
BEM2, a RHO GTPase Activating Protein That Regulates Morphogenesis in S. cerevisiae, Is a Downstream Effector of Fungicidal Action of Fludioxonil


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Screening of S. cerevisiae Strain BY4742 with mTn3-LacZ/LEU2 Mutagenized Library
2.3. Construction of BEM2 Mutants
2.4. Construction of MPK1 Mutants
2.5. Relative Growth Assay of mpk1 Mutants
2.6. Western Blotting
3. Results
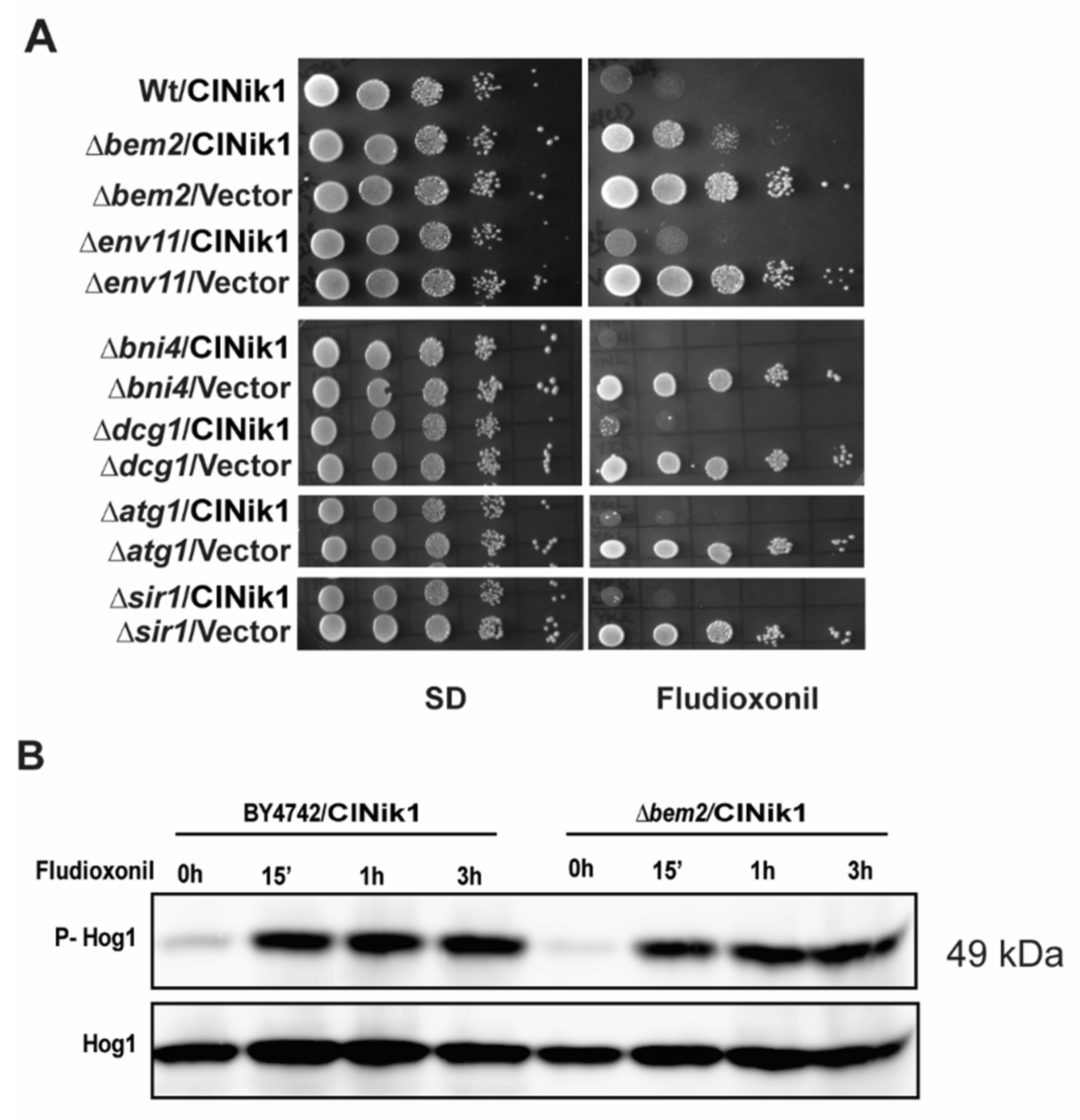
3.1. Identification of Fludioxonil Resistant bem2 Mutant Strain through mTn3-LacZ/LEU2 Mutagenized Library
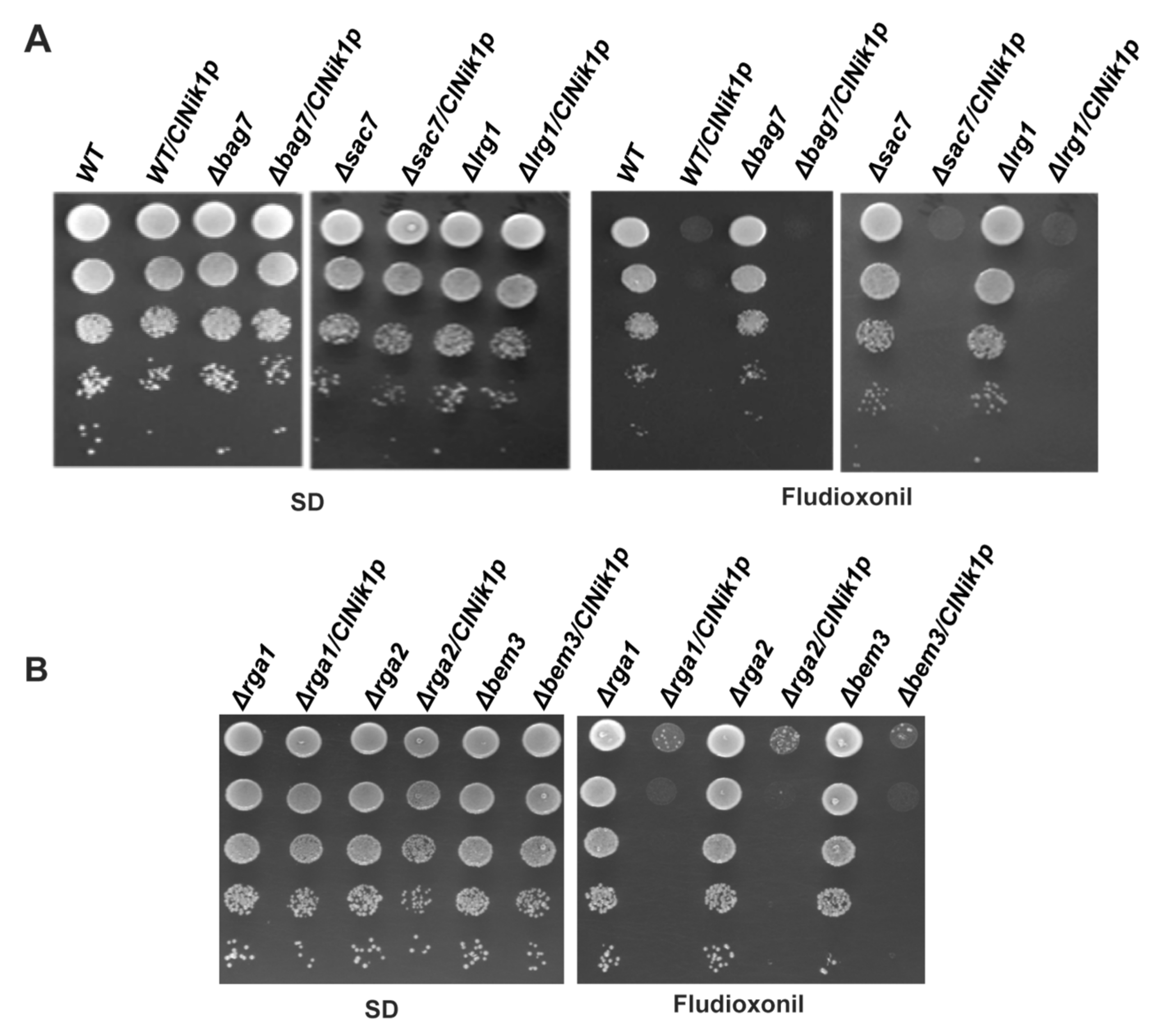
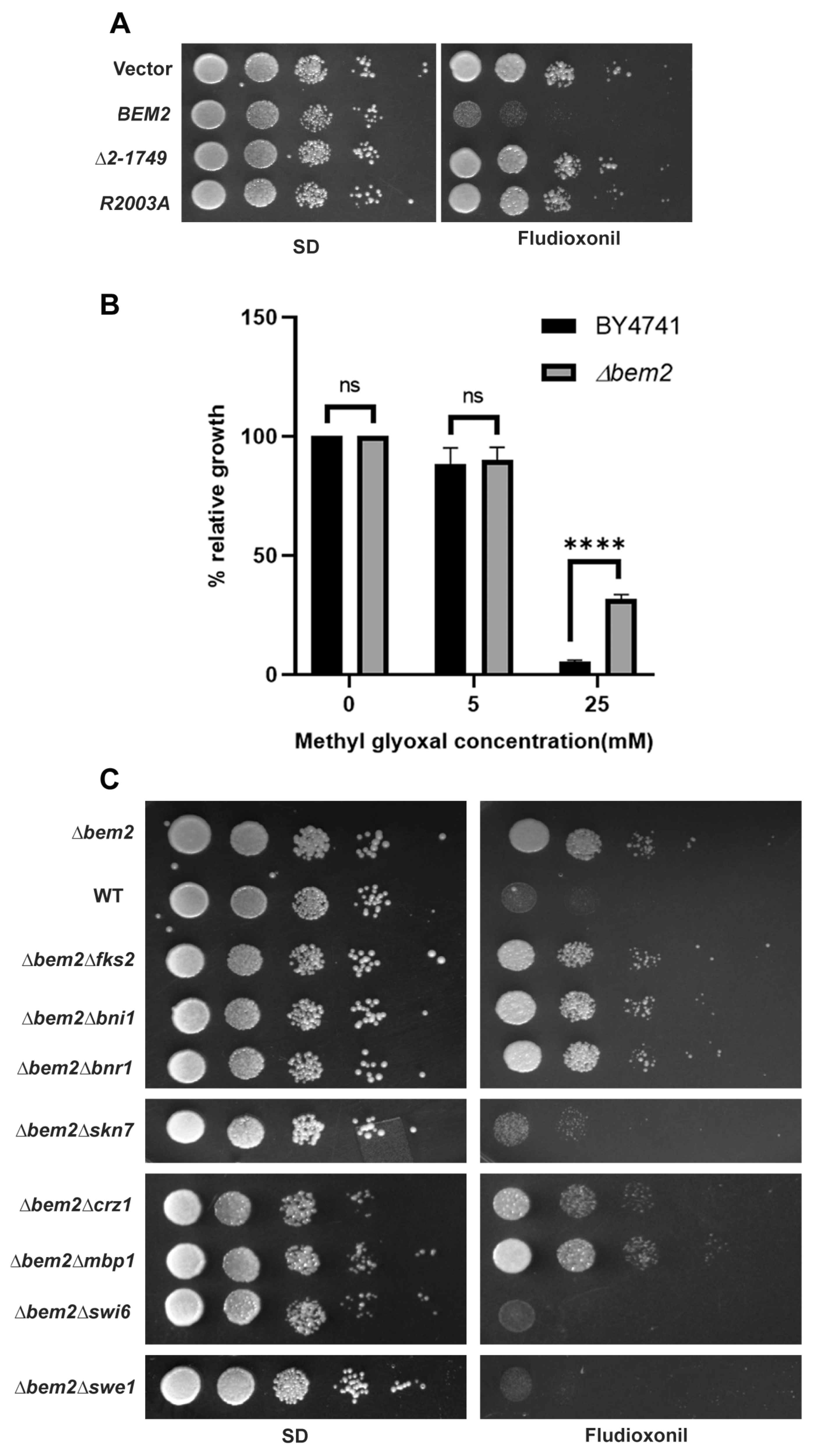
3.2. The Role of BEM2 in the Antifungal Activity of Fludioxonil
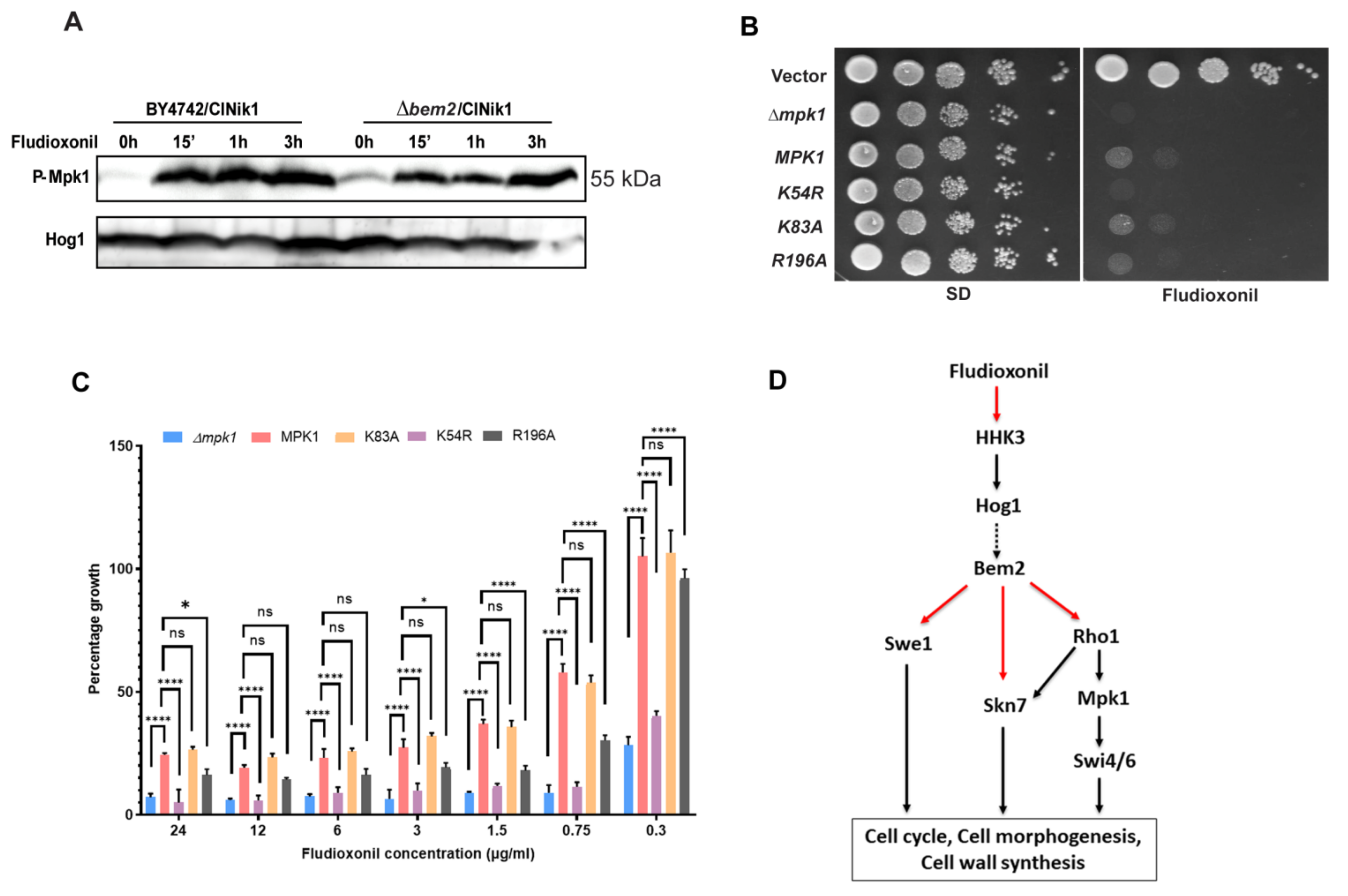
3.3. The Activation of CWI Pathway upon Fludioxonil Treatment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kilani, J.; Fillinger, S. Phenylpyrroles: 30 years, two molecules and (nearly) no resistance. Front. Microbiol. 2016, 7, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, T.; Izumitsu, K. Two-Component signaling system in filamentous fungi and the mode of action of dicarboximide and phenylpyrrole fungicides. In Fungicides; Carisse, O., Ed.; InTech: London, UK, 2010; ISBN 978-953-307-266-1. [Google Scholar]
- Bersching, K.; Jacob, S. The Molecular Mechanism of Fludioxonil Action Is Different to Osmotic Stress Sensing. J. Fungi 2021, 7, 393. [Google Scholar] [CrossRef] [PubMed]
- Brandhorst, T.T.; Kean, I.R.L.; Lawry, S.M.; Wiesner, D.L.; Klein, B.S. Phenylpyrrole fungicides act on triosephosphate isomerase to induce methylglyoxal stress and alter hybrid histidine kinase activity. Sci. Res. 2019, 9, 5047. [Google Scholar] [CrossRef]
- Cui, W.; Beever, R.E.; Parkes, S.L.; Weeds, P.L.; Templeton, M.D. An osmosensing histidine kinase mediates dicarboximide fungicide resistance in Botryotinia fuckeliana (Botrytis cinerea). Fungal Genet. Biol. 2002, 36, 187–198. [Google Scholar] [CrossRef]
- Fujimura, M.; Ochiai, N.; Ichiishi, A.; Usami, R.; Horikoshi, K.; Yamaguchi, I. Fungicide resistance and osmotic stress sensitivity in os mutants of Neurospora crassa. Pestic. Biochem. Phys. 2000, 67, 125–133. [Google Scholar] [CrossRef]
- Motoyama, T.; Ohira, T.; Kadokura, K.; Ichiishi, A.; Fujimura, M.; Yamaguchi, I.; Kudo, T. An Os-1 family histidine kinase from a filamentous fungus confers fungicide-sensitivity to yeast. Curr. Genet. 2005, 47, 298–306. [Google Scholar] [CrossRef]
- Yoshimi, A.; Tsuda, M.; Tanaka, C. Cloning and characterization of the histidine kinase gene Dic1 from Cochliobolus heterostrophus that confers dicarboximide resistance and osmotic adaptation. Mol. Genet. Genom. 2004, 271, 228–236. [Google Scholar] [CrossRef]
- Zhang, Y.; Lamm, R.; Pillonel, C.; Lam, S.; Xu, J.R. Osmoregulation and fungicide resistance: The Neurospora crassa os-2 gene encodes a HOG1 mitogen-activated protein kinase homologue. Appl. Environ. Microbiol. 2002, 68, 532–538. [Google Scholar] [CrossRef] [Green Version]
- Izumitsu, K.; Yoshimi, A.; Hamada, S.; Morita, A.; Saitoh, Y.; Tanaka, C. Dic2 and Dic3 loci confer osmotic adaptation and fungicidal sensitivity independent of the HOG pathway in Cochliobolus heterostrophus. Mycol. Res. 2009, 113 Pt 10, 1208–1215. [Google Scholar] [CrossRef]
- Bahn, Y.S.; Kojima, K.; Cox, G.M.; Heitman, J. A unique fungal two-component system regulates stress responses, drug sensitivity, sexual development, and virulence of Cryptococcus neoformans. Mol. Biol. Cell 2006, 17, 3122–3135. [Google Scholar] [CrossRef] [Green Version]
- Buschart, A.; Gremmer, K.; El-Mowafy, M.; van den Heuvel, J.; Mueller, P.P.; Bilitewski, U. A novel functional assay for fungal histidine kinases group III reveals the role of HAMP domains for fungicide sensitivity. J. Biotechnol. 2012, 157, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Randhawa, A.; Kaur, H.; Mondal, A.K.; Hohmann, S. Fungal fludioxonil sensitivity is diminished by a constitutively active form of the group III histidine kinase. FEBS Lett. 2012, 586, 2417–2422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randhawa, A.; Kundu, D.; Sharma, A.; Prasad, R.; Mondal, A.K. Overexpression of the CORVET complex alleviates the fungicidal effects of fludioxonil on the yeast Saccharomyces cerevisiae expressing hybrid histidine kinase 3. J. Biol. Chem. 2019, 294, 461–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gietz, R.D.; Woods, R.A. Transformation of yeast by lithium acetate/single-stranded carrier DNA/polyethylene glycol method. Methods Enzymol. 2002, 350, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Burns, N.; Grimwade, B.; Ross-Macdonald, P.B.; Choi, E.Y.; Finberg, K.; Roeder, G.S.; Snyder, M. Large-scale analysis of gene expression, protein localization, and gene disruption in Saccharomyces cerevisiae. Genes Dev. 1994, 8, 1087–1105. [Google Scholar] [CrossRef] [Green Version]
- Sikorski, R.S.; Hieter, P. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae. Genetics 1989, 122, 19–27. [Google Scholar] [CrossRef]
- Gueldener, U.; Heinisch, J.; Koehler, G.J.; Voss, D.; Hegemann, J.H. A second set of loxP marker cassettes for Cre-mediated multiple gene knockouts in budding yeast. Nucleic Acids Res. 2002, 30, e23. [Google Scholar] [CrossRef] [Green Version]
- Meena, N.; Kaur, H.; Mondal, A.K. Interactions among HAMP Domain repeats act as an osmosensing molecular switch in group III hybrid histidine kinases from fungi. J. Biol. Chem. 2010, 285, 12121–12132. [Google Scholar] [CrossRef] [Green Version]
- Randhawa, A.; Chawla, S.; Mondal, A.K. Functional dissection of HAMP domains in NIK1 ortholog from pathogenic yeast Candida lusitaniae. Gene 2016, 577, 251–257. [Google Scholar] [CrossRef]
- Bender, A.; Pringle, J.R. Multicopy suppression of the cdc24 budding defect in yeast by CDC42 and three newly identified genes including the ras-related gene RSR1. Proc. Natl. Acad. Sci. USA 1989, 86, 9976–9980. [Google Scholar] [CrossRef] [Green Version]
- Gong, T.; Liao, Y.; He, F.; Yang, Y.; Yang, D.D.; Chen, X.D.; Gao, X.D. Control of polarized growth by the Rho family GTPase Rho4 in budding yeast: Requirement of the N-terminal extension of Rho4 and regulation by the Rho GTPase-activating protein Bem2. Eukaryot. Cell 2013, 12, 368–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.J.; Francisco, L.; Chen, G.C.; Marcotte, E.; Chan, C.S. Control of cellular morphogenesis by the Ip12/Bem2 GTPase-activating protein: Possible role of protein phosphorylation. J. Cell Biol. 1994, 127, 1381–1394. [Google Scholar] [CrossRef] [PubMed]
- Knaus, M.; Pelli-Gulli, M.P.; van Drogen, F.; Springer, S.; Jaquenoud, M.; Peter, M. Phosphorylation of Bem2p and Bem3p may contribute to local activation of Cdc42p at bud emergence. EMBO J. 2007, 26, 4501–4513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marquitz, A.R.; Harrison, J.C.; Bose, I.; Zyla, T.R.; McMillan, J.N.; Lew, D.J. The Rho-GAP Bem2p plays a GAP-independent role in the morphogenesis checkpoint. EMBO J. 2002, 21, 4012–4025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, J.; Zheng, Y.; Bender, L.; Myers, A.; Cerione, R.; Bender, A. Interactions between the bud emergence proteins Bem1p and Bem2p and Rho-type GTPases in yeast. J. Cell Biol. 1994, 127, 1395–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, D.E. Regulation of cell wall biogenesis in Saccharomyces cerevisiae: The cell wall integrity signaling pathway. Genetics 2011, 189, 1145–1175. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, P.; Nishikawa, J.L.; Breitkreutz, B.J.; Tyers, M. Systematic identification of pathways that couple cell growth and division in yeast. Science 2002, 297, 395–400. [Google Scholar] [CrossRef]
- Kojima, K.; Bahn, Y.S.; Heitman, J. Calcineurin, Mpk1 and Hog1 MAPK pathways independently control fludioxonil antifungal sensitivity in Cryptococcus neoformans. Microbiology 2006, 152 Pt 3, 591–604. [Google Scholar] [CrossRef] [Green Version]
- Schruefer, S.; Bohmer, I.; Dichtl, K.; Spadinger, A.; Kleinemeier, C.; Ebel, F. The response regulator Skn7 of Aspergillus fumigatus is essential for the antifungal effect of fludioxonil. Sci. Rep. 2021, 11, 5317. [Google Scholar] [CrossRef]
- Baetz, K.; Moffat, J.; Haynes, J.; Chang, M.; Andrews, B. Transcriptional co-regulation by the cell integrity mitogen-activated protein kinase Slt2 and the cell cycle regulator Swi4. Mol. Cell. Biol. 2001, 21, 6515–6528. [Google Scholar] [CrossRef] [Green Version]
- Dodou, E.; Treisman, R. The Saccharomyces cerevisiae MADS-box transcription factor Rlm1 is a target for the Mpk1 mitogen-activated protein kinase pathway. Mol. Cell. Biol. 1997, 17, 1848–1859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madden, K.; Sheu, Y.J.; Baetz, K.; Andrews, B.; Snyder, M. SBF cell cycle regulator as a target of the yeast PKC-MAP kinase pathway. Science 1997, 275, 1781–1784. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Takaesu, G.; Hagiwara, M.; Irie, K.; Matsumoto, K. Characterization of a serum response factor-like protein in Saccharomyces cerevisiae, Rlm1, which has transcriptional activity regulated by the Mpk1 (Slt2) mitogen-activated protein kinase pathway. Mol. Cell. Biol. 1997, 17, 2615–2623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truman, A.W.; Kim, K.Y.; Levin, D.E. Mechanism of Mpk1 mitogen-activated protein kinase binding to the Swi4 transcription factor and its regulation by a novel caffeine-induced phosphorylation. Mol. Cell. Biol. 2009, 29, 6449–6461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkins, B.D.; Yoshida, S.; Saito, K.; Wu, C.F.; Lew, D.J.; Pellman, D. Inhibition of Cdc42 during mitotic exit is required for cytokinesis. J. Cell Biol. 2013, 202, 231–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cid, V.C.J.; Adamikova, L.; Sanchez, M.; Molina, M.A.; Nombela, C. Cell cycle control of septin ring dynamics in the budding yeast. Microbiology 2001, 147, 1437–1450. [Google Scholar] [CrossRef] [Green Version]
- Cid, V.J.; Cenamor, R.; Sanchez, M.; Nombela, C. A mutation in the Rho1-GAP-encoding gene BEM2 of Saccharomyces cerevisiae affects morphogenesis and cell wall functionality. Microbiology 1998, 144, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Seeley, E.S.; Kato, M.; Margolis, N.; Wickner, W.; Eitzen, G. Genomic analysis of homotypic vacuole fusion. Mol. Biol. Cell 2002, 13, 782–794. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Bretscher, A. The rho-GAP encoded by BEM2 regulates cytoskeletal structure in budding yeast. Mol. Biol. Cell 1995, 6, 1011–1024. [Google Scholar] [CrossRef]
- Sia, R.A.; Bardes, E.S.; Lew, D.J. Control of Swe1p degradation by the morphogenesis checkpoint. EMBO J. 1998, 17, 6678–6688. [Google Scholar] [CrossRef] [Green Version]
- Randhawa, A.; Mondal, A.K. The sixth HAMP domain negatively regulates the activity of the group III HHK containing seven HAMP domains. Biochem. Biophys. Res. Commun. 2013, 438, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Borah, S.; Shivarathri, R.; Kaur, R. The Rho1 GTPase-activating protein CgBem2 is required for survival of azole stress in Candida glabrata. J. Biol. Chem. 2001, 286, 34311–34324. [Google Scholar] [CrossRef] [Green Version]
- Martin, H.; Rodriguez-Pachon, J.M.; Ruiz, C.; Nombela, C.; Molina, M. Regulatory mechanisms for modulation of signaling through the cell integrity Slt2-mediated pathway in Saccharomyces cerevisiae. J. Biol. Chem. 2000, 275, 1511–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roncero, C.; Celador, R.; Sánchez, N.; García, P.; Sánchez, Y. The Role of the Cell Integrity Pathway in Septum Assembly in Yeast. J. Fungi 2021, 7, 729. [Google Scholar] [CrossRef]
- Mumberg, D.; Muller, R.; Funk, M. Yeast vectors for the controlled expression of heterologous proteins in different genetic backgrounds. Gene 1995, 156, 119–122. [Google Scholar] [CrossRef]
- Christianson, T.W.; Sikorski, R.S.; Dante, M.; Shero, J.H.; Hieter, P. Multifunctional yeast high-copy-number shuttle vectors. Gene 1992, 110, 119–122. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence |
|---|---|
| For cloning BEM2 and its mutants | |
| Bem2 PF | CGCTCGAGCGAGACGAAGAGCTGAGCACCAAGAG |
| Bem2 Kas R | GGCGCCGATTTGGATTTTTGCCATA |
| Bem2 Kas F | GGCGCCGCGGAAGCTATACTACATT |
| Bem2 ORF R | CGAGCTCGTTATTGCTTGAAATAATCATTTGGATTCT |
| Bem2 Sph1 F | GCATGCCATAATATCACCAGAGTCTAG |
| R2003A F | GTGGGATTGTACGCTATTCCTGGTTCCATCGG |
| R2003A R | GATGGAACCAGGAATAGCGTACAATCCCACTTCATCT |
| Δ2-1749 F | AGGAGTCTAATGATCTTGAAGAACTCTGCTGCTTTAC |
| Δ2-1749 R | AGAGTTCTTCAAGATCATTAGACTCCTGCTTCGTTATTT |
| For disruption of BEM2 | |
| Bem2 dis F | CTGGATAGACACAAAAAAAACAAATAACGAAGCAGGAGTCTAATGAAAGGCAGCTGAAGCTTCGTACGC |
| Bem2 dis R | CTCTCTCAGCAGTGGATTGTATACATTTACCACGAAAATTGTTTATTGCTGCATAGGCCACTAGTGGATCTG |
| Bem2 conf F | CACTGGTACTGTCTGCTAACTCAAA |
| Bem2 conf R | AAACATAACATTCAAAAGGCAAGAG |
| Lacz F | GAGGTCGACGGTATCGATAAGC |
| Lacz R | CCCAGTCACGACGTTGTAAAAC |
| For cloning MPK1 and its mutants | |
| MPK1 F | CCCTCGAGGGCGGTAACTATGGACACCTTACAGG |
| MPK1 R | CGGGATCCCGACGAGCTACAACAAGAGCACGTA |
| K54R F | CAGTTGCCATCAGAAAAGTGACAAACGTTTTTTCGA |
| K54R R | GTTTGTCACTTTTCTGATGGCAACTGTGGTATCTTCG |
| K83A F | TCAGAGGCCACGCTAATATTACATGTCTTTATGATATGGATATTG |
| K83A R | CATGTAATATTAGCGTGGCCTCTGAAATGTCTCAAA |
| R196A F | ACGTGGCCACTGCTTGGTATAGAGCTCCGGAAATAATG |
| R196A R | 5 GCTCTATACCAAGCAGTGGCCACGTACTCCGTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, A.; Martoliya, Y.; Mondal, A.K. BEM2, a RHO GTPase Activating Protein That Regulates Morphogenesis in S. cerevisiae, Is a Downstream Effector of Fungicidal Action of Fludioxonil. J. Fungi 2022, 8, 754. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8070754
Sharma A, Martoliya Y, Mondal AK. BEM2, a RHO GTPase Activating Protein That Regulates Morphogenesis in S. cerevisiae, Is a Downstream Effector of Fungicidal Action of Fludioxonil. Journal of Fungi. 2022; 8(7):754. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8070754
Chicago/Turabian StyleSharma, Anupam, Yogita Martoliya, and Alok K. Mondal. 2022. "BEM2, a RHO GTPase Activating Protein That Regulates Morphogenesis in S. cerevisiae, Is a Downstream Effector of Fungicidal Action of Fludioxonil" Journal of Fungi 8, no. 7: 754. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8070754