. Eng. 2018, 96, 2361–2368. [Google Scholar] [CrossRef]
- Pathak, A.P.; Rathod, M.G. A Review on Alkaline Protease Producers and Their Biotechnological Perspectives; NISCAIR-CSIR: New Delhi, India, 2018; pp. 1113–1119. [Google Scholar]
- Huynh, H.H.; Arioka, M. Functional expression and characterization of a glucuronoyl esterase from the fungus Neurospora crassa: Identification of novel consensus sequences containing the catalytic triad. J. Gen. Appl. Microbiol. 2016, 62, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Singh, B. Myceliophthora thermophila syn. Sporotrichum thermophile: A thermophilic mould of biotechnological potential. Crit. Rev. Biotechnol. 2016, 36, 59–69. [Google Scholar] [CrossRef]
- de Oliveira, T.B.; Rodrigues, A. Ecology of Thermophilic Fungi. Fungi in Extreme Environments: Ecological Role and Biotechnological Significance; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; pp. 39–57. [Google Scholar]
- Singh, B.; Satyanarayana, T. Phytase production by a thermophilic mould Sporotrichum thermophile in solid state fermentation and its potential applications. Bioresour. Technol. 2008, 99, 2824–2830. [Google Scholar] [CrossRef]
- Dilokpimol, A.; Mäkelä, M.R.; Cerullo, G.; Zhou, M.; Varriale, S.; Gidijala, L.; Brás, J.L.; Jütten, P.; Piechot, A.; Verhaert, R. Fungal glucuronoyl esterases: Genome mining based enzyme discovery and biochemical characterization. New Biotechnol. 2018, 40, 282–287. [Google Scholar] [CrossRef] [Green Version]
- Salwan, R.; Sharma, V. Proteases from Extremophilic Fungi: A Tool for White Biotechnology. In Recent Advancement in White Biotechnology Through Fungi; Springer: Berlin/Heidelberg, Germany, 2019; pp. 299–317. [Google Scholar]
- Kieliszek, M.; Kot, A.M.; Bzducha-Wróbel, A.; BŁażejak, S.; Gientka, I.; Kurcz, A. Biotechnological use of Candida yeasts in the food industry: A review. Fungal Biol. Rev. 2017, 31, 185–198. [Google Scholar] [CrossRef]
- Naveed, M.; Nadeem, F.; Mehmood, T.; Bilal, M.; Anwar, Z.; Amjad, F. Protease—a versatile and ecofriendly biocatalyst with multi-industrial applications: An updated review. Catal. Lett. 2021, 151, 307–323. [Google Scholar] [CrossRef]
- El-Khonezy, M.I.; Elgammal, E.W.; Ahmed, E.F.; Abd-Elaziz, A.M. Detergent stable thiol-dependant alkaline protease produced from the endophytic fungus Aspergillus ochraceus BT21: Purification and kinetics. Biocatal. Agric. Biotechnol. 2021, 35, 102046. [Google Scholar] [CrossRef]
- Niyonzima, F.N.; More, S.S. Screening and optimization of cultural parameters for an alkaline protease production by Aspergillus terreus gr. under submerged fermentation. Int. J. Pharm. Bio. Sci. 2013, 4, 1016–1028. [Google Scholar]
- Yadav, V.K.; Singh, V.; Mishra, V. Alkaline Protease: A Tool to Manage Solid Waste and Its Utility in Detergent Industry. In Microbial Genomics in Sustainable Agroecosystems; Springer: Berlin/Heidelberg, Germany, 2019; pp. 231–254. [Google Scholar]
- Martinelli, P.; Rugarli, E.I. Emerging roles of mitochondrial proteases in neurodegeneration. Biochim. Et Biophys. Acta (BBA)-Bioenerg. 2010, 1797, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hariharan, A.; Rajadurai, U.M.; Palanivel, I. Isolation, Purification and Mass Production of Protease from Bacillus subtilis. 2019. Available online: https://ssrn.com/abstract=3370124 (accessed on 15 July 2021).
- Devi, M.K.; Banu, A.R.; Gnanaprabhal, G.; Pradeep, B.; Palaniswamy, M. Purification, characterization of alkaline protease enzyme from native isolate Aspergillus niger and its compatibility with commercial detergents. Indian J. Sci. Technol. 2008, 1, 1–6. [Google Scholar] [CrossRef]
- Shahid, T.; Muhammad, S.; Ahmed, K. Enzyme Proteases Used in Laundry Detergents Engineering a Review. Sci. Int. 2016, 28, 2711–2717. [Google Scholar]
- Corvo, I.; Ferraro, F.; Merlino, A.; Zuberbühler, K.; O’Donoghue, A.J.; Pastro, L.; Pi-Denis, N.; Basika, T.; Roche, L.; McKerrow, J.H. Substrate specificity of cysteine proteases beyond the S2 Pocket: Mutagenesis and molecular dynamics investigation of Fasciola hepatica Cathepsins L. Front. Mol. Biosci. 2018, 5, 40. [Google Scholar] [CrossRef] [Green Version]
- Ou, J.-F.; Zhu, M.-J. An overview of current and novel approaches for microbial neutral protease improvement. Int. J. Mod. Biol. Med 2012, 2, 1–31. [Google Scholar]
- Theron, L.W.; Divol, B. Microbial aspartic proteases: Current and potential applications in industry. Appl. Microbiol. Biotechnol. 2014, 98, 8853–8868. [Google Scholar] [CrossRef]
- Dunn, B.M. Introduction to the aspartic proteinase family. Aspartic Acid Proteases Ther. Targets 2010, 45, 1–21. [Google Scholar]
- Jiang, L.; Zhang, X.; Zhou, Y.; Chen, Y.; Luo, Z.; Li, J.; Yuan, C.; Huang, M. Halogen bonding for the design of inhibitors by targeting the S1 pocket of serine proteases. RSC Adv. 2018, 8, 28189–28197. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Yuan, C.; Huang, M. A general strategy to inhibit serine protease by targeting its autolysis loop. FASEB J. 2021, 35, e21259. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J. [13] Evolutionary families of metallopeptidases. Methods Enzymol. 1995, 248, 183–228. [Google Scholar]
- Cheng, M.; Takenaka, S.; Aoki, S.; Murakami, S.; Aoki, K. Purification and characterization of an eggshell membrane decomposing protease from Pseudomonas aeruginosa strain ME-4. J. Biosci. Bioeng. 2009, 107, 373–378. [Google Scholar] [CrossRef]
- Sims, A.H.; Dunn-Coleman, N.S.; Robson, G.D.; Oliver, S.G. Glutamic protease distribution is limited to filamentous fungi. FEMS Microbiol. Lett. 2004, 239, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Kuan, C.S.; Ng, K.P.; Yew, S.M.; Meleh, H.U.; Seow, H.F.; How, K.N.; Yeo, S.K.; Jee, J.M.; Tan, Y.-C.; Yee, W.-Y. Comparative Genomic and Phenotypic Analyses of Pathogenic Fungi Neoscytalidium Dimidiatum and Bipolaris Papendorfii, Isolated From Human Skin Scraping. Res. Sq. 2020, 1–32. [Google Scholar] [CrossRef]
- Mamo, J.; Assefa, F. The role of microbial aspartic protease enzyme in food and beverage industries. J. Food Qual. 2018, 2018, 7957269. [Google Scholar] [CrossRef] [Green Version]
- Razzaq, A.; Shamsi, S.; Ali, A.; Ali, Q.; Sajjad, M.; Malik, A.; Ashraf, M. Microbial proteases applications. Front. Bioeng. Biotechnol. 2019, 7, 110. [Google Scholar] [CrossRef] [Green Version]
- Erjavec, J.; Kos, J.; Ravnikar, M.; Dreo, T.; Sabotič, J. Proteins of higher fungi–from forest to application. Trends Biotechnol. 2012, 30, 259–273. [Google Scholar] [CrossRef]
- Shaba, A.; Baba, J. Screening of Pleurotus ostreatus and Gleophylum sepiarium strains for extracellular protease enzyme production. Bayero J. Pure Appl. Sci. 2012, 5, 187–190. [Google Scholar] [CrossRef] [Green Version]
- Papagianni, M. Fungal morphology and metabolite production in submerged mycelial processes. Biotechnol. Adv. 2004, 22, 189–259. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ng, T. Pleureryn, a novel protease from fresh fruiting bodies of the edible mushroom Pleurotus eryngii. Biochem. Biophys. Res. Commun. 2001, 289, 750–755. [Google Scholar] [CrossRef]
- Musatti, A.; Ficara, E.; Mapelli, C.; Sambusiti, C.; Rollini, M. Use of solid digestate for lignocellulolytic enzymes production through submerged fungal fermentation. J. Environ. Manag. 2017, 199, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Burdsall, H.H., Jr.; Volk, T.J. Armillaria solidipes, an older name for the fungus called Armillaria ostoyae. North Am. Fungi 2008, 3, 261–267. [Google Scholar] [CrossRef]
- Faraco, V.; Palmieri, G.; Festa, G.; Monti, M.; Sannia, G.; Giardina, P. A new subfamily of fungal subtilases: Structural and functional analysis of a Pleurotus ostreatus member. Microbiology 2005, 151, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Nurika, I.; Suhartini, S.; Barker, G.C. Biotransformation of tropical lignocellulosic feedstock using the brown rot fungus Serpula lacrymans. Waste Biomass Valorization 2020, 11, 2689–2700. [Google Scholar] [CrossRef]
- Cha, W.-S.; Park, S.-S.; Kim, S.-J.; Choi, D. Biochemical and enzymatic properties of a fibrinolytic enzyme from Pleurotus eryngii cultivated under solid-state conditions using corn cob. Bioresour. Technol. 2010, 101, 6475–6481. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.B.; Wong, J.H.; Cheung, R.C.F.; Tse, T.F.; Tam, T.; Chan, H. Mushrooms: Proteins, polysaccharidepeptide complexes and polysaccharides with antiproliferative and anticancer activities. Int. J. Cancer Res. Prev. 2014, 7, 287. [Google Scholar]
- Lv, H.; Kong, Y.; Yao, Q.; Zhang, B.; Leng, F.-w.; Bian, H.-j.; Balzarini, J.; Van Damme, E.; Bao, J.-k. Nebrodeolysin, a novel hemolytic protein from mushroom Pleurotus nebrodensis with apoptosis-inducing and anti-HIV-1 effects. Phytomedicine 2009, 16, 198–205. [Google Scholar] [CrossRef]
- Berne, S.; Križaj, I.; Pohleven, F.; Turk, T.; Maček, P.; Sepčić, K. Pleurotus and Agrocybe hemolysins, new proteins hypothetically involved in fungal fruiting. Biochim. Et Biophys. Acta (BBA) Gen. Subj. 2002, 1570, 153–159. [Google Scholar] [CrossRef]
- Sumantha, A.; Larroche, C.; Pandey, A. Microbiology and industrial biotechnology of food-grade proteases: A perspective. Food Technol. Biotechnol. 2006, 44, 211. [Google Scholar]
- Sandhya, C.; Nampoothiri, K.M.; Pandey, A. Microbial Proteases. In Microbial Enzymes and Biotransformations; Springer: Berlin/Heidelberg, Germany, 2005; pp. 165–179. [Google Scholar]
- dos Santos Aguilar, J.G.; Sato, H.H. Microbial proteases: Production and application in obtaining protein hydrolysates. Food Res. Int. 2018, 103, 253–262. [Google Scholar] [CrossRef]
- Baird, T.T., Jr.; Wright, W.D.; Craik, C.S. Conversion of trypsin to a functional threonine protease. Protein Sci. 2006, 15, 1229–1238. [Google Scholar] [CrossRef] [Green Version]
- Jashni, M.K.; Dols, I.H.; Iida, Y.; Boeren, S.; Beenen, H.G.; Mehrabi, R.; Collemare, J.; de Wit, P.J. Synergistic action of a metalloprotease and a serine protease from Fusarium oxysporum f. sp. lycopersici cleaves chitin-binding tomato chitinases, reduces their antifungal activity, and enhances fungal virulence. Mol. Plant-Microbe Interact. 2015, 28, 996–1008. [Google Scholar] [CrossRef] [Green Version]
- Barrett, A.J. [1] Classification of peptidases. Methods Enzymol. 1994, 244, 1–15. [Google Scholar] [CrossRef]
- Brocklehurst, K.; Philpott, M.P. Cysteine proteases: Mode of action and role in epidermal differentiation. Cell Tissue Res. 2013, 351, 237–244. [Google Scholar] [CrossRef]
- Yegin, S.; Fernandez-Lahore, M.; Salgado, A.J.G.; Guvenc, U.; Goksungur, Y.; Tari, C. Aspartic proteinases from Mucor spp. in cheese manufacturing. Appl. Microbiol. Biotechnol. 2011, 89, 949–960. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, R.R.; Souto, T.B.; de Oliveira, T.B.; de Oliveira, L.C.G.; Karcher, D.; Juliano, M.A.; Juliano, L.; de Oliveira, A.H.; Rodrigues, A.; Rosa, J.C. Evaluation of the catalytic specificity, biochemical properties, and milk clotting abilities of an aspartic peptidase from Rhizomucor miehei. J. Ind. Microbiol. Biotechnol. 2016, 43, 1059–1069. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, F.S.; Voegeli, S.; Brachat, S.; Lerch, A.; Gates, K.; Steiner, S.; Mohr, C.; Pöhlmann, R.; Luedi, P.; Choi, S. The Ashbya gossypii genome as a tool for mapping the ancient Saccharomyces cerevisiae genome. Science 2004, 304, 304–307. [Google Scholar] [CrossRef] [Green Version]
- Feldmann, H. Génolevures—A Novel Approach to ‘Evolutionary Genomics’; Elsevier: Amsterdam, The Netherlands, 2000; Volume 487, pp. 1–2. [Google Scholar]
- Kellis, M.; Birren, B.W.; Lander, E.S. Proof and evolutionary analysis of ancient genome duplication in the yeast Saccharomyces cerevisiae. Nature 2004, 428, 617–624. [Google Scholar] [CrossRef]
- Neto, Y.A.A.H.; de Souza Motta, C.M.; Cabral, H. Optimization of metalloprotease production by Eupenicillium javanicum in both solid state and submerged bioprocesses. Afr. J. Biochem. Res. 2013, 7, 146–157. [Google Scholar]
- Erez, E.; Fass, D.; Bibi, E. How intramembrane proteases bury hydrolytic reactions in the membrane. Nature 2009, 459, 371–378. [Google Scholar] [CrossRef]
- Shafee, T. Evolvability of a Viral Protease: Experimental Evolution of Catalysis, Robustness and Specificity; University of Cambridge: Cambridge, UK, 2014. [Google Scholar]
- Madhavan, A.; Arun, K.; Binod, P.; Sirohi, R.; Tarafdar, A.; Reshmy, R.; Awasthi, M.K.; Sindhu, R. Design of novel enzyme biocatalysts for industrial bioprocess: Harnessing the power of protein engineering, high throughput screening and synthetic biology. Bioresour. Technol. 2021, 325, 124617. [Google Scholar] [CrossRef]
- Veloorvalappil, N.J.; Robinson, B.S.; Selvanesan, P.; Sasidharan, S.; Kizhakkepawothail, N.U.; Sreedharan, S.; Prakasan, P.; Moolakkariyil, S.J.; Sailas, B. Versatility of microbial proteases. Adv. Enzym. Res. 2013, 2013, 36957. [Google Scholar]
- Shankar, R.; Upadhyay, P.K.; Kumar, M. Protease Enzymes: Highlights on Potential of Proteases as Therapeutics Agents. Int. J. Pept. Res. Ther. 2021, 27, 1281–1296. [Google Scholar] [CrossRef]
- Chapman, J.; Ismail, A.E.; Dinu, C.Z. Industrial applications of enzymes: Recent advances, techniques, and outlooks. Catalysts 2018, 8, 238. [Google Scholar] [CrossRef] [Green Version]
- Timson, D.J. Four challenges for better biocatalysts. Fermentation 2019, 5, 39. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, G.; Ray, A.K. Impact of microbial proteases on biotechnological industries. Biotechnol. Genet. Eng. Rev. 2017, 33, 119–143. [Google Scholar] [CrossRef] [PubMed]
- Ademosun, M.T.; Omoba, O.S.; Olagunju, A.I. Antioxidant properties, glycemic indices, and carbohydrate hydrolyzing enzymes activities of formulated ginger-based fruit drinks. J. Food Biochem. 2021, 45, e13324. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Bhalla, T.C. Microbial proteases in peptide synthesis: Approaches and applications. Appl. Microbiol. Biotechnol. 2005, 68, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Ni, X.; Yue, L.; Chi, Z.; Li, J.; Wang, X.; Madzak, C. Alkaline protease gene cloning from the marine yeast Aureobasidium pullulans HN2-3 and the protease surface display on Yarrowia lipolytica for bioactive peptide production. Mar. Biotechnol. 2009, 11, 81–89. [Google Scholar] [CrossRef]
- Kanda, S.; Aimi, T.; Kano, S.; Ishihara, S.; Kitamoto, Y.; Morinaga, T. Ambient pH signaling regulates expression of the serine protease gene (spr1) in pine wilt nematode-trapping fungus, Monacrosporium megalosporum. Microbiol. Res. 2008, 163, 63–72. [Google Scholar] [CrossRef]
- Tzean, Y.; Chou, T.-H.; Hsiao, C.-C.; Shu, P.-Y.; Walton, J.D.; Tzean, S.-S. Cloning and characterization of cuticle-degrading serine protease from nematode-trapping fungus Arthrobotrys musiformis. Mycoscience 2016, 57, 136–143. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhang, K.-Q. Biological Control of Plant-Parasitic Nematodes by Nematophagous Fungi. In Nematode-Trapping Fungi; Springer: Berlin/Heidelberg, Germany, 2014; pp. 231–262. [Google Scholar]
- Ao, X.-L.; Yu, X.; Wu, D.-T.; Li, C.; Zhang, T.; Liu, S.-L.; Chen, S.-J.; He, L.; Zhou, K.; Zou, L.-K. Purification and characterization of neutral protease from Aspergillus oryzae Y1 isolated from naturally fermented broad beans. AMB Express 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Benmrad, M.O.; Mechri, S.; Jaouadi, N.Z.; Elhoul, M.B.; Rekik, H.; Sayadi, S.; Bejar, S.; Kechaou, N.; Jaouadi, B. Purification and biochemical characterization of a novel thermostable protease from the oyster mushroom Pleurotus sajor-caju strain CTM10057 with industrial interest. BMC Biotechnol. 2019, 19, 96. [Google Scholar] [CrossRef]
- Illuri, R.; Kumar, M.; Eyini, M.; Veeramanikandan, V.; Almaary, K.S.; Elbadawi, Y.B.; Biraqdar, M.; Balaji, P. Production, partial purification and characterization of ligninolytic enzymes from selected basidiomycetes mushroom fungi. Saudi J. Biol. Sci. 2021, 28, 7207–7218. [Google Scholar] [CrossRef]
- Jaouadi, B.; Jaouadi, N.Z.; Rekik, H.; Naili, B.; Beji, A.; Dhouib, A.; Bejar, S. Biochemical and molecular characterization of Pseudomonas aeruginosa CTM50182 organic solvent-stable elastase. Int. J. Biol. Macromol. 2013, 60, 165–177. [Google Scholar] [CrossRef]
- Mandujano-González, V.; Villa-Tanaca, L.; Anducho-Reyes, M.A.; Mercado-Flores, Y. Secreted fungal aspartic proteases: A review. Rev. Iberoam. De Micol. 2016, 33, 76–82. [Google Scholar] [CrossRef]
- Rantasalo, A.; Vitikainen, M.; Paasikallio, T.; Jäntti, J.; Landowski, C.P.; Mojzita, D. Novel genetic tools that enable highly pure protein production in Trichoderma reesei. Sci. Rep. 2019, 9, 5032. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Lv, G.; Tu, Y.; Cheng, X.; Duan, Y.; Zeng, B.; He, B. Applications of CRISPR/Cas9 in the Synthesis of Secondary Metabolites in Filamentous Fungi. Front. Microbiol. 2021, 12, 164. [Google Scholar] [CrossRef]
- Liu, Q.; Gao, R.; Li, J.; Lin, L.; Zhao, J.; Sun, W.; Tian, C. Development of a genome-editing CRISPR/Cas9 system in thermophilic fungal Myceliophthora species and its application to hyper-cellulase production strain engineering. Biotechnol. Biofuels 2017, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Adalsteinsson, B.T.; Kristjansdottir, T.; Merre, W.; Helleux, A.; Dusaucy, J.; Tourigny, M.; Fridjonsson, O.; Hreggvidsson, G.O. Efficient genome editing of an extreme thermophile, Thermus thermophilus, using a thermostable Cas9 variant. Sci. Rep. 2021, 11, 9586. [Google Scholar] [CrossRef]
- Katayama, T.; Tanaka, Y.; Okabe, T.; Nakamura, H.; Fujii, W.; Kitamoto, K.; Maruyama, J.-i. Development of a genome editing technique using the CRISPR/Cas9 system in the industrial filamentous fungus Aspergillus oryzae. Biotechnol. Lett. 2016, 38, 637–642. [Google Scholar] [CrossRef]
- Jin, F.-J.; Hu, S.; Wang, B.-T.; Jin, L. Advances in genetic engineering technology and its application in the industrial fungus Aspergillus oryzae. Front. Microbiol. 2021, 12, 353. [Google Scholar] [CrossRef]
- Zou, Z.; Liu, F.; Selin, C.; Fernando, W. Generation and characterization of a virulent Leptosphaeria maculans isolate carrying a mutated AvrLm7 gene using the CRISPR/Cas9 system. Front. Microbiol. 2020, 11, 1969. [Google Scholar] [CrossRef]
- Chen, B.-X.; Wei, T.; Ye, Z.-W.; Yun, F.; Kang, L.-Z.; Tang, H.-B.; Guo, L.-Q.; Lin, J.-F. Efficient CRISPR-Cas9 gene disruption system in edible-medicinal mushroom Cordyceps militaris. Front. Microbiol. 2018, 9, 1157. [Google Scholar] [CrossRef]
- Das, S.K.; Masuda, M.; Sakurai, A.; Sakakibara, M. Medicinal uses of the mushroom Cordyceps militaris: Current state and prospects. Fitoterapia 2010, 81, 961–968. [Google Scholar] [CrossRef]
- Gardiner, D.M.; Kazan, K. Selection is required for efficient Cas9-mediated genome editing in Fusarium graminearum. Fungal Biol. 2018, 122, 131–137. [Google Scholar] [CrossRef]
- Joshi, R. A review of Fusarium oxysporum on its plant interaction and industrial use. J. Med. Plants Stud. 2018, 6, 112–115. [Google Scholar] [CrossRef]
- Schuster, M.; Schweizer, G.; Kahmann, R. Comparative analyses of secreted proteins in plant pathogenic smut fungi and related basidiomycetes. Fungal Genet. Biol. 2018, 112, 21–30. [Google Scholar] [CrossRef]
- Olicón-Hernández, D.R.; Araiza-Villanueva, M.G.; Pardo, J.P.; Aranda, E.; Guerra-Sánchez, G. New insights of Ustilago maydis as yeast model for genetic and biotechnological research: A review. Curr. Microbiol. 2019, 76, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Matsu-Ura, T.; Baek, M.; Kwon, J.; Hong, C. Efficient gene editing in Neurospora crassa with CRISPR technology. Fungal Biol. Biotechnol. 2015, 2, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, H.; Xiao, H.; Zou, G.; Zhou, Z.; Zhong, J.-J. CRISPR-Cas9 assisted gene disruption in the higher fungus Ganoderma species. Process Biochem. 2017, 56, 57–61. [Google Scholar] [CrossRef]
- Yang, H.; Wu, T.; Zhang, K. Effects of Extracts of Chinese Medicines on Ganoderma lucidum in Submerged Culture. Acta Microbiol. Sin. 2003, 43, 519–522. [Google Scholar]
- Schuster, M.; Schweizer, G.; Reissmann, S.; Kahmann, R. Genome editing in Ustilago maydis using the CRISPR–Cas system. Fungal Genet. Biol. 2016, 89, 3–9. [Google Scholar] [CrossRef]
- Al Abdallah, Q.; Souza, A.C.O.; Martin-Vicente, A.; Ge, W.; Fortwendel, J.R. Whole-genome sequencing reveals highly specific gene targeting by in vitro assembled Cas9-ribonucleoprotein complexes in Aspergillus fumigatus. Fungal Biol. Biotechnol. 2018, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Xing, H.-L.; Wang, Z.-P.; Zhang, H.-Y.; Yang, F.; Wang, X.-C.; Chen, Q.-J. Potential high-frequency off-target mutagenesis induced by CRISPR/Cas9 in Arabidopsis and its prevention. Plant Mol. Biol. 2018, 96, 445–456. [Google Scholar] [CrossRef] [Green Version]
- Doench, J.G.; Fusi, N.; Sullender, M.; Hegde, M.; Vaimberg, E.W.; Donovan, K.F.; Smith, I.; Tothova, Z.; Wilen, C.; Orchard, R. Optimized sgRNA design to maximize activity and minimize off-target effects of CRISPR-Cas9. Nat. Biotechnol. 2016, 34, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Ouedraogo, J.-P.; Kolbusz, M.; Nguyen, T.T.M.; Tsang, A. Efficient genome editing using tRNA promoter-driven CRISPR/Cas9 gRNA in Aspergillus niger. PLoS ONE 2018, 13, e0202868. [Google Scholar] [CrossRef]
- Huang, L.; Dong, H.; Zheng, J.; Wang, B.; Pan, L. Highly efficient single base editing in Aspergillus niger with CRISPR/Cas9 cytidine deaminase fusion. Microbiol. Res. 2019, 223, 44–50. [Google Scholar] [CrossRef]
- Sharma, K.M.; Kumar, R.; Panwar, S.; Kumar, A. Microbial alkaline proteases: Optimization of production parameters and their properties. J. Genet. Eng. Biotechnol. 2017, 15, 115–126. [Google Scholar] [CrossRef]
- Ward, O.P. Production of recombinant proteins by filamentous fungi. Biotechnol. Adv. 2012, 30, 1119–1139. [Google Scholar] [CrossRef]
- Ravanelli, S.; den Brave, F.; Hoppe, T. Mitochondrial quality control governed by ubiquitin. Front. Cell Dev. Biol. 2020, 8, 270. [Google Scholar] [CrossRef]
- Jadiya, P.; Tomar, D. Mitochondrial protein quality control mechanisms. Genes 2020, 11, 563. [Google Scholar] [CrossRef]
- Deshwal, S.; Fiedler, K.U.; Langer, T. Mitochondrial proteases: Multifaceted regulators of mitochondrial plasticity. Annu. Rev. Biochem. 2020, 89, 501–528. [Google Scholar] [CrossRef] [Green Version]
- Quiros, P.M.; Langer, T.; Lopez-Otin, C. New roles for mitochondrial proteases in health, ageing and disease. Nat. Rev. Mol. Cell Biol. 2015, 16, 345–359. [Google Scholar] [CrossRef]
- Hofsetz, E.; Huesgen, P.F.; Trifunovic, A. Identification of Putative Mitochondrial Protease Substrates. In Mitochondrial Gene Expression; Springer: Berlin/Heidelberg, Germany, 2021; pp. 313–329. [Google Scholar]
- Voos, W. Chaperone–protease networks in mitochondrial protein homeostasis. Biochim. Et Biophys. Acta (BBA) Mol. Cell Res. 2013, 1833, 388–399. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, P.F.; Glaser, E. Processing peptidases in mitochondria and chloroplasts. Biochim. Et Biophys. Acta (BBA) Mol. Cell Res. 2013, 1833, 360–370. [Google Scholar] [CrossRef] [Green Version]
- Weiss-Sadan, T.; Gotsman, I.; Blum, G. Cysteine proteases in atherosclerosis. FEBS J. 2017, 284, 1455–1472. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Sukhova, G.K.; Yang, J.-T.; Sun, J.; Ma, L.; Ren, A.; Xu, W.-H.; Fu, H.; Dolganov, G.M.; Hu, C. Cathepsin L expression and regulation in human abdominal aortic aneurysm, atherosclerosis, and vascular cells. Atherosclerosis 2006, 184, 302–311. [Google Scholar] [CrossRef]
- Jaffer, F.A.; Vinegoni, C.; John, M.C.; Aikawa, E.; Gold, H.K.; Finn, A.V.; Ntziachristos, V.; Libby, P.; Weissleder, R. Real-time catheter molecular sensing of inflammation in proteolytically active atherosclerosis. Circulation 2008, 118, 1802–1809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weitoft, T.; Larsson, A.; Manivel, V.A.; Lysholm, J.; Knight, A.; Rönnelid, J. Cathepsin S and cathepsin L in serum and synovial fluid in rheumatoid arthritis with and without autoantibodies. Rheumatology 2015, 54, 1923–1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appelqvist, H.; Wäster, P.; Kågedal, K.; Öllinger, K. The lysosome: From waste bag to potential therapeutic target. J. Mol. Cell Biol. 2013, 5, 214–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
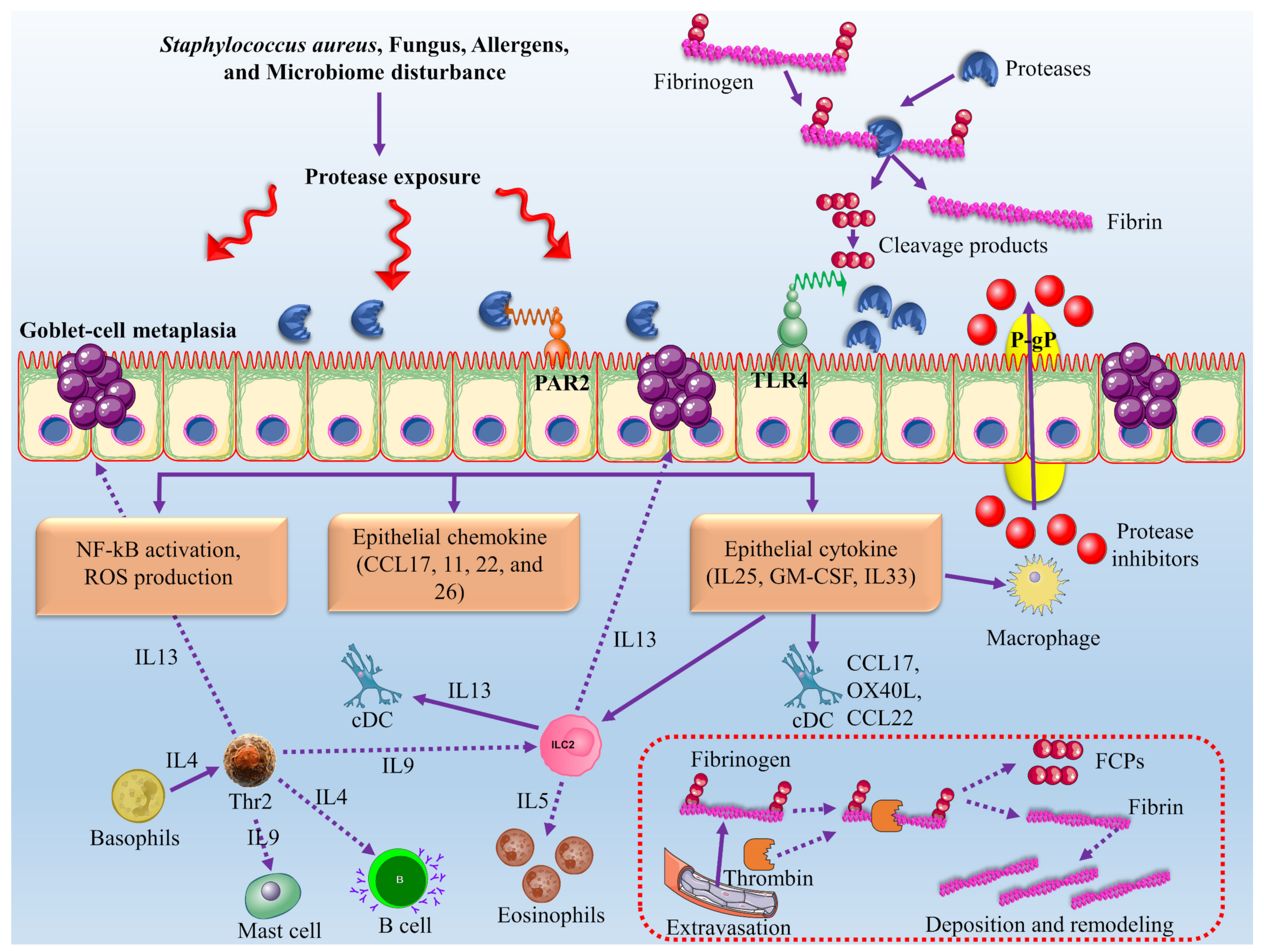
- Wu, D.; Wei, Y.; Bleier, B.S. Emerging role of proteases in the pathogenesis of chronic rhinosinusitis with nasal polyps. Front. Cell. Infect. Microbiol. 2018, 7, 538. [Google Scholar] [CrossRef] [Green Version]
- Stentzel, S.; Teufelberger, A.; Nordengrün, M.; Kolata, J.; Schmidt, F.; Van Crombruggen, K.; Michalik, S.; Kumpfmüller, J.; Tischer, S.; Schweder, T. Staphylococcal serine protease–like proteins are pacemakers of allergic airway reactions to Staphylococcus aureus. J. Allergy Clin. Immunol. 2017, 139, 492–500.e8. [Google Scholar] [CrossRef] [Green Version]
- Kale, S.L.; Agrawal, K.; Gaur, S.N.; Arora, N. Cockroach protease allergen induces allergic airway inflammation via epithelial cell activation. Sci. Rep. 2017, 7, 42341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teufelberger, A.R.; Nordengrün, M.; Braun, H.; Maes, T.; De Grove, K.; Holtappels, G.; O’Brien, C.; Provoost, S.; Hammad, H.; Gonçalves, A. The IL-33/ST2 axis is crucial in type 2 airway responses induced by Staphylococcus aureus–derived serine protease–like protein D. J. Allergy Clin. Immunol. 2018, 141, 549–559.e7. [Google Scholar] [CrossRef] [Green Version]
- López-Otín, C.; Matrisian, L.M. Emerging roles of proteases in tumour suppression. Nat. Rev. Cancer 2007, 7, 800–808. [Google Scholar] [CrossRef]
- Kwon, Y.T.; Ciechanover, A. The ubiquitin code in the ubiquitin-proteasome system and autophagy. Trends Biochem. Sci. 2017, 42, 873–886. [Google Scholar] [CrossRef]
- Liu, J.; Shaik, S.; Dai, X.; Wu, Q.; Zhou, X.; Wang, Z.; Wei, W. Targeting the ubiquitin pathway for cancer treatment. Biochim. Et Biophys. Acta (BBA)-Rev. Cancer 2015, 1855, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Green, P.H.; Lebwohl, B.; Greywoode, R. Celiac disease. J. Allergy Clin. Immunol. 2015, 135, 1099–1106. [Google Scholar] [CrossRef]
- Makharia, G.K.D. Current and emerging therapy for celiac disease. Front. Med. 2014, 1, 6. [Google Scholar] [CrossRef] [Green Version]
- Caruso, J.A.; Akli, S.; Pageon, L.; Hunt, K.K.; Keyomarsi, K. The serine protease inhibitor elafin maintains normal growth control by opposing the mitogenic effects of neutrophil elastase. Oncogene 2015, 34, 3556–3567. [Google Scholar] [CrossRef] [Green Version]
- Galipeau, H.J.; Wiepjes, M.; Motta, J.-P.; Schulz, J.D.; Jury, J.; Natividad, J.M.; Pinto-Sanchez, I.; Sinclair, D.; Rousset, P.; Martin-Rosique, R. Novel role of the serine protease inhibitor elafin in gluten-related disorders. Am. J. Gastroenterol. 2014, 109, 748. [Google Scholar] [CrossRef] [Green Version]
- Ghetti, B.; Tagliavini, F.; Kovacs, G.G.; Piccardo, P. 37 Gerstmann–Str ä ussler–Scheinker Disease. In Neurodegeneration: The Molecular Pathology of Dementia and Movement Disorders; Wiley: Hoboken, NJ, USA, 2011; p. 364. [Google Scholar]
- Iwasaki, Y. Creutzfeldt-Jakob disease. Neuropathology 2017, 37, 174–188. [Google Scholar] [CrossRef]
- Yoshioka, M.; Miwa, T.; Horii, H.; Takata, M.; Yokoyama, T.; Nishizawa, K.; Watanabe, M.; Shinagawa, M.; Murayama, Y. Characterization of a proteolytic enzyme derived from a Bacillus strain that effectively degrades prion protein. J. Appl. Microbiol. 2007, 102, 509–515. [Google Scholar] [CrossRef]
- Rajput, R.; Gupta, R. Thermostable keratinase from Bacillus pumilus KS12: Production, chitin crosslinking and degradation of Sup35NM aggregates. Bioresour. Technol. 2013, 133, 118–126. [Google Scholar] [CrossRef]
- Chauhan, B.; Gupta, R. Application of statistical experimental design for optimization of alkaline protease production from Bacillus sp. RGR-14. Process Biochem. 2004, 39, 2115–2122. [Google Scholar] [CrossRef]
- Reddy, C.C.; Khilji, I.A.; Gupta, A.; Bhuyar, P.; Mahmood, S.; AL-Japairai, K.A.S.; Chua, G.K. Valorization of keratin waste biomass and its potential applications. J. Water Process Eng. 2021, 40, 101707. [Google Scholar] [CrossRef]
- Matkawala, F.; Nighojkar, S.; Kumar, A.; Nighojkar, A. Microbial alkaline serine proteases: Production, properties and applications. World J. Microbiol. Biotechnol. 2021, 37, 63. [Google Scholar] [CrossRef]
- Chilakamarry, C.R.; Mahmood, S.; Saffe, S.N.B.M.; Arifin, M.A.B.; Gupta, A.; Sikkandar, M.Y.; Begum, S.S.; Narasaiah, B. Extraction and application of keratin from natural resources: A review. 3 Biotech 2021, 11, 220. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Wilkens, C.; Barrett, K.; Meyer, A.S. Microbial enzymes catalyzing keratin degradation: Classification, structure, function. Biotechnol. Adv. 2020, 44, 107607. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, S.; Poudel, D.K.; Marahatha, R.; Dawadi, S.; Khadayat, K.; Phuyal, S.; Shrestha, S.; Gaire, S.; Basnet, K.; Khadka, U. Microbial Enzymes Used in Bioremediation. J. Chem. 2021, 2021, 8849512. [Google Scholar] [CrossRef]
- Saranya, P.; Selvi, P.; Sekaran, G. Integrated thermophilic enzyme-immobilized reactor and high-rate biological reactors for treatment of palm oil-containing wastewater without sludge production. Bioprocess Biosyst. Eng. 2019, 42, 1053–1064. [Google Scholar] [CrossRef]
- Gradisar, H.; Friedrich, J.; Krizaj, I.; Jerala, R. Similarities and specificities of fungal keratinolytic proteases: Comparison of keratinases of Paecilomyces marquandii and Doratomyces microsporus to some known proteases. Appl. Environ. Microbiol. 2005, 71, 3420–3426. [Google Scholar]
- Espersen, R.; Huang, Y.; Falco, F.C.; Hägglund, P.; Gernaey, K.V.; Lange, L.; Svensson, B. Exceptionally rich keratinolytic enzyme profile found in the rare actinomycetes Amycolatopsis keratiniphila D2T. Appl. Microbiol. Biotechnol. 2021, 105, 8129–8138. [Google Scholar] [CrossRef]
- Barzkar, N.; Sohail, M.; Jahromi, S.T.; Nahavandi, R.; Khodadadi, M. Marine microbial L-glutaminase: From pharmaceutical to food industry. Appl. Microbiol. Biotechnol. 2021, 105, 4453–4466. [Google Scholar] [CrossRef]
- Bond, J.S. Proteases: History, discovery, and roles in health and disease. J. Biol. Chem. 2019, 294, 1643–1651. [Google Scholar] [CrossRef] [Green Version]
- Rudzińska, M.; Daglioglu, C.; Savvateeva, L.V.; Kaci, F.N.; Antoine, R.; Zamyatnin, A.A., Jr. Current Status and Perspectives of Protease Inhibitors and Their Combination with Nanosized Drug Delivery Systems for Targeted Cancer Therapy. Drug Des. Dev. Ther. 2021, 15, 9. [Google Scholar] [CrossRef]
- Awad, M.F.; El-Shenawy, F.S.; El-Gendy, M.M.A.A.; El-Bondkly, E.A.M. Purification, characterization, and anticancer and antioxidant activities of l-glutaminase from Aspergillus versicolor Faesay4. Int. Microbiol. 2021, 24, 169–181. [Google Scholar] [CrossRef]
- Qamar, S.A.; Asgher, M.; Bilal, M. Immobilization of alkaline protease from Bacillus brevis using Ca-alginate entrapment strategy for improved catalytic stability, silver recovery, and dehairing potentialities. Catal. Lett. 2020, 150, 3572–3583. [Google Scholar] [CrossRef]
- Bholay, A.; More, S.; Patil, V.; Niranjan, P. Bacterial extracellular alkaline proteases and its industrial applications. Int. Res. J. Biol. Sci. 2012, 1, 1–5. [Google Scholar]
- Abad, P.; Gouzy, J.; Aury, J.-M.; Castagnone-Sereno, P.; Danchin, E.G.; Deleury, E.; Perfus-Barbeoch, L.; Anthouard, V.; Artiguenave, F.; Blok, V.C. Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita. Nat. Biotechnol. 2008, 26, 909–915. [Google Scholar] [CrossRef] [Green Version]
- Nakpathom, M.; Somboon, B.; Narumol, N. Papain enzymatic degumming of Thai Bombyx mori silk fibers. J. Microsc. Soc. Thail. 2009, 23, 142–146. [Google Scholar]
- Miao, Y.; Chen, G.; Xi, X.; Ma, C.; Wang, L.; Burrows, J.F.; Duan, J.; Zhou, M.; Chen, T. Discovery and rational design of a novel bowman-birk related protease inhibitor. Biomolecules 2019, 9, 280. [Google Scholar] [CrossRef] [Green Version]
- Zhai, P.-b.; Qing, J.; Li, B.; Zhang, L.-q.; Ma, L.; Chen, L. GP205, a new hepatitis C virus NS3/4A protease inhibitor, displays higher metabolic stability in vitro and drug exposure in vivo. Acta Pharmacol. Sin. 2018, 39, 1746–1752. [Google Scholar] [CrossRef]
- Goris, T.; Pérez-Valero, Á.; Martínez, I.; Yi, D.; Fernández-Calleja, L.; San Leon, D.; Bornscheuer, U.T.; Magadán-Corpas, P.; Lombo, F.; Nogales, J. Repositioning microbial biotechnology against COVID-19: The case of microbial production of flavonoids. Microb. Biotechnol. 2021, 14, 94–110. [Google Scholar] [CrossRef]
- Cotabarren, J.; Lufrano, D.; Parisi, M.G.; Obregón, W.D. Biotechnological, biomedical, and agronomical applications of plant protease inhibitors with high stability: A systematic review. Plant Sci. 2020, 292, 110398. [Google Scholar] [CrossRef]
- Rawlings, N.D. Peptidase inhibitors in the MEROPS database. Biochimie 2010, 92, 1463–1483. [Google Scholar] [CrossRef]
- Garg, H.; Suri, P.; Gupta, J.C.; Talwar, G.; Dubey, S. Survivin: A unique target for tumor therapy. Cancer Cell Int. 2016, 16, 49. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, N.D.; Barrett, A.J.; Finn, R. Twenty years of the MEROPS database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Res. 2016, 44, D343–D350. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez-Gongora, D.; Geddes-McAlister, J. From Naturally-Sourced Protease Inhibitors to New Treatments for Fungal Infections. J. Fungi 2021, 7, 1016. [Google Scholar] [CrossRef]
- Kantyka, T.; Rawlings, N.D.; Potempa, J. Prokaryote-derived protein inhibitors of peptidases: A sketchy occurrence and mostly unknown function. Biochimie 2010, 92, 1644–1656. [Google Scholar] [CrossRef] [Green Version]
- Greenbaum, L.M.; Sutherland, J.H. Host cathepsin D response to tumor in the normal and pepstatin-treated mouse. Cancer Res. 1983, 43, 2584–2587. [Google Scholar]
- Kalchev, K.; Rabadjiev, Y.; Ganchev, D.; Tsenova, M.; Iliev, I.; Ivanova, I. Study of proteases and protease inhibitors from Streptomyces strains. Bulg. J. Agric. Sci. 2013, 19, 65–67. [Google Scholar]
- Manojlovic, N.T.; Solujic, S.; Sukdolak, S.; Milosev, M. Antifungal activity of Rubia tinctorum, Rhamnus frangula and Caloplaca cerina. Fitoterapia 2005, 76, 244–246. [Google Scholar] [CrossRef]
- Menéndez-Arias, L.; Tözsér, J. HIV-1 protease inhibitors: Effects on HIV-2 replication and resistance. Trends Pharmacol. Sci. 2008, 29, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Chandwani, A.; Shuter, J. Lopinavir/ritonavir in the treatment of HIV-1 infection: A review. Ther. Clin. Risk Manag. 2008, 4, 1023. [Google Scholar]
- Fear, G.; Komarnytsky, S.; Raskin, I. Protease inhibitors and their peptidomimetic derivatives as potential drugs. Pharmacol. Ther. 2007, 113, 354–368. [Google Scholar] [CrossRef]
- Purohit, R.; Rajendran, V.; Sethumadhavan, R. Studies on adaptability of binding residues flap region of TMC-114 resistance HIV-1 protease mutants. J. Biomol. Struct. Dyn. 2011, 29, 137–152. [Google Scholar] [CrossRef]
- van Maarseveen, N.M.; Andersson, D.; Lepšík, M.; Fun, A.; Schipper, P.J.; de Jong, D.; Boucher, C.A.; Nijhuis, M. Modulation of HIV-1 Gag NC/p1 cleavage efficiency affects protease inhibitor resistance and viral replicative capacity. Retrovirology 2012, 9, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu-Young, G.; Kozal, M.J. Hepatitis C protease and polymerase inhibitors in development. AIDS Patient Care STDs 2008, 22, 449–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onoda, T.; Ono, T.; Dhar, D.K.; Yamanoi, A.; Nagasue, N. Tetracycline analogues (doxycycline and COL-3) induce caspase-dependent and-independent apoptosis in human colon cancer cells. Int. J. Cancer 2006, 118, 1309–1315. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S.S.; Shepherd, F.A. Targeting angiogenesis: A review of angiogenesis inhibitors in the treatment of lung cancer. Lung Cancer 2003, 42, 81–91. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Tolle, D.P.; Barrett, A.J. Evolutionary families of peptidase inhibitors. Biochem. J. 2004, 378, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Clemente, M.; Corigliano, M.G.; Pariani, S.A.; Sánchez-López, E.F.; Sander, V.A.; Ramos-Duarte, V.A. Plant serine protease inhibitors: Biotechnology application in agriculture and molecular farming. Int. J. Mol. Sci. 2019, 20, 1345. [Google Scholar] [CrossRef] [Green Version]
- Peele, K.A.; Durthi, C.P.; Srihansa, T.; Krupanidhi, S.; Ayyagari, V.S.; Babu, D.J.; Indira, M.; Reddy, A.R.; Venkateswarulu, T. Molecular docking and dynamic simulations for antiviral compounds against SARS-CoV-2: A computational study. Inform. Med. Unlocked 2020, 19, 100345. [Google Scholar] [CrossRef]
- Macchiagodena, M.; Pagliai, M.; Procacci, P. Identification of potential binders of the main protease 3CLpro of the COVID-19 via structure-based ligand design and molecular modeling. Chem. Phys. Lett. 2020, 750, 137489. [Google Scholar] [CrossRef]
- Rut, W.; Groborz, K.; Zhang, L.; Sun, X.; Zmudzinski, M.; Pawlik, B.; Wang, X.; Jochmans, D.; Neyts, J.; Młynarski, W. SARS-CoV-2 M pro inhibitors and activity-based probes for patient-sample imaging. Nat. Chem. Biol. 2021, 17, 222–228. [Google Scholar] [CrossRef]
- Drag, M.; Salvesen, G.S. Emerging principles in protease-based drug discovery. Nat. Rev. Drug Discov. 2010, 9, 690–701. [Google Scholar] [CrossRef] [Green Version]
- Amin, S.A.; Banerjee, S.; Ghosh, K.; Gayen, S.; Jha, T. Protease targeted COVID-19 drug discovery and its challenges: Insight into viral main protease (Mpro) and papain-like protease (PLpro) inhibitors. Bioorganic Med. Chem. 2020, 29, 115860. [Google Scholar] [CrossRef]
- Lee, T.-W.; Cherney, M.M.; Huitema, C.; Liu, J.; James, K.E.; Powers, J.C.; Eltis, L.D.; James, M.N. Crystal structures of the main peptidase from the SARS coronavirus inhibited by a substrate-like aza-peptide epoxide. J. Mol. Biol. 2005, 353, 1137–1151. [Google Scholar] [CrossRef]
- Shen, S.; Wu, Y.; Liu, Y.; Wu, D. High drug-loading nanomedicines: Progress, current status, and prospects. Int. J. Nanomed. 2017, 12, 4085. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.; Du, X.-J.; Liu, J.; Sun, R.; Zhu, Y.-H.; Wang, J. Delivery of bortezomib with nanoparticles for basal-like triple-negative breast cancer therapy. J. Control. Release 2015, 208, 14–24. [Google Scholar] [CrossRef]
- Park, J.E.; Park, J.; Jun, Y.; Oh, Y.; Ryoo, G.; Jeong, Y.-S.; Gadalla, H.H.; Min, J.S.; Jo, J.H.; Song, M.G. Expanding therapeutic utility of carfilzomib for breast cancer therapy by novel albumin-coated nanocrystal formulation. J. Control. Release 2019, 302, 148–159. [Google Scholar] [CrossRef]
- Gotou, T.; Shinoda, T.; Mizuno, S.; Yamamoto, N. Purification and identification of proteolytic enzymes from Aspergillus oryzae capable of producing the antihypertensive peptide Ile-Pro-Pro. J. Biosci. Bioeng. 2009, 107, 615–619. [Google Scholar] [CrossRef]
- Song, R.; Qiao, W.; He, J.; Huang, J.; Luo, Y.; Yang, T. Proteases and Their Modulators in Cancer Therapy: Challenges and Opportunities. J. Med. Chem. 2021, 64, 2851–2877. [Google Scholar] [CrossRef]
- Cavaco, M.; Andreu, D.; Castanho, M.A. The challenge of peptide proteolytic stability studies: Scarce data, difficult readability, and the need for harmonization. Angew. Chem. Int. Ed. 2021, 60, 1686–1688. [Google Scholar] [CrossRef]
- Verma, S.; Goand, U.K.; Husain, A.; Katekar, R.A.; Garg, R.; Gayen, J.R. Challenges of peptide and protein drug delivery by oral route: Current strategies to improve the bioavailability. Drug Dev. Res. 2021, 82, 927–944. [Google Scholar] [CrossRef]
- Sharma, A.; Gupta, G.; Ahmad, T.; Mansoor, S.; Kaur, B. Enzyme engineering: Current trends and future perspectives. Food Rev. Int. 2021, 37, 121–154. [Google Scholar] [CrossRef]
- Nyika, J.M. The Use of Microorganism-Derived Enzymes for Bioremediation of Soil Pollutants. In Recent Advancements in Bioremediation of Metal Contaminants; IGI Global: Hershey/Pennsylvania, PA, USA, 2021; pp. 54–71. [Google Scholar]
- Saravanan, A.; Kumar, P.S.; Vo, D.-V.N.; Jeevanantham, S.; Karishma, S.; Yaashikaa, P. A review on catalytic-enzyme degradation of toxic environmental pollutants: Microbial enzymes. J. Hazard. Mater. 2021, 419, 126451. [Google Scholar] [CrossRef]
- Li, Q.; Yi, L.; Marek, P.; Iverson, B.L. Commercial proteases: Present and future. FEBS Lett. 2013, 587, 1155–1163. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Shen, H.; Shao, Y.; Ma, Q.; Niu, Y.; Shang, Z. A narrative review of proteolytic targeting chimeras (PROTACs): Future perspective for prostate cancer therapy. Transl. Androl. Urol. 2021, 10, 954. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme Class | Type of Strain | Production Method | Mol. Weight (kDa) | Opt. pH | Opt. Temperature (℃) | Substrate | Inhibitors | Enzyme Activity (%) | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Fungal amylase | Thermomyces lanuginosus | SSF | 33 | 5 | 48 | Wheat straw, guayule bagasse | SDS | 88 | [37,38] |
| Aspergillus fumigatus | SmF | 28 | 4.5 | 30 | Pomegranate peel, wheat bran | Ebselen | 90 | [39,40] | |
| Cryptococcus flavus | SSF | 70 | 5 | 45 | Starch, amylose | Mercury | 80–84 | [41] | |
| Aspergillus niger | SSF | 115 | 5 | 75 | Cowpeas, chickpeas | 88 | [42,43] | ||
| Mucor sp. | SSF | 38 | 4 | 30 | Kidney beans, lupine | EDTA | 84–86 | [44,45] | |
| Aspergillus oryzae | SSF | 102 | 8 | 30 | Groundnut oil, sesame oil | Copper | 85 | [46,47] | |
| Aspergillus kawachii | SSF | 110 | 3 | 32 | Pearl millet | Mercury | 88–90 | [48,49] | |
| Penicillium fterreus | SSF | 25 | 6 | 28 | Cowpeas, chickpeas | Lead | 78 | [43,50] | |
| Fungal cellulase | Trichoderma viride | SSF | 55 | 7 | 52 | CMC | Mercury | 86–88 | [51,52] |
| Peniophora sp. | SmF | 30 | 4 | 58 | SKT | EDTA | 80 | [53] | |
| Aspergillusniger IMMIS1 | SSF | 70 | 3.5 | 32 | RW, bread | Mercury | 90 | [54,55] | |
| T. harzianum | SSF | 40 | 5 | 70 | Sugarcane bagasse | EDTA | 85 | [13,56] | |
| Aspergillus niger VTCC-F021 | SSF | 29 | 4 | 52 | Sugar cane, CMC | Zinc | 95 | [57,58] | |
| Aspergillus terreus | SmF | 52 | 3.5 | 13 | Cowpeas, chickpeas | Mercury | 78 | [59] | |
| Fungal lipase | Aspergiillus niger | SSF | 30 | 3 | 40–48 | Coir waste, RH | Zinc | 88 | [42,60] |
| A. terreus | SmF | 35 | 3.5 | 48 | MOC | α-Glucosidase | 85 | [59,61] | |
| A. versicolor | SmF | 93 | 7 | 60 | EMO | Lipstatin | 90 | [62,63] | |
| A. tamarii | SSF | 5 | 4.5 | 55 | GOC, agrowastes | Ebelactone | 92 | [64,65] | |
| A. japonicus | SmF | 9 | 4.6 | 25 | SFO, casein | Caulerpenyne | 88 | [66,67] | |
| Mucor sp. | SmF | 7 | 4.2 | 28 | Kidney beans, lupine | Percyquinin | 66 | [45,68] | |
| Fungal protease | Scopulariopsis sp. | SSF | 38 | 8 | 56 | Glucose, peptone | NBS | 88 | [69,70] |
| Aspergillus niger | SSF | 47 | 7 | 45 | Cowpeas, chickpeas | EDTA | 88 | [42,71] | |
| Aspergillus fumigatus | SSF | 40 | 8 | 31 | PL, casein | DTT | 90 | [72,73] | |
| Rhizopus oryza | SmF | 55 | 5 | 32 | WBW | DTT | 75 | [74,75] | |
| Mucpr Sp. | SSF | 35 | 7 | 28 | Kidney beans, lupine | EDTA | 88 | [45,76] | |
| G. putredinis | SmF | 48 | 7–8 | 29 | Soya bean meal | IAA | 85 | [13] | |
| T. harzianum | SSF | 45 | 7 | 19 | Glutamine | PMSF | 80–82 | [13] |
| Fungal Strain | Nature | Genus | Opt. Temperature (°C) | Fungal Amylase | Fungal Cellulase | Fungal Lipase | Fungal Protease | Application | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Thermomyces lanuginosus | Thermophilic fungus | Thermomyces | 40–50 | √ | ✕ | √ | √ | Wastewater and pharmaceuticals waste treatment | [81,82] |
| Sporotrichum thermophile | Thermophilic fungus | Sporotrichum | 45–50 | ✕ | √ | ✕ | √ | Biomass degradation | [86,87] |
| Myceliophthora thermophila | Thermophilic fungus | Myceliophthora | 45–50 | ✕ | √ | ✕ | √ | Textile industries and bioremediation | [84] |
| Thermomyces ibadanensis | Thermophilic fungus | Thermomyces | 46–55 | ✕ | ✕ | √ | √ | Wastewater treatment | [85] |
| Neurospora crassa | Mesophilic fungus | Neurospora | 20–30 | ✕ | √ | ✕ | √ | As a model organism in an analysis of genetic recombination | [83] |
| Aspergillus niger | Mesophilic fungus | Aspergillus | 20–30 | √ | ✕ | ✕ | √ | Food industries | [88] |
| Aspergillus flavus | Mesophilic fungus | Aspergillus | 25–30 | √ | √ | ✕ | √ | Textile, detergent, and paper industries | [88] |
| Candida mogii | Psychrophilic fungus | Candida | 5–10 | √ | ✕ | √ | √ | Food industries | [89] |
| Enzyme Isolated | Enzyme Class | Active Site Residue (s) | Isolated Source | Reference |
|---|---|---|---|---|
| Clostripain, Streptopain | Cysteine proteases | Cysteine and histidine residues | C. histolyticum, S. griseus | [98,99] |
| Pepsins, proteases, rennet like proteases | Aspartic endoproteases | Two aspartate residues | A. niger, M. miehei | [100,101] |
| Chymotrypsins, subtilisins | Serine proteases | Serine residues | B. sphaericus | [102,103] |
| Collagenases, elastase | Metalloendoproteases | Metal ions | C. histolyticum, P. aeruginosa | [104,105] |
| Eqolisin protease | Glutamic proteases | Glutamate residues | S. lignicola, A. niger | [106,107] |
| Pepsins (A1), retropepsin (A2) | Acidic proteases | - | A. niger, A. saitoi. | [108,109] |
| Subtilisin, carlsberg | Alkaline proteases | - | A. salinivibrio, C. aureus | [109] |
| Neutrase, thermolysin | Neutral proteases | - | Bacillus sp. | [109] |
| Fungal Strain | Isolated Proteases | CRISPR System | Selective Marker | Promoter for sgRNA | Promoter for Cas9 | Delivery Method | Editing Method | Application | Efficiency (%) | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| A. oryzae | Aspartic acid protease | Cas-9 | pyrG | U6 | amyB | PMT | NHEJ | Genetic engineering, food industries | 10–30 | [158,159] |
| M. thermophila | Alkaline protease | Cas-9 | bar | U6 | tef1 | AMT | HDR | pharmaceuticals biomass/waste treatment | 20–95 | [86,156] |
| T. lanuginosus | Serine alkaline protease | Cas-9 | hph | U6 | act1 | AMT | NHEJ | Wastewater and pharmaceuticals waste treatment | 44–90 | [82,160] |
| C. militaris | Serine alkaline protease | Cas-9 | 5-FOA/blpR | U6 | tef1 | AMT | NHEJ | Pharmacecurials | 82–89 | [161,162] |
| F. graminearum | Acid proteases | Cas-9 | Fludioxonil | U6 | gpdA | AMT | HDR | Food indsuries, pests conrol | 2–12 | [163,164] |
| U. maydis | Metalloproteases | Cas-9 | Ip | U6 | otef | PMT | NHEJ | Biofuels, pharmacuetcals | 50–90 | [165,166] |
| N. crassa | Serine proteases | Cas-9 | bar | SNR52 | TrpC | AMT | HDR | Genetic recombination | 60–80 | [83,167] |
| G. lucidum | Metalloproteases | Cas-9 | ura3 | T7 | gpdA | PMT | NHEJ | Medicine | 28–33 | [168,169] |
| Type of Fungal Protease Inhibitors | Proteases Inhibited | Family | Activity | Inhibitory Mechanism | Application | Reference |
|---|---|---|---|---|---|---|
| Survivin (Cysteine inhibitor) | Caspases- specific cysteine proteases | C14 | Antifungal | Tight binding | Role as a mitotic regulator of cell division and as an inhibitor of caspase activation in the process of apoptosis. | [226,227] |
| Diosgenin (Metalloprotease inhibitor) | Metalloproteases | M15 | Antifungal | Tight binding | These inhibit the secreted metallopeptidase relevant in brain invasion by cryptococcal cells, causing meningoencephalitis. | [228,229] |
| Serpin (Serine inhibitor) | Serine Proteases | C1 and C14 | Antifungal | Trapping traps the serine protease in a covalent complex | Fungal serpins as a therapeutic benefit toward several inflammation-related complications. | [230] |
| Saccharo (Pepsin, aspartic acid inhibitor) (IA3) | Aspartic acid proteases | C1 | Antifungal | Tight binding | Role as an inhibitor in the process of apoptosis and cancer. | [231] |
| Streptomyces (Metallopeptidase inhibitor) | Metalloproteases | C14 | Antifungal | Tight binding | Role as an inhibitor in tumor invasion and metastasis (the most validated target for cancer). | [228,232] |
| RflP-1 (Rhamnus frangula inhibitor proteases) | Serine protease | C14 | Antifungal | Trapping | Role in free radical scavenging activities. | [233] |
| Name of Inhibitor | Targeted Enzyme | Target Disease | Clinical Trials Stage | Reference |
|---|---|---|---|---|
| RO033-4649 | Therapeutic agents have reduced the rate of mortality and are helpful for treatment | AIDS | Under the clinical phase I | [238] |
| VX-950 | Become the most common cause of liver cirrhosis | HCV | Under the clinical trial phase II | [239] |
| COL-3 | Therapeutic agents to control the different mutations in colon cancer | Colon cancer | Entered the phase II stage | [240] |
| AG3340 | Therapeutic agents to control the different mutations in lung cancer | Lung cancer | Entered the phase II stage | [241] |
| TMC-114 | Therapeutic agents have reduced the rate of mortality and are helpful for the treatment | AIDS | Under clinical phase III | [237] |
| Indinavir | These inhibitors bind to the HIV and inhibit the viral replication | HIV | FDA has approved this inhibitor | [234,235] |
| Ritonavir | These inhibitors bind to the HIV and inhibit the viral replication | HIV | FDA has approved this inhibitor | [234,235] |
| GP205 | GP205 inhibitor showed the biological activities in targeting the HCV virus, and ultimately, this novel inhibitor could be for possible treatment for Hepatitis C virus | HCV | GP205 inhibitor showed the biological activities in targeting the HCV virus, and ultimately, this novel inhibitor could be for possible treatment for Hepatitis C virus | [223] |
| Quercetin 3-β-d-glucoside and helichrysetin | The biochemical analysis of these compounds showed biological activities in the suppression of the MERS-COV 3Cl protease | Coronaviruses | These chemical compounds could be used as a possible treatment in targeting the coronaviruses | [224] |
| Mpro inhibitors | These inhibitors play a role in processing the replicase during the post-translational process’s viruses of the Coronaviridae family | Coronaviruses | These proteases can be used for antiviral drug and discovery | [244,245,246] |
| PLpro inhibitors | These inhibitors play a vital role in transcription by the processing of the two polyproteins, pp1a and pp1ab | Coronaviruses | These proteases also can be used for the discovery of novel protease inhibitors | [244,245,246] |
| NP-delivery systems-based carfilzomib and bortezomib | This nanotechnology-based approach could help reduce the side effects of drugs | These protease inhibitors can be designed with combinations with gold, PEGlycated, silica, liposomes and demonstrate the high efficacy rate | Future discovery of novel inhibitors based on modified NP-systems protecting the normal tissues and improving the quality of patients | [252] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naeem, M.; Manzoor, S.; Abid, M.-U.-H.; Tareen, M.B.K.; Asad, M.; Mushtaq, S.; Ehsan, N.; Amna, D.; Xu, B.; Hazafa, A. Fungal Proteases as Emerging Biocatalysts to Meet the Current Challenges and Recent Developments in Biomedical Therapies: An Updated Review. J. Fungi 2022, 8, 109. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8020109
Naeem M, Manzoor S, Abid M-U-H, Tareen MBK, Asad M, Mushtaq S, Ehsan N, Amna D, Xu B, Hazafa A. Fungal Proteases as Emerging Biocatalysts to Meet the Current Challenges and Recent Developments in Biomedical Therapies: An Updated Review. Journal of Fungi. 2022; 8(2):109. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8020109
Chicago/Turabian StyleNaeem, Muhammad, Saba Manzoor, Mashhud-Ul-Hasan Abid, Muhammad Burhan Khan Tareen, Mirza Asad, Sajida Mushtaq, Nazia Ehsan, Dua Amna, Baojun Xu, and Abu Hazafa. 2022. "Fungal Proteases as Emerging Biocatalysts to Meet the Current Challenges and Recent Developments in Biomedical Therapies: An Updated Review" Journal of Fungi 8, no. 2: 109. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8020109