
Gene Expression and Characterization of Iturin A Lipopeptide Biosurfactant from Bacillus aryabhattai for Enhanced Oil Recovery

 , , , ,
, , , ,
Abstract
:
1. Introduction
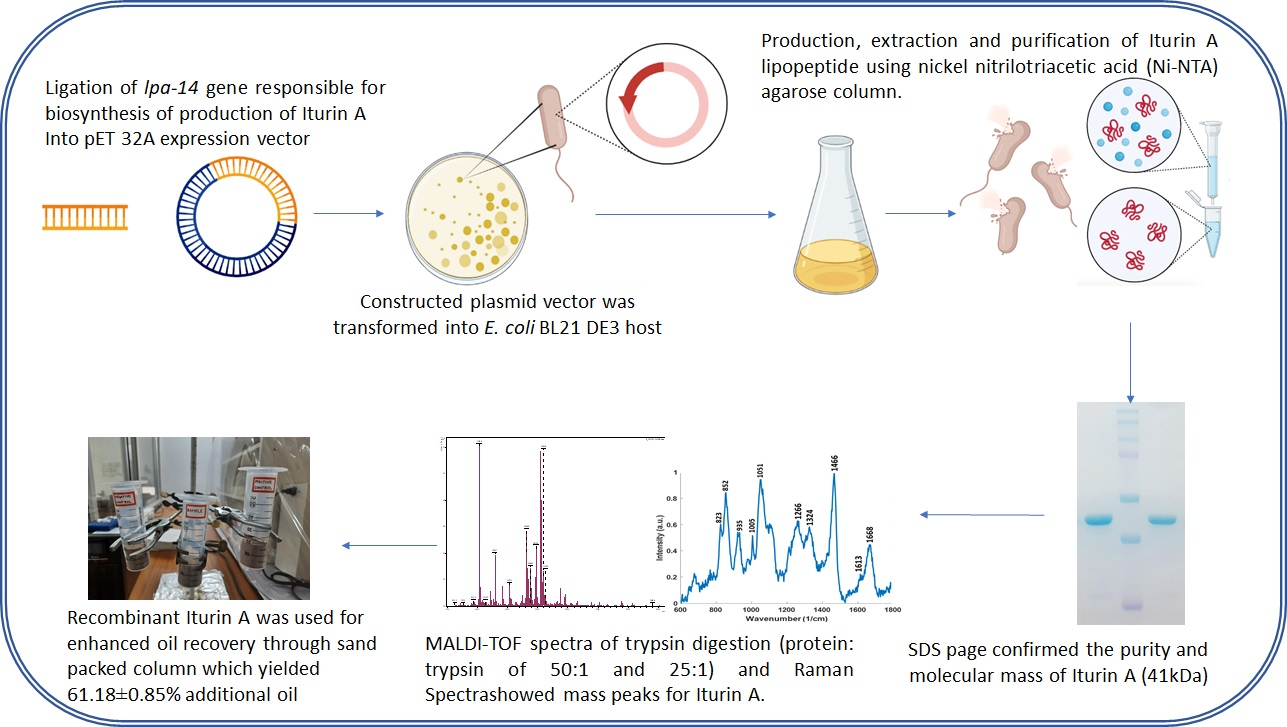
2. Materials and Methods
2.1. Chemicals, Vectors, and Bacterial Strains
2.2. Genome Sequence Identification
Gene Synthesis, Amplification, and Cloning
2.3. Expression of the Iturin A Lipopeptide
2.4. Purification of the Recombinant Iturin A Lipopeptide
2.5. Evaluation of Biosurfactant Activity
2.6. Analytical Characterization
2.6.1. Mass Spectrometry Analysis
2.6.2. Raman Spectroscopy Analysis
2.7. Sand-Packed Column Assay for Enhanced Oil Recovery
3. Results and Discussion
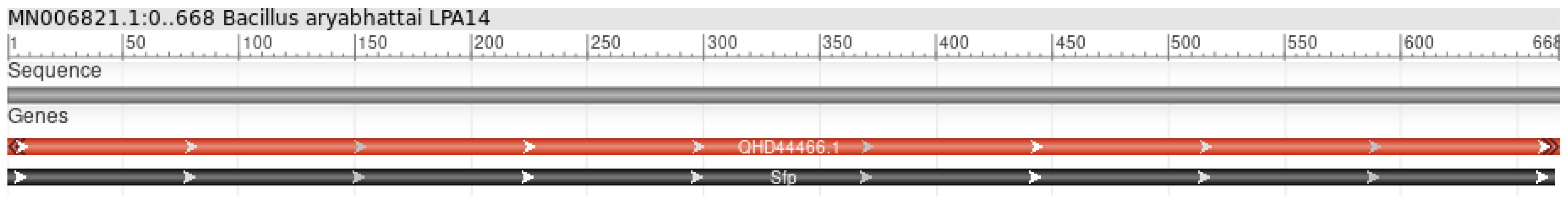
3.1. Genome Sequence Identification
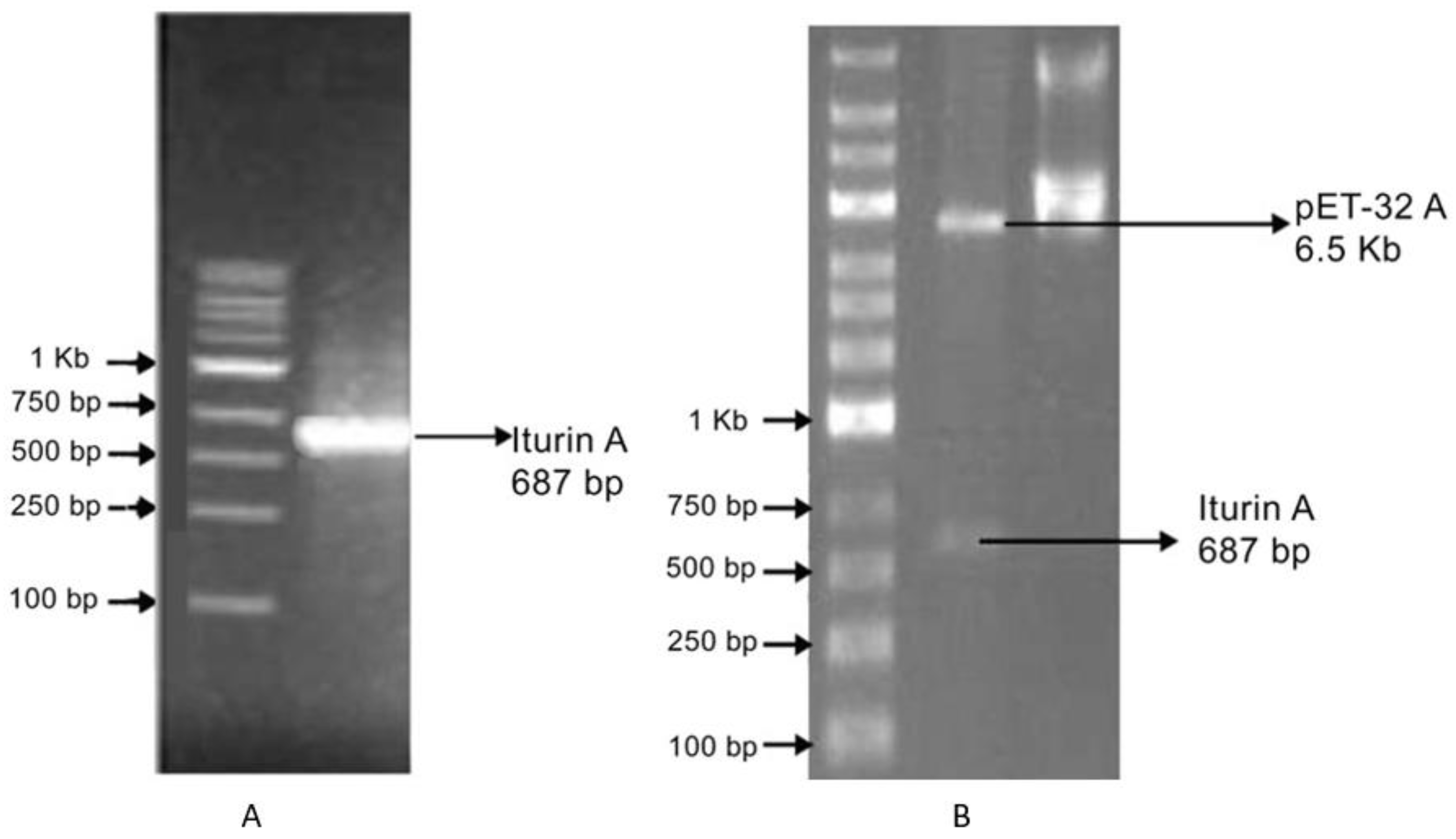
3.2. Cloning into the pET-32A Vector
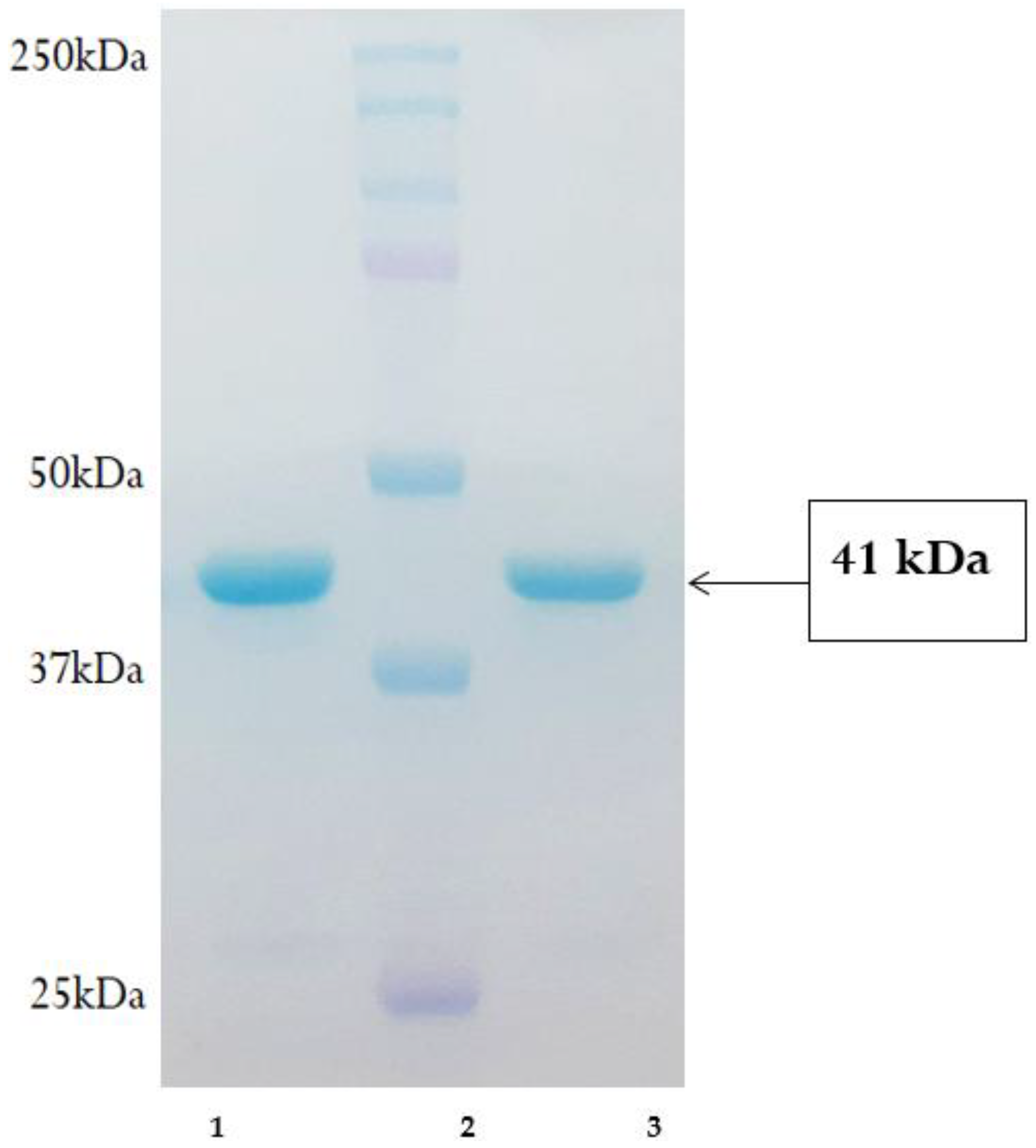
3.3. Expression and Purification of His–pET-32A-Iturin A Fusion Protein
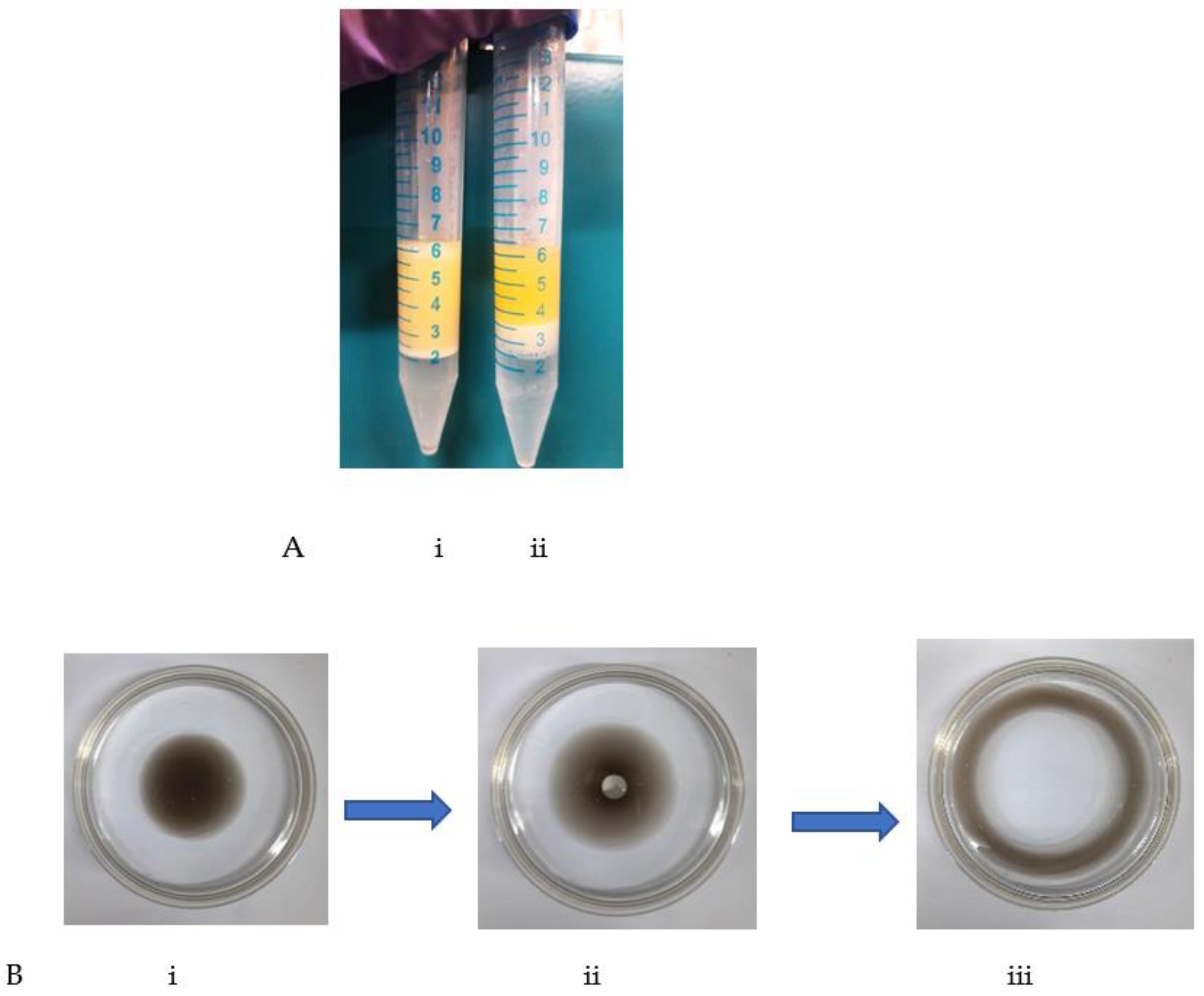
3.4. Evaluation of Biosurfactant Activity
3.5. Analytical Characterization
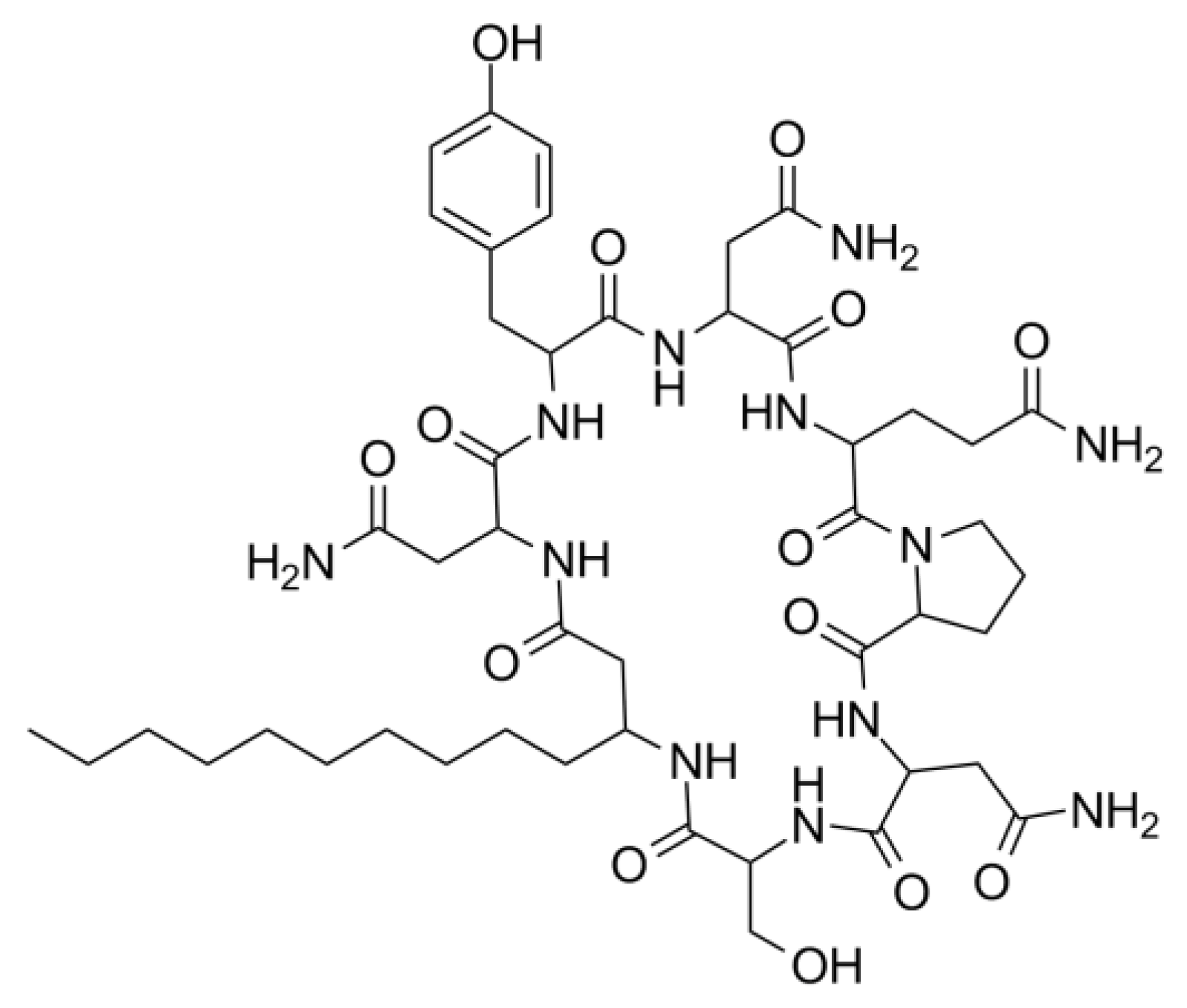
3.5.1. Mass Spectrometry Analysis
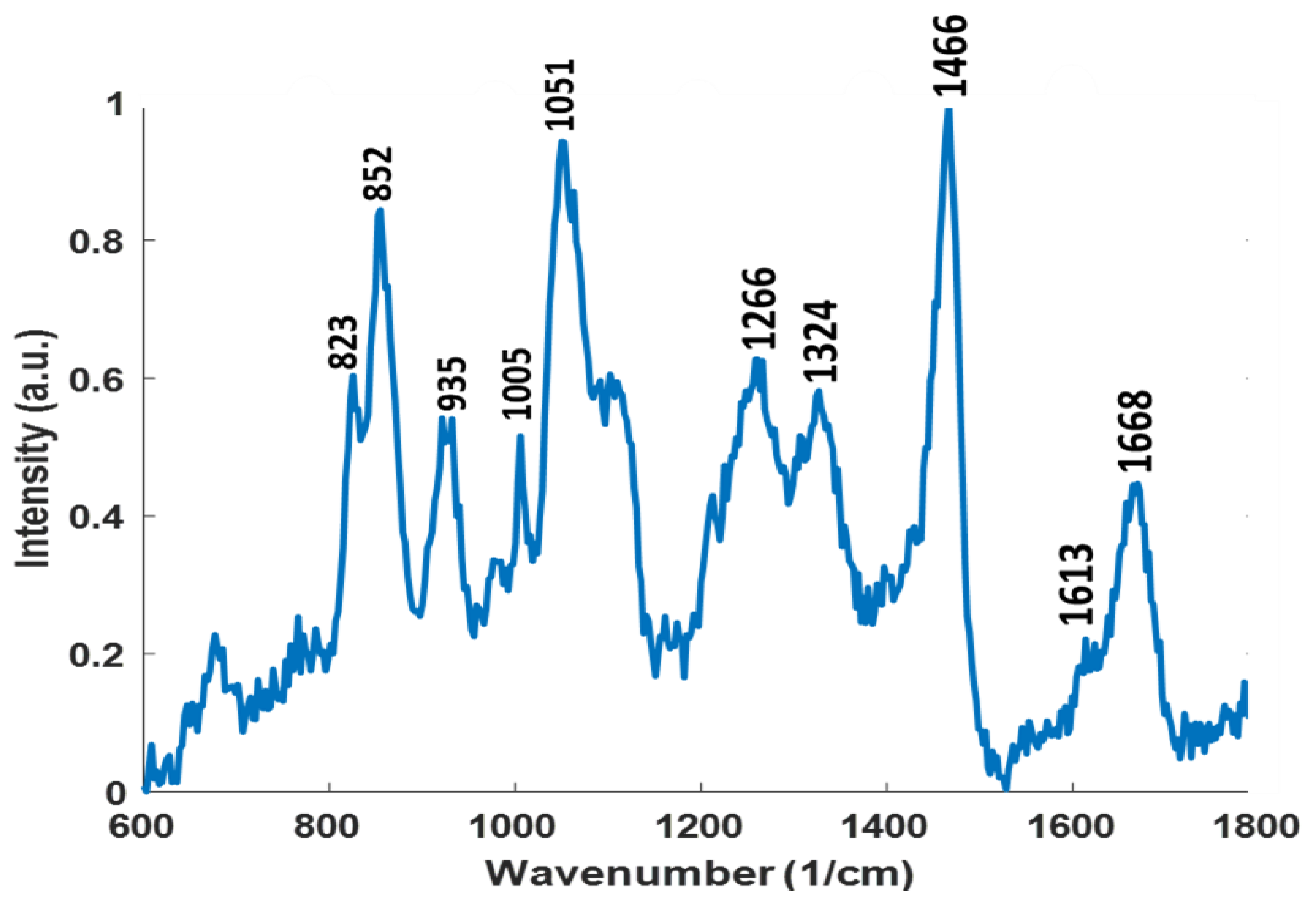
3.5.2. Raman Spectroscopy Analysis
3.6. Application of a Biosurfactant for Enhanced Oil Recovery
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sekhon, K.; Khanna, S.; Cameotra, S. Enhanced biosurfactant production through cloning of three genes and role of esterase in biosurfactant release. Microb. Cell Fact. 2011, 10, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapellos, G.E. Microbial Strategies for Oil Biodegradation. In Modeling of Microscale Transport in Biological Processes; Elsevier: Amsterdam, The Netherlands, 2017; pp. 19–39. [Google Scholar]
- Costa, J.A.V.; Treichel, H.; Santos, L.O.; Martins, V.G. Solid-State Fermentation for the Production of Biosurfactants and Their Applications. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2018; pp. 357–372. [Google Scholar]
- Meena, K.R.; Kanwar, S.S. Lipopeptides as the Antifungal and Antibacterial Agents: Applications in Food Safety and Therapeutics. Biomed Res. Int. 2015, 2015, 473050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zohora, U.S.; Rahman, M.S.; Ano, T. Biofilm formation and lipopeptide antibiotic iturin A production in different peptone media. J. Environ. Sci. 2009, 21, S24–S27. [Google Scholar] [CrossRef]
- Tsuge, K.; Akiyama, T.; Shoda, M. Cloning, Sequencing, and Characterization of the Iturin A Operon. J. Bacteriol. 2001, 183, 6265–6273. [Google Scholar] [CrossRef] [Green Version]
- Yaraguppi, D.A.; Bagewadi, Z.K.; Muddapur, U.M.; Mulla, S.I. Response surface methodology-based optimization of biosurfactant production from isolated Bacillus aryabhattai strain ZDY2. J. Pet. Explor. Prod. Technol. 2020, 10, 2483–2498. [Google Scholar] [CrossRef] [Green Version]
- Jimoh, A.A.; Lin, J. Biosurfactant: A new frontier for greener technology and environmental sustainability. Ecotoxicol. Environ. Saf. 2019, 184, 109607. [Google Scholar] [CrossRef]
- Bachmann, R.T.; Johnson, A.C.; Edyvean, R.G.J. Biotechnology in the petroleum industry: An overview. Int. Biodeterior. Biodegrad. 2014, 86, 225–237. [Google Scholar] [CrossRef]
- Sarubbo, L.A.; Silva, M.d.G.C.; Durval, I.J.B.; Bezerra, K.G.O.; Ribeiro, B.G.; Silva, I.A.; Twigg, M.S.; Banat, I.M. Biosurfactants: Production, properties, applications, trends, and general perspectives. Biochem. Eng. J. 2022, 181, 108377. [Google Scholar] [CrossRef]
- Roongsawang, N.; Thaniyavarn, J.; Thaniyavarn, S.; Kameyama, T.; Haruki, M.; Imanaka, T.; Morikawa, M.; Kanaya, S. Isolation and characterization of a halotolerant Bacillus subtilis BBK-1 which produces three kinds of lipopeptides: Bacillomycin L, plipastatin, and surfactin. Extremophiles 2002, 6, 499–506. [Google Scholar] [CrossRef]
- Luo, C.; Chen, Y.; Liu, X.; Wang, X.; Wang, X.; Li, X.; Zhao, Y.; Wei, L. Engineered biosynthesis of cyclic lipopeptide locillomycins in surrogate host Bacillus velezensis FZB42 and derivative strains enhance antibacterial activity. Appl. Microbiol. Biotechnol. 2019, 103, 4467–4481. [Google Scholar] [CrossRef]
- Ebadi, A.; Olamaee, M.; Khoshkholgh Sima, N.A.; Ghorbani Nasrabadi, R.; Hashemi, M. Isolation and Characterization of Biosurfactant Producing and Crude Oil Degrading Bacteria from Oil Contaminated Soils. Iran. J. Sci. Technol. Trans. A Sci. 2018, 42, 1149–1156. [Google Scholar] [CrossRef]
- Yaraguppi, D.A.; Bagewadi, Z.K.; Deshpande, S.H.; Chandramohan, V. In Silico Study on the Inhibition of UDP-N-Acetylglucosamine 1-Carboxy Vinyl Transferase from Salmonella typhimurium by the Lipopeptide Produced from Bacillus aryabhattai. Int. J. Pept. Res. Ther. 2022, 28, 80. [Google Scholar] [CrossRef]
- Duarte, C.; Gudiña, E.J.; Lima, C.F.; Rodrigues, L.R. Effects of biosurfactants on the viability and proliferation of human breast cancer cells. AMB Express 2014, 4, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Xu, P.; Zeng, G.; Yang, C.; Huang, D.; Zhang, J. Bioremediation of soils contaminated with polycyclic aromatic hydrocarbons, petroleum, pesticides, chlorophenols and heavy metals by composting: Applications, microbes and future research needs. Biotechnol. Adv. 2015, 33, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhu, X.; Lu, Y.; Zhao, H.; Lu, F.; Lu, Z. Improving Iturin a Production of Bacillus amyloliquefaciens by Genome Shuffling and Its Inhibition against Saccharomyces cerevisiae in Orange Juice. Front. Microbiol. 2018, 9, 2683. [Google Scholar] [CrossRef]
- Yaish, M.W. Draft Genome Sequence of the Endophytic Bacillus aryabhattai Strain SQU-R12, Identified from Phoenix dactylifera L. Roots. Genome Announc. 2017, 5, e00718-17. [Google Scholar] [CrossRef] [Green Version]
- Madeira, F.; Park, Y.m.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Ashish; Debnath, M. Application of biosurfactant produced by an adaptive strain of C.tropicalis MTCC230 in microbial enhanced oil recovery (MEOR) and removal of motor oil from contaminated sand and water. J. Pet. Sci. Eng. 2018, 170, 40–48. [Google Scholar] [CrossRef]
- Astuti, D.I.; Purwasena, I.A.; Putri, R.E.; Amaniyah, M.; Sugai, Y. Screening and characterization of biosurfactant produced by Pseudoxanthomonas sp. G3 and its applicability for enhanced oil recovery. J. Pet. Explor. Prod. Technol. 2019, 9, 2279–2289. [Google Scholar] [CrossRef] [Green Version]
- Das, A.J.; Kumar, R. Utilization of agro-industrial waste for biosurfactant production under submerged fermentation and its application in oil recovery from sand matrix. Bioresour. Technol. 2018, 260, 233–240. [Google Scholar] [CrossRef]
- Yaraguppi, D.A.; Deshpande, S.H.; Bagewadi, Z.K.; Kumar, S.; Muddapur, U.M. Genome Analysis of Bacillus aryabhattai to Identify Biosynthetic Gene Clusters and In Silico Methods to Elucidate its Antimicrobial Nature. Int. J. Pept. Res. Ther. 2021, 27, 1331–1342. [Google Scholar] [CrossRef]
- Flissi, A.; Ricart, E.; Campart, C.; Chevalier, M.; Dufresne, Y.; Michalik, J.; Jacques, P.; Flahaut, C.; Lisacek, F.; Leclère, V.; et al. Norine: Update of the nonribosomal peptide resource. Nucleic Acids Res. 2020, 48, D465–D469. [Google Scholar] [CrossRef]
- Gaur, V.K.; Sharma, P.; Gupta, S.; Varjani, S.; Srivastava, J.K.; Wong, J.W.C.; Ngo, H.H. Opportunities and challenges in omics approaches for biosurfactant production and feasibility of site remediation: Strategies and advancements. Environ. Technol. Innov. 2022, 25, 102132. [Google Scholar] [CrossRef]
- Xu, Y.; Cai, D.; Zhang, H.; Gao, L.; Yang, Y.; Gao, J.; Li, Y.; Yang, C.; Ji, Z.; Yu, J.; et al. Enhanced production of iturin A in Bacillus amyloliquefaciens by genetic engineering and medium optimization. Process Biochem. 2020, 90, 50–57. [Google Scholar] [CrossRef]
- Zhang, Z.; Ding, Z.T.; Zhong, J.; Zhou, J.Y.; Shu, D.; Luo, D.; Yang, J.; Tan, H. Improvement of iturin A production in Bacillus subtilis ZK0 by overexpression of the comA and sigA genes. Lett. Appl. Microbiol. 2017, 64, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Dang, Y.; Zhao, F.; Liu, X.; Fan, X.; Huang, R.; Gao, W.; Wang, S.; Yang, C. Enhanced production of antifungal lipopeptide iturin A by Bacillus amyloliquefaciens LL3 through metabolic engineering and culture conditions optimization. Microb. Cell Fact. 2019, 18, 68. [Google Scholar] [CrossRef] [PubMed]
- Jimoh, A.A.; Senbadejo, T.Y.; Adeleke, R.; Lin, J. Development and Genetic Engineering of Hyper-Producing Microbial Strains for Improved Synthesis of Biosurfactants. Mol. Biotechnol. 2021, 63, 267–288. [Google Scholar] [CrossRef]
- Bagewadi, Z.K.; Mulla, S.I.; Ninnekar, H.Z. Purification, characterization, gene cloning and expression of GH-10 xylanase (Penicillium citrinum isolate HZN13). 3 Biotech 2016, 6, 169. [Google Scholar] [CrossRef] [Green Version]
- Meena, K.R.; Dhiman, R.; Singh, K.; Kumar, S.; Sharma, A.; Kanwar, S.S.; Mondal, R.; Das, S.; Franco, O.L.; Mandal, A.K. Purification and identification of a surfactin biosurfactant and engine oil degradation by Bacillus velezensis KLP2016. Microb. Cell Fact. 2021, 20, 26. [Google Scholar] [CrossRef]
- Asgher, M.; Arshad, S.; Qamar, S.A.; Khalid, N. Improved biosurfactant production from Aspergillus niger through chemical mutagenesis: Characterization and RSM optimization. SN Appl. Sci. 2020, 2, 966. [Google Scholar] [CrossRef] [Green Version]
- Pele, M.A.; Ribeaux, D.R.; Vieira, E.R.; Souza, A.F.; Luna, M.A.C.; Rodríguez, D.M.; Andrade, R.F.S.; Alviano, D.S.; Alviano, C.S.; Barreto-Bergter, E.; et al. Conversion of renewable substrates for biosurfactant production by Rhizopus arrhizus UCP 1607 and enhancing the removal of diesel oil from marine soil. Electron. J. Biotechnol. 2019, 38, 40–48. [Google Scholar] [CrossRef]
- Simpson, R.J. Fragmentation of Protein Using Trypsin. Cold Spring Harb. Protoc. 2006, 2006, pdb.prot4550. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Li, Q.; Fu, G.; Yuan, G.; Miao, J.; Lin, W. Identification of Antifungal Substance (Iturin A2) Produced by Bacillus subtilis B47 and its Effect on Southern Corn Leaf Blight. J. Integr. Agric. 2012, 11, 90–99. [Google Scholar] [CrossRef]
- Labiadh, M.; Dhaouadi, S.; Chollet, M.; Chataigne, G.; Tricot, C.; Jacques, P.; Flahaut, S.; Kallel, S. Antifungal lipopeptides from Bacillus strains isolated from rhizosphere of Citrus trees. Rhizosphere 2021, 19, 100399. [Google Scholar] [CrossRef]
- Stincone, P.; Veras, F.F.; Pereira, J.Q.; Mayer, F.Q.; Varela, A.P.M.; Brandelli, A. Diversity of cyclic antimicrobial lipopeptides from Bacillus P34 revealed by functional annotation and comparative genome analysis. Microbiol. Res. 2020, 238, 126515. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Chávez, J.L.; Juárez-Campusano, Y.S.; Delgado, G.; Pacheco Aguilar, J.R. Identification of lipopeptides from Bacillus strain Q11 with ability to inhibit the germination of Penicillium expansum, the etiological agent of postharvest blue mold disease. Postharvest Biol. Technol. 2019, 155, 72–79. [Google Scholar] [CrossRef]
- San Keskin, N.O.; Han, D.; Devrim Ozkan, A.; Angun, P.; Onarman Umu, O.C.; Tekinay, T. Production and structural characterization of biosurfactant produced by newly isolated staphylococcus xylosus STF1 from petroleum contaminated soil. J. Pet. Sci. Eng. 2015, 133, 689–694. [Google Scholar] [CrossRef]
- Shi, L.; He, Y.; Lu, J.; Hou, G.; Liang, D. Anti-agglomeration evaluation and Raman spectroscopic analysis on mixed biosurfactants for preventing CH4 hydrate blockage in n-octane + water systems. Energy 2021, 229, 120755. [Google Scholar] [CrossRef]
- Zhu, F.; Isaacs, N.W.; Hecht, L.; Barron, L.D. Raman Optical Activity: A Tool for Protein Structure Analysis. Structure 2005, 13, 1409–1419. [Google Scholar] [CrossRef]
- Maiti, N.C.; Apetri, M.M.; Zagorski, M.G.; Carey, P.R.; Anderson, V.E. Raman Spectroscopic Characterization of Secondary Structure in Natively Unfolded Proteins: α-Synuclein. J. Am. Chem. Soc. 2004, 126, 2399–2408. [Google Scholar] [CrossRef]
- Elsevier. Biobased Surfactants; Elsevier: Amsterdam, The Netherlands, 2019; ISBN 9780128127056. [Google Scholar]
- Hashizume, H.; Nishimura, Y. Cyclic Lipopeptide Antibiotics. Stud. Nat. Prod. Chem. 2008, 35, 693–751. [Google Scholar]
- Pandey, R.; Krishnamurthy, B.; Singh, H.P.; Batish, D.R. Evaluation of a glycolipopepetide biosurfactant from Aeromonas hydrophila RP1 for bioremediation and enhanced oil recovery. J. Clean. Prod. 2022, 345, 131098. [Google Scholar] [CrossRef]
- Nasiri, M.A.; Biria, D. Extraction of the indigenous crude oil dissolved biosurfactants and their potential in enhanced oil recovery. Colloids Surf. A Physicochem. Eng. Asp. 2020, 603, 125216. [Google Scholar] [CrossRef]
- Sharma, J.; Kapley, A.; Sundar, D.; Srivastava, P. Characterization of a potent biosurfactant produced from Franconibacter sp. IITDAS19 and its application in enhanced oil recovery. Colloids Surf. B Biointerfaces 2022, 214, 112453. [Google Scholar] [CrossRef] [PubMed]
- Suthar, H.; Hingurao, K.; Desai, A.; Nerurkar, A. Evaluation of bioemulsifier mediated Microbial Enhanced Oil Recovery using sand pack column. J. Microbiol. Methods 2008, 75, 225–230. [Google Scholar] [CrossRef]
- Arora, P.; Kshirsagar, P.R.; Rana, D.P.; Dhakephalkar, P.K. Hyperthermophilic Clostridium sp. N-4 produced a glycoprotein biosurfactant that enhanced recovery of residual oil at 96 °C in lab studies. Colloids Surf. B Biointerfaces 2019, 182, 110372. [Google Scholar] [CrossRef]
- Wang, X.-T.; Liu, B.; Li, X.-Z.; Lin, W.; Li, D.-A.; Dong, H.; Wang, L. Biosurfactants produced by novel facultative-halophilic Bacillus sp. XT-2 with biodegradation of long chain n-alkane and the application for enhancing waxy oil recovery. Energy 2022, 240, 122802. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predicted Masses, [M + H]+ | Observed Masses in 50:1 Trypsin Digestion, [M + H]+ | Observed Masses in 25:1 Trypsin Digestion, [M + H]+ |
|---|---|---|
| 3904.9 | 3904.2 | 3904.9 |
| 2135.9 | - | 2135.9 |
| 2095.9 | 2096.0 | 2095.9 |
| 1970.9 | 1970.9 | 1970.9 |
| 1886.0 | 1886.0 | 1886.0 |
| 1819.8 | 1819.8 | 1819.8 |
| 1532.7 | 1532.7 | 1532.7 |
| 1290.7 | 1290.8 | 1290.7 |
| 1133.4 | 1133.6 | 1133.5 |
| 1073.6 | 1073.6 | 1073.5 |
| 1031.6 | 1031.6 | 1031.6 |
| 937.4 | 937.4 | 937.4 |
| 767.3 | 767.4 | 767.4 |
| 627.3 | - | - |
| 623.3 | 623.3 | 623.3 |
| 594.3 | - | - |
| Parameters | Sample | Positive Control | Negative Control |
|---|---|---|---|
| Pore volume (mL) | 10 ± 0.8 | 9.97 ± 0.09 | 10 ± 0.08 |
| Porosity (%) | 20 ± 0.16 | 19.93 ± 0.18 | 20 ± 00.16 |
| OOIP (mL) | 7.1± 0.08 | 7 ± 0.08 | 7.03 ± 0.08 |
| Soi (%) | 71 ± 0.73 | 70.23 ± 0.41 | 70.34 ± 1.45 |
| Swi (%) | 29 ± 0.73 | 29.76 ± 0.41 | 29.65 ± 0.46 |
| Sorwf (mL) | 4.53 ± 0.04 | 4.55 ± 0.04 | 4.52 ± 0.02 |
| Sor (%) | 35.85 ± 0.58 | 35 ± 0.69 | 35.77 ± 0.75 |
| Sorif (mL) | 1.51 ± 0.04 | 1.05 ± 0.05 | 0.80 ± 0.03 |
| AOR (%) | 61.18 ± 0.85 | 42.85 ± 0.84 | 32.06 ± 1.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yaraguppi, D.A.; Bagewadi, Z.K.; Mahanta, N.; Singh, S.P.; Khan, T.M.Y.; Deshpande, S.H.; Soratur, C.; Das, S.; Saikia, D. Gene Expression and Characterization of Iturin A Lipopeptide Biosurfactant from Bacillus aryabhattai for Enhanced Oil Recovery. Gels 2022, 8, 403. https://0-doi-org.brum.beds.ac.uk/10.3390/gels8070403
Yaraguppi DA, Bagewadi ZK, Mahanta N, Singh SP, Khan TMY, Deshpande SH, Soratur C, Das S, Saikia D. Gene Expression and Characterization of Iturin A Lipopeptide Biosurfactant from Bacillus aryabhattai for Enhanced Oil Recovery. Gels. 2022; 8(7):403. https://0-doi-org.brum.beds.ac.uk/10.3390/gels8070403
Chicago/Turabian StyleYaraguppi, Deepak A., Zabin K. Bagewadi, Nilkamal Mahanta, Surya P. Singh, T. M. Yunus Khan, Sanjay H. Deshpande, Chaitra Soratur, Simita Das, and Dimple Saikia. 2022. "Gene Expression and Characterization of Iturin A Lipopeptide Biosurfactant from Bacillus aryabhattai for Enhanced Oil Recovery" Gels 8, no. 7: 403. https://0-doi-org.brum.beds.ac.uk/10.3390/gels8070403