
Secretome Profiling by Proteogenomic Analysis Shows Species-Specific, Temperature-Dependent, and Putative Virulence Proteins of Pythium insidiosum

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism and Protein Extraction
2.2. Protein Digestion and Liquid Chromatography–Tandem Mass Spectrometry
2.3. Proteomic Data Analysis
2.4. Bioinformatic Analysis
2.5. Data Availability
3. Results
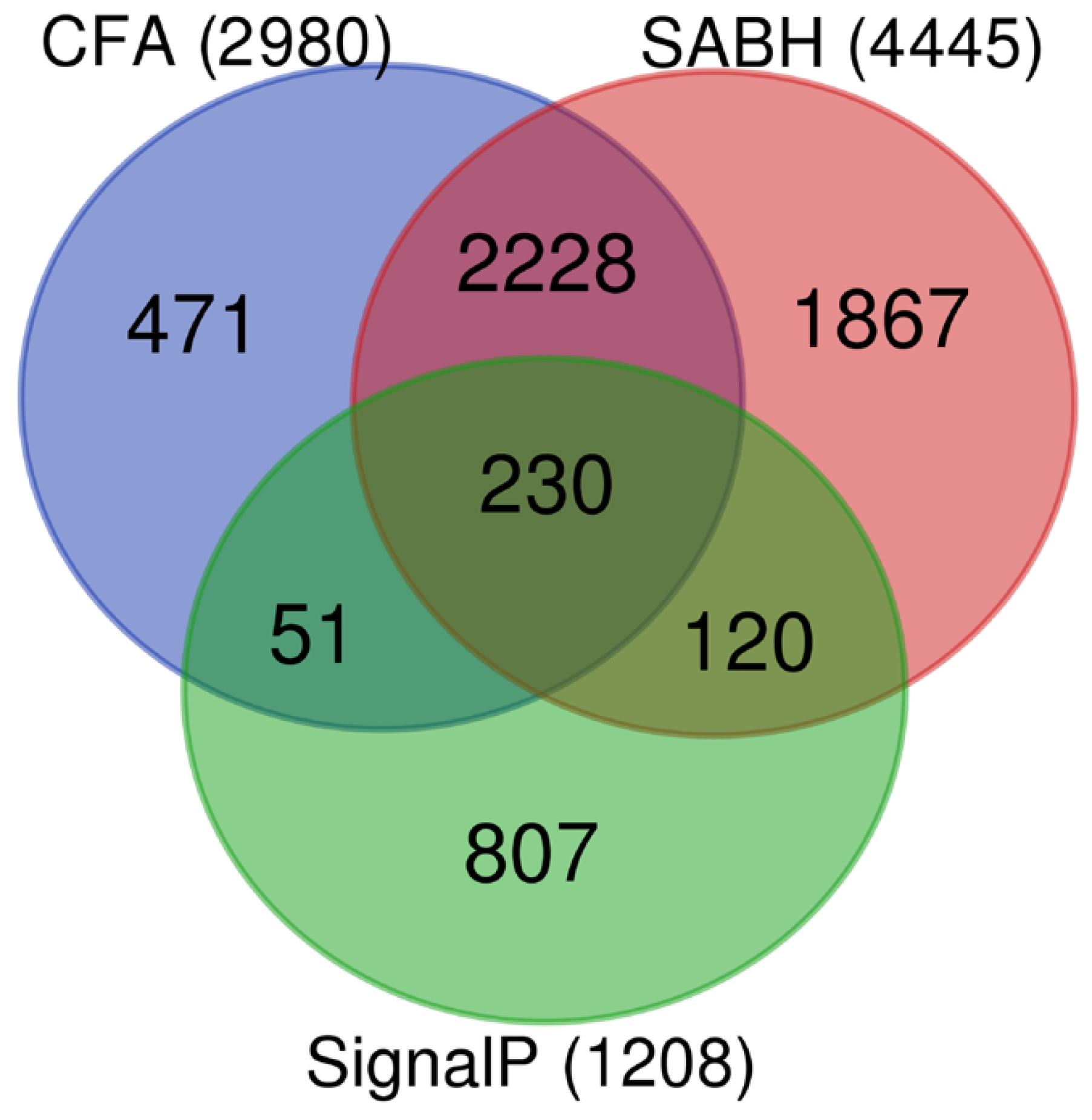
3.1. Identification and Validation of P. insidiosum Secretome
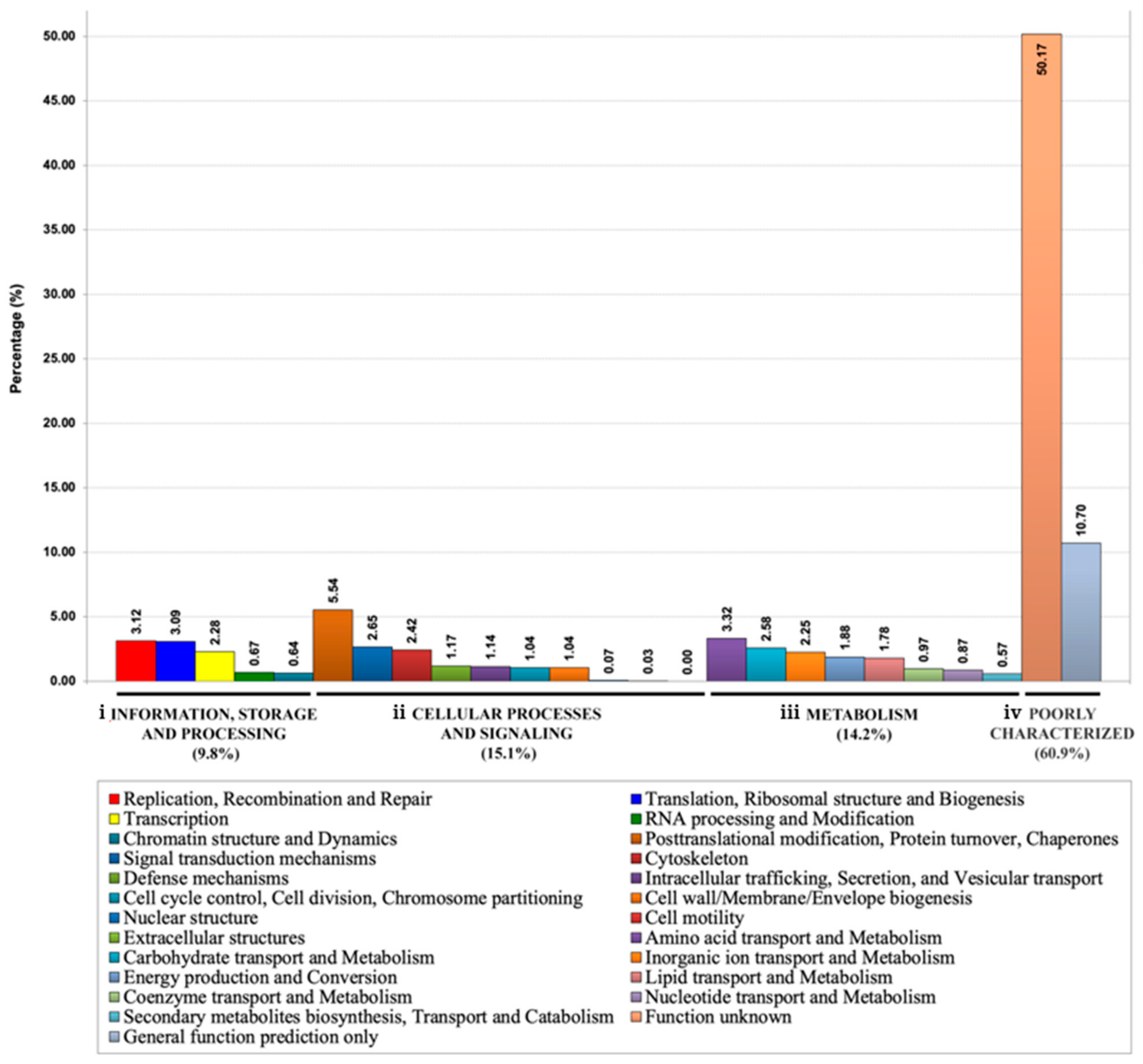
3.2. Functional Annotation of P. insidiosum’s Secretome
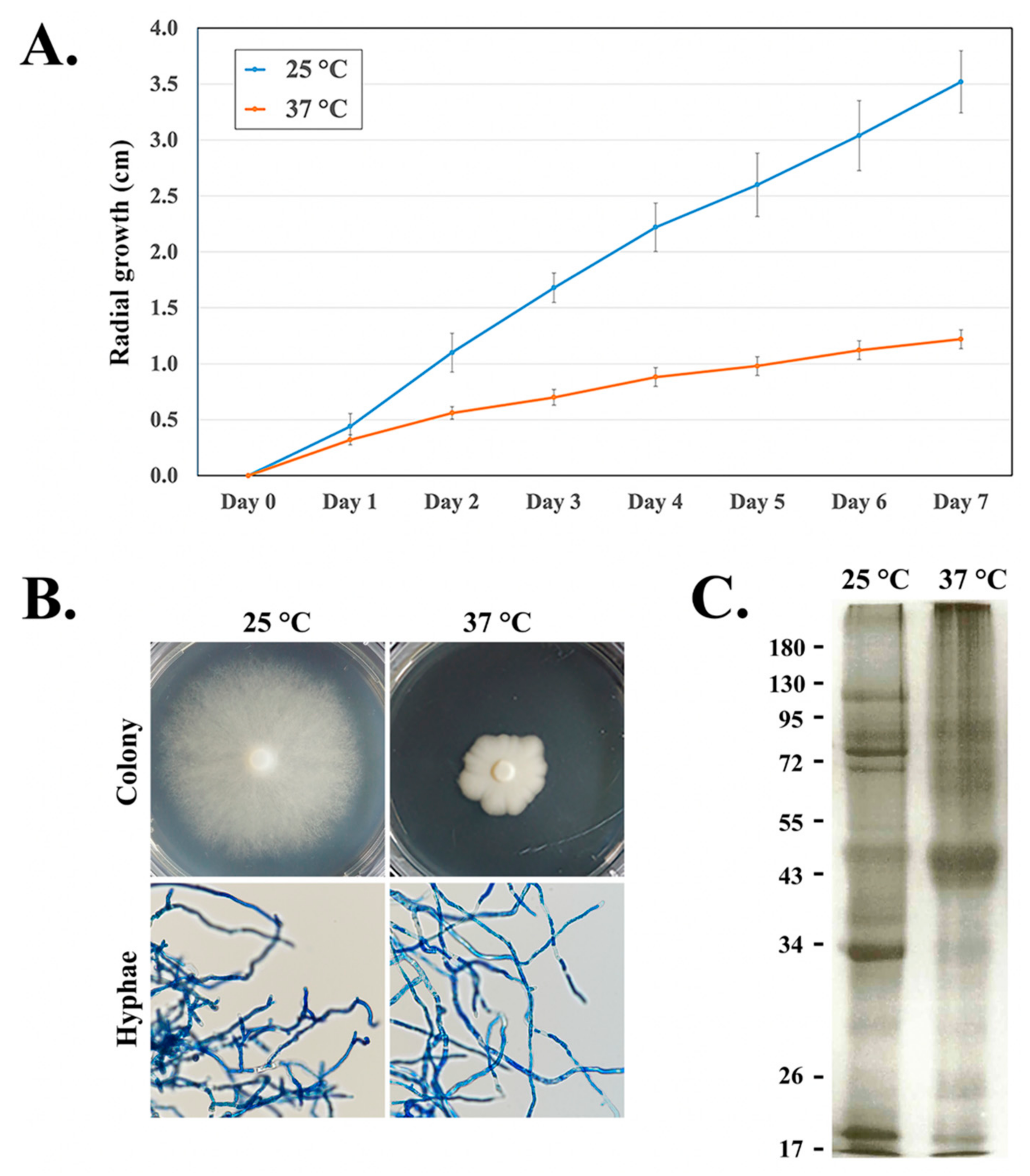
3.3. Effect of the Temperature on the Growth of the Organism and Expression of the Secretory/Extracellular Proteins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
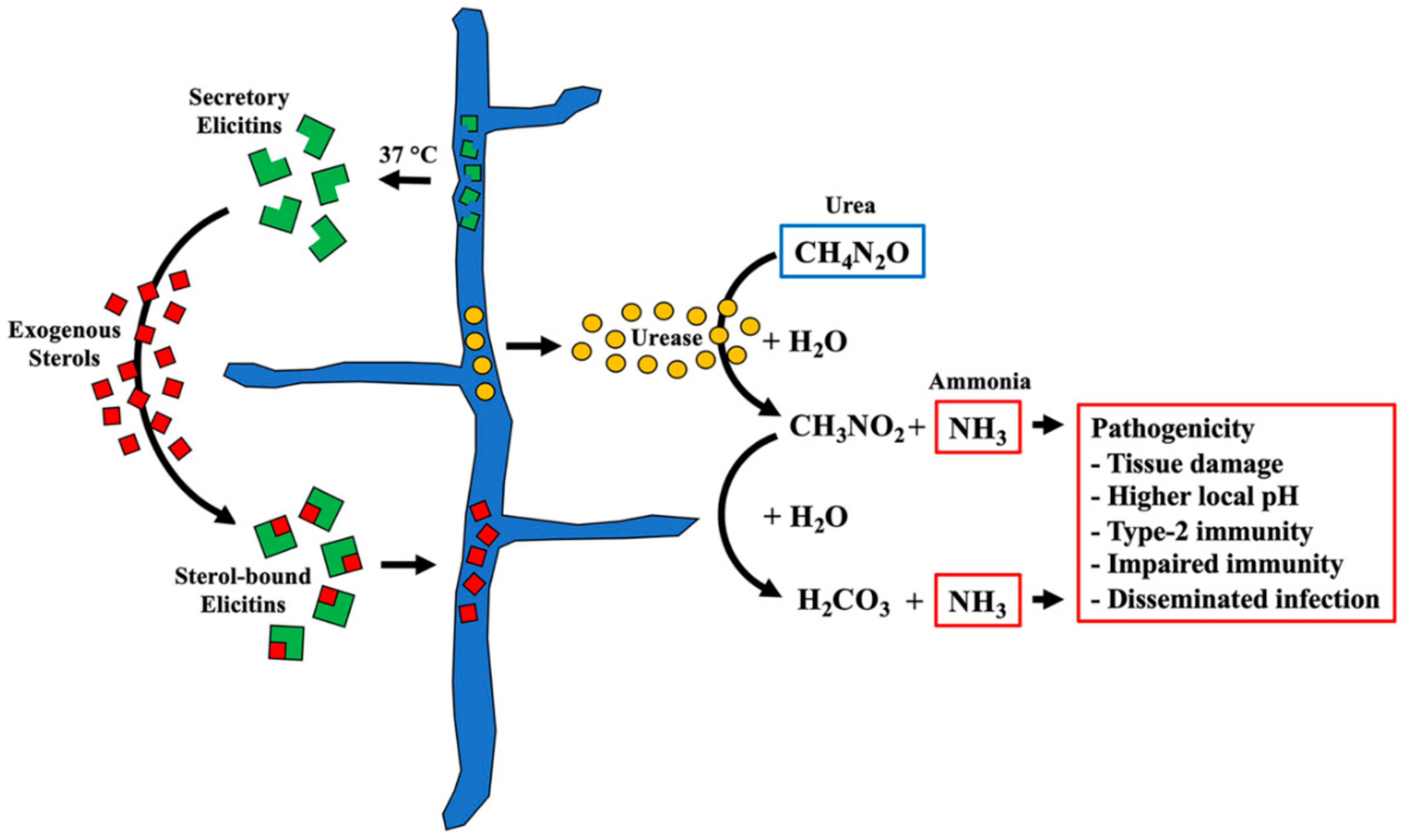{kind=link}
| P. insidiosum Protein ID | Functional Annotation a | Average Protein Abundance at 37 °C (emPAI b) | Average Protein Abundance at 25 °C (emPAI) | Relative Protein Abundance (37:25 °C) | Accession Number |
|---|---|---|---|---|---|
| PINS01640001A | Cyclophilin A | 50.18 | - | - | GAY01457.1 |
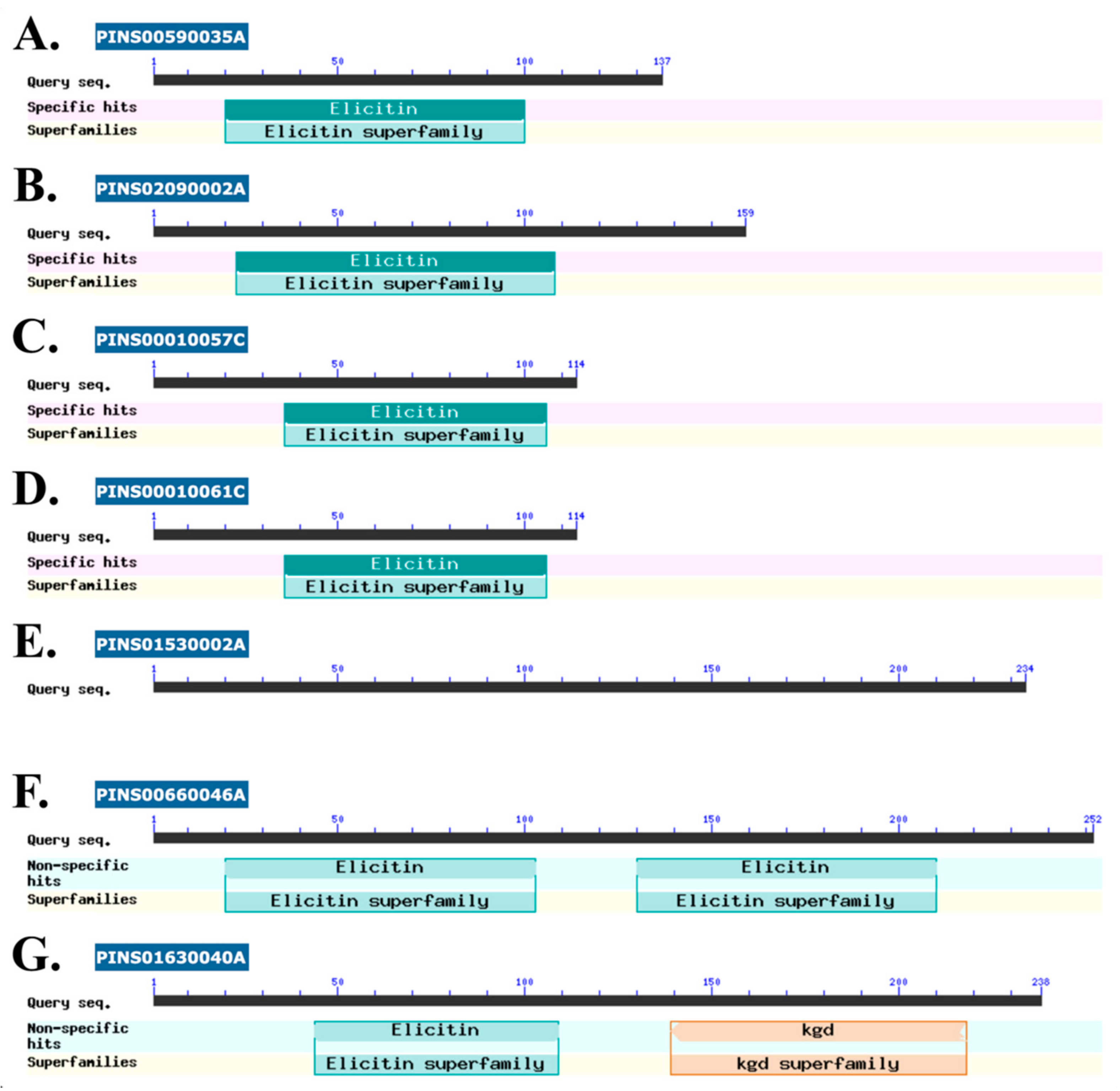
| PINS00590035A | Elicitin-like protein c,d | 9.94 | - | - | GAX97068.1 |
| PINS00010057C | Hypothetical protein c,d | 6.69 | - | - | GAX92118.1 |
| PINS00010061C | Hypothetical protein c,d | 3.50 | - | - | GAX92125.1 |
| PINS01530002A | Hypothetical protein d | 1.79 | - | - | GAY01151.1 |
| PINS00660046A | Hypothetical protein c,d | 1.69 | - | - | GAX97463.1 |
| PINS00380039A | ATP synthase subunit beta, mitochondrial | 1.44 | - | - | GAX95749.1 |
| PINS00660004A | Glycerophosphoryl diester phosphodiesterase | 2.34 | 0.22 | 10.61 | GAX97472.1 |
| PINS02090002A | Elicitin-like protein c,d | 71.97 | 7.82 | 9.21 | GAY02672.1 |
| PINS01630040A | Hypothetical protein c,d | 1.27 | 0.15 | 8.43 | GAY01431.1 |
| PINS02430017C | Cofilin/tropomyosin-type actin-binding protein | 0.11 | 0.02 | 7.33 | GAY03393.1 |
| PINS00560010C | Cofilin/tropomyosin-type actin-binding protein | 0.14 | 0.02 | 7.00 | GAX96878.1 |
| PINS00420001A | Malate dehydrogenase, NAD-dependent | 2.00 | 0.30 | 6.76 | GAX96042.1 |
| PINS06030002B | Glycoside hydrolase d | 0.11 | 0.02 | 5.25 | GAY06261.1 |
3.4. Putative Pathogenicity Proteins in P. insidiosum’s Secretome
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gaastra, W.; Lipman, L.J.; De Cock, A.W.; Exel, T.K.; Pegge, R.B.; Scheurwater, J.; Vilela, R.; Mendoza, L. Pythium insidiosum: An Overview. Vet. Microbiol. 2010, 146, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza, L.; Ajello, L.; McGinnis, M.R. Infection Caused by the Oomycetous Pathogen Pythium insidiosum. J. Mycol. Med. 1996, 6, 151–164. [Google Scholar]
- Krajaejun, T.; Sathapatayavongs, B.; Pracharktam, R.; Nitiyanant, P.; Leelachaikul, P.; Wanachiwanawin, W.; Chaiprasert, A.; Assanasen, P.; Saipetch, M.; Mootsikapun, P.; et al. Clinical and Epidemiological Analyses of Human Pythiosis in Thailand. Clin. Infect. Dis. 2006, 43, 569–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yolanda, H.; Krajaejun, T. Global Distribution and Clinical Features of Pythiosis in Humans and Animals. J. Fungi 2022, 8, 182. [Google Scholar] [CrossRef]
- Mendoza, L.; Hernandez, F.; Ajello, L. Life Cycle of the Human and Animal Oomycete Pathogen Pythium insidiosum. J. Clin. Microbiol. 1993, 31, 2967–2973. [Google Scholar] [CrossRef] [Green Version]
- Vanittanakom, N.; Szekely, J.; Khanthawong, S.; Sawutdeechaikul, P.; Vanittanakom, P.; Fisher, M.C. Molecular Detection of Pythium insidiosum from Soil in Thai Agricultural Areas. Int. J. Med. Microbiol. 2014, 304, 321–326. [Google Scholar] [CrossRef]
- Supabandhu, J.; Fisher, M.C.; Mendoza, L.; Vanittanakom, N. Isolation and Identification of the Human Pathogen Pythium insidiosum from Environmental Samples Collected in Thai Agricultural Areas. Med. Mycol. 2008, 46, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Mar Htun, Z.; Laikul, A.; Pathomsakulwong, W.; Yurayart, C.; Lohnoo, T.; Yingyong, W.; Kumsang, Y.; Payattikul, P.; Sae-Chew, P.; Rujirawat, T.; et al. Identification and Biotyping of Pythium insidiosum Isolated from Urban and Rural Areas of Thailand by Multiplex PCR, DNA Barcode, and Proteomic Analyses. J. Fungi 2021, 7, 242. [Google Scholar] [CrossRef]
- Keeratijarut, A.; Lohnoo, T.; Yingyong, W.; Sriwanichrak, K.; Krajaejun, T. A Peptide ELISA to Detect Antibodies against Pythium insidiosum Based on Predicted Antigenic Determinants of Exo-1,3-Beta-Glucanase. Southeast Asian J. Trop. Med. Public Health 2013, 44, 672–680. [Google Scholar]
- Chareonsirisuthigul, T.; Khositnithikul, R.; Intaramat, A.; Inkomlue, R.; Sriwanichrak, K.; Piromsontikorn, S.; Kitiwanwanich, S.; Lowhnoo, T.; Yingyong, W.; Chaiprasert, A.; et al. Performance Comparison of Immunodiffusion, Enzyme-Linked Immunosorbent Assay, Immunochromatography and Hemagglutination for Serodiagnosis of Human Pythiosis. Diagn. Microbiol. Infect. Dis. 2013, 76, 42–45. [Google Scholar] [CrossRef]
- Inkomlue, R.; Larbcharoensub, N.; Karnsombut, P.; Lerksuthirat, T.; Aroonroch, R.; Lohnoo, T.; Yingyong, W.; Santanirand, P.; Sansopha, L.; Krajaejun, T. Development of an Anti-Elicitin Antibody-Based Immunohistochemical Assay for Diagnosis of Pythiosis. J. Clin. Microbiol. 2016, 54, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Keeratijarut, A.; Lohnoo, T.; Yingyong, W.; Rujirawat, T.; Srichunrusami, C.; Onpeaw, P.; Chongtrakool, P.; Brandhorst, T.T.; Krajaejun, T. Detection of the Oomycete Pythium insidiosum by Real-Time PCR Targeting the Gene Coding for Exo-1,3-β-Glucanase. J. Med. Microbiol. 2015, 64, 971–977. [Google Scholar] [CrossRef]
- Keeratijarut, A.; Lohnoo, T.; Yingyong, W.; Nampoon, U.; Lerksuthirat, T.; Onpaew, P.; Chongtrakool, P.; Krajaejun, T. PCR Amplification of a Putative Gene for Exo-1,3-Beta-Glucanase to Identify the Pathogenic Oomycete Pythium insidiosum. Asian Biomed. 2014, 8, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Rujirawat, T.; Sridapan, T.; Lohnoo, T.; Yingyong, W.; Kumsang, Y.; Sae-Chew, P.; Tonpitak, W.; Krajaejun, T. Single Nucleotide Polymorphism-Based Multiplex PCR for Identification and Genotyping of the Oomycete Pythium insidiosum from Humans, Animals and the Environment. Infect. Genet. Evol. 2017, 54, 429–436. [Google Scholar] [CrossRef]
- Intaramat, A.; Sornprachum, T.; Chantrathonkul, B.; Chaisuriya, P.; Lohnoo, T.; Yingyong, W.; Jongruja, N.; Kumsang, Y.; Sandee, A.; Chaiprasert, A.; et al. Protein A/G-Based Immunochromatographic Test for Serodiagnosis of Pythiosis in Human and Animal Subjects from Asia and Americas. Med. Mycol. 2016, 54, 641–647. [Google Scholar] [CrossRef] [Green Version]
- Jaturapaktrarak, C.; Payattikul, P.; Lohnoo, T.; Kumsang, Y.; Laikul, A.; Pathomsakulwong, W.; Yurayart, C.; Tonpitak, W.; Krajaejun, T. Protein A/G-Based Enzyme-Linked Immunosorbent Assay for Detection of Anti-Pythium insidiosum Antibodies in Human and Animal Subjects. BMC Res. Notes 2020, 13, 135. [Google Scholar] [CrossRef] [Green Version]
- Krajaejun, T.; Lohnoo, T.; Jittorntam, P.; Srimongkol, A.; Kumsang, Y.; Yingyong, W.; Rujirawat, T.; Reamtong, O.; Mangmee, S. Assessment of Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry for Identification and Biotyping of the Pathogenic Oomycete Pythium insidiosum. Int. J. Infect. Dis. 2018, 77, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Chaiprasert, A.; Samerpitak, K.; Wanachiwanawin, W.; Thasnakorn, P. Induction of Zoospore Formation in Thai Isolates of Pythium insidiosum. Mycoses 1990, 33, 317–323. [Google Scholar] [CrossRef]
- Chitasombat, M.N.; Jongkhajornpong, P.; Lekhanont, K.; Krajaejun, T. Recent Update in Diagnosis and Treatment of Human Pythiosis. PeerJ 2020, 8, e8555. [Google Scholar] [CrossRef] [Green Version]
- Grooters, A.M.; Gee, M.K. Development of a Nested Polymerase Chain Reaction Assay for the Detection and Identification of Pythium insidiosum. J. Vet. Intern. Med. 2002, 16, 147–152. [Google Scholar] [CrossRef]
- Thongsri, Y.; Wonglakorn, L.; Chaiprasert, A.; Svobodova, L.; Hamal, P.; Pakarasang, M.; Prariyachatigul, C. Evaluation for the Clinical Diagnosis of Pythium insidiosum Using a Single-Tube Nested PCR. Mycopathologia 2013, 176, 369–376. [Google Scholar] [CrossRef] [Green Version]
- Pracharktam, R.; Changtrakool, P.; Sathapatayavongs, B.; Jayanetra, P.; Ajello, L. Immunodiffusion Test for Diagnosis and Monitoring of Human Pythiosis Insidiosi. J. Clin. Microbiol. 1991, 29, 2661–2662. [Google Scholar] [CrossRef] [Green Version]
- Yolanda, H.; Krajaejun, T. Review of Methods and Antimicrobial Agents for Susceptibility Testing against Pythium insidiosum. Heliyon 2020, 6, e03737. [Google Scholar] [CrossRef]
- Yolanda, H.; Krajaejun, T. History and Perspective of Immunotherapy for Pythiosis. Vaccines 2021, 9, 1080. [Google Scholar] [CrossRef]
- Klein, B.S. Molecular Basis of Pathogenicity in Blastomyces Dermatitidis: The Importance of Adhesion. Curr. Opin. Microbiol. 2000, 3, 339–343. [Google Scholar] [CrossRef]
- Brandhorst, T.; Wüthrich, M.; Finkel-Jimenez, B.; Klein, B. AC-Terminal EGF-like Domain Governs BAD1 Localization to the Yeast Surface and Fungal Adherence to Phagocytes, but Is Dispensable in Immune Modulation and Pathogenicity of Blastomyces Dermatitidis. Mol. Microbiol. 2003, 48, 53–65. [Google Scholar] [CrossRef]
- Geddes, J.M.H.; Croll, D.; Caza, M.; Stoynov, N.; Foster, L.J.; Kronstad, J.W. Secretome Profiling of Cryptococcus Neoformans Reveals Regulation of a Subset of Virulence-Associated Proteins and Potential Biomarkers by Protein Kinase A. BMC Microbiol. 2015, 15, 206. [Google Scholar] [CrossRef] [Green Version]
- Cadieux, B.; Lian, T.; Hu, G.; Wang, J.; Biondo, C.; Teti, G.; Liu, V.; Murphy, M.E.P.; Creagh, A.L.; Kronstad, J.W. The Mannoprotein Cig1 Supports Iron Acquisition from Heme and Virulence in the Pathogenic Fungus Cryptococcus Neoformans. J. Infect. Dis. 2013, 207, 1339–1347. [Google Scholar] [CrossRef] [Green Version]
- Kamoun, S. A Catalogue of the Effector Secretome of Plant Pathogenic Oomycetes. Annu. Rev. Phytopathol. 2006, 44, 41–60. [Google Scholar] [CrossRef] [Green Version]
- Severino, V.; Farina, A.; Fleischmann, F.; Dalio, R.J.D.; Di Maro, A.; Scognamiglio, M.; Fiorentino, A.; Parente, A.; Osswald, W.; Chambery, A. Molecular Profiling of the Phytophthora Plurivora Secretome: A Step towards Understanding the Cross-Talk between Plant Pathogenic Oomycetes and Their Hosts. PLoS ONE 2014, 9, e112317. [Google Scholar] [CrossRef] [Green Version]
- Mateos, F.V.; Rickauer, M.; Esquerré-Tugayé, M.T. Cloning and Characterization of a CDNA Encoding an Elicitor of Phytophthora Parasitica Var. Nicotianae That Shows Cellulose-Binding and Lectin-like Activities. Mol. Plant Microbe Interact. 1997, 10, 1045–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meijer, H.J.G.; van de Vondervoort, P.J.I.; Yin, Q.Y.; de Koster, C.G.; Klis, F.M.; Govers, F.; de Groot, P.W.J. Identification of Cell Wall-Associated Proteins from Phytophthora Ramorum. Mol. Plant Microbe Interact. 2006, 19, 1348–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grenville-Briggs, L.J.; Avrova, A.O.; Hay, R.J.; Bruce, C.R.; Whisson, S.C.; van West, P. Identification of Appressorial and Mycelial Cell Wall Proteins and a Survey of the Membrane Proteome of Phytophthora Infestans. Fungal Biol. 2010, 114, 702–723. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.J.; Lanter, K.; Makselan, S.; Bonati, C.; Asbrock, P.; Ravishankar, J.P.; Money, N.P. Relationship between Temperature Optima and Secreted Protease Activities of Three Pythium Species and Pathogenicity toward Plant and Animal Hosts. Mycol. Res. 2006, 110, 96–103. [Google Scholar] [CrossRef] [PubMed]
- van der Plaats-Niterink, A.J. Monograph of the Genus Pythium. Stud. Mycol. 1981, 21, 1–242. [Google Scholar]
- Kamoun, S. Molecular Genetics of Pathogenic Oomycetes. Eukaryot. Cell 2003, 2, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Krajaejun, T.; Khositnithikul, R.; Lerksuthirat, T.; Lowhnoo, T.; Rujirawat, T.; Petchthong, T.; Yingyong, W.; Suriyaphol, P.; Smittipat, N.; Juthayothin, T.; et al. Expressed Sequence Tags Reveal Genetic Diversity and Putative Virulence Factors of the Pathogenic Oomycete Pythium insidiosum. Fungal Biol. 2011, 115, 683–696. [Google Scholar] [CrossRef]
- Krajaejun, T.; Lerksuthirat, T.; Garg, G.; Lowhnoo, T.; Yingyong, W.; Khositnithikul, R.; Tangphatsornruang, S.; Suriyaphol, P.; Ranganathan, S.; Sullivan, T.D. Transcriptome Analysis Reveals Pathogenicity and Evolutionary History of the Pathogenic Oomycete Pythium insidiosum. Fungal Biol. 2014, 118, 640–653. [Google Scholar] [CrossRef]
- Rujirawat, T.; Patumcharoenpol, P.; Lohnoo, T.; Yingyong, W.; Lerksuthirat, T.; Tangphatsornruang, S.; Suriyaphol, P.; Grenville-Briggs, L.J.; Garg, G.; Kittichotirat, W.; et al. Draft Genome Sequence of the Pathogenic Oomycete Pythium insidiosum Strain Pi-S, Isolated from a Patient with Pythiosis. Genome Announc. 2015, 3, e00574-15. [Google Scholar] [CrossRef] [Green Version]
- Kittichotirat, W.; Patumcharoenpol, P.; Rujirawat, T.; Lohnoo, T.; Yingyong, W.; Krajaejun, T. Draft Genome and Sequence Variant Data of the Oomycete Pythium insidiosum Strain Pi45 from the Phylogenetically-Distinct Clade-III. Data Brief 2017, 15, 896–900. [Google Scholar] [CrossRef]
- Krajaejun, T.; Kittichotirat, W.; Patumcharoenpol, P.; Rujirawat, T.; Lohnoo, T.; Yingyong, W. Data on Whole Genome Sequencing of the Oomycete Pythium insidiosum Strain CBS 101555 from a Horse with Pythiosis in Brazil. BMC Res. Notes 2018, 11, 880. [Google Scholar] [CrossRef] [Green Version]
- Krajaejun, T.; Kittichotirat, W.; Patumcharoenpol, P.; Rujirawat, T.; Lohnoo, T.; Yingyong, W. Draft Genome Sequence of the Oomycete Pythium Destruens Strain ATCC 64221 from a Horse with Pythiosis in Australia. BMC Res. Notes 2020, 13, 329. [Google Scholar] [CrossRef]
- Patumcharoenpol, P.; Rujirawat, T.; Lohnoo, T.; Yingyong, W.; Vanittanakom, N.; Kittichotirat, W.; Krajaejun, T. Draft Genome Sequences of the Oomycete Pythium insidiosum Strain CBS 573.85 from a Horse with Pythiosis and Strain CR02 from the Environment. Data Brief 2018, 16, 47–50. [Google Scholar] [CrossRef]
- Krajaejun, T.; Kunakorn, M.; Pracharktam, R.; Chongtrakool, P.; Sathapatayavongs, B.; Chaiprasert, A.; Vanittanakom, N.; Chindamporn, A.; Mootsikapun, P. Identification of a Novel 74-KiloDalton Immunodominant Antigen of Pythium insidiosum Recognized by Sera from Human Patients with Pythiosis. J. Clin. Microbiol. 2006, 44, 1674–1680. [Google Scholar] [CrossRef] [Green Version]
- Rujirawat, T.; Patumcharoenpol, P.; Lohnoo, T.; Yingyong, W.; Kumsang, Y.; Payattikul, P.; Tangphatsornruang, S.; Suriyaphol, P.; Reamtong, O.; Garg, G.; et al. Probing the Phylogenomics and Putative Pathogenicity Genes of Pythium insidiosum by Oomycete Genome Analyses. Sci. Rep. 2018, 8, 4135. [Google Scholar] [CrossRef] [Green Version]
- Ishihama, Y.; Oda, Y.; Tabata, T.; Sato, T.; Nagasu, T.; Rappsilber, J.; Mann, M. Exponentially Modified Protein Abundance Index (EmPAI) for Estimation of Absolute Protein Amount in Proteomics by the Number of Sequenced Peptides per Protein. Mol. Cell Proteom. 2005, 4, 1265–1272. [Google Scholar] [CrossRef] [Green Version]
- Dowle, A.A.; Wilson, J.; Thomas, J.R. Comparing the Diagnostic Classification Accuracy of ITRAQ, Peak-Area, Spectral-Counting, and EmPAI Methods for Relative Quantification in Expression Proteomics. J. Proteome Res. 2016, 15, 3550–3562. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating Signal Peptides from Transmembrane Regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Zhou, C.; Smith, J.; Lam, M.; Zemla, A.; Dyer, M.; Slezak, T. MvirDB—A Microbial Database of Protein Toxins, Virulence Factors and Antibiotic Resistance Genes for Bio-Defence Applications. Nucleic Acids Res. 2007, 35, D391–D394. [Google Scholar] [CrossRef]
- Tatusov, R.L.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Kiryutin, B.; Koonin, E.V.; Krylov, D.M.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; et al. The COG Database: An Updated Version Includes Eukaryotes. BMC Bioinform. 2003, 4, 41. [Google Scholar] [CrossRef] [Green Version]
- Galperin, M.Y.; Makarova, K.S.; Wolf, Y.I.; Koonin, E.V. Expanded Microbial Genome Coverage and Improved Protein Family Annotation in the COG Database. Nucleic Acids Res. 2015, 43, D261–D269. [Google Scholar] [CrossRef]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A Web-Based Tool for the Analysis of Sets through Venn Diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; et al. CDD: Specific Functional Annotation with the Conserved Domain Database. Nucleic Acids Res. 2009, 37, D205–D210. [Google Scholar] [CrossRef] [Green Version]
- Marchler-Bauer, A.; Lu, S.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; et al. CDD: A Conserved Domain Database for the Functional Annotation of Proteins. Nucleic Acids Res. 2011, 39, D225–D229. [Google Scholar] [CrossRef] [Green Version]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE Database Resources in 2022: A Hub for Mass Spectrometry-Based Proteomics Evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]
- Rujirawat, T.; Patumcharoenpol, P.; Kittichotirat, W.; Krajaejun, T. Oomycete Gene Table: An Online Database for Comparative Genomic Analyses of the Oomycete Microorganisms. Database 2019, 2019, baz082. [Google Scholar] [CrossRef]
- Nickel, W.; Seedorf, M. Unconventional Mechanisms of Protein Transport to the Cell Surface of Eukaryotic Cells. Annu. Rev. Cell Dev. Biol. 2008, 24, 287–308. [Google Scholar] [CrossRef]
- Sae-Chew, P.; Rujirawat, T.; Kumsang, Y.; Payattikul, P.; Lohnoo, T.; Yingyong, W.; Jaturapaktrarak, C.; Rotchanapreeda, T.; Reamtong, O.; Srisuk, T.; et al. Automated Cell-Free Multiprotein Synthesis Facilitates the Identification of a Secretory, Oligopeptide Elicitor-Like, Immunoreactive Protein of the Oomycete Pythium insidiosum. mSystems 2020, 5, e00196-20. [Google Scholar] [CrossRef] [PubMed]
- De Cock, A.W.; Mendoza, L.; Padhye, A.A.; Ajello, L.; Kaufman, L. Pythium insidiosum sp. Nov., the Etiologic Agent of Pythiosis. J. Clin. Microbiol. 1987, 25, 344–349. [Google Scholar] [CrossRef] [Green Version]
- Krajaejun, T.; Chongtrakool, P.; Angkananukul, K.; Brandhorst, T.T. Effect of Temperature on Growth of the Pathogenic Oomycete Pythium insidiosum. Southeast Asian J. Trop. Med. Public Health 2010, 41, 1462–1466. [Google Scholar]
- Calvano, T.P.; Blatz, P.J.; Vento, T.J.; Wickes, B.L.; Sutton, D.A.; Thompson, E.H.; White, C.E.; Renz, E.M.; Hospenthal, D.R. Pythium Aphanidermatum Infection Following Combat Trauma. J. Clin. Microbiol. 2011, 49, 3710–3713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farmer, A.R.; Murray, C.K.; Driscoll, I.R.; Wickes, B.L.; Wiederhold, N.; Sutton, D.A.; Sanders, C.; Mende, K.; Enniss, B.; Feig, J.; et al. Combat-Related Pythium Aphanidermatum Invasive Wound Infection: Case Report and Discussion of Utility of Molecular Diagnostics. J. Clin. Microbiol. 2015, 53, 1968–1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.; Winter, M.; Zouhar, M.; Ryšánek, P. Cyclophilins: Less Studied Proteins with Critical Roles in Pathogenesis. Phytopathology 2018, 108, 6–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bzdyl, N. Characterisation of Cyclophilin Proteins from Burkholderia Pseudomallei and Their Role in Pathogenesis. Ph.D. Thesis, The University of Western Australia, Perth, Australia, 1 November 2021. [Google Scholar] [CrossRef]
- Gan, P.H.P.; Shan, W.; Blackman, L.M.; Hardham, A.R. Characterization of Cyclophilin-Encoding Genes in Phytophthora. Mol. Genet. Genom. 2009, 281, 565. [Google Scholar] [CrossRef]
- Jiang, R.H.Y.; Tyler, B.M.; Whisson, S.C.; Hardham, A.R.; Govers, F. Ancient Origin of Elicitin Gene Clusters in Phytophthora Genomes. Mol. Biol. Evol. 2006, 23, 338–351. [Google Scholar] [CrossRef] [Green Version]
- PanabiÈRes, F.; Ponchet, M.; Allasia, V.; Cardin, L.; Ricci, P. Characterization of Border Species among Pythiaceae: Several Pythium Isolates Produce Elicitins, Typical Proteins from Phytophthora spp. Mycol. Res. 1997, 101, 1459–1468. [Google Scholar] [CrossRef]
- Nürnberger, T.; Brunner, F.; Kemmerling, B.; Piater, L. Innate Immunity in Plants and Animals: Striking Similarities and Obvious Differences. Immunol. Rev. 2004, 198, 249–266. [Google Scholar] [CrossRef]
- Gaulin, E.; Bottin, A.; Dumas, B. Sterol Biosynthesis in Oomycete Pathogens. Plant Signal. Behav. 2010, 5, 258–260. [Google Scholar] [CrossRef] [Green Version]
- Madoui, M.-A.; Bertrand-Michel, J.; Gaulin, E.; Dumas, B. Sterol Metabolism in the Oomycete Aphanomyces Euteiches, a Legume Root Pathogen. New Phytol. 2009, 183, 291–300. [Google Scholar] [CrossRef]
- Lerksuthirat, T.; Sangcakul, A.; Lohnoo, T.; Yingyong, W.; Rujirawat, T.; Krajaejun, T. Evolution of the Sterol Biosynthetic Pathway of Pythium insidiosum and Related Oomycetes Contributes to Antifungal Drug Resistance. Antimicrob. Agents Chemother. 2017, 61, e02352-16. [Google Scholar] [CrossRef] [Green Version]
- Lerksuthirat, T.; Lohnoo, T.; Inkomlue, R.; Rujirawat, T.; Yingyong, W.; Khositnithikul, R.; Phaonakrop, N.; Roytrakul, S.; Sullivan, T.D.; Krajaejun, T. The Elicitin-like Glycoprotein, ELI025, Is Secreted by the Pathogenic Oomycete Pythium insidiosum and Evades Host Antibody Responses. PLoS ONE 2015, 10, e0118547. [Google Scholar] [CrossRef] [Green Version]
- Dobbin, C.A.; Smith, N.C.; Johnson, A.M. Heat Shock Protein 70 Is a Potential Virulence Factor in Murine Toxoplasma Infection Via Immunomodulation of Host NF-ΚB and Nitric Oxide. J. Immunol. 2002, 169, 958–965. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, S.; Tatu, U. Heat Shock Protein 90 Localizes to the Surface and Augments Virulence Factors of Cryptococcus Neoformans. PLoS Negl. Trop. Dis. 2017, 11, e0005836. [Google Scholar] [CrossRef] [Green Version]
- Neckers, L.; Tatu, U. Molecular Chaperones in Pathogen Virulence: Emerging New Targets for Therapy. Cell Host Microbe 2008, 4, 519–527. [Google Scholar] [CrossRef] [Green Version]
- Bajaj, D.; Batra, J.K. Heat Shock Proteins in Mycobacterium Tuberculosis: Involvement in Survival and Virulence of the Pathogen. In Understanding Tuberculosis—Deciphering the Secret Life of the Bacilli; Cardona, P.-J., Ed.; InTech: London, UK, 2012; ISBN 978-953-307-946-2. [Google Scholar]
- Alam, A.; Bröms, J.E.; Kumar, R.; Sjöstedt, A. The Role of ClpB in Bacterial Stress Responses and Virulence. Front. Mol. Biosci. 2021, 8, 668910. [Google Scholar] [CrossRef]
- Kupper, M.; Gupta, S.K.; Feldhaar, H.; Gross, R. Versatile Roles of the Chaperonin GroEL in Microorganism-Insect Interactions. FEMS Microbiol. Lett. 2014, 353, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Cox, G.M.; Mukherjee, J.; Cole, G.T.; Casadevall, A.; Perfect, J.R. Urease as a Virulence Factor in Experimental Cryptococcosis. Infect. Immun. 2000, 68, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Rutherford, J.C. The Emerging Role of Urease as a General Microbial Virulence Factor. PLoS Pathog. 2014, 10, e1004062. [Google Scholar] [CrossRef] [Green Version]
- Mora, D.; Arioli, S. Microbial Urease in Health and Disease. PLoS Pathog. 2014, 10, e1004472. [Google Scholar] [CrossRef] [Green Version]
- Kusters, J.G.; van Vliet, A.H.M.; Kuipers, E.J. Pathogenesis of Helicobacter Pylori Infection. Clin. Microbiol. Rev. 2006, 19, 449–490. [Google Scholar] [CrossRef] [Green Version]
- Scott, D.R.; Marcus, E.A.; Weeks, D.L.; Sachs, G. Mechanisms of Acid Resistance Due to the Urease System of Helicobacter Pylori. Gastroenterology 2002, 123, 187–195. [Google Scholar] [CrossRef]





| VFID | MvirDB-Defined Virulence Factors | Organisms | Virulence Types | P. insidiosum Protein ID | Accession Number | Identity (%) | E-Value |
|---|---|---|---|---|---|---|---|
| 26381 | Heat shock protein 70 | Cryptosporidium parvum | Virulence protein | PINS00530003B | GAX96752.1 | 71.24 | 0 |
| 26468 | Organellar heat shock protein | Eimeria tenella | Virulence protein | PINS01150027A | GAX99732.1 | 65.0 | 0 |
| 12025 | Urease | Oryza sativa | Virulence protein | PINS00920015C a | GAX98700.1 | 64.5 | 0 |
| 15128 | Chaperone protein ClpB | Francisella tularensis | Virulence protein | PINS01620006A | GAY01393.1 | 55.1 | 0 |
| 26434 | Carbamoyl phosphate synthetase II | Toxoplasma gondii | Virulence protein | PINS04750002A | GAY05754.1 | 53.3 | 0 |
| 11169 | Tripeptidyl-peptidase 2 | Mus musculus | Protein toxin | PINS01420022C | GAY00792.1 | 34.8 | 0 |
| 26447 | Heat shock protein 70 | Toxoplasma gondii | Virulence protein | PINS00020096A | GAX92393.1 | 57.6 | 0 |
| 26455 | Eukaryotic translation initiation factor 4A | Toxoplasma gondii | Virulence protein | PINS00130063A | GAX93656.1 | 72.9 | 1 × 10−173 |
| 8350 | Chaperonin GroEL | Legionella pneumophila | Virulence protein | PINS00020065A | GAX92364.1 | 57.0 | 3 × 10−159 |
| 8728 | Phosphoglucomutase | Brucella melitensis | Virulence protein | PINS00550030A | GAX96838.1 | 50.4 | 7 × 10−158 |
| 12413 | Transcription factor site-1 protease | Homo sapiens | Protein toxin | PINS02120014C | GAY02722.1 | 40.2 | 5 × 10−142 |
| 7573 | Nonribosomal peptide synthetase Dhbf | Bacillus anthracis | Virulence protein | PINS01020025B | GAX99146.1 | 29.0 | 1 × 10−113 |
| 20164 | Asparaginyl-tRNA synthetase | Salmonella enterica | Pathogenicity island | PINS00170005A | GAX93976.1 | 46.7 | 6 × 10−113 |
| 13372 | Pyruvate kinase | Salmonella typhimurium | Virulence protein | PINS00750004A | GAX97930.1 | 45.9 | 3 × 10−107 |
| 26400 | cGMP dependent protein kinase | Toxoplasma gondii | Virulence protein | PINS00410016C | GAX95955.1 | 33.0 | 6 × 10−105 |
| 26980 | AP65-1 adhesin | Trichomonas vaginalis | Virulence protein | PINS01980002A | GAY02392.1 | 37.9 | 5 × 10−101 |
| 26382 | Heat shock protein 90 | Cryptosporidium parvum | Virulence protein | PINS00460042A | GAX96321.1 | 42.1 | 3 × 10−100 |
| 26459 | Peroxisomal catalase | Toxoplasma gondii | Virulence protein | PINS00770005C | GAX98049.1 | 40.6 | 2 × 10−92 |
| 20579 | Recombination factor protein RarA | Salmonella enterica | Pathogenicity island | PINS00070081C | GAX93094.1 | 44.8 | 5 × 10−91 |
| 14026 | Lysil-tRNA synthetase LysU | Escherichia coli | Virulence protein | PINS00380044C | GAX95759.1 | 38.5 | 5 × 10−91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krajaejun, T.; Rujirawat, T.; Lohnoo, T.; Yingyong, W.; Sae-Chew, P.; Reamtong, O.; Kittichotirat, W.; Patumcharoenpol, P. Secretome Profiling by Proteogenomic Analysis Shows Species-Specific, Temperature-Dependent, and Putative Virulence Proteins of Pythium insidiosum. J. Fungi 2022, 8, 527. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050527
Krajaejun T, Rujirawat T, Lohnoo T, Yingyong W, Sae-Chew P, Reamtong O, Kittichotirat W, Patumcharoenpol P. Secretome Profiling by Proteogenomic Analysis Shows Species-Specific, Temperature-Dependent, and Putative Virulence Proteins of Pythium insidiosum. Journal of Fungi. 2022; 8(5):527. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050527
Chicago/Turabian StyleKrajaejun, Theerapong, Thidarat Rujirawat, Tassanee Lohnoo, Wanta Yingyong, Pattarana Sae-Chew, Onrapak Reamtong, Weerayuth Kittichotirat, and Preecha Patumcharoenpol. 2022. "Secretome Profiling by Proteogenomic Analysis Shows Species-Specific, Temperature-Dependent, and Putative Virulence Proteins of Pythium insidiosum" Journal of Fungi 8, no. 5: 527. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050527