
Morphological and Phylogenetic Characterization Reveals Five New Species of Samsoniella (Cordycipitaceae, Hypocreales)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation
2.2. Morphological Observations
2.3. DNA Extraction, PCR and Sequencing for Nuclear Genes
2.4. Phylogenetic Analyses
3. Results
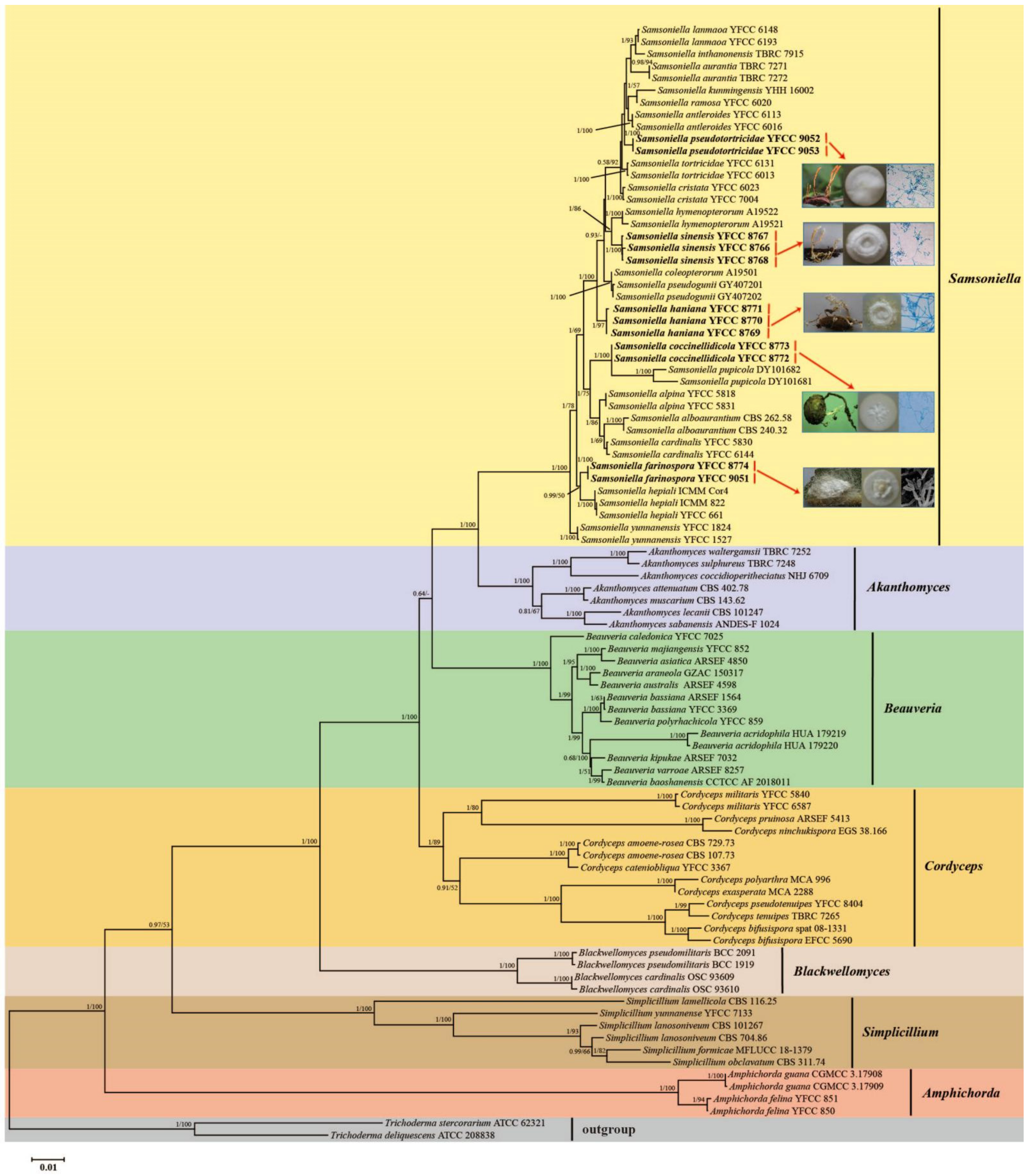
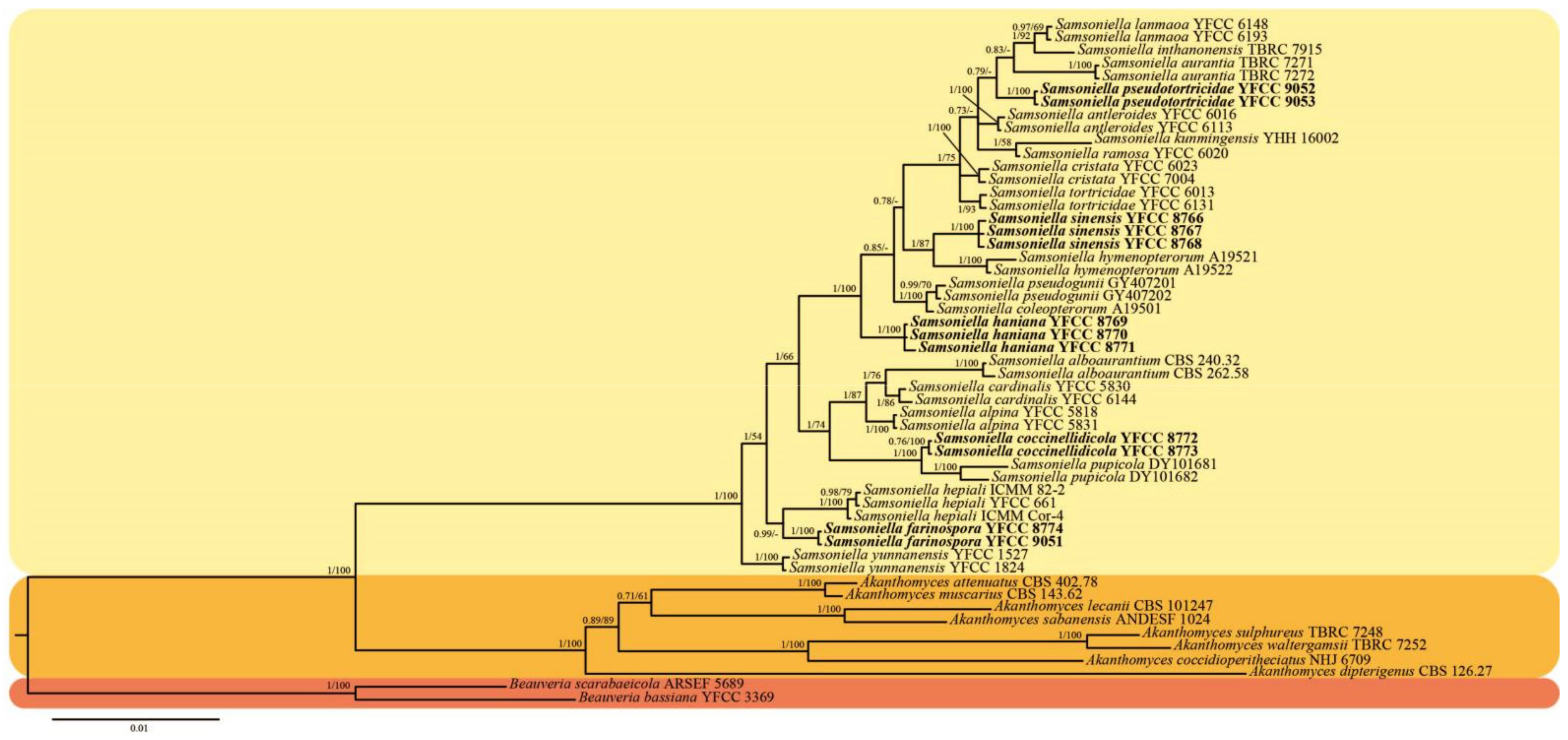
3.1. Sequencing and Phylogenetic Analyses
- Samsoniella alboaurantium
- Samsoniella alpina
- Samsoniella antleroides
- Samsoniella aurantia
- Samsoniella cardinalis
- Samsoniella coccinellidicola
- Samsoniella coleopterorum
- Samsoniella cristata
- Samsoniella farinospora
- Samsoniella haniana
- Samsoniella hepiali
- Samsoniella hymenopterorum
- Samsoniella inthanonensis
- Samsoniella kunmingensis
- Samsoniella lanmaoa
- Samsoniella lepidopterorum
- Samsoniella pseudogunii
- Samsoniella pseudotortricidae
- Samsoniella pupicola
- Samsoniella ramosa
- Samsoniella sinensis
- Samsoniella tortricidae
- Samsoniella yunnanensis
| Teleomorph characters |
| Insect host |
| 1. Lepidoptera (pupa,larva) .................................................................................................3, 5, 8, 13, 14, 15, 18, 20, 22 |
| Stromata |
| 1. Number |
| a. Fasciculate ................................................................................................................................................3, 13, 20, 22 b. Several ...................................................................................................................................................................5 c. Solitary or two ........................................................................................................................................................8 d. Solitary or Several ...................................................................................................................................................18 e. Solitary ....................................................................................................................................................................14 f. Two to five ................................................................................................................................................................15 |
| 2. Size (long) |
| a. 10~20 mm ..................................................................................................................................................................... 5 b. 15~40 mm .......................................................................................................................................................8, 14, 20 c. 20~70 mm ..............................................................................................................................................3, 13, 15, 18, 22 |
| 3. Shape |
| a. Cylindrical to clavate, branches .........................................................................................................................3, 13 b. Cylindrical, unbranches .............................................................................................................................................5 c. Cylindrical, unbranches or dichotomous ..............................................................................................................18 d. Crista-like, much branched .......................................................................................................................................8 e. Cylindrical to clavate, bifurcated ...........................................................................................................................14 f. Palmately branched ..................................................................................................................................................15 g. Fascicular, multi-branched, often confluent at the base .....................................................................................20 h. Unbranched or dichotomous ...................................................................................................................................22 |
| Fertile parts |
| 1. Shape |
| a. Clavate to fake-like ....................................................................................................................................................3 b. Clavate ...............................................................................................................................................................5, 14, 15 c. Clavate to subulate ....................................................................................................................................................18 d. Crista-like or subulate .........................................................................................................................................8, 22 e. Having no obvious boundary with stipes, with a tapering sterile part ...........................................................20 |
| 2. Colour |
| a. Orange red ................................................................................................................................................................13 b. Orange to orange red ................................................................................................................................................3 c. Scarlet ..........................................................................................................................................................................5 d. Reddish orange ........................................................................................................................................8, 14, 15, 18 e. White to pale brown ..........................................................................................................................................20, 22 |
| Perithecia |
| 1. shape |
| a. Superficial, ovoid ........................................................................................................................................................13 b. Superficial, fusiform ................................................................................................................................................3 c. Superficial, oblong-ovate to fusiform ...................................................................................................................5 d. Superficial, narrowly ovoid ..................................................................................................................................8 e. Superficial, narrowly ovoid to fusiform ....................................................................................14, 15, 18, 20, 22 |
| 2. Length |
| a. 280~450 µm ................................................................................................................................................3, 14, 18, 20 b. 330~470 µm ..........................................................................................................................................................15, 22 c. 370~490 µm ........................................................................................................................................................5, 8, 13 |
| 3. Width |
| a. 110~210 µm ......................................................................................................................................... 14, 15, 18, 20 b. 130~250 µm ................................................................................................................................................3, 5, 8, 22 c. 200~260 µm ..............................................................................................................................................................13 |
| Asci |
| 1. Shape |
| a. Cylindrical, eight-spored, hyaline ...........................................................................................3, 5, 8, 13, 14, 15, 22 |
| 2. Length |
| a. 150~300 µm ............................................................................................................................................3, 13, 14, 22 b. 160~360 µm .....................................................................................................................................................5, 8, 15 |
| 3. Width |
| a. 2~3 µm ................................................................................................................................................................3, 13 b. 2.5~4 µm .................................................................................................................................................................22 c. 3~5 µm .......................................................................................................................................................5, 8, 14, 15 |
| Ascospores |
| 1. Shape |
| a. Bola-shaped, septate, central part fliform, terminal part narrowly fusiform .....................3, 5, 8, 13, 14, 15, 22 |
| 2. Length |
| a. 110~185 µm ................................................................................................................................................................3 b. 120~300 µm .....................................................................................................................................5, 8, 13, 14, 15, 22 |
| 3. Width |
| a. 0.5~1.0 µm ......................................................................................................................................................... 5, 13 b. 0.8~1.5 µm ............................................................................................................................................3, 8, 14, 15, 22 |
| Anamorph characters |
| Insect host |
| 1. Lepidoptera (pupa, larva) .................................................................................1, 2, 4, 9, 10, 11, 16, 17, 19, 21, 23 2. Dermaptera ................................................................................................................................................................21 3. Coleoptera (Snout beetle) .....................................................................................................................................6, 7 4. Hymenoptera (bee) ....................................................................................................................................................12 5. Araneae .......................................................................................................................................................................9 |
| Synnemata |
| 1. Present ......................................................................................................................................2, 4, 6, 10, 11, 21, 23 |
| Irregularly branched ................................................................................................................................2, 4, 6, 10, 21 b. Branched or unbranched ........................................................................................................................................11 c. Gregarious ................................................................................................................................................................23 |
| 2. Not observed ..............................................................................................................................1, 7, 9, 12, 16, 17, 19 |
| Cultural characteristics on PDA |
| 1. Growth rate on PDA at 25 °C at 2 wk |
| a. Relatively rapid (>60 mm diam) ..........................................................................................................................12 b. Moderate (30–60 mm diam) ...........................................................................................2, 6, 7, 9, 11, 16, 17, 21, 23 c. Slow (<30 mm diam) ......................................................................................................................................4, 10, 19 |
| 2. Conidiophores |
| a. Biverticillate ............................................................................................................................................................2 b. Solitary .............................................................................................................................................................11, 19 c. Solitary or verticillate .........................................................................................................6, 7, 9, 10, 12, 16, 21, 23 d. Verticillate .......................................................................................................................................................1, 4, 17 |
| 3. Phialides number in a whorls |
| a. 2–4 ..............................................................................................................................................................1, 4, 7, 9, 16 b. 2–5 ...........................................................................................................................................................6, 10, 11, 21 c. 2–7 .......................................................................................................................................................................2, 23 b. 2–9 .....................................................................................................................................................................17, 19 e. 3–4 .............................................................................................................................................................................12 |
| 4. Shape of conidia |
| a. fusiform or oval ............................................................................................................................2, 4, 6, 10, 11, 12, 23 b. fusiform, ellipsoidal or subglobose ..........................................................................................................................7 c. fusiform to subglobose ............................................................................................................................................16 d. fusiform ..............................................................................................................................................................17, 19 e. ellipsoidal to fusiform, sometimes lemon-shaped ...................................................................................................1 f. oblong to cylindrical ..................................................................................................................................................9 g. spherical, elliptical or fusiform .................................................................................................................................21 |
| Key toSamsoniellaspecies |
| Sexual state present .........................................................................................................................................................1 |
| Sexual state not observed ................................................................................................................................................5 |
| 1a. Stromata fasciculate ...................................................................................................................................................2 1b. Stromata not fasciculate ...........................................................................................................................................4 2a. Stromata cylindrical to clavate, branches ..........................................................................................................................3 2b. Stromata fascicular, multi-branched, often confluent at the base ..................................................................S. ramosa 2c. Stromata unbranched or dichotomous .........................................................................................................S. tortricidae 3a. Fertile parts clavate to fake-like, orange to orange red; Perithecia superficial, fusiform .......................S. antleroides 3b. Fertile parts clavate, orange red; Perithecia superficial, ovoid ..............................................................S. inthanonensis 4a. Stromata several, cylindrical, unbranches; Fertile parts clavate, scarlet .....................................................S. cardinalis 4b. Stromata solitary or two, crista-like, much branched; Fertile parts crista-like or subulate, reddish orange; Ascospores 155–290 × 1.0–1.3 μm ...................................................................................................................................S. cristata 4c. Stromata solitary or several, cylindrical, unbranches or dichotomous; Fertile parts clavate to subulate, reddish orange; No mature ascospores were observed ........................................................................................S. pseudotortricidae 4d. Stromata solitary, cylindrical to clavate, bifurcated; Fertile parts clavate, reddish orange; Ascospores 127–190 × 0.5–1.5 μm ..........................................................................................................................................................S. kunmingensis 4e. Stromata two to five, palmately, branched; Fertile parts clavate, reddish orange; Ascospores 135–260 × 0.9–1.4 μm .................................................................................................................................................................................S. lanmaoa 5a. Synnemata Present .................................................................................................................................................................6 5b. Synnemata not observed ......................................................................................................................................................11 6a. Synnemata irregularly branched ..........................................................................................................................................7 6b. Synnemata branched or unbranched; Conidiophores solitary, with phialides in whorls of two to five ...................................................................................................................................................................................S. hepiali 6c. Synnemata gregarious; Conidiophores solitary or verticillate, with phialides in whorls of two to seven .....................................................................................................................................................................S. yunnanensis 7a. Colonies on PDA at 25 °C at 2 wk growing moderate (30–60 mm diam) ......................................................................8 7b. Colonies on PDA at 25 °C at 2 wk growing slow (<30 mm diam) ................................................................................10 8a. Conidiophores biverticillate....................................................................................................................................S. alpina 8b. Conidiophores solitary or verticillate ................................................................................................................................9 9a. Conidia fusiform or oval, 1.8–3.0 × 1.3–2.0 μm .....................................................................................S. coccinellidicola 9b. Conidia spherical, elliptical or fusiform, 2.0–3.1 × 1.3–1.9 μm .........................................................................S. sinensis 10a. Conidiophores solitary or verticillate, with phialides in whorls of two to five; Conidia fusiform or oval, 2.3–3.7 × 1.2–2.8 μm ...................................................................................................................................................................S. haniana 10b. Conidiophores verticillate, with phialides in whorls of two to four; Conidia fusiform or oval, 2.5–3.5 × 1.5 μm .................................................................................................................................................................................S. aurantia 11a. Colonies on PDA at 25 °C at 2 wk growing relatively rapid (>60 mm diam) ..............................S. hymenopterorum 11b. Colonies on PDA at 25 °C at 2 wk growing moderate (30–60 mm diam) ..................................................................12 11c. Colonies on PDA at 25 °C at 2 wk growing slow (<30 mm diam) ...............................................................S. pupicola 12a. Conidiophores verticillate ...............................................................................................................................................13 12b. Conidiophores solitary or verticillate ............................................................................................................................14 13a. Have phialides in whorls of two to four; Conidia ellipsoidal to fusiform, sometimes lemon-shaped, 2.0–3.0 × 1.0–1.8 μm ..........................................................................................................................................................S. alboaurantium 13b. Have phialides in whorls of two to nine; Conidia fusiform, 2.8–3.2 × 1.7–2.1 μm .............................S. pseudogunii 14a. Conidia fusiform, ellipsoidal or subglobose, 1.7–2.5 × 1.2–1.8 μm ...................................................S. coleopterorum 14b. Conidia oblong to cylindrical, 1.6–2.8 × 0.6–1.2 μm ..................................................................................S. farinospora 14c. Conidia fusiform to subglobose, 2.0–2.5 × 1.2–2.0 μm .......................................................................S. lepidopterorum |
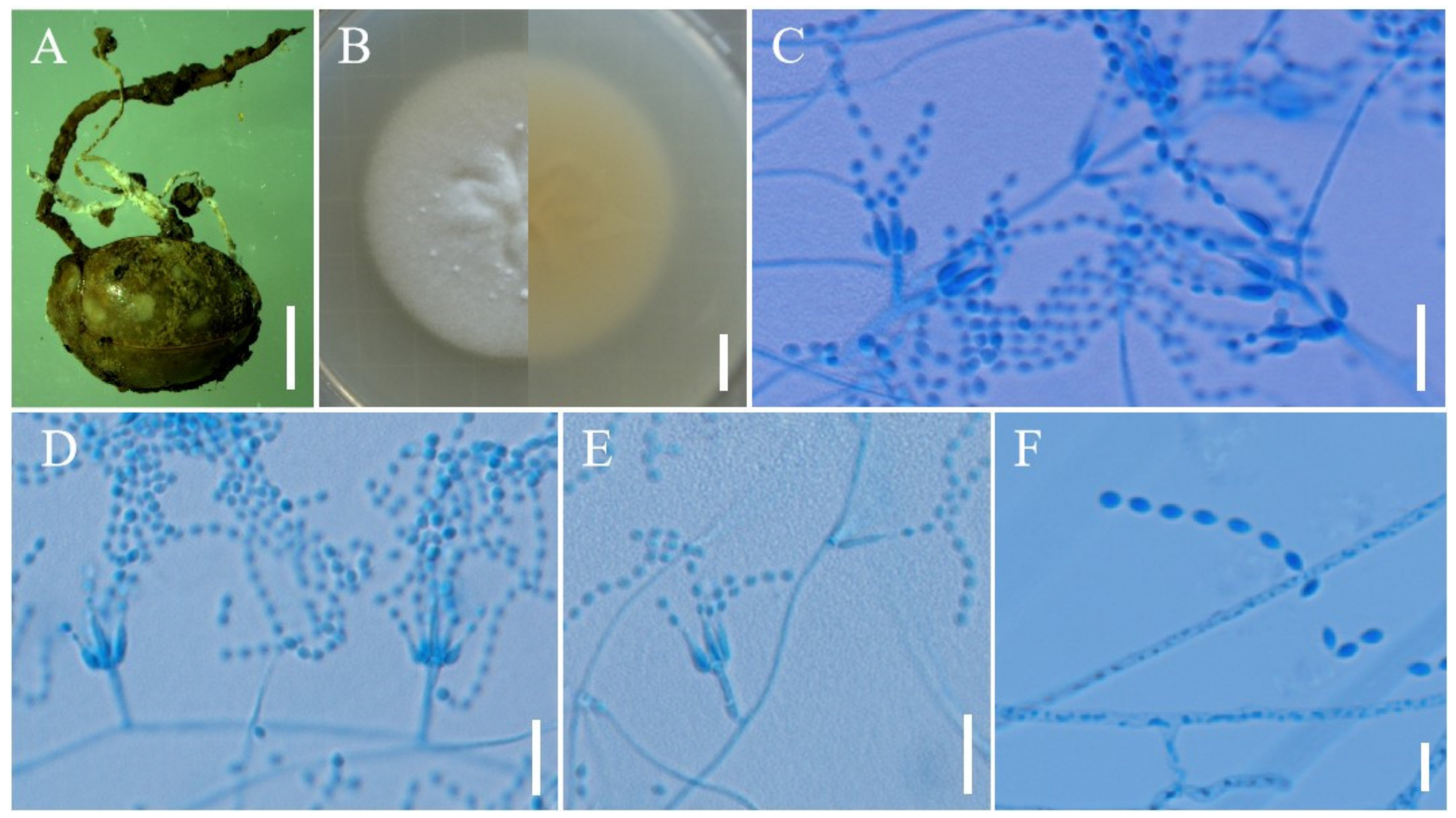
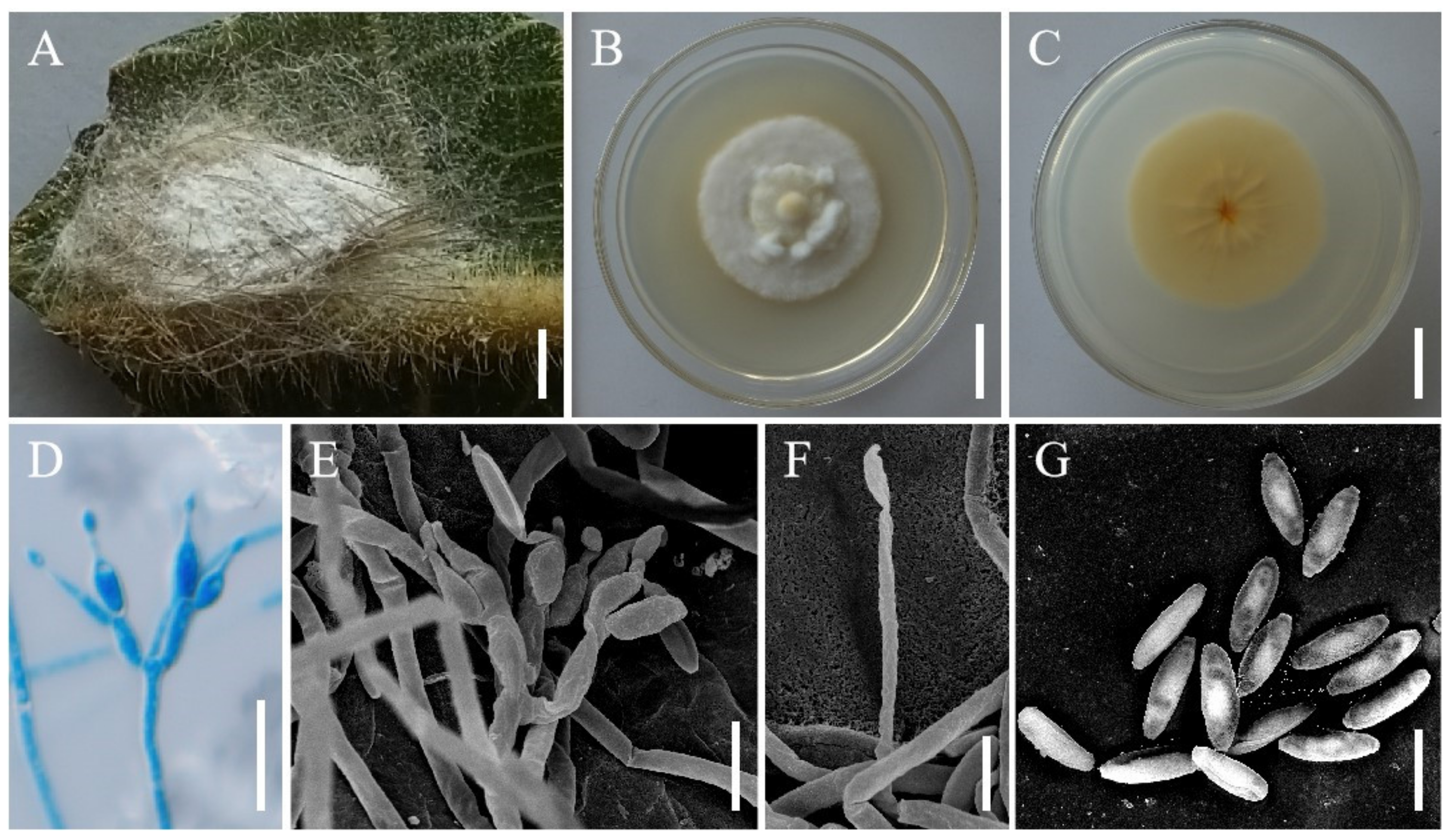
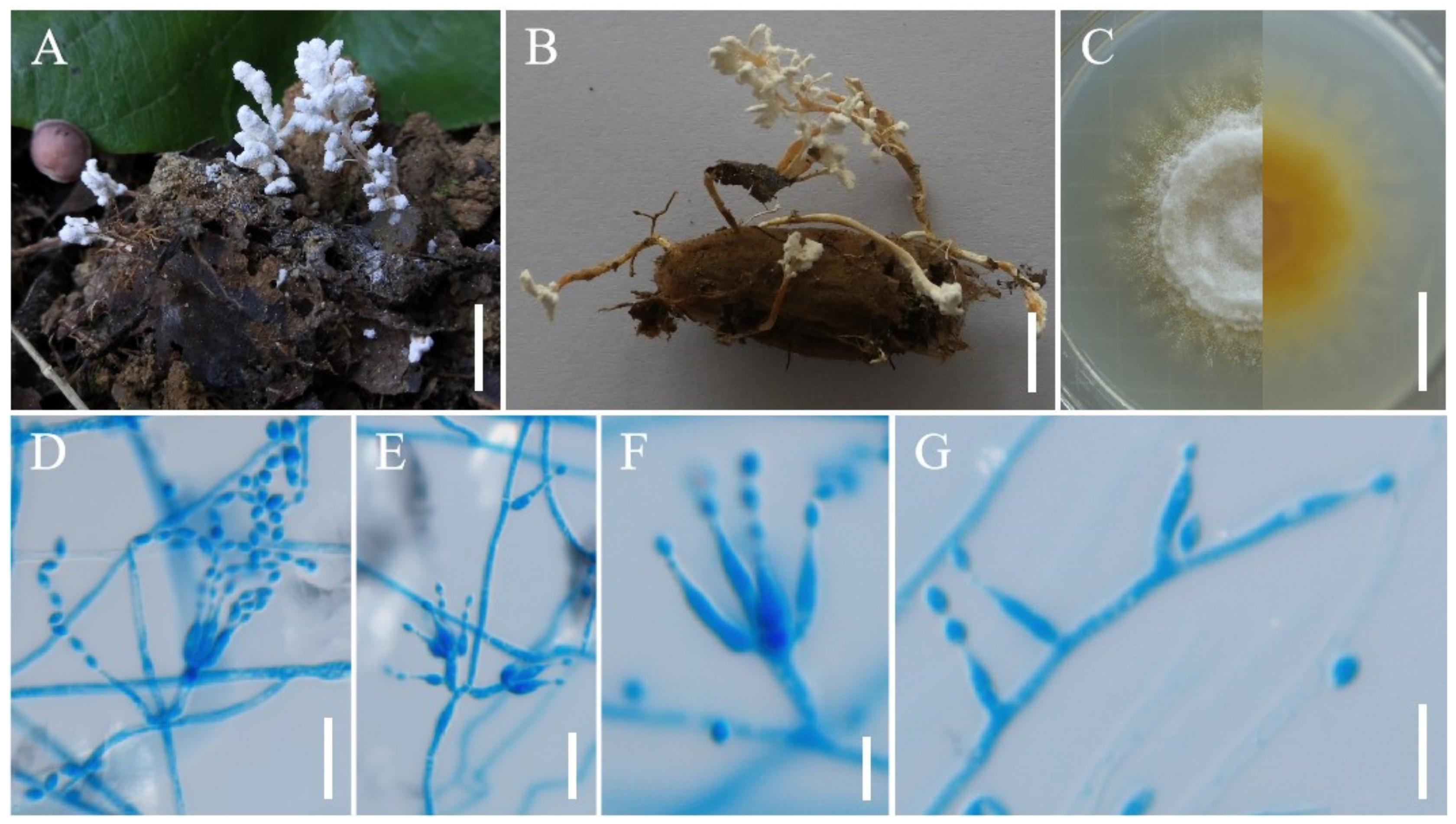
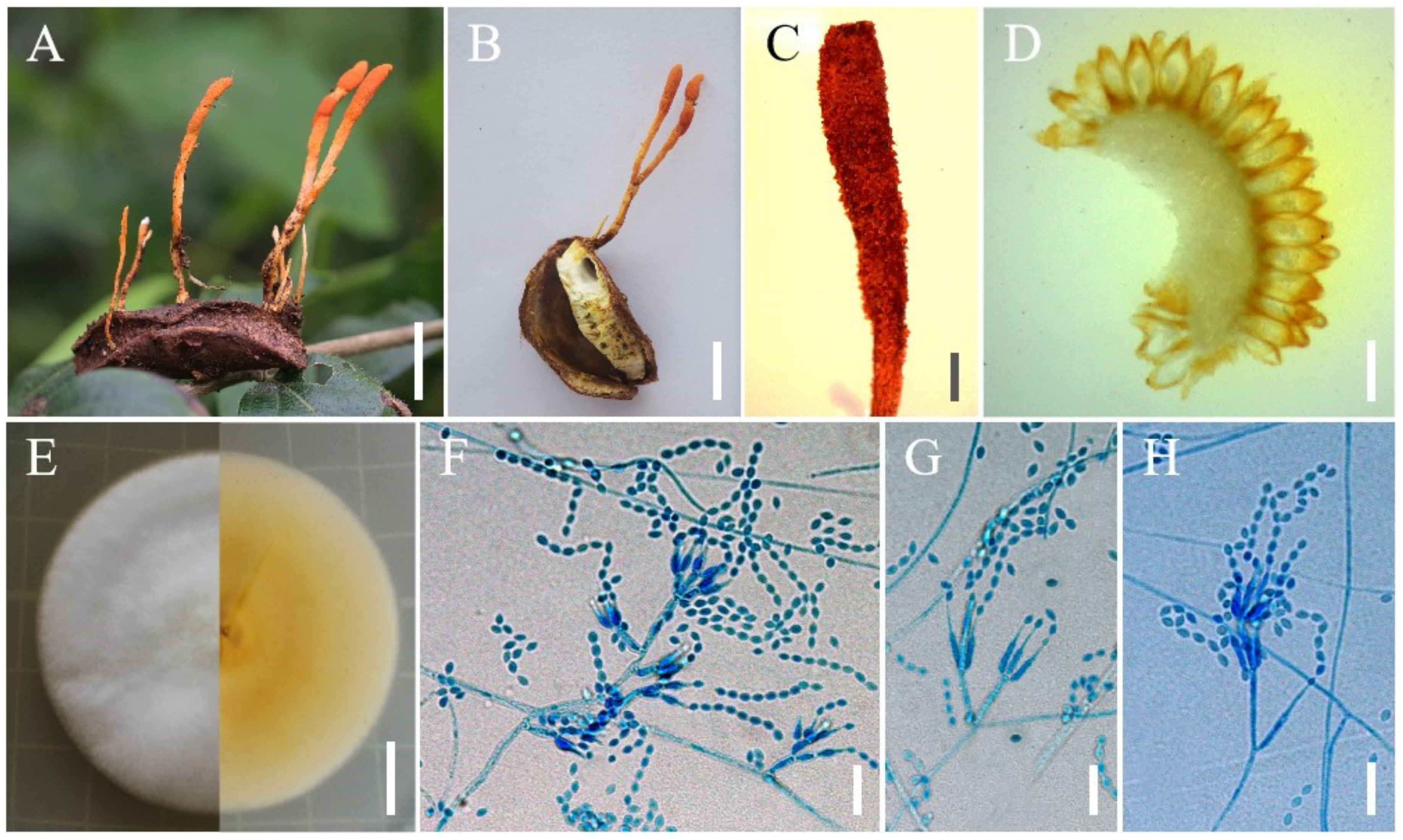
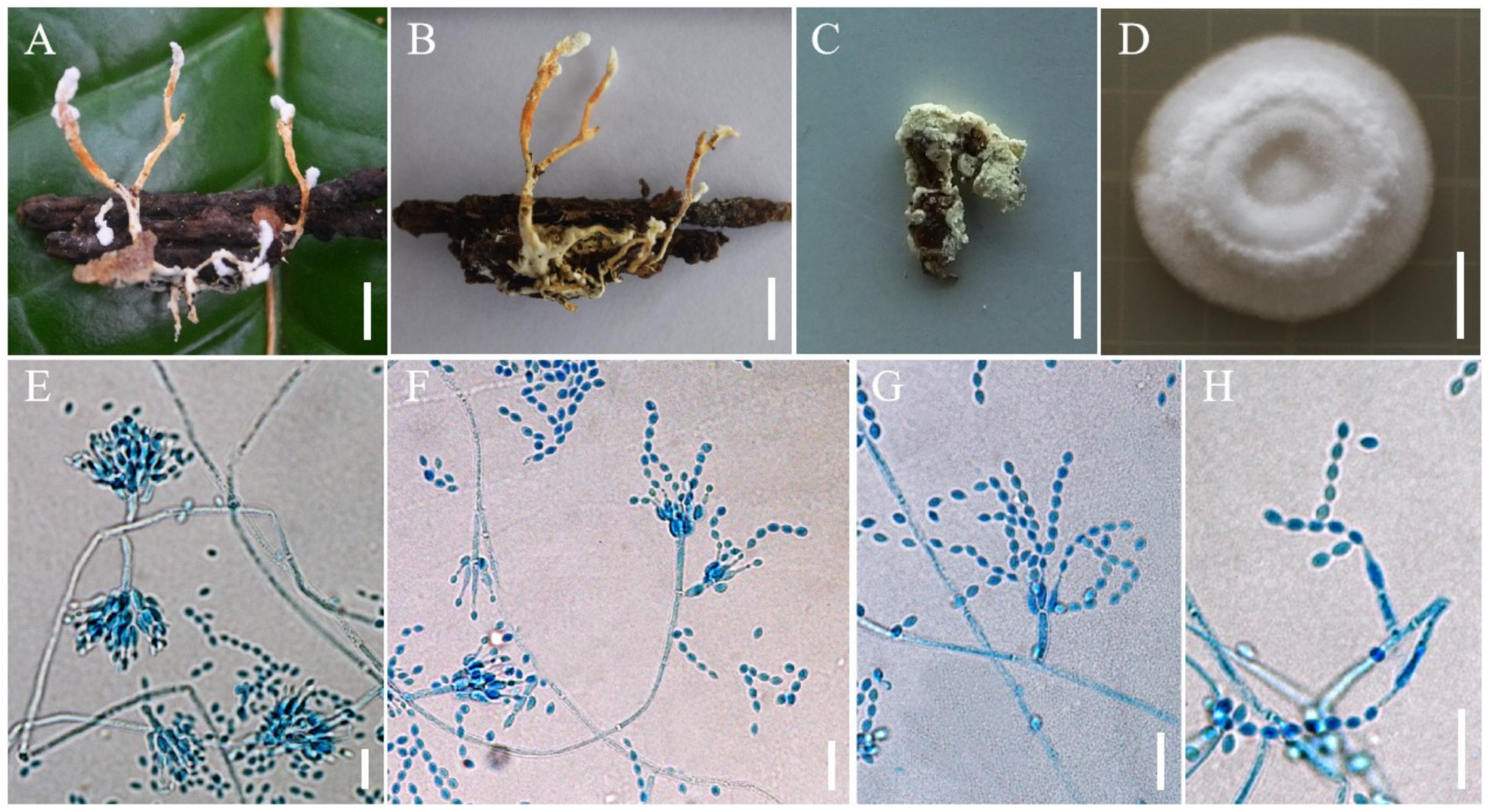
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mongkolsamrit, S.; Noisripoom, W.; Thanakitpipattana, D.; Wutikhun, T.; Spatafora, J.W.; Luangsa-ard, J. Disentangling cryptic species with Isaria-like morphs in Cordycipitaceae. Mycologia 2018, 110, 230–257. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.B.; Wang, Y.; Fan, Q.; Duan, D.E.; Zhang, G.D.; Dai, R.Q.; Dai, Y.D.; Zeng, W.B.; Chen, Z.H.; Li, D.D.; et al. Multigene phylogeny of the family Cordycipitaceae (Hypocreales): New taxa and the new systematic position of the Chinese cordycipitoid fungus Paecilomyces hepiali. Fungal Divers. 2020, 103, 1–46. [Google Scholar] [CrossRef]
- Luangsa-ard, J.J.; Hywel-Jones, N.L.; Samson, R.A. The order level polyphyletic nature of Paecilomyces sensu lato as revealed through 18S-generated rRNA phylogeny. Mycologia 2004, 96, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Kepler, R.M.; Luangsa-ard, J.J.; Hywel-Jones, N.L.; Quandt, C.A.; Sung, G.H.; Rehner, S.A.; Aime, M.C.; Henkel, T.W.; Sanjuan, T.; Zare, R.; et al. A phylogenetically-based nomenclature for Cordycipitaceae (Hypocreales). IMA Fungus 2017, 8, 335–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.P.; Xu, J.Y.; Li, H.C.; Song, L.P.; Feng, C.Q. The complete mitochondrial genome of Paecilomyces hepiali (Ascomycota, Eurotiomycetes). Mitochondrial DNA J. Dna Mapp. Seq. Anal. 2014, 27, 916–917. [Google Scholar] [CrossRef]
- Wang, Y.B.; Yu, H.; Dai, Y.D.; Wu, C.K.; Zeng, W.B.; Yuan, F.; Liang, Z.Q. Polycephalomyces agaricus, a new hyperparasite of Ophiocordyceps sp. infecting melolonthid larvae in southwestern China. Mycol. Prog. 2015, 14, 70. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identifcation and mapping of enzymatically amplifed ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Bischof, J.F.; Rehner, S.A.; Humber, R.A. Metarhizium frigidum sp. nov.: A cryptic species of M. anisopliae and a member of the M. favoviride Complex. Mycologia 2006, 98, 737–745. [Google Scholar] [CrossRef]
- Sung, G.H.; Hywel-Jones, N.L.; Sung, J.M.; Luangsa-Ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classifcation of Cordyceps and the clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.B.; Yu, H.; Dai, Y.D.; Chen, Z.H.; Zeng, W.B.; Yuan, F.; Liang, Z.Q. Polycephalomyces yunnanensis (Hypocreales), a new species of Polycephalomyces parasitizing Ophiocordyceps nutans and stink bugs (Hemipteran adults). Phytotaxa 2015, 208, 34–44. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML Web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: Moremodels, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.; Sung, G.H.; Hywel-Jones, N.L.; Luangsa-Ard, J.J.; Bischoff, J.F.; Kepler, R.M.; Spatafora, J.W. Systematics and evolution of the genus Torrubiella (Hypocreales, Ascomycota). Mycol. Res. 2009, 113, 279–289. [Google Scholar] [CrossRef]
- Spatafora, J.W.; Sung, G.H.; Hywel-Jones, N.L.; White, J.F. Phylogenetic evidence for an animal pathogen origin of ergot and the grass endophytes. Mol. Ecol. 2007, 16, 1701–1711. [Google Scholar] [CrossRef]
- Chiriví-Salomón, J.S.; Danies, G.; Restrepo, S.; Sanjuan, T. Lecanicillium sabanense sp. nov. (Cordycipitaceae) a new fungal entomopathogen of coccids. Phytotaxa 2015, 234, 63–74. [Google Scholar] [CrossRef]
- Wang, Y.; Fan, Q.; Wang, D.; Zou, W.Q.; Tang, D.X.; Hongthong, P.; Yu, H. Species Diversity and Virulence Potential of the Beauveria bassiana Complex and Beauveria scarabaeidicola Complex. Front. Microbiol. 2022, 13, 841604. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.Z.; Liu, S.J.; Cai, L. Culturable mycobiota from Karst caves in China, with descriptions of 20 new species. Pers. Mol. Phylogeny Evol. Fungi 2017, 39, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Sanjuan, T.; Tabima, J.; Restrepo, S.; Læssøe, T.; Spatafora, J.W.; FrancoMolano, A.E. Entomopathogens of Amazonian stick insects and locusts are members of the Beauveria species complex (Cordyceps sensu stricto). Mycologia 2014, 106, 260–275. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.H.; Han, Y.F.; Liang, Z.Q.; Jin, D.C. A new araneogenous fungus in the genus Beauveria from Guizhou, China. Phytotaxa 2017, 302, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S.A.; Minnis, A.M.; Sung, G.H.; Luangsa-ard, J.J.; Devotto, L.; Humber, R.A. Phylogeny and systematics of the anamorphic, entomopathogenic genus Beauveria. Mycologia 2011, 103, 1055–1073. [Google Scholar] [CrossRef]
- Chen, Z.H.; Chen, K.; Dai, Y.D.; Zheng, Y.; Xu, L. Beauveria species diversity in the Gaoligong Mountains of China. Mycol. Prog. 2019, 18, 933–943. [Google Scholar] [CrossRef]
- Luangsa-ard, J.J.; Hywel-Jones, N.L.; Manoch, L.; Samson, R.A. On the relationships of Paecilomyces sect. Isarioidea species. Mycol. Res. 2005, 109, 581–589. [Google Scholar] [CrossRef] [Green Version]
- Dong, Q.Y.; Wang, Y.; Wang, Z.Q.; Tang, D.X.; Zhao, Z.Y.; Wu, H.J.; Yu, H. Morphology and phylogeny reveal five novel species in the genus Cordyceps (Cordycipitaceae, Hypocreales) from Yunnan, China. Front. Microbiol. 2022, 13, 846909. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.H.; Han, Y.F.; Liang, J.D.; Tian, W.Y.; Liang, Z.Q. Morphological and phylogenetic characterisations reveal three new species of Samsoniella (Cordycipitaceae, Hypocreales) from Guizhou, China. MycoKeys 2020, 74, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.H.; Liang, J.D.; Ren, X.X.; Zhao, J.H.; Han, Y.F.; Liang, Z.Q. Cryptic diversity of isaria-like species in Guizhou, China. Life 2021, 11, 1093. [Google Scholar] [CrossRef]
- Wei, D.P.; Wanasinghe, D.N.; Hyde, K.D.; Mortimer, P.E.; To-Anun, C. The genus Simplicillium. MycoKeys 2019, 60, 69–92. [Google Scholar] [CrossRef]
- Currie, C.R.; Wong, B.; Stuart, A.E.; Schultz, T.R.; Rehner, S.A.; Mueller, U.G.; Sung, G.H.; Spatafora, J.W.; Straus, N.A. Ancient Tripartite Coevolution in the Attine Ant-Microbe Symbiosis. Science 2003, 299, 386–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, G. Some new and interesting species of micro-fungi. Trans. Br. Mycol. Soc. 1957, 40, 481–488. [Google Scholar] [CrossRef]
- Lou, Y.Q.; Liao, X.M.; Lu, Y.C. Cardiovascular pharmacological studies of ethanol extracts of Cordyceps mycelia and Cordyceps fermentation solution. Chin. Tradit. Herb. Drugs 1986, 17, 17–21. [Google Scholar]
- Huang, M.M.; Zhang, J.F.; Pang, L.; Jiang, Z.; Wang, D.W. Studies on immunopharmacology of Cordyceps (Fr.) Link IV. Observations on the immunosuppressive activity of artifcial substance of Paecilomyces hapiali Chen. Acta Univ. Med. Tongji 1988, 5, 329–331. [Google Scholar]
- Wang, D.W.; Huang, M.M. Studies on immunopharmacology of Cordyceps (Fr.) Link V. Infuence of artifcial fermentative substance of Paecilomyces hapiali Chen on the function of T cell and its subgroup in mice. Acta Univ. Med. Tongji 1988, 5, 332–334. [Google Scholar]
- Dai, R.Q.; Lan, J.L.; Chen, W.H.; Li, X.M.; Chen, Q.T.; Shen, C.Y. Research on Paecilomyces hepiali Chen et Dai, sp. nov. Acta Agric. Univ. Pekin 1989, 15, 221–224. [Google Scholar]
- Zou, W.P.; Huang, M.M. Primary studies on the mechanism of Paecilomyces hepiali Chen against the rejection reaction. Acta Univ. Med. Tongji 1993, 22, 282–284. [Google Scholar]
- Xiang, M.; Tang, J.; Chu, T.; Zhang, C.L.; Zou, X.L. Hypoglycemic effect and mechanism study on streptozocin induced diabetes in mice by Paecilomyces hepiali Chen mycelium. Chin. J. Hosp. Pharm. 2006, 26, 556–559. [Google Scholar]
- Jiang, L.; Bao, H.Y.; Yang, M. Antitumor activity of a petroleum ether extract from Paecilomyces hepiali mycelium. Acta Edulis Fungi 2010, 17, 58–60. [Google Scholar]
- Dai, R.Q.; Shen, C.Y.; Li, X.M.; Lan, J.L.; Lin, S.F.; Shao, A.J. Response to neotypification of Paecilomyces hepiali (Hypocreales) (Wang & al., 2015). Taxon 2018, 67, 784–786. [Google Scholar] [CrossRef] [Green Version]
- Samson, R.A. Paecilomyces and some allied hyphomycetes. Stud. Mycol. 1974, 6, 1–119. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | 5′-Sequence-3′ | References |
|---|---|---|---|
| nrSSU | nrSSU-CoF | TCTCAAAGATTAAGCCATGC | [6] |
| nrSSU-CoR | TCACCAACGGAGACCTTG | ||
| nrLSU | LR5 | ATCCTGAGGGAAACTTC | [7,8] |
| LR0R | GTACCCGCTGAACTTAAGC | ||
| tef-1α | EF1α-EF | GCTCCYGGHCAYCGTGAYTTYAT | [9,10] |
| EF1α-ER | ATGACACCRACRGCRACRGTYTG | ||
| rpb1 | RPB1-5′F | CAYCCWGGYTTYATCAAGAA | [9,10] |
| RPB1-5′R | CCNGCDATNTCRTTRTCCATRTA | ||
| rpb2 | RPB2-5′F | CCCATRGCTTGTYYRCCCAT | [9,10] |
| RPB2-5′R | GAYGAYMGWGATCAYTTYGG |
| Taxon | Voucher Information | Host | GenBank Accession Number | References | ||||
|---|---|---|---|---|---|---|---|---|
| nrSSU | nrLSU | tef-1α | rpb1 | rpb2 | ||||
| Akanthomyces attenuatus | CBS 402.78 | Leaf litter | AF339614 | AF339565 | EF468782 | EF468888 | EF468935 | [10] |
| coccidioperitheciatus | NHJ 6709 | Araneae | EU369110 | EU369042 | EU369025 | EU369067 | EU369086 | [17] |
| A. dipterigenus | CBS 126.27 | Hemiptera: Monophlebidae | AF339605 | AF339556 | KM283820 | KR064300 | KR064303 | Unpublished |
| A. lecanii | CBS 101247 | Hemiptera: Coccidae | AF339604 | AF339555 | DQ522359 | DQ522407 | DQ522466 | [18] |
| A. muscarius | CBS 143.62 | Hemiptera: Aleyrodidae | KM283774 | KM283798 | KM283821 | KM283841 | KM283863 | Unpublished |
| A. sabanensis | ANDES-F 1024 | Hemiptera: Coccidae | KC633251 | KC875225 | KC633266 | KC633249 | [19] | |
| A. sulphureus | TBRC 7248 | Araneae | MF140722 | MF140843 | MF140787 | MF140812 | [1] | |
| A. waltergamsii | TBRC 7252 | Araneae | MF140714 | MF140834 | MF140782 | MF140806 | [1] | |
| Amphichorda felina | YFCC 850 | Bird droppings | MW181774 | MW173986 | MW168227 | MW168193 | MW168210 | [20] |
| A. felina | YFCC 851 | Bird droppings | MW181775 | MW173987 | MW168228 | MW168194 | MW168211 | [20] |
| A. guana | CGMCC 3.17908 | Bat guano | KY883262 | KU746711 | KX855211 | KY883202 | KY883228 | [21] |
| A. guana | CGMCC 3.17909 | Bat guano | KY883263 | KU746712 | KX855212 | KY883203 | [21] | |
| Beauveria acridophila | HUA 179219 | Orthoptera: Acrididae | JQ895541 | JQ958613 | JX003857 | JX003841 | [22] | |
| B. acridophila | HUA 179220 | Orthoptera: Acrididae | JQ895527 | JQ895536 | JQ958614 | JX003852 | JX003842 | [22] |
| B. araneola | GZAC 150317 | Araneae | KT961699 | KT961701 | [23] | |||
| B. asiatica | ARSEF 4850 | Coleoptera: Cerambycidae | AY531937 | HQ880859 | HQ880931 | [24] | ||
| B. australis | ARSEF 4598 | Soil | HQ880995 | HQ880861 | HQ880933 | [24] | ||
| B. bassiana | YFCC 3369 | Coleoptera: Scarabaeidae | MN576768 | MN576824 | MN576994 | MN576884 | MN576938 | [2] |
| B. baoshanensis | CCTCC AF 2018011 | Coleoptera: Chrysomelidae | MG642882 | MG642840 | MG642897 | MG642854 | MG642867 | [25] |
| B. caledonica | YFCC 7025 | MN576771 | MN576827 | MN576997 | MN576887 | MN576941 | [2] | |
| B. kipukae | ARSEF 7032 | Homoptera: Delphacidae | HQ881005 | HQ880875 | HQ880947 | [24] | ||
| B. majiangensis | YFCC 852 | Hemiptera: Pentatomidae | MW181776 | MW173988 | MW168229 | MW168195 | MW168212 | [20] |
| B. polyrhachicola | YFCC 859 | Hymenoptera: Formicidae | MW181783 | MW173995 | MW168236 | MW168202 | MW168219 | [20] |
| B.scarabaeidicola | ARSEF 5689 | Coleoptera: Scarabaeidae | AF339574 | AF339524 | DQ522335 | DQ522380 | DQ522431 | [20] |
| B. varroae | ARSEF 8257 | Coleoptera: Curculionidae | HQ881002 | HQ880872 | HQ880944 | [24] | ||
| Blackwellomyces cardinalis | OSC 93609 | Lepidoptera: Tineidae | AY184973 | AY184962 | DQ522325 | DQ522370 | DQ522422 | [18] |
| B. cardinalis | OSC 93610 | Lepidoptera: Tineidae | AY184974 | AY184963 | EF469059 | EF469088 | EF469106 | [18] |
| B. pseudomilitaris | BCC 1919 | Lepidoptera (Larva) | MF416588 | MF416534 | MF416478 | MF416440 | [4] | |
| B. pseudomilitaris | BCC 2091 | Lepidoptera (Larva) | MF416589 | MF416535 | MF416479 | MF416441 | [4] | |
| Cordyceps amoene-rosea | CBS 107.73 | Coleoptera (Pupa) | AY526464 | MF416550 | MF416494 | MF416651 | MF416445 | [26] |
| C. amoene-rosea | CBS 729.73 | Coleoptera: Nitidulidae | MF416604 | MF416551 | MF416495 | MF416652 | MF416446 | [26] |
| C. bifusispora | EFCC 5690 | Lepidoptera (Pupa) | EF468952 | EF468806 | EF468746 | EF468854 | EF468909 | [10] |
| C. bifusispora | spat 08-133.1 | Lepidoptera (Pupa) | MF416577 | MF416524 | MF416469 | MF416631 | MF416434 | [4] |
| C. cateniobliqua | YFCC 3367 | Coleoptera adult | MN576765 | MN576821 | MN576991 | MN576881 | MN576935 | [2] |
| C. exasperata | MCA 2288 | Lepidoptera (Larva) | MF416592 | MF416538 | MF416482 | MF416639 | [4] | |
| C. militaris | YFCC 6587 | Lepidoptera (Pupa) | MN576762 | MN576818 | MN576988 | MN576878 | MN576932 | [2] |
| C. militaris | YFCC 5840 | Lepidoptera (Pupa) | MN576763 | MN576819 | MN576989 | MN576879 | MN576933 | [2] |
| C. ninchukispora | EGS 38.166 | Plant (Beilschmiedia erythrophloia) | EF468992 | EF468847 | EF468794 | EF468901 | [10] | |
| C. polyarthra | MCA 996 | Lepidoptera | MF416597 | MF416543 | MF416487 | MF416644 | [4] | |
| C. pruinosa | ARSEF 5413 | Lepidoptera: Limacodidae | AY184979 | AY184968 | DQ522351 | DQ522397 | DQ522451 | [18] |
| C. pseudotenuipes | YFCC 8404 | Lepidoptera | OL468559 | OL468579 | OL473527 | OL739573 | OL473538 | [27] |
| C. tenuipes | TBRC 7265 | Lepidoptera (Pupa) | MF140707 | MF140827 | MF140776 | MF140800 | [1] | |
| Samsoniella alboaurantium | CBS 240.32 | Lepidoptera (Pupa) | JF415958 | JF415979 | JF416019 | JN049895 | JF415999 | [1] |
| S. alboaurantium | CBS 262.58 | Soil | MF416497 | MF416654 | MF416448 | [1] | ||
| S. alpina | YFCC 5818 | Hepialidae (Hepialus baimaensis) | MN576753 | MN576809 | MN576979 | MN576869 | MN576923 | [2] |
| S. alpina | YFCC 5831 | Hepialidae (Hepialus baimaensis) | MN576754 | MN576810 | MN576980 | MN576870 | MN576924 | [2] |
| S. antleroides | YFCC 6016 | Noctuidae (Larvae) | MN576747 | MN576803 | MN576973 | MN576863 | MN576917 | [2] |
| S. antleroides | YFCC 6113 | Noctuidae (Larvae) | MN576748 | MN576804 | MN576974 | MN576864 | MN576918 | [2] |
| S. aurantia | TBRC 7271 | Lepidoptera | MF140728 | MF140846 | MF140791 | MF140818 | [1] | |
| S. aurantia | TBRC 7272 | Lepidoptera | MF140727 | MF140845 | MF140817 | [1] | ||
| S. cardinalis | YFCC 5830 | Limacodidae (Pupa) | MN576732 | MN576788 | MN576958 | MN576848 | MN576902 | [2] |
| S. cardinalis | YFCC 6144 | Limacodidae (Pupa) | MN576730 | MN576786 | MN576956 | MN576846 | MN576900 | [2] |
| S. coccinellidicola | YFCC 8772 | Coccinellidae | ON563166 | ON621670 | ON676514 | ON676502 | ON568685 | This study |
| S. coccinellidicola | YFCC 8773 | Coccinellidae | ON563167 | ON621671 | ON676515 | ON676503 | ON568686 | This study |
| S. coleopterorum | A19501 | Curculionidae (Snout beetle) | MN101586 | MT642600 | MN101585 | [28] | ||
| S. cristata | YFCC 6021 | Saturniidae (Pupa) | MN576735 | MN576791 | MN576961 | MN576851 | MN576905 | [2] |
| S. cristata | YFCC 7004 | Saturniidae (Pupa) | MN576737 | MN576793 | MN576963 | MN576853 | MN576907 | [2] |
| S. farinospora | YFCC 8774 | Araneae(Spider) | ON563168 | ON621672 | ON676516 | ON676504 | ON568687 | This study |
| S. farinospora | YFCC 9051 | Lepidoptera:Hepialus | ON563169 | ON621673 | ON676517 | ON676505 | ON568688 | This study |
| S. haniana | YFCC 8769 | Lepidoptera(pupa) | ON563170 | ON621674 | ON676518 | ON676506 | ON568689 | This study |
| S. haniana | YFCC 8770 | Lepidoptera(pupa) | ON563171 | ON621675 | ON676519 | ON676507 | ON568690 | This study |
| S. haniana | YFCC 8771 | Lepidoptera(pupa) | ON563172 | ON621676 | ON676520 | ON676508 | ON568691 | This study |
| S. hepiali | ICMM 82-2 | Fungi (O. sinensis) | MN576738 | MN576794 | MN576964 | MN576854 | MN576908 | [2] |
| S. hepiali | YFCC 661 | Fungi (O. sinensis) | MN576739 | MN576795 | MN576965 | MN576855 | MN576909 | [2] |
| S. hepiali | Cor-4 | Fungi (O. sinensis) | MN576743 | MN576799 | MN576969 | MN576859 | MN576913 | [2] |
| S. hymenopterorum | A19521 | Vespidae (Bee) | MN101588 | MT642603 | MT642604 | [28] | ||
| S. hymenopterorum | A19522 | Vespidae (Bee) | MN101591 | MN101589 | MN101590 | [28] | ||
| S. inthanonensis | TBRC 7915 | Lepidoptera (Pupa) | MF140725 | MF140849 | MF140790 | MF140815 | [1] | |
| S. kunmingensis | YHH 16002 | Lepidoptera (Pupa) | MN576746 | MN576802 | MN576972 | MN576862 | MN576916 | [2] |
| S. lanmaoa | YFCC 6148 | Lepidoptera (Pupa) | MN576733 | MN576789 | MN576959 | MN576849 | MN576903 | [2] |
| S. lanmaoa | YFCC 6193 | Lepidoptera (Pupa) | MN576734 | MN576790 | MN576960 | MN576850 | MN576904 | [2] |
| S. pseudogunii | GY407201 | Lepidoptera (Larvae) | MZ827010 | MZ855233 | MZ855239 | [29] | ||
| S. pseudogunii | GY407202 | Lepidoptera (Larvae) | MZ831865 | MZ855234 | MZ855240 | [29] | ||
| S. pseudotortricidae | YFCC 9052 | Lepidoptera(pupa) | ON563173 | ON621677 | ON676521 | ON676509 | ON568692 | This study |
| S. pseudotortricidae | YFCC 9053 | Lepidoptera(pupa) | ON563174 | ON621678 | ON676522 | ON676510 | ON568693 | This study |
| S. pupicola | DY101681 | Lepidoptera (Pupa) | MZ827009 | MZ855231 | MZ855237 | [29] | ||
| S. pupicola | DY101682 | Lepidoptera (Pupa) | MZ827635 | MZ855232 | MZ855238 | [29] | ||
| S. ramosa | YFCC 6020 | Limacodidae (Pupa) | MN576749 | MN576805 | MN576975 | MN576865 | MN576919 | [2] |
| S. sinensis | YFCC 8766 | Lepidoptera(Larvae) | ON563175 | ON621679 | ON676523 | ON676511 | ON568694 | This study |
| S. sinensis | YFCC 8767 | Dermaptera | ON563176 | ON621680 | ON676524 | ON676512 | ON568695 | This study |
| S. sinensis | YFCC 8768 | Dermaptera | ON563177 | ON621681 | ON676525 | ON676513 | ON568696 | This study |
| S. tortricidae | YFCC 6013 | Tortricidae (Pupa) | MN576751 | MN576807 | MN576977 | MN576867 | MN576921 | [2] |
| S. tortricidae | YFCC 6131 | Tortricidae (Pupa) | MN576750 | MN576806 | MN576976 | MN576866 | MN576920 | [2] |
| S. yunnanensis | YFCC 1527 | Fungi (Cordyceps cicadae) | MN576756 | MN576812 | MN576982 | MN576872 | MN576926 | [2] |
| S. yunnanensis | YFCC 1824 | Fungi (Cordyceps cicadae) | MN576757 | MN576813 | MN576983 | MN576873 | MN576927 | [2] |
| Simplicillium formicae | MFLUCC 18-1379 | Hymenoptera: Formicidae | MK765046 | MK766512 | MK926451 | MK882623 | [30] | |
| S. lamellicola | CBS 116.25 | Fungi (Agaricus bisporus) | AF339601 | AF339552 | DQ522356 | DQ522404 | DQ522462 | [18] |
| S. lanosoniveum | CBS 704.86 | Fungi (Hemileia vastatrix) | AF339602 | AF339553 | DQ522358 | DQ522406 | DQ522464 | [18] |
| S. lanosoniveum | CBS 101267 | Fungi (Hemileia vastatrix) | AF339603 | AF339554 | DQ522357 | DQ522405 | DQ522463 | [18] |
| S. obclavatum | CBS 311.74 | Air above sugarcane field | AF339567 | AF339517 | EF468798 | [10] | ||
| S. yunnanense | YFCC 7133 | Fungi (A. waltergamsii) | MN576728 | MN576784 | MN576954 | MN576844 | [2] | |
| Trichoderma deliquescens | ATCC 208838 | On decorticated conifer wood | AF543768 | AF543791 | AF543781 | AY489662 | DQ522446 | [31] |
| T. stercorarium | ATCC 62321 | Cow dung | AF543769 | AF543792 | AF543782 | AY489633 | EF469103 | [31] |
| Species | Stromata (mm) | Fertile Part (mm) | Perithecia (μm) | Asci (μm) | Ascospores (μm) | Reference |
|---|---|---|---|---|---|---|
| Samsoniella antleroides | fasciculate, antler-like, cylindrical to clavate, long 22.3–57.8, oblate terminal branches, long 4.6–26.2 | clavate to fake-like, lateral sides have a longitudinal ditch without producing perithecia, 6.3–9.5 × 0.6–2.3 | superficial, fusiform, 294–442 × 131–216 | cylindrical, 8-spored, 160–248 × 2.1–2.7 | bola-shaped, septate, 110–184 × 0.8–1.3 | [2] |
| S. cardinalis | several, cylindrical, long 11.5–18.6 | clavate, lateral sides have a longitudinal ditch without producing perithecia, 2.5–6.8 × 0.5–2.6 | superficial, oblong-ovate to fusiform, 370–485 × 140–238 | cylindrical, 8-spored, 163–320 × 3.2–4.3 | bola-shaped, septate, 165–230 × 0.5–0.9 | [2] |
| S. cristata | solitary or two, crista-like, long 25–40, much branched | crista-like or subulate, 3.1–18.5 × 0.9–8.0 | superficial, narrowly ovoid, 370–485 × 150–245 | cylindrical, 8-spored, 180–356 × 3.0–4.8 | bola-shaped, septate, 155–290 × 1.0–1.3 | [2] |
| S. inthanonensis | gregarious, cylindrical to clavate, long 20–50, 1–1.5 broad | 8–15 long, 1.5–2 broad | superficial, ovoid, (380–)417.5–474.5(–500) × (150–)205–260(–265) | cylindrical, 8-spored, 300 × 2–2.5 | bola-shaped, 3 or 4septate, 221.5–267 × 0.5–1 | [1] |
| S. kunmingensis | solitary, cylindrical to clavate, long 23, bifurcated, | clavate, lateral sides usually have a longitudinal ditch without producing perithecia, 3.3–4.2 × 0.8–1.2 | superficial, narrowly ovoid to fusiform, 330–395 × 110–185 | cylindrical, 8-spored, 150–297 × 3.0–4.6 | bola-shaped, septate, 127–190 × 0.8–1.5 | [2] |
| S. pseudotortricidae | solitary to several, long 20–65, unbranched or dichotomous | clavate to subulate, lateral side usually have a longitudinal section without producing perithecia, 10–17 × 1.5–4.2 | superficial, narrowly ovoid to fusiform, 285.7–313.2 × 149.2–154.9 | This study | ||
| S. lanmaoa | two to five, orange, long 38–69, palmately branched | clavate, lateral sides usually have a longitudinal ditch without producing perithecia, 8.5–11.2 × 0.6–2.3 | superficial, narrowly ovoid to fusiform, 360–467 × 124–210 | cylindrical, 8-spored, 160–325 × 3.3–4.8 | bola-shaped, septate, 135–260 × 0.9–1.4 | [2] |
| S. ramosa | fascicular, 15–32 × 0.8–1.5, multi-branched, often confluent at the base | having no obvious boundary with stipes | superficial, narrowly ovoid to fusiform, 340–435 × 130–197 | [2] | ||
| S. tortricidae | gregarious, long 25–60, unbranched or dichotomous | clavate to subulate, lateral side usually has a longitudinal section without producing perithecia, 5–15 × 1.2–2.3 | superficial, narrowly ovoid to fusiform, 350–468 × 140–225 | cylindrical, 8-spored, 170–285 × 2.8–4.0 | bola-shaped, septate, 120–235 × 0.8–1.3 | [2] |
| Species | Synnemata (mm) | Conidiophores (μm) | Phialides | Phialides Size (μm) | Conidia (μm) | References |
|---|---|---|---|---|---|---|
| Samsoniella alboaurantium | 30–400 × 2–2.5 | 5–8 × 2, tapering fairly abruptly at the tip | ovate to lemon-shaped, 2.3–2.5(–3) × 1.5–1.8 | [32] | ||
| S. alpina | irregularly branched, 3–20 long, cylindrical or clavate stipes with white powdery heads | 3.1–6.5 × 1.6–2.8 | verticillate on conidiophores, solitary or verticillate on hyphae | 4.7–9.5 × 1.9–3.1, wide (apex) 0.5–1.1, basal portion cylindrical to narrowly lageniform, tapering abruptly toward the apex | fusiform or oval, 2.0–3.1 × 1.3–2.1 | [2] |
| S. antleroides | 3.5–9.7 × 1.3–3.2 | verticillate, in whorls of 2 to 9, sometimes solitary on hyphae | 3.5–16.3 × 1.7–2.9, wide (apex) 0.5–1.0, basal portion cylindrical to narrowly lageniform, tapering abruptly toward the apex | fusiform or oval, 2.3–3.5 × 1.6–2.5 | [2] | |
| S. aurantia | irregularly branched starting 15–40 above the ground and continuously to the apex, 25–75 × 1–1.5 | verticillate, in whorls of 2 to 4 | (5–)5.5–8.5(–13) × 2–3, basal portion cylindrical to ellipsoidal, neck 2–4 × 1 | fusiform, (2–)2.5–3.5(–4) × (1–)1.5(–2) | [1] | |
| S. cardinalis | 3.1–9.5 × 1.3–2.0 | verticillate, in whorls of 2 to 5, sometimes solitary on hyphae | 4.1–43.5 × 1.3–2.4, wide (apex) 0.6–1.2, basal portion cylindrical to narrowly lageniform, tapering gradually or abruptly toward the apex | fusiform or oval, 2.4–3.2 × 1.4–2.2 | [2] | |
| S. coccinellidicola | Irregularly branched, starting 2–2.5 above the cocoons of insect host, 15–25 × 0.8–1.2 | 4.8–15 × 1.0–1.9 | verticillate, usually in whorls of 2 to 5, or solitary on hyphae | 6.0–14.1 × 1.0–2.0 wide (apex) 0.3–0.8, basal portion cylindrical to narrowly lageniform, tapering gradually or abruptly toward the apex | fusiform or oval, 1.8–3.0 × 1.3–2.0 | This study |
| S. coleopterorum | verticillate, in whorls of 2 to 4 | 5.4–9.7 × 1.2–1.8, a cylindrical to ellipsoidal basal portion, tapering into a short distinct neck | fusiform, ellipsoidal or subglobose, 1.7–2.5 × 1.2–1.8 | [28] | ||
| S. cristata | 3.6–11.5 × 1.7–2.5 | verticillate, in whorls of 2 to 5, usually solitary on hyphae | 4.5–23.2 × 1.6–2.7, wide (apex) 0.5–1.1, basal portion cylindrical to narrowly lageniform, tapering gradually or abruptly toward the apex | fusiform or oval, 2.4–3.2 × 1.6–2.3 | [2] | |
| S. farinospora | 2.4–14.0 × 0.9–1.8 | verticillate, usually in whorls of 2 to 4, or solitary on hyphae | 3.0–13.5 × 0.6–1.6, basal portion cylindrical to narrowly lageniform, tapering gradually or abruptly toward the apex | oblong to cylindrical, 1.6–2.8 × 0.6–1.2 | This study | |
| S. haniana | usually unbranched or irregularly branched at the apex, 20–40 × 1–1.8 | 3.8–10.2 × 1.1–2.9 | verticillate, usually in whorls of 2 to 5, or solitary on hyphae | 5.4–12.1 × 1.2–2.9, wide (apex) 0.3–1.1 basal portion cylindrical to narrowly lageniform, tapering gradually or abruptly toward the apex | fusiform or oval,2.3–3.7 × 1.2–2.8 | This study |
| S. hepiali | branched or unbranched, 5–41 long | 4.0–7.6 × 1.4–2.2 | verticillate, in whorls of 2 to 5, solitary or opposite on hyphae | 3.5–13.6 × 1.3–2.1, wide (apex) 0.5–1.0, basal portion cylindrical to narrowly lageniform, tapering gradually or abruptly toward the apex | fusiform or oval, 1.8–3.3 × 1.4–2.2 | [2] |
| S. hymenopterorum | Verticillate, in whorls of 3 to 4 | 6.5–10.6 × 1.2–2.0, a cylindrical basal portion, tapering to a distinct neck | fusiform to ovoid, 1.9–2.5 × 1.5–2.1 | [28] | ||
| S. inthanonensis | verticillate in whorls of 2 to 5, sometimes solitary on hyphae | (4–)6.5–10(–12) × (1–)1.5–2(–3), cylindrical basal portion, tapering into a long neck, (1–)2.5(–4) × 0.5–1 | short fusiform, (2–)3(–3.5) × 1.5–2 | [1] | ||
| S. lanmaoa | 3.8–13.3 × 1.5–2.1 | verticillate, in whorls of 2 to 6, usually solitary on hyphae | 3.5–20.7 × 1.7–2.6, wide (apex) 0.5–1.1, basal portion cylindrical to narrowly lageniform, tapering gradually or abruptly toward the apex | fusiform or oval, 1.9–2.7 × 1.4–2.0 | [2] | |
| S. lepidopterorum | Verticillate, in whorls of 2 to 4 | 5.2–8.5 (–13.1) × 1.1–1.7, ellipsoidal basal portion, tapering into a distinct neck | fusiform to subglobose, 2.0–2.5 × 1.2–2.0 | [28] | ||
| S. pseudotortricidae | 6.6–26.5 × 1.1–2.5 | verticillate, in whorls of 2 to 5, usually solitary on hyphae | 5.4–6.9 × 1.0–1.6, wide (apex) 0.5–0.8 basal portion cylindrical to narrowly lageniform, tapering gradually or abruptly toward the apex | fusiform or oval, 0.9–1.5 × 0.8–1.3 | This study | |
| S. pseudogunnii | solitary or in whorls of 2 to 9 | 6.8–11.0 × 2.2–2.4, cylindrical basal portion, tapering into a short distinct neck | fusiform 2.8–3.2 × 1.7–2.1 | [29] | ||
| S. pupicola | solitary or in whorls of 2 to 9 | 7.0–9.2 × 2.5–3.3, a cylindrical basal portion, tapering into a short distinct neck | fusiform, 2.5–3.3 × 2.2–2.6 | [29] | ||
| S. ramosa | 4.3–10.5 × 1.3–2.4 | verticillate, in whorls of 2 to 6, usually solitary on hyphae | 5.3–14.6 × 1.3–2.8, wide (apex) 0.6–1.2, basal portion cylindrical to narrowly lageniform, tapering gradually or abruptly toward the apex | fusiform or oval, 2.0–3.6 × 1.5–2.2 | [2] | |
| S. sinensis | 3.5–5long, branched, conidia in abundance at the apex. | 6.4–10.5 × 1.7–2.1 | verticillate, in whorls of 2 to 5, sometimes solitary on hyphae | 5.6–9.3 × 1.5–2.1, wide (apex) 0.6–1.0 basal portion cylindrical to narrowly lageniform, tapering gradually or abruptly toward the apex | spherical, elliptical or fusiform, 2.0–3.1 × 1.3–1.9 | This study |
| S. tortricidae | 4.2–12.5 × 1.4–2.4 | verticillate, in whorls of 2 to 5, usually solitary on hyphae | 3.6–42.4 × 1.1–2.6, wide (apex) 0.4–0.9, basal portion cylindrical to narrowly lageniform, tapering gradually or abruptly toward the apex | fusiform or oval, 2.1–3.0 × 1.3–1.7 | [2] | |
| S. yunnanensis | gregarious, flexuous, fleshy, 4–18 long, with terminal branches of 3–7 × 1.0–2.0 | 4.2–23.5 × 1.4–2.3 | verticillate, in whorls of 2 to 7, usually solitary on hyphae | 4.5–11.6 × 1.2–2.4, wide (apex) 0.6–1.0, basal portion cylindrical to narrowly lageniform, tapering gradually or abruptly toward the apex | fusiform or oval, 2.0–3.3 × 1.1–2.2 | [2] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Wang, Y.; Dong, Q.; Fan, Q.; Dao, V.-M.; Yu, H. Morphological and Phylogenetic Characterization Reveals Five New Species of Samsoniella (Cordycipitaceae, Hypocreales). J. Fungi 2022, 8, 747. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8070747
Wang Z, Wang Y, Dong Q, Fan Q, Dao V-M, Yu H. Morphological and Phylogenetic Characterization Reveals Five New Species of Samsoniella (Cordycipitaceae, Hypocreales). Journal of Fungi. 2022; 8(7):747. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8070747
Chicago/Turabian StyleWang, Zhiqin, Yao Wang, Quanying Dong, Qi Fan, Van-Minh Dao, and Hong Yu. 2022. "Morphological and Phylogenetic Characterization Reveals Five New Species of Samsoniella (Cordycipitaceae, Hypocreales)" Journal of Fungi 8, no. 7: 747. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8070747