
Genomic Based Analysis of the Biocontrol Species Trichoderma harzianum: A Model Resource of Structurally Diverse Pharmaceuticals and Biopesticides

Abstract
:1. Introduction
2. Materials and Methods
2.1. Trichoderma Species Genome Scan
2.2. Validation of the M10 v1.0 Isolate and Annotation of the Core Enzymes of Its Secondary Metabolites (SMs)
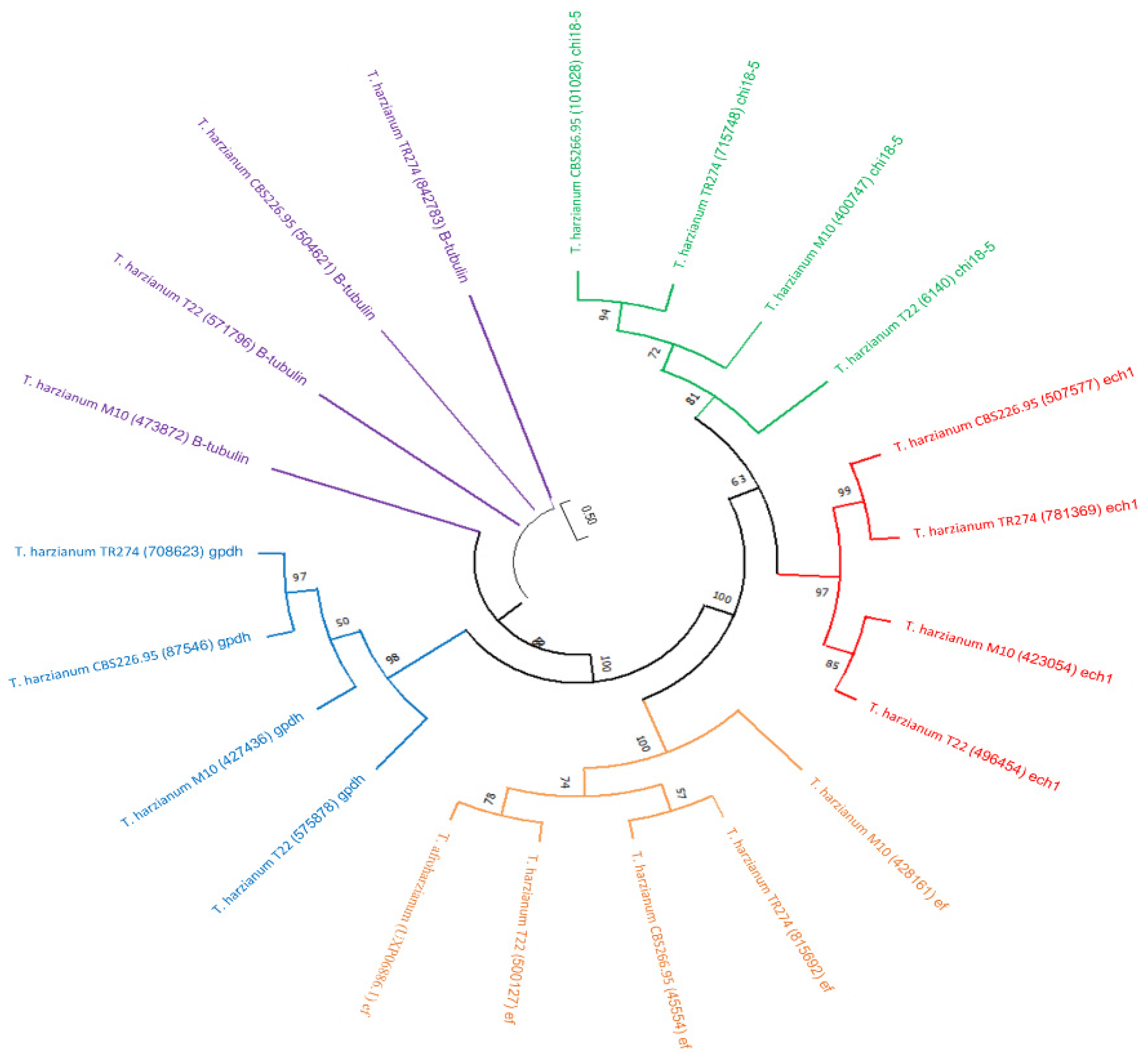
2.3. Secondary Metabolites Phylogenetic Analysis
2.4. Annotation of the Predicted Biosynthetic Gene Clusters
3. Results
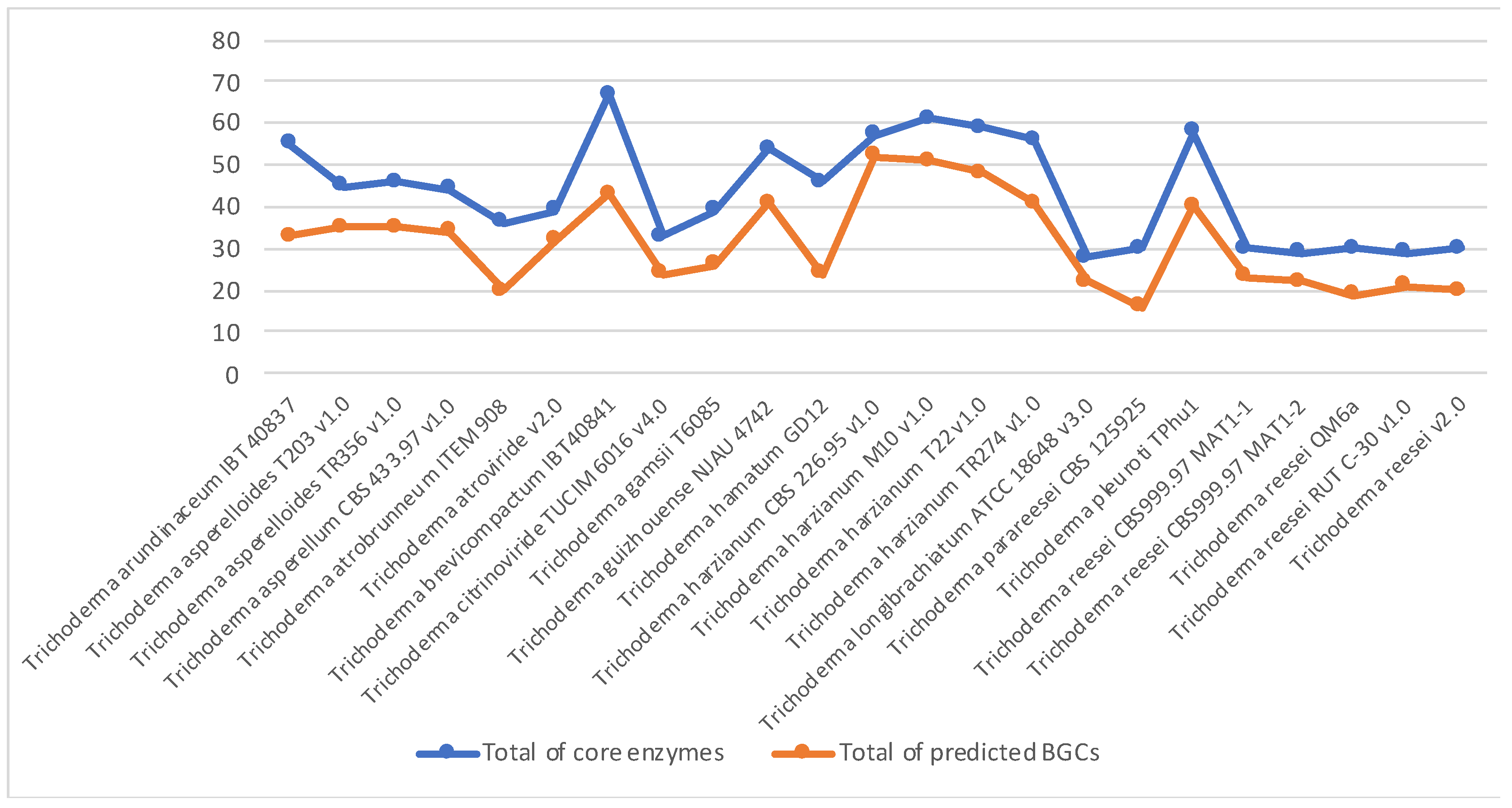
3.1. Genome Scan
3.2. Trichoderma Harzianum Genomes Comparison
3.2.1. NRPS and NRPS-Like Phylogenetic Analysis and Biosynthetic Gene Clusters Annotation
3.2.2. PKS and PKS-like Phylogenetic Analysis and Biosynthetic Gene Clusters Annotation
3.2.3. Terpene Cyclase Phylogenetic Analysis and Biosynthetic Gene Clusters Annotation
3.3. Clinker Investigation
3.3.1. Core Enzymes Cblaster
3.3.2. Biosynthetic Gene Clusters Cblaster
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adams, P.; De-Leij, F.A.; Lynch, J.M. Trichoderma harzianum Rifai 1295-22 mediates growth promotion of crack willow (Salix fragilis) saplings in both clean and metal-contaminated soil. Microb. Ecol. 2007, 54, 306–313. [Google Scholar] [CrossRef]
- Barratt, B.I.P.; Moran, V.C.; Bigler, F.; van Lenteren, J.C. The status of biological control and recommendations for improving uptake for the future. BioControl 2018, 63, 155–167. [Google Scholar] [CrossRef]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of action of microbial biological control agents against plant diseases: Relevance beyond efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, P.; Casadevall, A. Quorum sensing in fungi—A review. Med. Mycol. 2012, 50, 337–345. [Google Scholar] [CrossRef]
- Weindling, R.; Fawcett, H. Experiments in the control of Rhizoctonia damping-off of citrus seedlings. Hilgardia 1936, 10, 1–16. [Google Scholar] [CrossRef]
- Rossman, A.Y. The impact of invasive fungi on agricultural ecosystems in the United States. In Ecological Impacts of Non-Native Invertebrates and Fungi on Terrestrial Ecosystems; Springer: Dordrecht, The Netherlands, 2008; pp. 97–107. [Google Scholar]
- Meena, M.; Swapnil, P.; Zehra, A.; Aamir, M.; Dubey, M.K.; Goutam, J.; Upadhyay, R.S. Beneficial microbes for disease suppression and plant growth promotion. In Plant-Microbe Interactions in Agro-Ecological Perspective; Springer: Singapore, 2017; pp. 395–432. [Google Scholar]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of arbuscular mycorrhizal fungi in plant growth regulation: Implications in abiotic stress tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [PubMed]
- Kulimushi, S.M.; Muiru, W.M.; Mutitu, E.W. Potential of Trichoderma spp., Bacillus subtilis and Pseudomonas fluorescens in the management of early blight in tomato. Biocontrol Sci. Technol. 2021, 31, 912–923. [Google Scholar] [CrossRef]
- Benjamin, E.O.; Wesseler, J.H. A socioeconomic analysis of biocontrol in integrated pest management: A review of the effects of uncertainty, irreversibility and flexibility. NJAS-Wagening J. Life Sci. 2016, 77, 53–60. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.; Kannangara, S.D.; Promputtha, I. Fungi vs. fungi in biocontrol: An overview of fungal antagonists applied against fungal plant pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 604923. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef]
- Rokas, A.; Mead, M.E.; Steenwyk, J.L.; Raja, H.A.; Oberlies, N.H. Biosynthetic gene clusters and the evolution of fungal chemodiversity. Nat. Prod. Rep. 2020, 37, 868–878. [Google Scholar] [CrossRef] [PubMed]
- Al-Salihi, S.A.; Alberti, F. Naturally occurring terpenes: A promising class of organic molecules to address influenza pandemics. Nat. Prod. Bioprospect. 2021, 11, 405–419. [Google Scholar] [CrossRef]
- Keswani, C.; Singh, H.B.; Hermosa, R.; García-Estrada, C.; Caradus, J.; He, Y.W.; Mezaache-Aichour, S.; Glare, T.R.; Borriss, R.; Vinale, F.; et al. Antimicrobial secondary metabolites from agriculturally important fungi as next biocontrol agents. Appl. Microbiol. Biotechnol. 2019, 103, 9287–9303. [Google Scholar] [CrossRef] [PubMed]
- Claydown, K.L.; Emerson, O.H.; Sauthwell, R.J. The isolation of a toxic substance from the culture filtrate of Trichoderma. Phytopathology 1987, 36, 7. [Google Scholar]
- Hjeljord, L.; Tronsmo, A.; Harman, G.E.; Kubicek, C.P. Trichoderma and Gliocladium in biological control: An overview. In Trichoderma & Gliocladium: Enzymes, Biological Control and Commercial Applications; Harman, G.E., Kubice, C.P., Eds.; Taylor & Francis: Abingdon, UK, 1998; Volume 2, pp. 131–151. [Google Scholar]
- Tamizi, A.A.; Mat-Amin, N.; Weaver, J.A.; Olumakaiye, R.T.; Akbar, M.A.; Jin, S.; Bunawan, H.; Alberti, F. Genome Sequencing and Analysis of Trichoderma (Hypocreaceae) Isolates Exhibiting Antagonistic Activity against the Papaya Dieback Pathogen, Erwinia mallotivora. J. Fungi 2022, 8, 246. [Google Scholar] [CrossRef]
- Kumar, M.; Ashraf, S. Role of Trichoderma spp. as a biocontrol agent of fungal plant pathogens. In Probiotics and Plant Health; Springer: Singapore, 2017; pp. 497–506. [Google Scholar]
- Guo, K.; Sui, Y.; Li, Z.; Huang, Y.; Zhang, H. Trichoderma viride Tv-1511 colonizes Arabidopsis leaves and promotes Arabidopsis growth by modulating the MAP Kinase 6-mediated activation of plasma membrane H+-ATPase. J. Plant Growth Regul. 2020, 39, 1261–1276. [Google Scholar] [CrossRef]
- Shafique, S.; Shafique, S.; Javed, A.; Akhtar, N.; Bibi, S. Analysis of antagonistic potential of secondary metabolites and organic fractions of Trichoderma species against Alternaria Alternata. Biocontrol Sci. 2019, 24, 81–88. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Del-Val, E.K.; Larsen, J. Ecological functions of Trichoderma spp. and their secondary metabolites in the rhizosphere: Interactions with plants. FEMS Microbiol. Ecol. 2016, 92, fiw036. [Google Scholar] [CrossRef]
- Nordberg, H.; Cantor, M.; Dusheyko, S.; Hua, S.; Poliakov, A.; Shabalov, I.; Smirnova, T.; Grigoriev, I.V.; Dubchak, I. The genome portal of the Department of Energy Joint Genome Institute: 2014 updates. Nucleic Acids Res. 2014, 42, D26–D31. [Google Scholar] [CrossRef] [PubMed]
- Al-Salihi, S.A.; Dao, T.T.; Williams, K.; Bailey, A.M.; Foster, G.D. The biogenetic origin of the biologically active naematolin of Hypholoma species involves an unusual sesquiterpene synthase. Mol. Biotechnol. 2019, 61, 754–762. [Google Scholar] [CrossRef]
- Al-Salihi, S.A.; Bull, I.D.; Al-Salhi, R.; Gates, P.J.; Salih, K.S.; Bailey, A.M.; Foster, G.D. Further biochemical profiling of Hypholoma fasciculare metabolome reveals its chemogenetic diversity. Front. Bioeng. Biotechnol. 2021, 316, 567384. [Google Scholar] [CrossRef]
- Baroncelli, R.; Zapparata, A.; Piaggeschi, G.; Sarrocco, S.; Vannacci, G. Draft whole-genome sequence of Trichoderma gamsii T6085, a promising biocontrol agent of Fusarium head blight on wheat. Genome Announc. 2016, 4, e01747-15. [Google Scholar] [CrossRef] [PubMed]
- Bravo, D.; Cailleau, G.; Bindschedler, S.; Simon, A.; Job, D.; Verrecchia, E.; Junier, P. Isolation of oxalotrophic bacteria able to disperse on fungal mycelium. FEMS Microbiol. Lett. 2013, 348, 157–166. [Google Scholar] [CrossRef]
- Chadha, S.; Mehetre, S.T.; Bansal, R.; Kuo, A.; Aerts, A.; Grigoriev, I.V.; Druzhinina, I.S.; Mukherjee, P.K. Genome-wide analysis of cytochrome P450s of Trichoderma spp.: Annotation and evolutionary relationships. Fungal Biol. Biotechnol. 2018, 5, 12. [Google Scholar] [CrossRef]
- Druzhinina, I.S.; Chenthamara, K.; Zhang, J.; Atanasova, L.; Yang, D.; Miao, Y.; Rahimi, M.J.; Grujic, M.; Cai, F.; Pourmehdi, S.; et al. Massive lateral transfer of genes encoding plant cell wall-degrading enzymes to the mycoparasitic fungus Trichoderma from its plant-associated hosts. PLoS Genet. 2018, 14, e1007322. [Google Scholar] [CrossRef] [PubMed]
- Fanelli, F.; Liuzzi, V.C.; Logrieco, A.F.; Altomare, C. Genomic characterization of Trichoderma atrobrunneum (T. harzianum species complex) ITEM 908: Insight into the genetic endowment of a multi-target biocontrol strain. BMC Genom. 2018, 19, 662. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, K.R.P.; de Souza, A.Q.L.; dos Santos, L.A.; Nogueira, F.C.S.; Evaristo, J.A.M.; Carneiro, G.R.A.; da Silva, G.F.; da Cruz, J.C.; Sousa, T.F.; da Silva, S.R.S.; et al. Asperelines Produced by the Endophytic Fungus Trichoderma asperelloides from the Aquatic Plant Victoria amazonica. Rev. Bras. Farmacogn. 2021, 31, 667–675. [Google Scholar] [CrossRef]
- Gortikov, M.; Wang, Z.; Steindorff, A.S.; Grigoriev, I.V.; Druzhinina, I.S.; Townsend, J.P.; Yarden, O. Sequencing and Analysis of the Entire Genome of the Mycoparasitic Bioeffector Fungus Trichoderma asperelloides Strain T 203 (Hypocreales). Microbiol. Resour. Announc. 2022, 11, e00995-21. [Google Scholar] [CrossRef]
- Gruber, S.; Vaaje-Kolstad, G.; Matarese, F.; López-Mondéjar, R.; Kubicek, C.P.; Seidl-Seiboth, V. Analysis of subgroup C of fungal chitinases containing chitin-binding and LysM modules in the mycoparasite Trichoderma atroviride. Glycobiology 2011, 21, 122–133. [Google Scholar] [CrossRef]
- Koike, H.; Aerts, A.; LaButti, K.; Grigoriev, I.V.; Baker, S.E. Comparative Genomics Analysis of Trichoderma reesei Strains. Ind. Biotechnol. 2013, 9, 352–367. [Google Scholar] [CrossRef]
- Junier, P.; Cailleau, G.; Palmieri, I.; Vallotton, C.; Trautschold, O.C.; Junier, T.; Paul, C.; Bregnard, D.; Palmieri, F.; Estoppey, A.; et al. Democratization of fungal highway columns as a tool to investigate bacteria associated with soil fungi. FEMS Microbiol. Ecol. 2021, 97, fiab003. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Herrera-Estrella, A.; Seidl-Seiboth, V.; Martinez, D.A.; Druzhinina, I.S.; Thon, M.; Zeilinger, S.; Casas-Flores, S.; Horwitz, B.A.; Mukherjee, P.K.; et al. Comparative genome sequence analysis underscores mycoparasitism as the ancestral life style of Trichoderma. Genome Biol. 2011, 12, R40. [Google Scholar] [CrossRef] [PubMed]
- Kubicek, C.P.; Steindorff, A.S.; Chenthamara, K.; Manganiello, G.; Henrissat, B.; Zhang, J.; Cai, F.; Kopchinskiy, A.G.; Kubicek, E.M.; Kuo, A.; et al. Evolution and comparative genomics of the most common Trichoderma species. BMC Genom. 2019, 20, 485. [Google Scholar] [CrossRef] [PubMed]
- Li, W.C.; Huang, C.H.; Chen, C.L.; Chuang, Y.C.; Tung, S.Y.; Wang, T.F. Trichoderma reesei complete genome sequence, repeat-induced point mutation, and partitioning of CAZyme gene clusters. Biotechnol. Biofuels 2017, 10, 170. [Google Scholar] [CrossRef] [PubMed]
- Li, W.C.; Liu, H.C.; Lin, Y.J.; Tung, S.Y.; Wang, T.F. Third-generation sequencing-based mapping and visualization of single nucleotide polymorphism, meiotic recombination, illegitimate mutation and repeat-induced point mutation. NAR Genom. Bioinform. 2020, 2, lqaa056. [Google Scholar] [CrossRef]
- Li, W.C.; Lee, C.Y.; Lan, W.H.; Woo, T.T.; Liu, H.C.; Yeh, H.Y.; Chang, H.Y.; Chuang, Y.C.; Chen, C.Y.; Chuang, C.N.; et al. Trichoderma reesei Rad51 tolerates mismatches in hybrid meiosis with diverse genome sequences. Proc. Natl. Acad. Sci. USA 2021, 118, e2007192118. [Google Scholar] [CrossRef]
- Li, W.C.; Lin, T.C.; Chen, C.L.; Liu, H.C.; Lin, H.N.; Chao, J.L.; Hsieh, C.H.; Ni, H.F.; Chen, R.S.; Wang, T.F. Complete genome sequences and genome-wide characterization of Trichoderma biocontrol agents provide new insights into their evolution and variation in genome organization, sexual development, and fungal-plant interactions. Microbiol. Spectr. 2021, 9, e00663-21. [Google Scholar] [CrossRef]
- Lindo, L.; McCormick, S.P.; Cardoza, R.E.; Brown, D.W.; Kim, H.S.; Alexander, N.J.; Proctor, R.H.; Gutiérrez, S. Effect of deletion of a trichothecene toxin regulatory gene on the secondary metabolism transcriptome of the saprotrophic fungus Trichoderma arundinaceum. Fungal Genet. Biol. 2018, 119, 29–46. [Google Scholar] [CrossRef]
- Marik, T.; Urbán, P.; Tyagi, C.; Szekeres, A.; Leitgeb, B.; Vágvölgyi, M.; Manczinger, L.; Druzhinina, I.S.; Vágvölgyi, C.; Kredics, L. Diversity profile and dynamics of peptaibols produced by green mould Trichoderma species in interactions with their hosts Agaricus bisporus and Pleurotus ostreatus. Chem. Biodivers. 2017, 14, e1700033. [Google Scholar] [CrossRef]
- Martinez, D.; Berka, R.M.; Henrissat, B.; Saloheimo, M.; Arvas, M.; Baker, S.E.; Chapman, J.; Chertkov, O.; Coutinho, P.M.; Cullen, D.; et al. Genome sequencing and analysis of the biomass-degrading fungus Trichoderma reesei (syn. Hypocrea jecorina). Nat. Biotechnol. 2008, 26, 553–560. [Google Scholar] [CrossRef]
- Proctor, R.H.; McCormick, S.P.; Kim, H.S.; Cardoza, R.E.; Stanley, A.M.; Lindo, L.; Kelly, A.; Brown, D.W.; Lee, T.; Vaughan, M.M.; et al. Evolution of structural diversity of trichothecenes, a family of toxins produced by plant pathogenic and entomopathogenic fungi. PLoS Pathog. 2018, 14, e1006946. [Google Scholar] [CrossRef]
- Simon, A.; Bindschedler, S.; Job, D.; Wick, L.Y.; Filippidou, S.; Kooli, W.M.; Verrecchia, E.P.; Junier, P. Exploiting the fungal highway: Development of a novel tool for the in situ isolation of bacteria migrating along fungal mycelium. FEMS Microbiol. Ecol. 2015, 91, fiv116. [Google Scholar] [CrossRef]
- Studholme, D.J.; Harris, B.; Le Cocq, K.; Winsbury, R.; Perera, V.; Ryder, L.; Ward, J.L.; Beale, M.H.; Thornton, C.R.; Grant, M. Investigating the beneficial traits of Trichoderma hamatum GD12 for sustainable agriculture—Insights from genomics. Front. Plant Sci. 2013, 4, 258. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Pomraning, K.; Kopchinskiy, A.; Karimi Aghcheh, R.; Atanasova, L.; Chenthamara, K.; Baker, S.E.; Zhang, R.; Shen, Q.; Freitag, M.; et al. Genome sequence and annotation of Trichoderma parareesei, the ancestor of the cellulase producer Trichoderma reesei. Genome Announc. 2015, 3, e00885-15. [Google Scholar] [CrossRef] [PubMed]
- Chaverri, P.; Branco-Rocha, F.; Jaklitsch, W.; Gazis, R.; Degenkolb, T.; Samuels, G.J. Systematics of the Trichoderma harzianum species complex and the re-identification of commercial biocontrol strains. Mycologia 2015, 107, 558–590. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Zang, X.; Cheng, W.; Zhang, Z.; Zhou, J.; Chen, M.; Tang, Y. Harzianic acid from Trichoderma afroharzianum is a natural product inhibitor of acetohydroxyacid synthase. J. Am. Chem. Soc. 2021, 143, 9575–9584. [Google Scholar] [CrossRef] [PubMed]
- Pfordt, A.; Schiwek, S.; Karlovsky, P.; von Tiedemann, A. Trichoderma afroharzianum ear rot—A new disease on maize in Europe. Front. Agron. 2020, 11, 547758. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. AntiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Terlouw, B.R.; Blin, K.; Navarro-Muñoz, J.C.; E Avalon, N.; Chevrette, M.G.; Egbert, S.; Lee, S.; Meijer, D.; Recchia, M.J.J.; Reitz, Z.L.; et al. MIBiG 3.0: A community-driven effort to annotate experimentally validated biosynthetic gene clusters. Nucleic Acids Res. 2023, 51, D603–D610. [Google Scholar] [CrossRef]
- Rush, T.A.; Shrestha, H.K.; Gopalakrishnan Meena, M.; Spangler, M.K.; Ellis, J.C.; Labbé, J.L.; Abraham, P.E. Bioprospecting Trichoderma: A systematic roadmap to screen genomes and natural products for biocontrol applications. Front. Fungal Biol. 2021, 2, 716511. [Google Scholar] [CrossRef]
- Gilchrist, C.L.; Booth, T.J.; van Wersch, B.; van Grieken, L.; Medema, M.H.; Chooi, Y.H. Cblaster: A remote search tool for rapid identification and visualization of homologous gene clusters. Bioinform. Adv. 2021, 1, vbab016. [Google Scholar] [CrossRef] [PubMed]
- Steindorff, A.S.; Ramada, M.H.S.; Coelho, A.S.G.; Miller, R.N.G.; Pappas, G.J.; Ulhoa, C.J.; Noronha, E.F. Identification of mycoparasitism-related genes against the phytopathogen Sclerotinia sclerotiorum through transcriptome and expression profile analysis in Trichoderma harzianum. BMC Genom. 2014, 15, 204. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Liu, Q.; Gao, S.S.; Young, A.E.; Jacobsen, S.E.; Tang, Y. Genome mining and biosynthesis of a polyketide from a biofertilizer fungus that can facilitate reductive iron assimilation in plant. Proc. Natl. Acad. Sci. USA 2019, 116, 5499–5504. [Google Scholar] [CrossRef]
- Huang, K.; Tang, J.; Zou, Y.; Sun, X.; Lan, J.; Wang, W.; Xu, P.; Wu, X.; Ma, R.; Wang, Q.; et al. Whole genome sequence of Alternaria alternata, the causal agent of black spot of kiwifruit. Front. Microbiol. 2021, 12, 713462. [Google Scholar] [CrossRef]
- Iorio, M.; Davatgarbenam, S.; Serina, S.; Criscenzo, P.; Zdouc, M.M.; Simone, M.; Maffioli, S.I.; Ebright, R.H.; Donadio, S.; Sosio, M. Blocks in the pseudouridimycin pathway unlock hidden metabolites in the Streptomyces producer strain. Sci. Rep. 2021, 11, 5827. [Google Scholar] [CrossRef]
- Pohle, S.; Appelt, C.; Roux, M.; Fiedler, H.P.; Süssmuth, R.D. Biosynthetic gene cluster of the non-ribosomally synthesized cyclodepsipeptide skyllamycin: Deciphering unprecedented ways of unusual hydroxylation reactions. J. Am. Chem. Soc. 2011, 33, 6194–6205. [Google Scholar] [CrossRef]
- Sun, C.; Yang, Z.; Zhang, C.; Liu, Z.; He, J.; Liu, Q.; Zhang, T.; Ju, J.; Ma, J. Genome mining of Streptomyces atratus SCSIO ZH16: Discovery of atratumycin and identification of its biosynthetic gene cluster. Org. Lett. 2019, 21, 1453–1457. [Google Scholar] [CrossRef]
- Shi, J.; Liu, C.L.; Zhang, B.; Guo, W.J.; Zhu, J.; Chang, C.Y.; Zhao, E.J.; Jiao, R.H.; Tan, R.X.; Ge, H.M. Genome mining and biosynthesis of kitacinnamycins as a STING activator. Chem. Sci. 2019, 10, 4839–4846. [Google Scholar] [CrossRef]
- Zhang, C.; Seyedsayamdost, M.R. Discovery of a cryptic depsipeptide from streptomyces ghanaensis via MALDI-MS-guided high-throughput elicitor screening. Angew. Chem. Int. Ed. 2020, 59, 23005–23009. [Google Scholar] [CrossRef]
- Stachelhaus, T.; Mootz, H.D.; Marahiel, M.A. The specificity-conferring code of adenylation domains in nonribosomal peptide synthetases. Chem. Biol. 1999, 6, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Conti, E.; Stachelhaus, T.; Marahiel, M.A.; Brick, P. Structural basis for the activation of phenylalanine in the non-ribosomal biosynthesis of gramicidin S. EMBO J. 1997, 16, 4174–4183. [Google Scholar] [CrossRef]
- Marner, M.; Patras, M.A.; Kurz, M.; Zubeil, F.; Förster, F.; Schuler, S.; Bauer, A.; Hammann, P.; Vilcinskas, A.; Schäberle, T.F.; et al. Molecular Networking-Guided Discovery and Characterization of Stechlisins, a Group of Cyclic Lipopeptides from a Pseudomonas sp. J. Nat. Prod. 2020, 83, 2607–2617. [Google Scholar] [CrossRef]
- Steele, J.A.; Uchytil, T.F.; Durbin, R.D.; Bhatnagar, P.; Rich, D.H. Chloroplast coupling factor 1: A species-specific receptor for tentoxin. Proc. Natl. Acad. Sci. USA 1976, 73, 2245–2248. [Google Scholar] [CrossRef]
- Li, Y.H.; Han, W.J.; Gui, X.W.; Wei, T.; Tang, S.Y.; Jin, J.M. Putative nonribosomal peptide synthetase and cytochrome P450 genes responsible for tentoxin biosynthesis in Alternaria alternata ZJ33. Toxins 2016, 8, 234. [Google Scholar] [CrossRef]
- Caesar, L.K.; Robey, M.T.; Swyers, M.; Islam, M.N.; Ye, R.; Vagadia, P.P.; Schiltz, G.E.; Thomas, P.M.; Wu, C.C.; Kelleher, N.L.; et al. Heterologous expression of the unusual terreazepine biosynthetic gene cluster reveals a promising approach for identifying new chemical scaffolds. mBio 2020, 11, e01691-20. [Google Scholar] [CrossRef]
- Wight, W.D.; Labuda, R.; Walton, J.D. Conservation of the genes for HC-toxin biosynthesis in Alternaria jesenskae. BMC Microbiol. 2013, 13, 165. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Kang, Q.; Lu, Y.; Bai, L.; Wang, C. Unveiling the biosynthetic puzzle of destruxins in Metarhizium species. Proc. Natl. Acad. Sci. USA 2012, 109, 1287–1292. [Google Scholar] [CrossRef]
- Pedras, M.S.C.; Zaharia, L.I.; Ward, D.E. The destruxins: Synthesis, biosynthesis, biotransformation, and biological activity. Phytochemistry 2002, 59, 579–596. [Google Scholar] [CrossRef]
- Ruiz-Sanchez, E.; Lange, A.B.; Orchard, I. Effects of the mycotoxin destruxin A on Locusta migratoria visceral muscles. Toxicon 2010, 56, 1043–1051. [Google Scholar] [CrossRef] [PubMed]
- Sarges, R.; Witkop, B. Gramicidin AV The structure of valine-and isoleucine-gramicidin A. J. Am. Chem. Soc. 1965, 87, 2011–2020. [Google Scholar] [CrossRef]
- Takada, Y.; Itoh, H.; Paudel, A.; Panthee, S.; Hamamoto, H.; Sekimizu, K.; Inoue, M. Discovery of gramicidin A analogues with altered activities by multidimensional screening of a one-bead-one-compound library. Nat. Commun. 2020, 11, 4935. [Google Scholar] [CrossRef]
- Hamano, K.; Kinoshita, M.; Tanzawa, K.; Yoda, K.; Ohki, Y.; Nakamura, T.; Kinoshita, T. Leualacin, a novel calcium blocker from Hapsidospora irregularis II. Structure determination. J. Antibiot. 1992, 45, 906–913. [Google Scholar] [CrossRef] [PubMed]
- Raymond, K.N.; Dertz, E.A.; Kim, S.S. Enterobactin: An archetype for microbial iron transport. Proc. Natl. Acad. Sci. USA 2003, 100, 3584–3588. [Google Scholar] [CrossRef] [PubMed]
- Pollack, J.R.; Neilands, J.B. Enterobactin, an iron transport compound from Salmonella typhimurium. Biochem. Biophys. Res. Commun. 1970, 38, 989–992. [Google Scholar] [CrossRef]
- Royles, B.J. Naturally occurring tetramic acids: Structure, isolation, and synthesis. Chem. Rev. 1995, 95, 1981–2001. [Google Scholar] [CrossRef]
- Sims, J.W.; Fillmore, J.P.; Warner, D.D.; Schmidt, E.W. Equisetin biosynthesis in Fusarium heterosporum. Chem. Commun. 2005, 2, 186–188. [Google Scholar] [CrossRef]
- Tobert, J.A. Lovastatin and beyond: The history of the HMG-CoA reductase inhibitors. Nat. Rev. Drug Discov. 2003, 2, 517–526. [Google Scholar] [CrossRef]
- Campbell, C.D.; Vederas, J.C. Biosynthesis of lovastatin and related metabolites formed by fungal iterative PKS enzymes. Biopolymers 2010, 93, 755–763. [Google Scholar] [CrossRef]
- Koumoutsi, A.; Chen, X.H.; Henne, A.; Liesegang, H.; Hitzeroth, G.; Franke, P.; Vater, J.; Borriss, R. Structural and functional characterization of gene clusters directing nonribosomal synthesis of bioactive cyclic lipopeptides in Bacillus amyloliquefaciens strain FZB42. J. Bacteriol. 2004, 186, 1084–1096. [Google Scholar] [CrossRef]
- Díaz-Sánchez, V.; Avalos, J.; Limón, M.C. Identification and regulation of fusA, the polyketide synthase gene responsible for fusarin production in Fusarium fujikuroi. Appl. Environ. Microbiol. 2012, 78, 7258–7266. [Google Scholar] [CrossRef]
- Chiang, Y.M.; Szewczyk, E.; Davidson, A.D.; Keller, N.; Oakley, B.R.; Wang, C.C. A gene cluster containing two fungal polyketide synthases encodes the biosynthetic pathway for a polyketide, asperfuranone, in Aspergillus nidulans. J. Am. Chem. Soc. 2009, 131, 2965–2970. [Google Scholar] [CrossRef]
- Li, Y.P.; Xu, Y.; Huang, Z.B. Isolation and characterization of the citrinin biosynthetic gene cluster from Monascus aurantiacus. Biotechnol. Lett. 2012, 34, 131–136. [Google Scholar] [CrossRef]
- Wang, C.M.; Hopson, R.; Lin, X.; Cane, D.E. Biosynthesis of the Sesquiterpene Botrydial in Botrytis cinerea. Mechanism and Stereochemistry of the Enzymatic Formation of Presilphiperfolan-8β-ol. J. Am. Chem. Soc. 2009, 131, 8360–8361. [Google Scholar] [CrossRef]
- Xu, H.; Dickschat, J.S. Germacrene A–A central intermediate in sesquiterpene biosynthesis. Chem. Eur. J. 2020, 26, 17318–17341. [Google Scholar] [CrossRef]
- Aaron, J.A.; Lin, X.; Cane, D.E.; Christianson, D.W. Structure of epi-isozizaene synthase from Streptomyces coelicolor A3 (2), a platform for new terpenoid cyclization templates. Biochemistry 2010, 49, 1787–1797. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Most Similar BGC on antiSMASH | BGC Type | Trichoderma harzianum CBS 226.95 v1.0 (Triha1) | Trichoderma harzianum T22 v1.0 (TriharT22) | Trichoderma harzianum TR274 v1.0 (Trihar1) | Trichoderma harzianum M10 v1.0 (TriharM10) |
|---|---|---|---|---|---|
| Metachelin A-C | NRPS | 62% | 75% | 62% | 62% |
| Ochratoxin | NRPS-PKS | X | X | X | 100% |
| Dichlorodiapothin | PKS | 50% | 50% | 50% | 33% |
| Trichoxide | PKS | 75% | 83% | 75% | 83% |
| Harzianopyridone | NRPS-PKS | 60% | 60% | 60% | 70% |
| Clavaric acid | Terpene | 100% | X | 100% | 100% |
| Harziphilone | PKS | 80% | 80% | 80% | 80% |
| Squalestatin | Terpene | 40% | 40% | X | 40% |
| Depudecin | PKS | 33% | 33% | 33% | 33% |
| Trichobrasilenol | Terpene | 60% | 60% | 60% | X |
| Tricholignan | PKS | 100% | 100% | 100% | 100% |
| Choline | NRPS | 100% | 100% | 100% | 100% |
| Peramine | NRPS | X | 100% | X | 100% |
| Lucilactaene | PKS | X | X | X | 38% |
| BGC of Interest | No. of Organisms Searched | No. of Genomic Scaffold Searched | No. of Scaffolds Identified with the BGC | No. of Trichoderma Species with Similar BGC |
|---|---|---|---|---|
| Ochratoxin | 416 | 566 | 573 | 18 |
| Clavaric acid | 513 | 568 | 559 | None |
| Harziphilon | 631 | 2971 | 735 | 9 |
| Tricholignan | 93 | 390 | 96 | 7 |
| Choline | 1088 | 1134 | 1133 | None |
| Peramine | 346 | 628 | 512 | 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Salihi, S.A.A.; Alberti, F. Genomic Based Analysis of the Biocontrol Species Trichoderma harzianum: A Model Resource of Structurally Diverse Pharmaceuticals and Biopesticides. J. Fungi 2023, 9, 895. https://0-doi-org.brum.beds.ac.uk/10.3390/jof9090895
Al-Salihi SAA, Alberti F. Genomic Based Analysis of the Biocontrol Species Trichoderma harzianum: A Model Resource of Structurally Diverse Pharmaceuticals and Biopesticides. Journal of Fungi. 2023; 9(9):895. https://0-doi-org.brum.beds.ac.uk/10.3390/jof9090895
Chicago/Turabian StyleAl-Salihi, Suhad A. A., and Fabrizio Alberti. 2023. "Genomic Based Analysis of the Biocontrol Species Trichoderma harzianum: A Model Resource of Structurally Diverse Pharmaceuticals and Biopesticides" Journal of Fungi 9, no. 9: 895. https://0-doi-org.brum.beds.ac.uk/10.3390/jof9090895