
Exploring the Relationships between Four New Species of Boletoid Fungi from Northern China and Their Related Species


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samplings and Morphological Analyses
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Data Analysis
3. Results
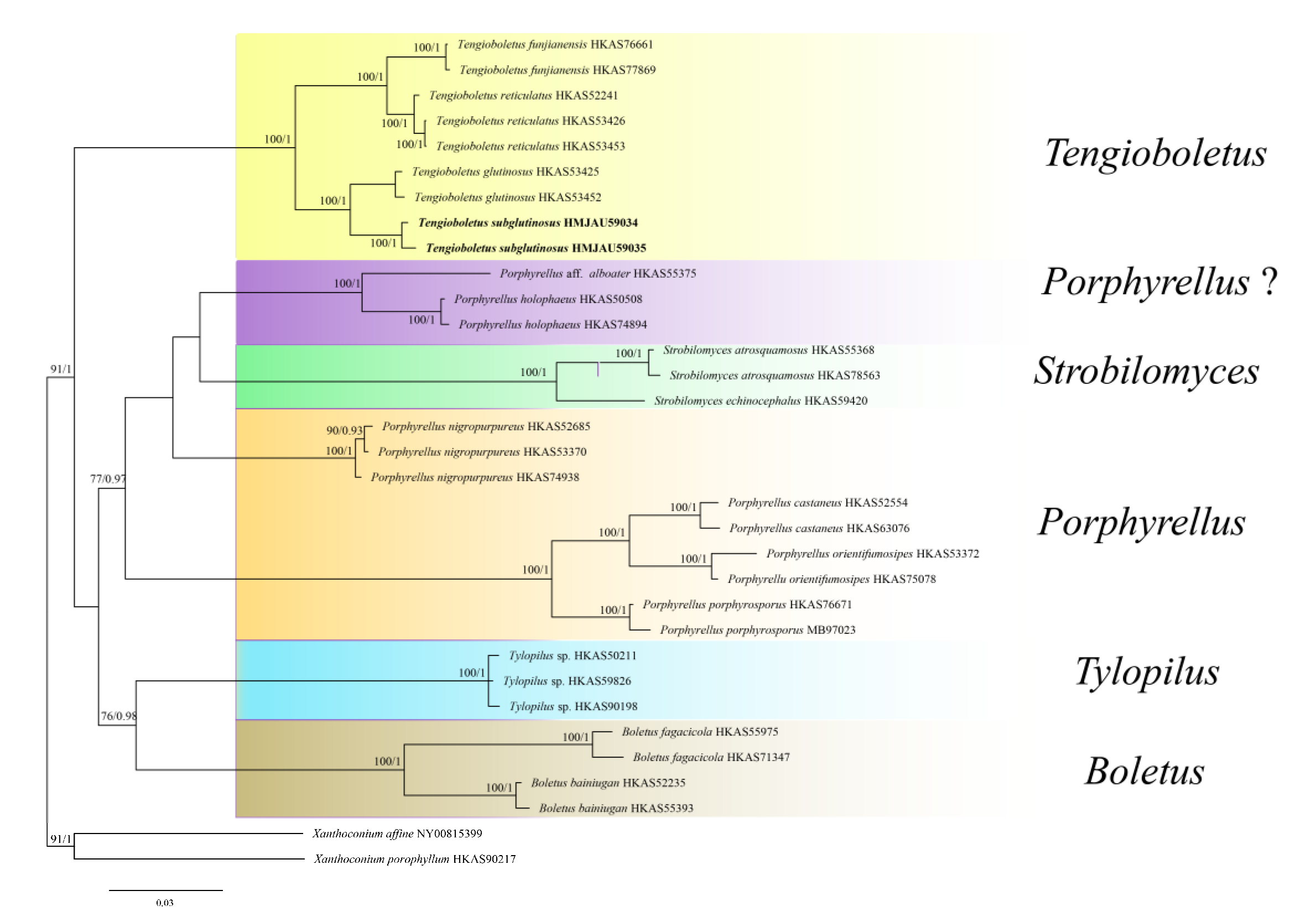
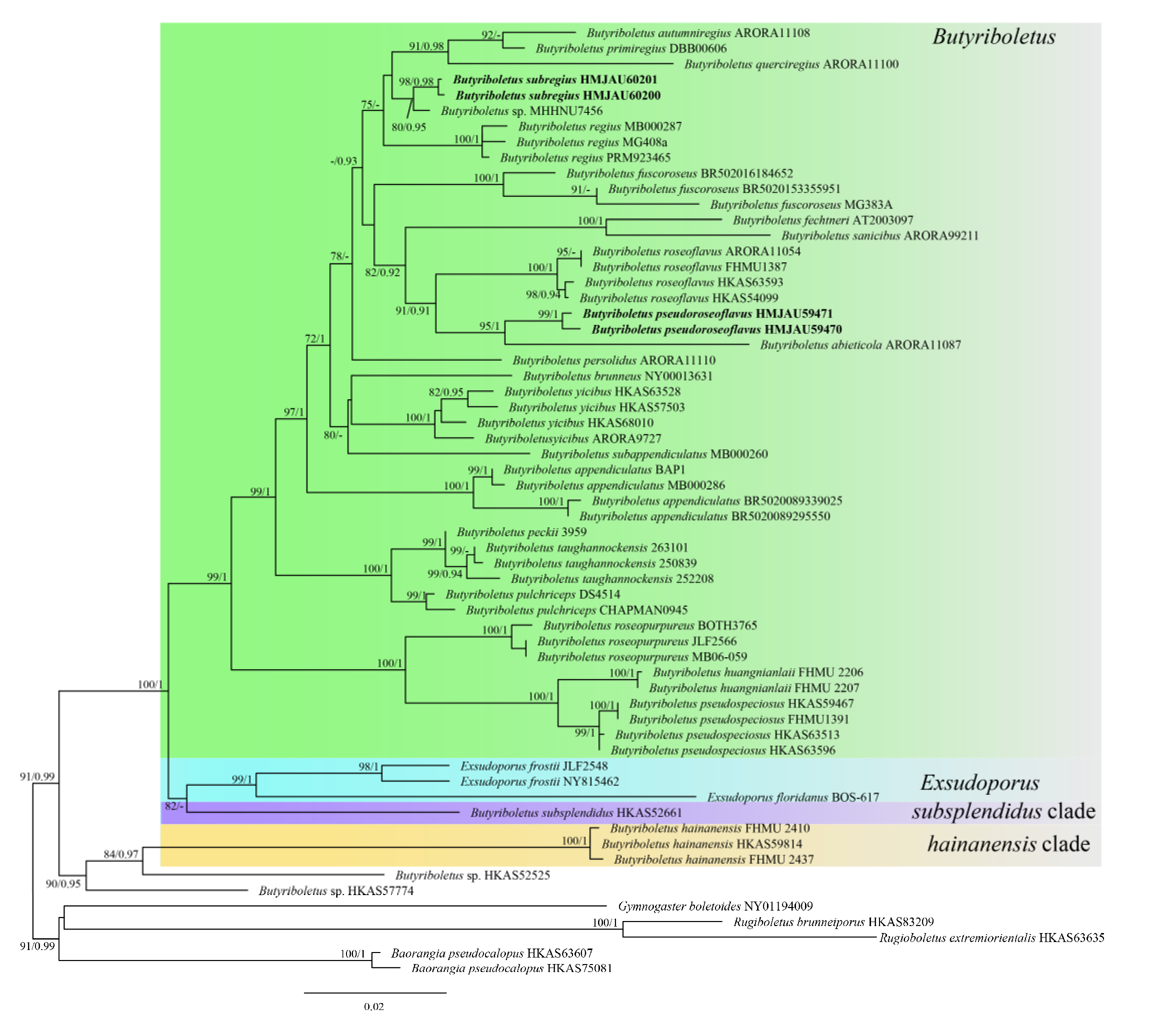
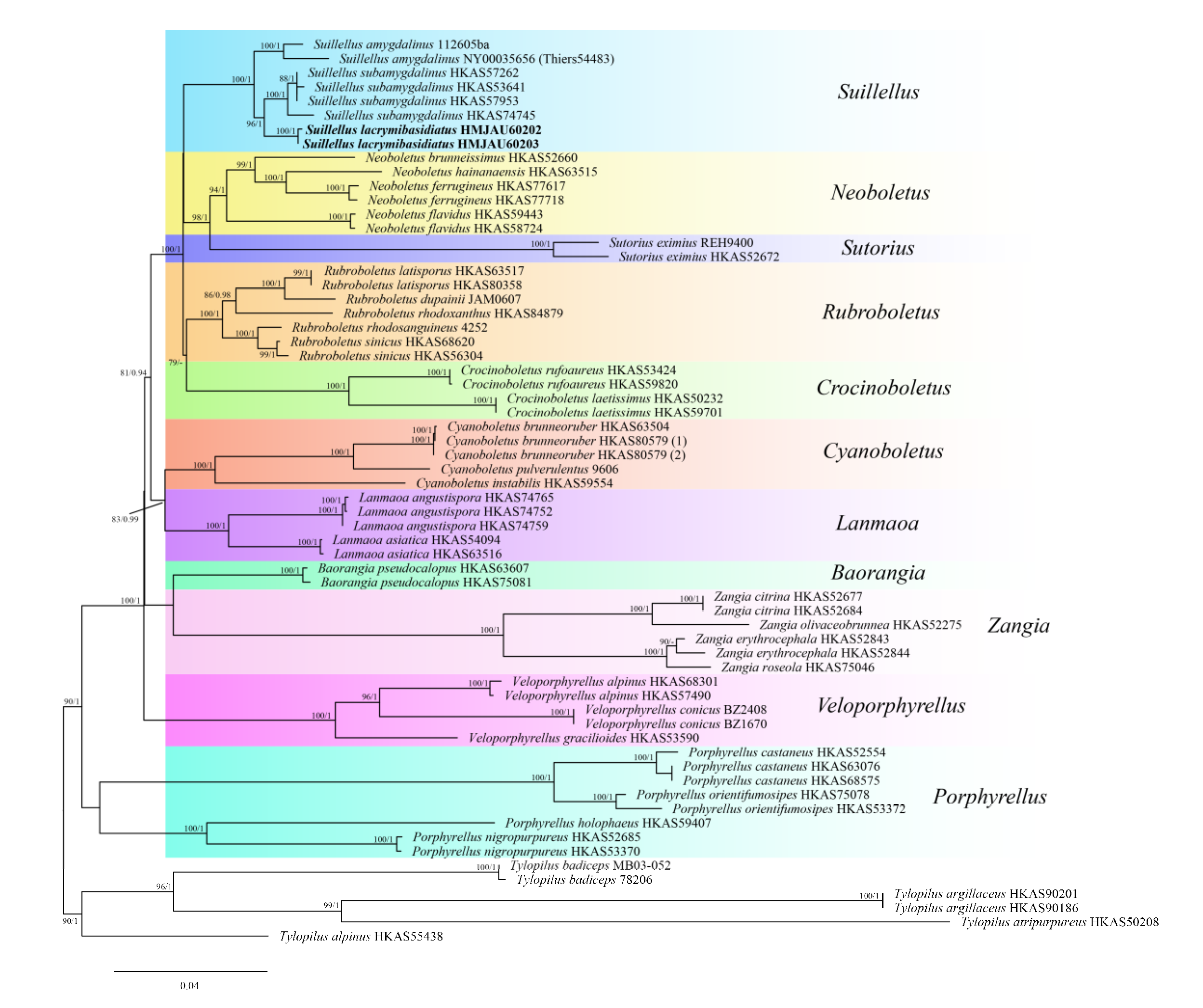
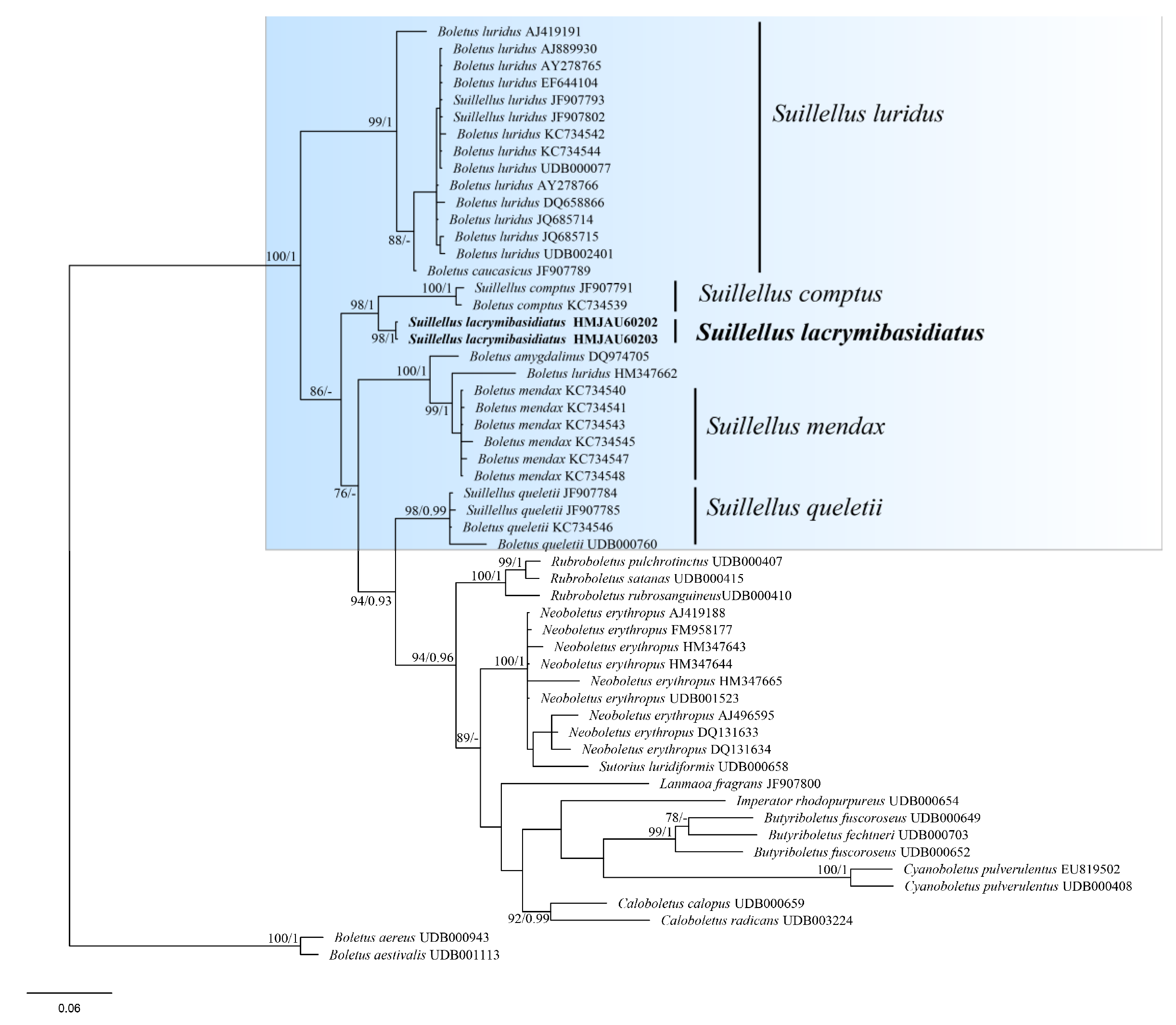
3.1. Molecular Phylogeny
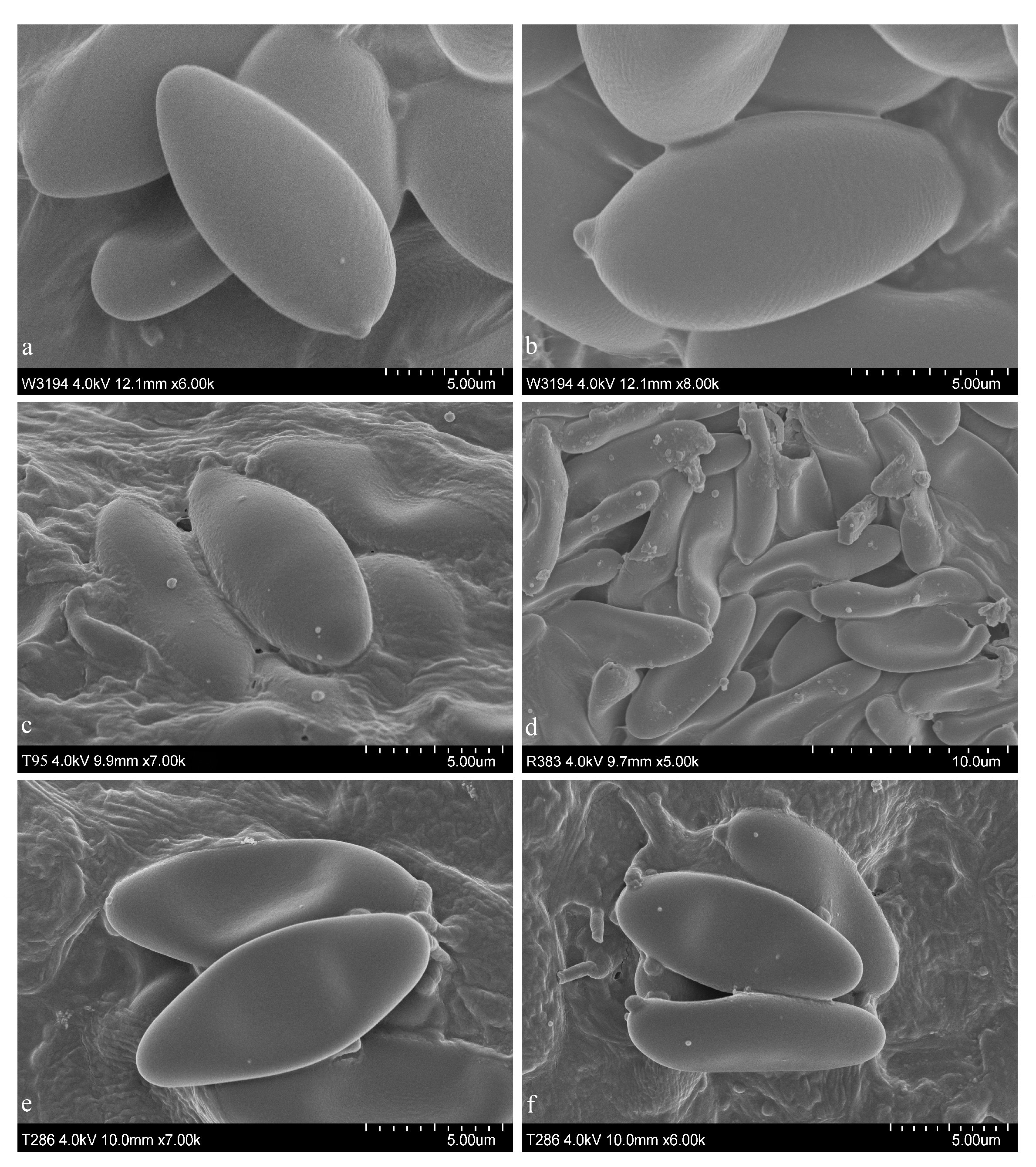
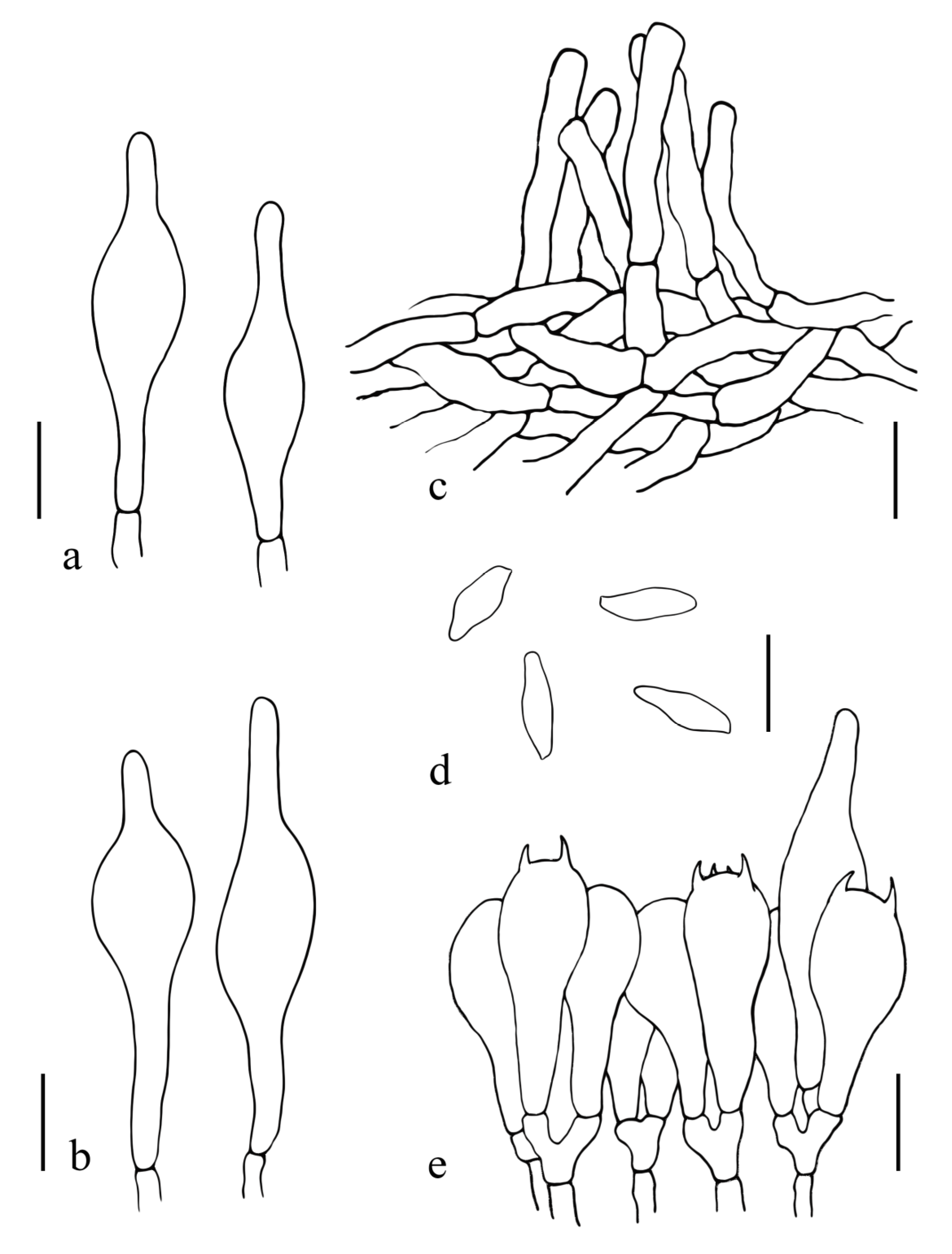
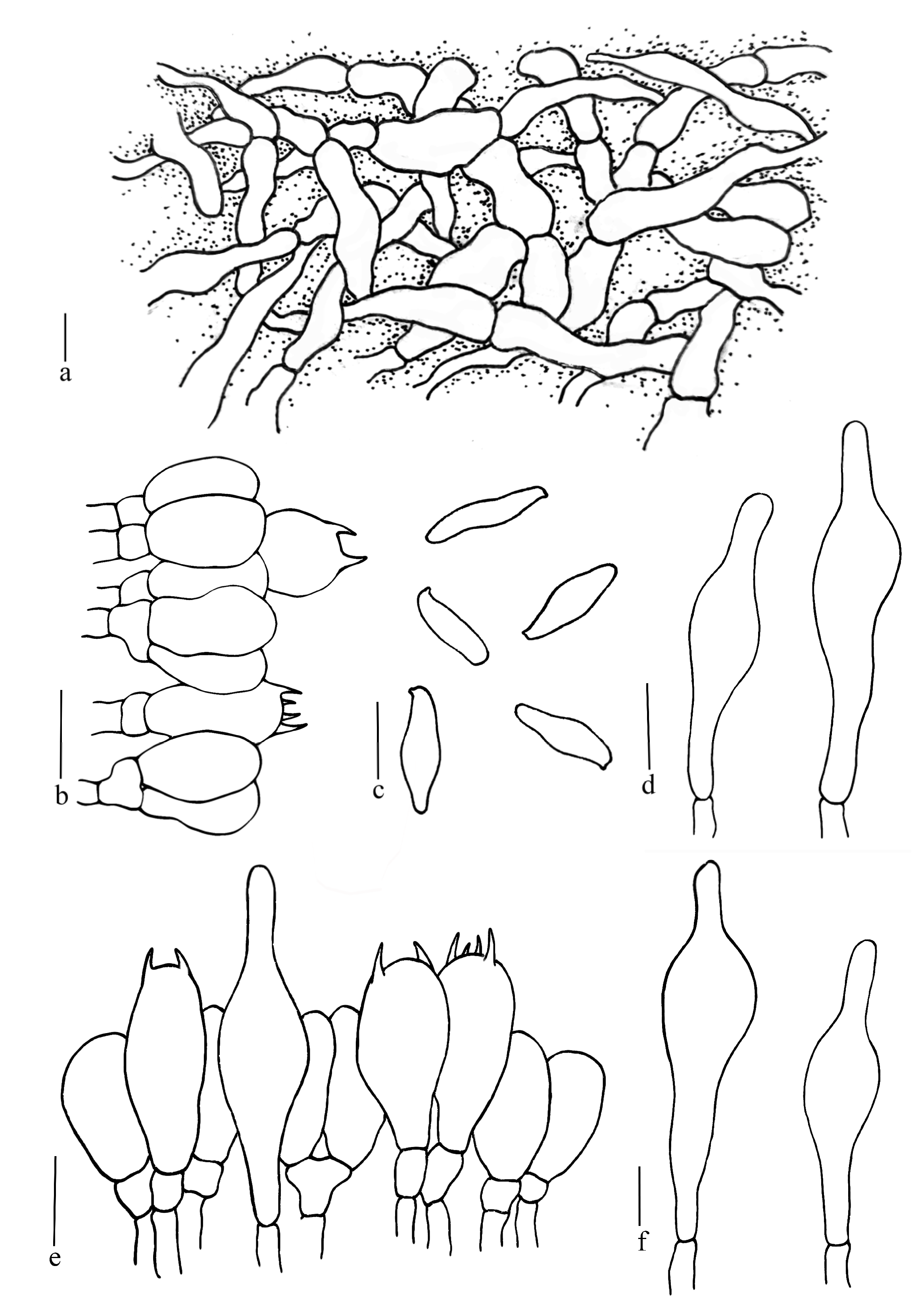
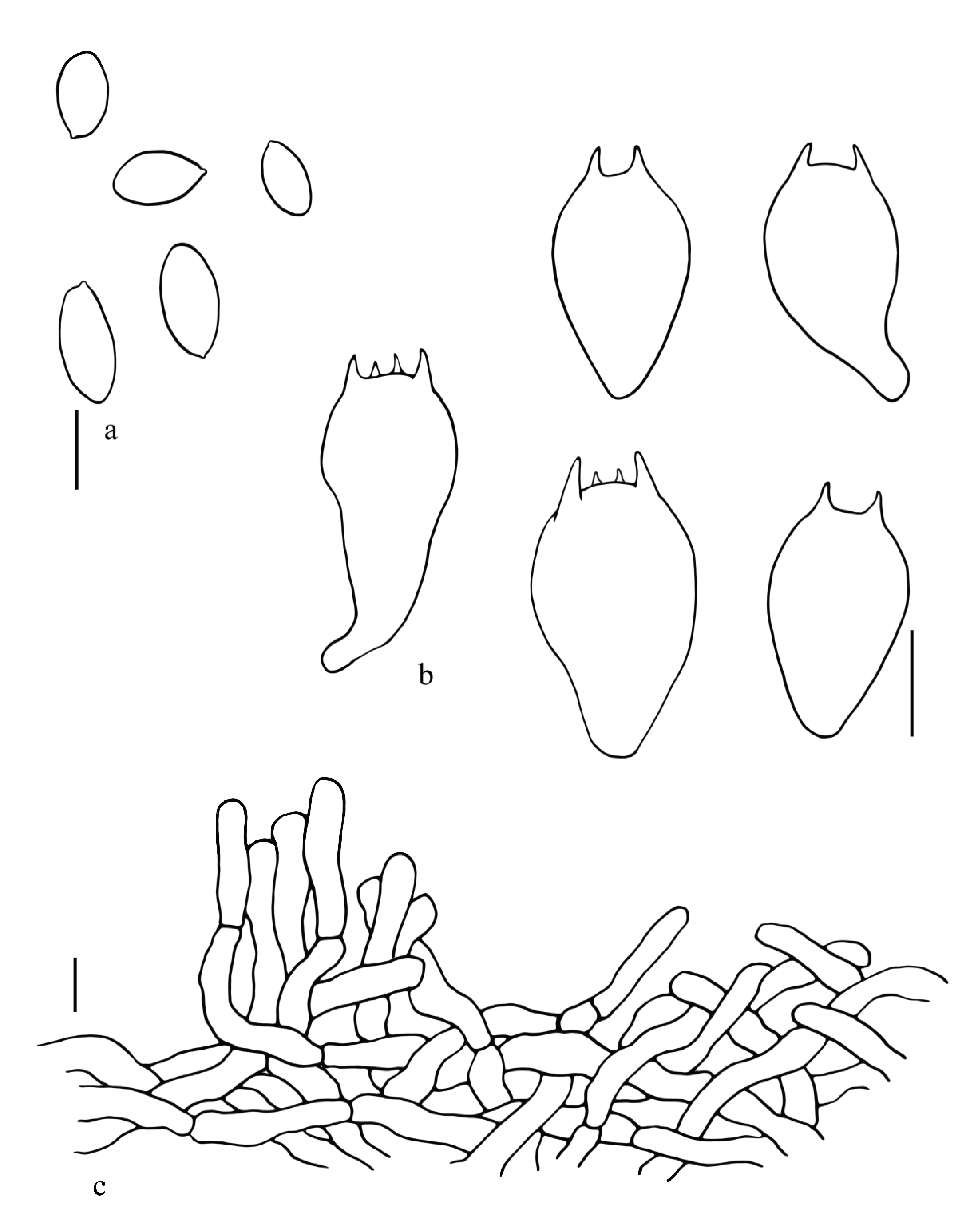
3.2. Taxonomy
| 1. Pores changing color when bruised | 2 |
| 1. Pores unchanging color when bruised | 3 |
| 2. Pores staining blue when bruised, pileipellis an ixotrichodermium | T. subglutinosus |
| 2. Pores staining brown when bruised, pileipellis an trichodermium | T. fujianensis |
| 3. Stipe covered with distinct reticulations, basidiospores larger, 12.0–14.5 × 4.5–6.0 μm, pileipellis an trichodermium | T. reticulatus |
| 3. Stipe nearly glabrous, basidiospores 10.0–12.0 × 3.5–4.5 μm, pileipellis, an ixotrichodermium | T. glutinosus |
| 1. Basidiospores usually longer than 15 μm | 2 |
| 1. Basidiospores shorter than or equal to 15 μm | 4 |
| 2. Stipe covered with pruinose or granulose, but without any trace of reticulation | S. amygdalinus |
| 2. Stipe covered with reticulation | 3 |
| 3. Basidia lacrymoid, Q = 1.5–2.1 | S. lacrymibasidiatus |
| 3. Basidia clavate, Q = 2.0–2.6 | S. subamygdalinus |
| 4. Surface of stipe rough, but without reticulation | 5 |
| 4. Stipe covered with reticulation | 6 |
| 5. Stipe covered with distinct pruinose, basidiospores less than 14 μm, basidia broad clavate | S. adonis |
| 5. Stipe covered with reddish to brown granules, basidiospores can be longer than 14 μm, hymenophoral basidia clavate | S. queletii |
| 6. Stipe covered with prominent, red to orange reticulation. Basidiospores 11–14 × 4.5–6 μm | S. luridus |
| 6. Stipe with indistinctly or finely reticulation, usually distributed erratically | 7 |
| 7. Q value higher than 2.6 | S. mendax |
| 7. Q value less than or equal to 2.6 | 8 |
| 8. Basidiospores dextrinoid, Q value can reach 2.6, reticulation yellow and fine at the upper portion of stipe | S. atlanticus |
| 8. Q value less than or equal to 2.2, pores red to orange red, stipe covered with very fine yellow, pale orange, orange, reddish orange, or pale red granules | S. comptus |
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chevallier, F.F. Flore Générale des Environs de Paris. Available online: https://bibdigital.rjb.csic.es/idurl/1/11834 (accessed on 20 December 2021).
- Zhang, M. Molecular Phylogenetic Studies on the Family Boletaceae in Southern China, and Taxonomic Study on the Genus Aureoboletus in China. Ph.D. Thesis, South China University of Technology, Guangzhou, China, 2016. [Google Scholar]
- Li, Y.; Li, T.H.; Yang, Z.L.; Bau, T.; Dai, Y.C. Atlas of Chinese Macrofungal Resources; Central Plains Farmers Press: Zhengzhou, China, 2016; pp. 1068–1148. [Google Scholar]
- Roman, M.D.; Claveria, V.; Miguel, A.M. A revision of the descriptions of ectomycorrhizas published since 1961. Mycol. Res. 2005, 109, 1063–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Li, Y.C.; Zhu, X.T.; Zhao, K.; Han, L.H.; Cui, Y.Y.; Li, F.; Xu, J.P.; Yang, Z.L. One hundred noteworthy boletes from China. Fungal Divers. 2016, 81, 25–188. [Google Scholar] [CrossRef]
- Yang, Z.L.; Wu, G.; Li, Y.C.; Wang, X.H.; Cai, Q. Common Edible and Poisonous Mushrooms of Southwestern China; Science Press: Beijing, China, 2021. [Google Scholar]
- Murrill, W.A. The Boletaceae of North America—I. Mycologia 1909, 1, 4–18. [Google Scholar] [CrossRef]
- Smith, A.H.; Thiers, H.D. Boletes of Michigan; The University of Michigan Press: Ann Arbor, MI, USA, 1971. [Google Scholar]
- Singer, R.; Williams, R. Some boletes from Florida. Mycologia 1992, 84, 724–728. [Google Scholar] [CrossRef]
- Baroni, T.J. Boletus aurantiosplendens sp. nov. from the southern Appalachian Mountains with notes on Pulveroboletus auriflammeus, Pulveroboletus melleouluteus and Boletus auripes. Bull. Buffalo Soc. Nat. Sci. 1998, 36, 245–255. [Google Scholar]
- Baroni, T.J.; Bessette, A.E.; Roody, W.C. Boletus patrioticus—A new species from the eastern United States. Bull. Buffalo Soc. Nat. Sci. 1998, 36, 265–268. [Google Scholar]
- Farid, A.; Gelardi, M.; Angelini, C.; Franck, A.; Costanzo, F.; Kaminsky, L.; Ercole, E.; Baroni, T.; White, A.; Garey, J. Phylloporus and Phylloboletellus are no longer alone: Phylloporopsis gen. nov. (Boletaceae), a new smooth-spored lamellate genus to accommodate the American species Phylloporus Boletinoides. Fungal Syst. Evol. 2018, 2, 341. [Google Scholar] [CrossRef]
- Vizzini, A.; Simonini, G.; Ercole, E.; Voyron, S. Boletus mendax, a new species of Boletus sect. Luridi from Italy and insights on the B. luridus complex. Mycol. Prog. 2014, 13, 95–109. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Santana, B.; Roody, W.C.; Both, E.E. A new arenicolous Boletus from the Gulf Coast of northern Florida. Mycotaxon 2009, 107, 243–247. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Santana, B.; Bessette, A.E.; McConnell, O.L. Boletus durhamensis sp. nov. from North Carolina. Mycotaxon 2016, 131, 703–715. [Google Scholar] [CrossRef]
- Frank, J.; Siegel, N.; Schwarz, C.; Araki, B.; Vellinga, E. Xerocomellus (Boletaceae) in western North America. Fungal Syst. Evol. 2020, 6, 265. [Google Scholar] [CrossRef]
- Crous, P.; Wingfield, M.; Lombard, L.; Roets, F.; Swart, W.; Alvarado, P.; Carnegie, A.; Moreno, G.; Luangsaard, J.; Thangavel, R. Fungal Planet description sheets: 951–1041. Persoonia 2019, 43, 223. [Google Scholar] [CrossRef]
- Farid, A.; Franck, A.R.; Bolin, J.; Garey, J.R. Expansion of the genus Imleria in North America to include Imleria floridana, sp. nov., and Imleria pallida, comb. nov. Mycologia 2020, 112, 423–437. [Google Scholar] [CrossRef]
- Farid, A.; Bessette, A.R.; Bolin, J.A.; Kudzma, L.V.; Franck, A.R.; Garey, J.R. Investigations in the boletes (Boletaceae) of southeastern USA: Four novel species and three novel combinations. Mycosphere 2021, 12, 1038–1076. [Google Scholar] [CrossRef]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibbett, D.S.; Fisher, M.C. Phylogenetic species recognition and species concepts in fungi. Fungal Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Nuhn, M.E.; Binder, M.; Taylor, A.F.; Halling, R.E.; Hibbett, D.S. Phylogenetic overview of the Boletineae. Fungal Biol. 2013, 117, 479–511. [Google Scholar] [CrossRef]
- Wu, G.; Feng, B.; Xu, J.P.; Zhu, X.T.; Li, Y.C.; Zeng, N.K.; Hosen, M.I.; Yang, Z.L. Molecular phylogenetic analyses redefine seven major clades and reveal 22 new generic clades in the fungal family Boletaceae. Fungal Divers. 2014, 69, 93–115. [Google Scholar] [CrossRef]
- Wilson, A.W.; Binder, M.; Hibbett, D.S. Diversity and evolution of ectomycorrhizal host associations in the Sclerodermatineae (Boletales, Basidiomycota). New Phytol. 2012, 194, 1079–1095. [Google Scholar] [CrossRef]
- Wu, G.; Miyauchi, S.; Morin, E.; Kuo, A.; Drula, E.; Varga, T.; Kohler, A.; Feng, B.; Cao, Y.; Lipzen, A. Evolutionary innovations through gain and loss of genes in the ectomycorrhizal Boletales. New Phytol. 2022, 233, 1383–1400. [Google Scholar] [CrossRef]
- Zhao, K.; Wu, G.; Yang, Z.L. A new genus, Rubroboletus, to accommodate Boletus sinicus and its allies. Phytotaxa 2014, 188, 61–77. [Google Scholar] [CrossRef]
- Zhu, X.T.; Wu, G.; Zhao, K.; Halling, R.E.; Yang, Z.L. Hourangia, a new genus of Boletaceae to accommodate Xerocomus cheoi and its allied species. Mycol. Prog. 2015, 14, 1–10. [Google Scholar] [CrossRef]
- Wu, G.; Zhao, K.; Li, Y.C.; Zeng, N.K.; Feng, B.; Halling, R.E.; Yang, Z.L. Four new genera of the fungal family Boletaceae. Fungal Divers. 2016, 81, 1–24. [Google Scholar] [CrossRef]
- Chai, H.; Liang, Z.Q.; Xue, R.; Jiang, S.; Luo, S.H.; Wang, Y.; Wu, L.L.; Tang, L.P.; Chen, Y.; Hong, D. New and noteworthy boletes from subtropical and tropical China. MycoKeys 2019, 46, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.X.; Wu, G.; Yang, Z.L. Four New Species of Hemileccinum (Xerocomoideae, Boletaceae) from Southwestern China. J. Fungi 2021, 7, 823. [Google Scholar] [CrossRef]
- Gelardi, M.; Vizzini, A.; Ercole, E.; Horak, E.; Ming, Z.; Li, T.H. Circumscription and taxonomic arrangement of Nigroboletus roseonigrescens gen. et sp. nov., a new member of Boletaceae from tropical South–Eastern China. PLoS ONE 2015, 10, e0134295. [Google Scholar] [CrossRef]
- Cui, Y.Y.; Feng, B.; Wu, G.; Xu, J.P.; Yang, Z.L. Porcini mushrooms (Boletus sect. Boletus) from China. Fungal Divers. 2016, 81, 189–212. [Google Scholar] [CrossRef]
- Liang, Z.Q.; An, D.Y.; Juang, S.; Su, M.S.; Zeng, N.K. Butyriboletus hainanensis (Boletaceae, Boletales), a new species from tropical China. Phytotaxa 2016, 267, 256–262. [Google Scholar] [CrossRef]
- Arora, D.; Frank, J.L. Clarifying the butter Boletes: A new genus, Butyriboletus, is established to accommodate Boletus sect. Appendiculati, and six new species are described. Mycologia 2014, 106, 464–480. [Google Scholar] [CrossRef]
- Zeng, N.K.; Chai, H.; Jiang, S.; Xue, R.; Wang, Y.; Hong, D.; Liang, Z.Q. Retiboletus nigrogriseus and Tengioboletus fujianensis, two new boletes from the south of China. Phytotaxa 2018, 367, 45–54. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Pavey, D., Ed.; Eyre Methuen: London, UK, 1978. [Google Scholar]
- Imler, L. Recherches sur les bolets. Bull. Soc. Mycol. Fr. 1950, 66, 177–203. [Google Scholar]
- Biketova, A.Y.; Gelardi, M.; Smith, M.E.; Simonini, G.; Healy, R.A.; Taneyama, Y.; Vasquez, G.; Kovács, A.; Nagy, L.G.; Wasser, S.P.; et al. Reappraisal of the Genus Exsudoporus (Boletaceae) Worldwide Based on Multi-Gene Phylogeny, Morphology and Biogeography, and Insights on Amoenoboletus. J. Fungi 2022, 8, 101. [Google Scholar] [CrossRef]
- Zhu, X.T.; Li, Y.C.; Wu, G.; Feng, B.; Zhao, K.; Gelardi, M.; Kost, G.W.; Yang, Z.L. The genus Imleria (Boletaceae) in East Asia. Phytotaxa 2014, 191, 81–98. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Cubeta, M.; Echandi, E.; Abernethy, T.; Vilgalys, R. Characterization of anastomosis groups of binucleate Rhizoctonia species using restriction analysis of an amplified ribosomal RNA gene. Phytopathology 1991, 81, 1395–1400. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Zhang, M.; Li, T.H.; Song, B. Two new species of Chalciporus (Boletaceae) from southern China revealed by morphological characters and molecular data. Phytotaxa 2017, 327, 47–56. [Google Scholar] [CrossRef]
- Kuo, M.; Ortiz-Santana, B. Revision of leccinoid fungi, with emphasis on North American taxa, based on molecular and morphological data. Mycologia 2020, 112, 197–211. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Binder, M.; Hibbett, D.S. Molecular systematics and biological diversification of Boletales. Mycologia 2006, 98, 971–981. [Google Scholar] [CrossRef]
- Feng, B.; Xu, J.P.; Wu, G.; Zeng, N.K.; Li, Y.C.; Tolgor, B.; Kost, G.W.; Yang, Z.L. DNA sequence analyses reveal abundant diversity, endemism and evidence for Asian origin of the porcini mushrooms. PLoS ONE 2012, 7, e37567. [Google Scholar] [CrossRef]
- Binder, M.; Bresinsky, A. Retiboletus, a new genus for a species-complex in the Boletaceae producing retipolides. Feddes Repert. Z. Bot. Taxon. Geobot. 2002, 113, 30–40. [Google Scholar] [CrossRef]
- Zhao, K.; Wu, G.; Halling, R.E.; Yang, Z.L. Three new combinations of Butyriboletus (Boletaceae). Phytotaxa 2015, 234, 51–62. [Google Scholar] [CrossRef]
- Zhao, K.; Wu, G.; Feng, B.; Yang, Z.L. Molecular phylogeny of Caloboletus (Boletaceae) and a new species in East Asia. Mycol. Prog. 2014, 13, 1127–1136. [Google Scholar] [CrossRef]
- Halling, R.E.; Nuhn, M.; Fechner, N.A.; Osmundson, T.W.; Soytong, K.; Arora, D.; Hibbett, D.S.; Binder, M. Sutorius: A new genus for Boletus eximius. Mycologia 2012, 104, 951–961. [Google Scholar] [CrossRef] [Green Version]
- Šutara, J.; Janda, V.; Kříž, M.; Graca, M.; Kolařík, M. Contribution to the study of genus Boletus, section Appendiculati: Boletus roseogriseus sp. nov. and neotypification of Boletus fuscoroseus Smotl. Czech Mycol. 2014, 66, 1–37. [Google Scholar] [CrossRef]
- Li, Y.C.; Ortiz-Santana, B.; Zeng, N.K.; Feng, B.; Yang, Z.L. Molecular phylogeny and taxonomy of the genus Veloporphyrellus. Mycologia 2014, 106, 291–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.C.; Feng, B.; Yang, Z.L. Zangia, a new genus of Boletaceae supported by molecular and morphological evidence. Fungal Divers. 2011, 49, 125–143. [Google Scholar] [CrossRef]
- Krpata, D.; Peintner, U.; Langer, I.; Fitz, W.J.; Schweiger, P. Ectomycorrhizal communities associated with Populus tremula growing on a heavy metal contaminated site. Mycol. Res. 2008, 112, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Osmundson, T.W.; Robert, V.A.; Schoch, C.L.; Baker, L.J.; Smith, A.; Robich, G.; Mizzan, L.; Garbelotto, M.M. Filling gaps in biodiversity knowledge for macrofungi: Contributions and assessment of an herbarium collection DNA barcode sequencing project. PLoS ONE 2013, 8, e62419. [Google Scholar] [CrossRef] [Green Version]
- Iotti, M.; Barbieri, E.; Stocchi, V.; Zambonelli, A. Morphological and molecular characterisation of mycelia of ectomycorrhizal fungi in pure culture. Fungal Divers. 2005, 19, 51–68. [Google Scholar]
- Nygren, C.M.; Edqvist, J.; Elfstrand, M.; Heller, G.; Taylor, A.F. Detection of extracellular protease activity in different species and genera of ectomycorrhizal fungi. Mycorrhiza 2007, 17, 241–248. [Google Scholar] [CrossRef]
- Martin, M.P.; Raidl, S. The taxonomic position of Rhizopogon melanogastroides (Boletales). Mycotaxon 2002, 84, 221–228. [Google Scholar]
- Smith, M.E.; Douhan, G.W.; Rizzo, D.M. Ectomycorrhizal community structure in a xeric Quercus woodland based on rDNA sequence analysis of sporocarps and pooled roots. New Phytol. 2007, 174, 847–863. [Google Scholar] [CrossRef]
- Mello, A.; Ghignone, S.; Vizzini, A.; Sechi, C.; Ruiu, P.; Bonfante, P. ITS primers for the identification of marketable boletes. J. Biotechnol. 2006, 121, 318–329. [Google Scholar] [CrossRef]
- Palmer, J.M.; Lindner, D.L.; Volk, T.J. Ectomycorrhizal characterization of an American chestnut (Castanea dentata)-dominated community in Western Wisconsin. Mycorrhiza 2008, 19, 27–36. [Google Scholar] [CrossRef]
- Li, H.; Wei, H.; Peng, H.; Ding, H.; Wang, L.; He, L.; Fu, L. Boletus roseoflavus, a new species of Boletus in section Appendiculati from China. Mycol. Prog. 2014, 13, 21–31. [Google Scholar] [CrossRef]
- Takahashi, H.; Taneyama, Y.; Degawa, Y. Notes on the boletes of Japan 1. Four new species of the genus Boletus from central Honshu, Japan. Mycoscience 2013, 54, 458–468. [Google Scholar] [CrossRef]
- Janda, V.; Kříž, M.; Kolařík, M. Butyriboletus regius and Butyriboletus fechtneri: Typification of two well-known species. Czech Mycol. 2019, 71, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Thiers, H.D. California Mushrooms—A Field Guide to the Boletes; Hafner Press: New York, NY, USA, 1975; p. 261. [Google Scholar]
- Muñoz, J.A. Boletus s.l. (excl. Xerocomus). In Fungi Europaei 2; Candusso Editrice: Bardolino, Italy, 2005; pp. 428–432. [Google Scholar]
- Heykoop, M. Morphology and taxonomy of Boletus queletii var. discolor, a rare bolete resembling Boletus erythropus. Mycotaxon 1995, 56, 115–123. [Google Scholar]
- Marsico, O.; Musumeci, E. Boletus adalgisae sp. nov. Boll. Assoc. Micol. Ecol. Romana 2011, 27, 3–15. [Google Scholar]
- Seaver, F.J.; John, N.C.; Murrill, W.A.; George, L.Z.; Fred, W.; Singer, R. Notes and Brief Articles. Mycologia 1945, 37, 792–799. [Google Scholar] [CrossRef]
- Pilát, A. Boletus gabretae sp. nov. bohemica ex affinitate Boleti junguillei (Quél.) Boud. Czech Mycol. 1968, 22, 167–170. [Google Scholar]
- Murill, W.A. More Florida fungi. Lloydia 1946, 8, 263–290. [Google Scholar]
- New York State Museum. Bulletin of the New York State Museum; University of the State of New York: New York, NY, USA, 1889; Volume 2, pp. 142–143. [Google Scholar]
- De Queiroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Truong, C.; Mujic, A.B.; Healy, R.; Kuhar, F.; Furci, G.; Torres, D.; Niskanen, T.; Sandoval-Leiva, P.A.; Fernández, N.; Escobar, J.M. How to know the fungi: Combining field inventories and DNA-barcoding to document fungal diversity. New Phytol. 2017, 214, 913–919. [Google Scholar] [CrossRef] [Green Version]
- Bozok, F.; Assyov, B.; Taşkin, H. First records of Exsudoporus permagnificus and Pulchroboletus roseoalbidus (Boletales) in association with non-native Fagaceae, with taxonomic remarks. Phytol. Balc. 2019, 25, 13–27. [Google Scholar]
- Liu, H.Y. Taxonomy and Resource Evaluation of Boletes in Northeastern China. Master’s Thesis, Jilin Agricultural University, Changchun, China, 2020. [Google Scholar]
- Muñoz, J.; Boletus, S.L. (Excl. Xerocomus): Strobilomycetaceae, Gyroporaceae, Gyrodontaceae, Suillaceae, Boletaceae. In Fungi Europaei 2; Edizioni Candusso: Alassio, Italy, 2005. [Google Scholar]
- Han, L.H.; Wu, G.; Horak, E.; Halling, R.; Xu, J.P.; Ndolo, E.; Sato, H.; Fechner, N.; Sharma, Y.; Yang, Z.L. Phylogeny and species delimitation of Strobilomyces (Boletaceae), with an emphasis on the Asian species. Persoonia 2020, 44, 113–139. [Google Scholar] [CrossRef] [PubMed]
- Klofac, W. Schlüssel zur Bestimmung von Frischfunden der europäischen Arten der Boletales mit röhrigem Hymenophor. Osterr. Z. Pilzkd. 2007, 16, 187–279. [Google Scholar]
- Vesterholt, J. Funga Nordica, Agaricoid, Boletoid, Cyphelloid and Gasteroid Genera; Nordsvamp: Copenhagen, Denmark, 2012; p. 9811021. [Google Scholar]
- Gelardi, M. Contribution to the knowledge of Chinese boletes. II: Aureoboletus thibetanus sl, Neoboletus brunneissimus, Pulveroboletus macrosporus and Retiboletus kauffmanii (Part I). Riv. Micol. Romana 2017, 102, 13–30. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Voucher ID | ITS | 28S | TEF1 | RPB1 | RPB2 | References |
|---|---|---|---|---|---|---|---|
| Tengioboletus glutinosus | HKAS53425 | – | KF112341 | KF112204 | KF112578 | KF112800 | [22] |
| T. glutinosus | HKAS53452 | – | KT990655 | KT990844 | KT990994 | KT990480 | [5] |
| T. reticulatus | HKAS53426 | – | KF112491 | KF112313 | KF112649 | KF112828 | [22] |
| T. reticulatus | HKAS52241 | – | KT990657 | KT990845 | KT990995 | KT990481 | [5] |
| T. reticulatus | HKAS53453 | – | KT990656 | KT990846 | – | KT990482 | [5] |
| T. funjianensis | HKAS76661 | – | KF112342 | KF112205 | – | KF112801 | [22] |
| T. funjianensis | HKAS77869 | – | KT990658 | KT990847 | KT990996 | KT990483 | [5] |
| T. subglutinosus | HMJAU59034 (T286) | – | OL588198 | OL739119 | OL739121 | – | this study |
| T. subglutinosus | HMJAU59035 (T293) | – | OL588197 | OL739120 | OL739122 | OL739118 | this study |
| Porphyrellus porphyrosporus | MB97-023 | – | DQ534643 | GU187734 | GU187475 | GU187800 | [50] |
| P. porphyrosporus | HKAS76671 | – | KF112482 | KF112243 | KF112611 | KF112718 | [22] |
| Tylopilus sp. | HKAS50211 | – | KT990552 | KT990752 | KT990920 | KT990389 | [22] |
| Tylopilus sp. | HKAS59826 | – | KT990558 | – | – | – | [5] |
| Tylopilus sp. | HKAS90198 | – | KT990559 | – | – | – | [5] |
| Strobilomyces atrosquamosus | HKAS55368 | – | KT990648 | KT990839 | KT990989 | KT990476 | [5] |
| S. atrosquamosus | HKAS78563 | – | KT990649 | KT990833 | KT990983 | KT990470 | [5] |
| S. echinocephalus | HKAS59420 | – | KF112463 | KF112256 | KF112600 | KF112810 | [22] |
| P. aff. alboater | HKAS55375 | – | KT990622 | KT990816 | KT990969 | – | [5] |
| P. nigropurpureus | HKAS74938 | – | KF112466 | KF112246 | – | KF112763 | [22] |
| P. nigropurpureus | HKAS52685 | – | KT990627 | KT990821 | KT990973 | KT990459 | [5] |
| P. nigropurpureus | HKAS53370 | – | KT990628 | KT990822 | KT990974 | KT990460 | [5] |
| P. holophaeus | HKAS50508 | – | KF112465 | KF112244 | KF112553 | – | [22] |
| P. holophaeus | HKAS74894 | – | KF112474 | KF112245 | KF112554 | – | [22] |
| P. castaneus | HKAS63076 | – | KT990548 | KT990749 | KT990916 | KT990386 | [5] |
| P. castaneus | HKAS52554 | – | KT990697 | KT990883 | KT991026 | KT990502 | [5] |
| P. orientifumosipes | HKAS75078 | – | KF112481 | KF112242 | – | KF112717 | [22] |
| P. orientifumosipes | HKAS53372 | – | KT990629 | KT990823 | KT990975 | KT990461 | [5] |
| Boletus bainiugan | HKAS52235 | – | KF112457 | KF112203 | KF112587 | KF112705 | [22] |
| B. bainiugan | HKAS55393 | – | JN563852 | – | JN563868 | – | [51] |
| B. fagacicola | HKAS55975 | – | JN563853 | – | JN563879 | – | [51] |
| B. fagacicola | HKAS71347 | – | JQ172790 | – | JQ172791 | – | [51] |
| Xanthoconium affine | NY00815399 (REH8660) | – | KT990661 | KT990850 | KT990999 | KT990486 | [5] |
| X. porophyllum | HKAS90217 | – | KT990662 | KT990851 | KT991000 | KT990487 | [5] |
| Baorangia pseudocalopus | HKAS63607 | – | KF112355 | KF112167 | – | – | [22] |
| Ba. pseudocalopus | HKAS75081 | – | KF112356 | KF112168 | – | – | [22] |
| Butyriboletus abieticola | Arora11087 | KC184412 | KC184413 | – | – | – | [33] |
| Bu. appendiculatus | Bap1 | KJ419923 | AF456837 | JQ327025 | – | – | [52] |
| Bu. appendiculatus | BR50200893390-25 | KT002598 | KT002609 | KT002633 | – | – | [53] |
| Bu. appendiculatus | BR50200892955-50 | KJ605668 | KJ605677 | KJ619472 | – | KP055030 | [54] |
| Bu. appendiculatus | MB000286 | KT002599 | KT002610 | KT002634 | – | – | [53] |
| Bu. autumniregius | Arora11108 | KC184423 | KC184424 | – | – | – | [33] |
| Bu. brunneus | NY00013631 | KT002600 | KT002611 | KT002635 | – | – | [53] |
| Bu. fechtneri | AT2003097 | KC584784 | KF030270 | – | – | – | [21] |
| Exsudoporus frostii | JLF2548 | KC812303 | KC812304 | – | – | – | [33] |
| E. frostii | NY815462 | – | JQ924342 | KF112164 | – | KF112675 | [22] |
| E. floridanus | BOS 617, BZ 3170 | MN250222 | MK601725 | MK721079 | – | MK766287 | [43] |
| Bu. hainanensis | N.K. Zeng 1197 (FHMU 2410) | KU961653 | KU961651 | – | – | KU961658 | [32] |
| Bu. hainanensis | N.K. Zeng 2418 (FHMU 2437) | KU961654 | KU961652 | KU961656 | – | KX453856 | [32] |
| Bu. huangnianlaii | N.K. Zeng 3245 (FHMU 2206) | MH885350 | MH879688 | MH879717 | – | MH879740 | [28] |
| Bu. huangnianlaii | N.K. Zeng 3246 (FHMU 2207) | MH885351 | MH879689 | MH879718 | – | MH879741 | [28] |
| Bu. peckii | 3959 | – | JQ326999 | JQ327026 | – | – | [55] |
| Bu. persolidus | Arora11110 | KC184444 | – | – | – | – | [33] |
| Bu. primiregius | DBB00606 | – | KC184451 | – | – | – | [33] |
| Bu. fuscoroseus | BR50201618465-02 | KT002602 | KT002613 | KT002637 | – | – | [53] |
| Bu. fuscoroseus | BR50201533559-51 | KT002603 | KT002614 | KT002638 | – | – | [53] |
| Bu. pseudospeciosus | HKAS59467 | – | KF112331 | KF112176 | – | KF112672 | [22] |
| Bu. pseudospeciosus | HKAS63513 | – | KT990541 | KT990743 | – | KT990380 | [5] |
| Bu. pseudospeciosus | HKAS63596 | – | KT990542 | KT990744 | – | KT990381 | [5] |
| Bu. pseudospeciosus | N.K. Zeng 2127 (FHMU 1391) | MH885349 | MH879687 | MH879716 | – | – | [28] |
| Bu. fuscoroseus | MG383a | KC184458 | – | – | – | – | [33] |
| Bu. pulchriceps | DS4514 | – | KF030261 | KF030409 | – | – | [21] |
| Bu. pulchriceps | R. Chapman 0945 | KT002604 | KT002615 | KT002639 | – | – | [53] |
| Bu. querciregius | Arora11100 | KC184461 | – | – | – | – | [33] |
| Bu. regius | MB000287 | KT002605 | KT002616 | KT002640 | – | – | [53] |
| Bu. regius | MG408a | KC584789 | KC584790 | – | – | – | [33] |
| Bu. regius | PRM:923465 | KJ419920 | KJ419931 | – | – | – | [56] |
| Bu. roseoflavus | Arora11054 | KC184434 | KC184435 | – | – | – | [33] |
| Bu. roseoflavus | HKAS63593 | KJ909517 | KJ184559 | KJ184571 | – | – | [53] |
| Bu. roseoflavus | HKAS54099 | KJ909519 | KF739665 | KF739779 | – | – | [53] |
| Bu. roseoflavus | N.K. Zeng 2123 (FHMU 1387) | MH885348 | MH879686 | MH879715 | – | – | [28] |
| Bu. pseudoroseoflavus | HMJAU59470 (T274) | OL604164 | OL587853 | OL739124 | – | OL739126 | this study |
| Bu. pseudoroseoflavus | HMJAU59471 (R383) | OL604165 | OL587852 | OL739123 | – | OL739125 | this study |
| Bu. Roseopurpureus | E.E. Both3765 | KT002606 | KT002617 | KT002641 | – | – | [53] |
| Bu. Roseopurpureus | JLF2566 | KC184466 | KC184467 | – | – | – | [33] |
| Bu. Roseopurpureus | MB06-059 | KC184464 | KF030262 | KF030410 | – | – | [21] |
| Bu. sanicibus | Arora99211 | KC184469 | KC184470 | – | – | – | [33] |
| Bu. subregius | HMJAU60200 (T95) | OM237336 | OM237339 | OM285111 | – | OM285109 | this study |
| Bu. subregius | HMJAU60201 (T198) | OM237337 | OM237340 | OM285112 | – | OM285110 | this study |
| Butyriboletus sp. | MHHNU7456 | – | KT990539 | KT990741 | – | KT990378 | [5] |
| Butyriboletus sp. | HKAS52525 | – | KF112337 | KF112163 | – | KF112671 | [22] |
| Butyriboletus sp. | HKAS57774 | – | KF112330 | KF112155 | – | KF112670 | [22] |
| Bu. hainanensis | HKAS59814 | – | KF112336 | KF112199 | – | KF112699 | [22] |
| Butyriboletus yicibus | HKAS63528 | – | KF112332 | KF112156 | – | KF112673 | [22] |
| Bu. Subappendiculatus | MB000260 | KT002607 | KT002618 | KT002642 | – | – | [53] |
| Bu. subsplendidus | HKAS52661 | – | KF112339 | KF112169 | – | KF112676 | [5] |
| Bu. taughannockensis | 263101 | MH257559 | MH236172 | – | – | – | |
| Bu. taughannockensis | 250839 | MH234472 | MH234473 | – | – | – | |
| Bu. taughannockensis | 252208 | MH236100 | MH236099 | – | – | – | |
| Bu. yicibus | Arora9727 | KC184474 | KC184475 | – | – | – | [33] |
| Bu. yicibus | HKAS57503 | KT002608 | KT002620 | KT002644 | – | – | [53] |
| Bu. yicibus | HKAS68010 | KJ909521 | KT002619 | KT002643 | – | – | [53] |
| Gymnogaster boletoides | NY01194009 (REH9455) | – | KT990572 | KT990768 | – | KT990406 | [5] |
| Rugiboletus brunneiporus | HKAS83209 | – | KM605134 | KM605144 | – | KM605168 | [27] |
| R. extremiorientalis | HKAS63635 | – | KF112403 | KF112198 | – | KF112720 | [22] |
| Crocinoboletus laetissimus | HKAS50232 | — | KT990567 | KT990762 | KT990925 | — | [5] |
| C. laetissimus | HKAS59701 | — | KF112436 | — | — | KF112711 | [22] |
| C. rufoaureus | HKAS53424 | — | KF112435 | KF112206 | KF112533 | KF112710 | [22] |
| C. rufoaureus | HKAS59820 | — | KF112434 | — | KF112532 | KF112709 | [22] |
| Cyanoboletus brunneoruber | HKAS63504 | — | KF112368 | KF112194 | KF112531 | KF112702 | [22] |
| Cy. brunneoruber | HKAS80579 (1) | — | KT990568 | KT990763 | KT990926 | KT990401 | [5] |
| Cy. brunneoruber | HKAS80579 (2) | — | KT990569 | KT990764 | KT990927 | KT990402 | [5] |
| Cy. instabilis | HKAS59554 | — | KF112412 | KF112186 | KF112528 | KF112698 | [22] |
| Cy. pulverulentus | 9606 | — | KF030313 | KF030418 | KF030364 | — | [21] |
| Baorangia pseudocalopus | HKAS63607 | — | KF112355 | KF112167 | KF112519 | KF112677 | [22] |
| Ba. pseudocalopus | HKAS75081 | — | KF112356 | KF112168 | KF112520 | KF112678 | [22] |
| Lanmaoa angustispora | HKAS74765 | — | KF112322 | KF112159 | KF112521 | KF112680 | [22] |
| L. angustispora | HKAS74752 | — | KM605139 | KM605154 | KM605166 | KM605177 | [27] |
| L. angustispora | HKAS74759 | — | KM605140 | KM605155 | KM605167 | KM605178 | [27] |
| L. asiatica | HKAS54094 | — | KF112353 | KF112161 | KF112522 | KF112682 | [22] |
| L. asiatica | HKAS63516 | — | KT990584 | KT990780 | KT990935 | KT990419 | [5] |
| L. fragrans | 18555 | JF907800 | – | – | – | – | |
| Neoboletus brunneissimus | HKAS52660 | — | KF112314 | KF112143 | KF112492 | KF112650 | [22] |
| N. hainanaensis | HKAS63515 | — | KT990614 | KT990808 | KT990964 | KT990449 | [5] |
| N. ferrugineus | HKAS77617 | — | KT990595 | KT990788 | KT990943 | KT990430 | [5] |
| N. ferrugineus | HKAS77718 | — | KT990596 | KT990789 | KT990944 | KT990431 | [5] |
| N. flavidus | HKAS59443 | — | KU974139 | KU974136 | KU974142 | KU974144 | [5] |
| N. flavidus | HKAS58724 | — | KU974140 | KU974137 | KU974143 | KU974145 | [5] |
| Porphyrellus castaneus | HKAS52554 | — | KT990697 | KT990883 | KT991026 | KT990502 | [5] |
| P. castaneus | HKAS63076 | — | KT990548 | KT990749 | KT990916 | KT990386 | [5] |
| P. castaneus | HKAS68575 | — | KT990560 | — | — | — | [5] |
| P. holophaeus | HKAS59407 | — | KT990708 | KT990888 | KT991030 | KT990506 | [5] |
| P. nigropurpureus | HKAS52685 | — | KT990627 | KT990821 | KT990973 | KT990459 | [5] |
| P. nigropurpureus | HKAS53370 | — | KT990628 | KT990822 | KT990974 | KT990460 | [5] |
| P. orientifumosipes | HKAS75078 | — | KF112481 | KF112242 | — | KF112717 | [22] |
| P. orientifumosipes | HKAS53372 | — | KT990629 | KT990823 | KT990975 | KT990461 | [5] |
| Rubroboletus dupainii | JAM0607 | — | — | KF030413 | KF030361 | — | [21] |
| R. latisporus | HKAS63517 | — | KP055022 | KP055019 | KP055025 | KP055028 | [25] |
| R. latisporus | HKAS80358 | — | KP055023 | KP055020 | KP055026 | KP055029 | [25] |
| R. rhodosanguineus | 4252 | — | KF030252 | KF030412 | — | — | [21] |
| R. rhodoxanthus | HKAS84879 | — | KT990637 | KT990831 | KT990981 | KT990468 | [5] |
| R. sinicus | HKAS68620 | — | KF112319 | KF112146 | KF112504 | KF112661 | [22] |
| R. sinicus | HKAS56304 | — | KJ605673 | KJ619483 | KJ619482 | KP055031 | [54] |
| Suillellus amygdalinus | 112605ba | — | JQ326996 | JQ327024 | KF030360 | — | [55] |
| S. amygdalinus | NY00035656 (Thiers54483) | — | KT990650 | KT990840 | KT990990 | KT990477 | [5] |
| S. subamygdalinus | HKAS57262 | — | KF112316 | KF112174 | KF112501 | KF112660 | [22] |
| S. subamygdalinus | HKAS53641 | — | KT990651 | KT990841 | KT990991 | KT990478 | [5] |
| S. subamygdalinus | HKAS57953 | — | KT990652 | KT990842 | KT990992 | — | [5] |
| S. subamygdalinus | HKAS74745 | — | KT990653 | KT990843 | KT990993 | KT990479 | [5] |
| S. lacrymibasidiatus | HMJAU60202 (W3194) | OM237315 | OM230174 | OM285117 | OM285113 | OM285115 | this study |
| S. lacrymibasidiatus | HMJAU60203 (W3229) | OM237338 | OM230172 | OM285116 | – | OM285114 | this study |
| Sutorius eximius | REH9400 | — | JQ327004 | JQ327029 | — | — | [55] |
| Su. eximius | HKAS52672 | — | KF112399 | KF112207 | KF112584 | KF112802 | [22] |
| Su. luridiformis | AT1998054 | UDB000658 | – | – | – | – | |
| Tylopilus alpinus | HKAS55438 | — | KF112404 | KF112191 | KF112538 | KF112687 | [22] |
| Ty. argillaceus | HKAS90201 | — | KT990588 | KT990783 | — | — | [5] |
| Ty. argillaceus | HKAS90186 | — | KT990589 | KT990784 | — | KT990424 | [5] |
| Ty. atripurpureus | HKAS50208 | — | KF112472 | KF112283 | KF112620 | KF112799 | [22] |
| Ty. badiceps | MB03-052 | — | KF030336 | — | — | — | [21] |
| Ty. badiceps | 78206 | — | KF030335 | KF030429 | — | — | [21] |
| Veloporphyrellus alpinus | HKAS68301 | — | JX984538 | JX984550 | — | — | [57] |
| V. alpinus | HKAS57490 | — | KF112380 | KF112209 | KF112555 | KF112733 | [22] |
| V. conicus | BZ2408 | — | JX984545 | — | — | — | [57] |
| V. conicus | BZ1670 | — | JX984543 | JX984555 | — | — | [57] |
| V. gracilioides | HKAS53590 | — | KF112381 | KF112210 | KF112556 | KF112734 | [22] |
| Zangia citrina | HKAS52677 | — | HQ326940 | HQ32687 | — | — | [58] |
| Z. citrina | HKAS52684 | — | HQ326941 | HQ326872 | — | — | [58] |
| Z. erythrocephala | HKAS52843 | — | HQ326943 | — | — | — | [58] |
| Z. erythrocephala | HKAS52844 | — | HQ326944 | — | — | — | [58] |
| Z. olivaceobrunnea | HKAS52275 | — | HQ326947 | HQ326875 | — | — | [58] |
| Z. roseola | HKAS75046 | — | KF112414 | KF112269 | KF112579 | KF112791 | [22] |
| S. luridus | IB2004270 | EF644104 | – | – | – | – | [59] |
| S. luridus | 18902 | JF907802 | – | – | – | – | [60] |
| S. luridus | 18182 | JF907793 | – | – | – | – | [60] |
| S. luridus | Blu3 | AY278765 | – | – | – | – | [61] |
| S. luridus | AMB12636 | KC734542 | – | – | – | – | [13] |
| S. luridus | AMB12638 | KC734544 | – | – | – | – | [13] |
| S. luridus | TL-6877 | AJ889930 | – | – | – | – | |
| S. luridus | TL-6877 | UDB000077 | – | – | – | – | |
| S. luridus | 1968 | AY278766 | – | – | – | – | [61] |
| S. luridus | BL2-VII-10 | JQ685714 | – | – | – | – | [61] |
| S. luridus | AT-04 | UDB002401 | – | – | – | – | |
| S. luridus | UP12 | DQ658866 | – | – | – | – | [62] |
| S. luridus | 17696 | JF907789 | – | – | – | – | [60] |
| S. luridus | BL1-VII-09 | JQ685715 | – | – | – | – | |
| S. luridus | MA-Fungi 47706 | AJ419191 | – | – | – | – | [63] |
| S. mendax | AMB12632 | KC734547 | – | – | – | – | [13] |
| S. mendax | AMB12633 | KC734548 | – | – | – | – | [13] |
| S. mendax | AMB12634 | KC734543 | – | – | – | – | [13] |
| S. mendax | AMB12635 | KC734545 | – | – | – | – | [13] |
| S. mendax | AMB12637 | KC734540 | – | – | – | – | [13] |
| S. mendax | AMB12640 | KC734541 | – | – | – | – | [13] |
| Boletus luridus | UF107 | HM347662 | – | – | – | – | |
| B. amygdalinus | src491 | DQ974705 | – | – | – | – | [64] |
| B. comptus | 17827 | JF907791 | – | – | – | – | [60] |
| B. comptus | AMB12639 | KC734539 | – | – | – | – | [13] |
| B. queletii | 17196 | JF907784 | – | – | – | – | [60] |
| B. queletii | 17208 | JF907785 | – | – | – | – | [60] |
| B. queletii | AMB12641 | KC734546 | – | – | – | – | [13] |
| B. queletii | JV01-231 | UDB000760 | – | – | – | – | |
| N. erythropus | MA-Fungi 47702 | AJ419188 | – | – | – | – | [63] |
| N. erythropus | BOER_TO_2 (AAM630/06) | FM958177 | – | – | – | – | |
| N. erythropus | UF278 | HM347644 | – | – | – | – | |
| N. erythropus | UF276 | HM347643 | – | – | – | – | |
| N. erythropus | UF269 | HM347665 | – | – | – | – | |
| N. erythropus | DG05-54 | UDB001523 | – | – | – | – | |
| N. erythropus | SU46 | DQ131633 | – | – | – | – | [65] |
| N. erythropus | SU47 | DQ131634 | – | – | – | – | [65] |
| N. erythropus | Daniels 582 | AJ496595 | – | – | – | – | [63] |
| Caloboletus calopus | AT1998059 | UDB000659 | – | – | – | – | |
| Ca. radicans | TUF106003 | UDB003224 | – | – | – | – | |
| Bu. fuscoroseus | AH96025 | UDB000649 | – | – | – | – | |
| Bu. fuscoroseus | AT1996017 | UDB000652 | – | – | – | – | |
| Bu. fechtneri | AT2003097 | UDB000703 | – | – | – | – | [21] |
| Imperator rhodopurpureus | AT1996058 | UDB000654 | – | – | – | – | |
| R. pulchrotinctus | GS0860 | UDB000407 | – | – | – | – | |
| R. satanas | AT1998051 | UDB000415 | – | – | – | – | |
| R. rubrosanguineus | GS0405 | UDB000410 | – | – | – | – | |
| R. rhodoxanthus | AT2000182 | UDB001116 | – | – | – | – | |
| Cyanoboletus pulverulentus | RT00004 | EU819502 | – | – | – | – | |
| Cyanoboletus pulverulentus | AH97030 | UDB000408 | – | – | – | – | [66] |
| B. aestivalis | AT2004040 | UDB001113 | – | – | – | – | |
| B. aereus | AT2000198 | UDB000943 | – | – | – | – |
| Species | Pileus | Context | Hymenophore | Stipe | Spores | Cystidia |
|---|---|---|---|---|---|---|
| Butyriboletus huangnianlaii | Surface dry, finely tomentose, brown to reddish brown | Yellowish to yellow, changing blue quickly when injured | Adnate or slightly depressed, changing blue quickly when injured | Stipitipellis, fertile hymeniform, fusiform, or subfusiform terminal cells | (7.0) 7.5–10.5 (11.0) × 3.0–4.0 μm, olive-brown to yellowish brown | Fusiform or subfusiform |
| Bu. pseudospeciosus | Purplish tint, staining dark blue quickly when bruised | Yellowish, staining blue to grayish blue promptly when injured | Adnate, rapidly bluing when bruised | Stipitipellis consisting of tufts of lageniform caulocystidia | 9.0–11.0 (12.0) × 3.5–4.0 μm | Narrowly lageniform to lageniform |
| Bu. roseoflavus | Pinkish to purplish red or rose-red | Yellowish or light yellow, turning blue slowly or unchanging when bruised | Adnate, staining blue quickly when hurt | Stipe trama composed of parallel hyphae | 9.0–12.0 (13.0) × 3.0–4.0 μm | Narrowly lageniform to lageniform |
| Bu. sanicibus | Dull brown | Pale yellow, usually turning blue when cut | Depressed, bruising blue | – | 11.0–15.0 × 4.0–5.0 μm | Fusoid-ventricose |
| Bu. subregius | Pastel pink | Yellowish green, turning blue when cut | weakly decurrent, covered with a layer of whitish mycelium when young, surface yellowish green | Stipitipellis fertile, hymeniform, caulocystidia narrowly lageniform, caulobasidia subclavate, with yellowish intracellular pigments. | (10.0) 11.1–11.5 (13.0) × (3.0) 4.0–4.2 (5.0) μm | narrowly lageniform |
| Bu. yicibus | Covered with fibrillose squamules, ochreous, brown to dark brown | Nearly white, staining light blue very slowly when injured | Adnate, degrading bluish slowly when injured | Stipitipellis consisting of tufts of lageniform caulocystidia | (11.0) 13.0–15.0 (16.0) × 4.0–5.0 (5.5) μm | Narrowly lageniform to lageniform |
| Bu. pseudoroseoflavus | Tomentose, pink to greyish rose | Light yellow, unchanging in color when injured. | Adnate to decurrent, staining blue when bruised | Stipitipellis hymeniform, with terminal inflated cells | (7.0) 10.2–11.0 (16.0) × (2.0) 3.1–3.7 (4.0) μm | Narrowly lageniform |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Tuo, Y.-L.; Wu, D.-M.; Gao, N.; Zhang, Z.-H.; Rao, G.; Wang, X.-M.; Wang, J.; Dai, D.; Li, Y.; et al. Exploring the Relationships between Four New Species of Boletoid Fungi from Northern China and Their Related Species. J. Fungi 2022, 8, 218. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8030218
Wang Y, Tuo Y-L, Wu D-M, Gao N, Zhang Z-H, Rao G, Wang X-M, Wang J, Dai D, Li Y, et al. Exploring the Relationships between Four New Species of Boletoid Fungi from Northern China and Their Related Species. Journal of Fungi. 2022; 8(3):218. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8030218
Chicago/Turabian StyleWang, Yang, Yong-Lan Tuo, Dong-Mei Wu, Neng Gao, Zhen-Hao Zhang, Gu Rao, Xiao-Min Wang, Jing Wang, Dan Dai, Yu Li, and et al. 2022. "Exploring the Relationships between Four New Species of Boletoid Fungi from Northern China and Their Related Species" Journal of Fungi 8, no. 3: 218. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8030218