Non-Coding RNAs in Breast Cancer: Intracellular and Intercellular Communication


Department of Biochemistry & Molecular Genetics, University of Louisville School of Medicine, Louisville, KY 40292, USA
Non-Coding RNA 2018, 4(4), 40; https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna4040040
Submission received: 29 October 2018
/
Revised: 29 November 2018
/
Accepted: 4 December 2018
/
Published: 12 December 2018
(This article belongs to the Special Issue Non-Coding RNA in Reproductive Organ Cancers)
Abstract
:Non-coding RNAs (ncRNAs) are regulators of intracellular and intercellular signaling in breast cancer. ncRNAs modulate intracellular signaling to control diverse cellular processes, including levels and activity of estrogen receptor α (ERα), proliferation, invasion, migration, apoptosis, and stemness. In addition, ncRNAs can be packaged into exosomes to provide intercellular communication by the transmission of microRNAs (miRNAs) and long non-coding RNAs (lncRNAs) to cells locally or systemically. This review provides an overview of the biogenesis and roles of ncRNAs: small nucleolar RNA (snRNA), circular RNAs (circRNAs), PIWI-interacting RNAs (piRNAs), miRNAs, and lncRNAs in breast cancer. Since more is known about the miRNAs and lncRNAs that are expressed in breast tumors, their established targets as oncogenic drivers and tumor suppressors will be reviewed. The focus is on miRNAs and lncRNAs identified in breast tumors, since a number of ncRNAs identified in breast cancer cells are not dysregulated in breast tumors. The identity and putative function of selected lncRNAs increased: nuclear paraspeckle assembly transcript 1 (NEAT1), metastasis-associated lung adenocarcinoma transcript 1 (MALAT1), steroid receptor RNA activator 1 (SRA1), colon cancer associated transcript 2 (CCAT2), colorectal neoplasia differentially expressed (CRNDE), myocardial infarction associated transcript (MIAT), and long intergenic non-protein coding RNA, Regulator of Reprogramming (LINC-ROR); and decreased levels of maternally-expressed 3 (MEG3) in breast tumors have been observed as well. miRNAs and lncRNAs are considered targets of therapeutic intervention in breast cancer, but further work is needed to bring the promise of regulating their activities to clinical use.
Keywords:
ncRNA; miRNA; lncRNA; breast cancer; transcription; estrogen receptor; resistance; cancer stem cells1. Introduction
Breast cancer is the most commonly diagnosed cancer and second leading cause of cancer death among women in the United States (U.S.). Breast tumors are heterogeneous, and are pathologically classified according to their expression of key proteins by immunohistochemical (IHC) staining (at a minimum 1% [1]): estrogen receptor α (ERα), gene ESR1, tumors are termed ER positive (ER+), progesterone receptor (PR, gene PGR), and human epidermal growth factor receptor 2 (HER2, gene ERBB2). Tumors that lack these three protein markers are “basal-like” and referred to as triple negative breast cancer (TNBC). Most primary breast tumors are ER+/PR+/HER2-, and patients are treated with surgery, radiation, and endocrine therapies (also referred to as antiestrogen therapies) Endocrine therapies employ aromatase inhibitors (AI), e.g., letrozole, to block the conversion of androgens to estrogens, or tamoxifen (TAM), which is a selective ER modulator (SERM) that competes with estrogens, including estradiol (E2) for binding ER. Most postmenopausal women with ER+ breast tumors receive AI therapy, while American Society of Clinical Oncology (ASCO) guidelines recommend 10 years of TAM for premenopausal women [2]. Unfortunately, 30–40% of patients develop resistance to endocrine therapies and develop metastatic disease [3,4]. Multiple mechanisms are involved in acquired endocrine resistant breast cancer [5]. Approximately 25–40% of metastatic tumors in breast cancer patients treated with AIs show ESR1 mutations within the ligand binding domain (LBD) [6]. These mutations result in the ligand-independent transcriptional activity of the mutant ERα and reduce the efficacy of ER antagonists, including the selective ER downregulators (SERDs) fulvestrant, AZD9496, RU-58688, and GDC-0810 [7]. Both ESR1 Y537S and D538G mutations are associated with more aggressive disease biology and shorter survival [8]. Recent clinical data suggest that the addition of the CDK4/6 inhibitor palbociclib in combination with letrozole provides a benefit in advanced disease [9,10,11,12]. Gene expression analysis in primary breast tumors for 50 genes in the PAM50 test has allowed the further dissection of molecular phenotypes that have clinical implications for individualizing patient treatment [13]. As of yet, no non-coding RNAs (ncRNAs) are used in commercial diagnostic tests; however, there is great interest in identifying circulating microRNAs (miRNAs) [14,15,16] and long non-coding RNAs (lncRNAs) [17,18,19] in breast cancer diagnosis and for monitoring therapeutic response.
Over the past two decades, we have learned that 99% of the total cellular RNA content of human cells consists of ncRNAs that are classified by size and function [20]. Transfer RNA (tRNA) (89%) and ribosomal RNA (rRNA) (8.9%) constitute the majority of ncRNAs, followed in abundance by messenger RNAs (mRNAs) (0.9%). Thus, the remaining ncRNAs, including circular RNA (circRNA), small nuclear RNA (snRNA), small nucleolar RNA (snoRNA), miRNA, and lncRNA together account for ~1% of total ncRNA. Despite their low abundance, these ncRNAs play critical roles in transcription, post-transcriptional processing, and translation [21]. In addition, because ncRNA can be packaged into extracellular vesicles (EV), including exosomes [22], they provide a mechanism for intercellular communication by the transfer of miRNA and lncRNA to recipient cells both locally and systemically [23]. It is important to note that the levels of ncRNA expression, their post-transcriptional modification (particularly lncRNAs), and their subcellular distribution are important to consider in assigning their potential function [24]. As concluded by Palazzo and Lee, it is ultimately critical to examine the biological function of each identified ncRNA on a case-by-case basis [24]. Table 1 lists the best-characterized regulatory ncRNAs with roles in breast cancer, as well as their sizes and function.
LncRNAs and miRNAs are considered to be epigenetic regulators of human cancers [25]. Both pri-miRNAs and lncRNAs are post-transcriptionally modified, e.g., by the methylation on N6 of adenosine (m6A), which alters their activities, providing an additional layer of regulation termed “epitranscriptomics” [26]. In addition, there is one report that a pool of inactive miR-34 lacking the 5′-phosphate in cancer cells, including MCF-7 cells, is activated by 5′-end phosphorylation by ATM serine/threonine kinase (ATM) and cleavage and polyadenylation factor I subunit 1 (CLP1) in response to DNA damage, in turn allowing AGO2 loading [27]. At least three times more reports are listed in PubMed on miRNAs than lncRNAs in endocrine cancers [28].
2. Small Nuclear RNAs and Small Nucleolar RNAs in Breast Cancer
snRNAs and snoRNAs play roles in mRNA splicing and rRNA maturation, respectively (reviewed in [67]). Elevated snoRNA biogenesis is required for the high rRNA expression that is needed for ribosomal biogenesis and enhanced protein synthesis in breast cancer (reviewed in [68]). Some snoRNAs are expressed in a cell-specific manner, including SNORD46 and SNORD42A in mammary glands and lymphoblastoid cells in the ENCODE (Encyclopedia of DNA elements) dataset (reviewed in [69]). Thirteen snoRNAs were identified as having prognostic relevance in breast cancer, including the downregulation of SNORD46 and SNORD89 [70]. The clonal evolution of successful cancer cells is guided by intracellular and intercellular mutations that provide advantages or disadvantages to cell lineages. This evolution is mediated in part by ribosomal alterations that are driven by mutations in uL18/RPL5 ribosomal protein genes in ~34% of breast tumors [71]. Many snoRNAs are processed into stable miRNA-like oligomers that are called ‘small nucleolar RNA-derived RNAs’ (sdRNAs) [72]. sdRNA-93 was reported to play a role in MDA-MB-231 cell invasiveness, and is overexpressed in Luminal B/HER2+ breast tumors relative to other types of breast tumors or normal breast tissue [73]. Other studies show that alterations in snoRNAs may promote carcinogenesis and favor caner stem cell phenotypes [71].
3. Circular RNAs
During the splicing of exons and removal of introns from heterogeneous nuclear RNA (hnRNA) to generate mRNA, “backsplicing” can generate circular RNAs (circRNAs) that are usually exported to the cytoplasm. CircRNAs are considered stable, although they are expressed at <10% of their associated linear mRNA transcripts (reviewed in [74]). circRNAs are evolutionarily conserved, suggesting that they are critical for cellular activities. Several factors regulate circRNA levels, including exon-skipping events, complementary sequences of flanking introns, RNA-binding proteins, and the amount of more than 25 of the ~170 proteins that constitute the spliceosome [74]. circRNAs can act as competing endogenous RNA (ceRNA, also called ‘sponge’) for microRNAs, thus reducing miRNA repression of their target mRNAs [75]. circRNAs modulate the stability of mRNAs, interact with RNA binding proteins, and regulate gene transcription by interaction with RNA pol II [29]. In addition, recent studies show that some circRNAs are translated, including circ-SHPRH, which generates a 17 kDa SHPRH-146aa protein that acts as a novel tumor suppressor protein and a protective decoy for its full-length SHPRH protein in glioblastoma [76,77].
At least six circRNAs have been reported as having roles in breast cancer (Table 1). Examples of circRNAs that are increased in breast tumors and classified as having oncogenic activity in breast cancer cell lines by acting as ceRNA for miRNAs include: hsa_circ_ABCB10 (ceRNA for miR-1271 [34]); hsa_circ_0011946 (ceRNA for miR-26a/b and thus upregulating RFC3 in MCF-7 cells) [78], hsa_circ_0052112 (ceRNA for miR-125a-5p in MDA-MB-231 cells) [79], and hsa_circ_0008039 (ceRNA for miR-432-5p and thus increasing E2F3 expression in MCF-7 and BT-20 cells) [38]. Recently, hsa_circ_0007294 (circANKS1B) was shown to be upregulated in TNBC tumors compared with all other breast tumor types, and circANKS1B acts as a ceRNA for miR-148a-3p and miR-152-3p, resulting in the increased expression of USF1, which increases transforming growth factor β1 (TGF-β1) expression to stimulate epithelial-to-mesenchymal transformation (EMT) [80]. It is likely that the further examination of RNA profiling data will identify more circRNAs in breast tumors, which will necessitate further molecular and functional studies about their roles in tumor formation, progression, and metastasis.
4. PIWI-Interacting RNAs
piRNAs work in PIWI–piRNA complexes to regulate gene expression at the epigenetic and post-transcriptional levels. piRNAs are the guardians of genome integrity by epigenetically silencing transposable elements by DNA methylation [81]. PIWI proteins are aberrantly expressed in breast cancer, including a high expression of PIWIL2, which is a member of the PIWI/AGO gene family [82], and PIWIL4 is highly expressed in TNBC tumors and cell lines, and correlates with distant metastasis fatality [83]. Experiments in MDA-MB-231 TNBC cells demonstrated that PIWIL4 activates TGF-β, MAPK/ERK, and fibroblast growth factor (FGF) signaling, and suppresses the expression of MHC class II genes [83]. The analysis of existing small RNAs in–RNA seq data identified >100 piRNAs in breast cancer cells, some of which were differentially expressed in MCF-7, ZR-75.1, and SKBR3 breast cancer cells relative to MCF-10A immortalized ‘normal’ breast epithelial cells [41]. The role of piRNAs in breast tumors remains to be thoroughly examined.
5. MicroRNAs in Breast Cancer
Reflecting the intense interest in the role of miRNAs in breast cancer initiation, progression, and metastasis, there are currently ~3662 publications on miRNAs in breast cancer in PubMed. The reason behind this interest is that miRNAs are dysregulated in breast cancer (reviewed in [56,57,58,59,60,84]). Since each miRNA has the theoretical capacity to regulate multiple gene targets post-transcriptionally, alteration in the expression levels of just one miRNA has the potential to affect hundreds of target mRNAs. There are a number of programs to identify targets of a selected miRNA, and conversely, the miRNAs that may regulate a specific target gene (mRNA). Examples include miRTarBase, PicTar, miRanda, and MirAncesTar, although these programs predict false positives and false negatives (reviewed in [85]). Thus, experimental verification of the ability of a miRNA to downregulate a target requires cloning the 3′UTR of the target mRNA downstream of a luciferase reporter driven by an active promoter, e.g., CMV, mutating the miRNA recognition element in the reporter, and performing transient transfection assays in cells using the overexpression of wild-type miRNA and knockdown the miRNA expression. In addition, westerns and/or immunohistochemical staining (IHC) are needed to validate the decrease in authenticated target protein expression.
miRNAs are short (~22 nt) single-stranded RNAs that regulate mRNA stability and/or translation by base-pairing between the seed sequences at 5′ positions 2–7 or 2–8 of the miRNA with ~7 bp miRNA recognition elements (MREs) in the 3’UTR of their target mRNAs within the RNA-induced silencing complex (RISC) [86]. The current miRBase (release 22) reports 2654 mature miRNAs (http://www.mirbase.org/ [87]). Additional complexity is offered by the production of isomiRs that are generated from a single miRNA locus by template and non-template variants, and are differentially expressed in different types of human breast tumors [88]. The biogenesis of miRNAs has been extensively reviewed [28,89]. In summary, most miRNAs are transcribed as primary (pri)-miRNAs by RNA polymerase II, either cotranscribed within introns of host genes or as independent genes [90], and processed within the nucleus by the DROSHA-DGCR8 microprocessor complex [91]. DROSHA cleaves the hairpin-loop pri-miRNA yielding a 60–70 nt precursor (pre)-miRNA that is exported from the nucleus by the Exportin (XPO5) and Ran-GTP (RAN) or Exportin1 (XPO1, also called CRM1) [92]. The high/middle methylation of XPO5 was associated with reduced breast cancer risk, and XPO5 expression is increased in breast tumors [93]. An epidemiological study of key miRNA processing genes in the blood of male U.S. veterans reported that the DNA methylation of DROSHA and TNRC6B may play a role in early carcinogenesis [94]. DROSHA has tumor suppressor or oncogenic activity, depending on the type of tumor (reviewed in [95]). Single nucleotide polymorphisms (SNPs) in DROSHA are associated with increased breast cancer risk [96]. In the cytoplasm, the DICER-TRBP complex unwinds the double-stranded precursor miRNA (pre-miRNA) to allow the incorporation of one strand of the miRNA (called the guide strand) into the RISC complex that includes the catalytic Argonaut proteins, e.g., AGO2 [97]. The non-incorporated passenger strand of miRNA is degraded [98]. DICER is considered a tumor suppressor in breast cancer [95]. Increased DICER was associated with TAM resistance in metastatic breast tumors and tumor xenografts [99]. The high expression of AGO2 was reported to correlate with the luminal B subtype of breast cancer [100]. Interestingly, the overexpression of AGO2 in MCF-7 human breast cancer cells increased the expression of an ERα variant called ERα36, and stimulated E2-induced xenograft tumor growth in vivo in severe combined immunodeficiency (SCID)/Beige female mice [100]. This observation is of interest, since ERα36 plays a role in endocrine resistance and cancer stem cells (CSC) in breast cancer [101].
Next-generation sequencing approaches have identified ‘mitomiRs’ that function in mitochondria, whether by being imported from the repertoire of nuclear-encoded miRNAs or transcribed from the mitochondrial DNA (mtDNA) [102]. miRNAs may act as retrograde and anterograde signaling molecules between mitochondria and the nucleus to regulate energy homeostasis and apoptosis [102]. However, caution is needed, since the cytosolic contamination of mitochondrial extracts can lead to artifacts [102]. The role of mitomiRs in breast cancer is unknown. It is intriguing to note that PyMT mammary tumor metastasis was regulated by mtDNA in transgenic “mitochondrial-nuclear exchange (MNX) mice”, i.e., mice that had BL/6 mitochondria had longer tumor latency compared to those with FVB or BALB/c mitochondria, and mice with BALB/c mitochondria showed higher metastatic areas in lung [103,104]. At present, the precise mechanism(s) for this observation remain to be determined.
miRNAs that are overexpressed in breast tumor are called ‘oncomiRs’, since they promote carcinogenesis and progression by downregulating tumor suppressor genes. One example of an oncomiR in breast, as well as many cancers, is miR-21 [105,106,107,108,109,110,111,112,113,114,115]. Some examples of validated targets of miR-21 are indicated in Table 2. On the other hand, miRNAs that are downregulated in breast tumors are called tumor suppressor miRNAs. Notably, it is incorrect to assume that a miRNA is always an oncomiR or a tumor suppressor miRNA, since the activity of each miRNA depends on the cellular context, which includes mitigating factors such as lncRNAs and circRNAs (reviewed in [116]). It is important to note that many of the miRNAs that have been studied in breast cancer cell lines (reviewed in [53,59,117,118]) are not dysregulated in human breast tumors; thus, the focus here is on those identified in human breast tumors.
In breast cancer, miRNAs regulate genes involved in apoptosis, cell-signaling pathways (e.g., TGFβ [220], epithelial-to-mesenchymal transformation (EMT) [221], metastasis [222], the expression of ERα [117] and other nuclear receptors (NRs) [50]), regulation of the tumor microenvironment, and stemness, which includes transfer or exosomal miRNAs to adjacent normal fibroblasts forming cancer-associated fibroblasts (CAFs) (reviewed in [223]). Figure 1, Figure 2 and Figure 3 highlight the pathways of some of the miRNAs dysregulated in breast cancer, as summarized in Table 2 and Table 3. In addition, the lncRNAs that have been reported to ‘sponge’ some of the dysregulated miRNAs are shown as competing endogenous RNA (ceRNA).
Croce’s group first identified miRNAs dysregulated in breast cancer in 2005 [233]. There are numerous reviews covering 13 years of publications on the identity, targets, and regulation of miRNAs in breast cancer, and these references are not fully inclusive of all the published work [43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62]. The identity and possible roles of miRNA dysregulated in specific subtypes of breast cancer, including HER2+ [261,262], TNBC [263,264,265], and endocrine-resistant breast cancer [16,52,54,59,60] have been reviewed.
6. Long Non-Coding RNAs in Breast Cancer
The current GENECODE (version 28, GRCh38.p12) of the human genome includes 58,381 genes, 15,779 long non-coding RNAs (lncRNAs), and 1881 miRNAs (https://www.gencodegenes.org/stats/archive.html#a28). LncRNAs are defined as ncRNAs of >200 nucleotides [64]. They are transcribed from genomic DNA by RNA pol II, and are classified according to the genomic organization: (1) intergenic lncRNAs (lincRNAs) are transcribed between two protein-coding genes; (2) intronic lncRNAs are transcribed from the introns of protein-coding genes; (3) overlapping lncRNAs, which overlap protein-coding genes; and 4) antisense (as) lncRNAs, which are transcribed in a direction opposite to that of the protein-coding gene [65]. lncRNA expression is a low percent of total cellular RNA, i.e., ~0.03–0.20% [266]. Thus, while 15,779 lncRNAs have been identified, very few have been characterized with respect to cell type specificity and function. There is a need to develop tools for an improved analysis of differentially expressed lncRNAs. Since lncRNAs are low in expression, their expression levels are “very noisy”, which reflects their low counts in RNA-seq data [267]. Overall, quantitative examination of the identity of lncRNAs and their roles is needed, including in breast cancer.
In addition to nuclear-encoded lncRNAs, seven lncRNAs were identified as mitochondrial DNA transcripts mtDNA [268]. Two lncRNAs are encoded by mitochondrial D-loop regions: MDL1 and MDL1AS [269]. There is evidence of mt-encoded lncRNAs in the nucleus, suggesting a potential role in retrograde signaling [268]; however, there are no reports on mtDNA-encoded lncRNAs in breast cancer.
Functionally, lncRNAs regulate transcription by associating with enhancer regions, in cis, i.e., at adjacent sites relative to their own transcription, or in trans, i.e., at more distal sites [270]. lncRNAs have heterogeneous complex 3D structures, which allows them to assume different shapes and interact with a wide variety of intracellular components. lncRNAs interact with the Mediator complex, forming loops between enhancer and promoter regions [266]. lncRNAs interact with proteins and other RNAs to influence their activities and cellular location. lncRNAs regulate development and differentiation, gene imprinting, and antiviral responses; and assist in chromatin modification, mRNA splicing, and protein stability [271]. Another function of lncRNAs is to act as ‘sponges’ (ceRNAs) for miRNAs, thus blocking the repressive activity of miRNAs for binding to the 3’UTR of their target transcripts. A network analysis of ncRNAs in cancer drug resistance-associated lncRNAs–miRNAs, TAM resistance (including lncRNAs MALAT1 and CCAT2; miR-221, miR-222, miR-26a, miR29a, miR-29b), and Trastuzumab resistance (lncRNA GAS5, miR-16, and miR-155) has been described [272]. lncRNAs act as scaffolds, e.g., HOTAIR links the PRC2 and LSD1 histone-modifying complexes to promote histone H3K27 methylation and H3K4 demethylation to silence target genes and promote breast cancer metastasis [273].
The identity and roles of lncRNAs in breast cancer have been reviewed [28,52,274,275,276,277,278,279,280,281,282]. lncRNA expression profiles have been correlated with hormone status and intrinsic tumor type [283,284]. At present, there is no consistent identification of which lncRNAs in primary tumors are the best signature for predicting patient outcomes. For example, using four Gene Expression Omnibus (GEO) datasets (n = 473 breast cancer patients), one group identified a 12-lncRNA predictive signature for recurrence (RP1-34M23.5, RP11-202K23.1, RP11-560G2.1, RP4-591L5.2, RP13-104F24.2, RP11-506D12.5, ERVH48-1, RP4-613B23.1, RP11-360F5.1, CTD-2031P19.5, RP11-247A12.8, and SNHG7) [285], and another group using TCGA (n = 1064) identified a different set of three lncRNAs as prognostic markers (CAT104, LINC01234, and STXBP5-AS1) [286]. The differences may be the result of different platforms—the Affymetrix HG-U133 Plus 2.0 platform versus RNA-seq—in addition to the different tumors analyzed, the heterogeneity of breast tumors, the method of lncRNA identification and data interrogation, and, as stated earlier, the low level of lncRNA expression that confounds statistically relevant data interpretation. For this review, a selection of nuclear encoded lncRNAs that have been identified as dysregulated in breast tumors will be briefly summarized.
The lncRNA RMRP (RNA component of mitochondrial RNA processing endoribonuclease) binds RNA-binding proteins GRSF1, HUR, and PNPASE for transport into mitochondria, where RMRP plays a role in RNA processing and mtDNA replication [287]. Mutations were identified in the promoters of lncRNAs RMRP and nuclear paraspeckle assembly transcript 1 (NEAT1) that increased their expression in human breast tumors [288]. However, the expression of NEAT1 showed no correlation with clinical gene signatures associated with higher grade, stage, metastasis, tumor aggression, or TAM resistance. Thus, the role of NEAT1 in breast cancer is unclear [289]. Patients whose primary breast tumors showed a high expression of NEAT1, colon cancer associated transcript 2 (CCAT2), or metastasis associated lung adenocarcinoma transcript 1 (MALAT1) had shorter overall survival (OS) [282].
NEAT1 is involved in the organization of nuclear architecture called paraspeckles for gene transcription and splicing [66]. Nuclear speckles are dynamic punctate compartments in the nucleus that contain components of the pre-mRNA spliceosome, including SRSFs, small nuclear ribonucleoproteins (snRNPs), RNA Pol II subunits, 3’ end processing proteins, m6A writers METTL3/14 and reader YTHDC1, and various protein kinases that regulate the pool of proteins in the speckles [290,291]. NEAT1 was identified as an essential component of the FOXN3–SIN3A repressor complex, and the overexpression of NEAT1 promoted EMT in ERα+ MCF-7 breast cancer cells and promoted the lung metastasis of MCF-7 when orthotopically implanted in the mammary fat pad, suggesting that NEAT1 has oncogenic activity [292]. Bioinformatic analysis of sample-matched miRNA-seq and RNA-SeqV2 data of breast cancer from The Cancer Genome Atlas (TCGA) revealed that NEAT1 was overexpressed in luminal A, luminal B, HER2+, and basal-like (TNBC) tumors [293]. Further, the authors identified a putative ceRNA network for NEAT1, as well as lncRNAs OPI5-AS1 and AC008124.1 in all breast tumors and each subtype [293]. NEAT1 was also identified in a gene (ESR1, DKC1)–lncRNA (TERC and TUG1) interaction network in breast tumors from TCGA [294].
Another integrative analysis of RNA-seq data of ~1000 breast tumors in TCGA identified GATA3-AS1 (ENSG00000197308), RP11-279F6 (ENSG00000245750), and AC017048 as highly expressed in ERα-positive versus ERα-negative breast tumors and normal breast tissue samples [295]. However, there are no confirming reports on these three lncRNAs or their function in breast cancer. Another analysis of TGCA RNA-seq data of human breast tumors identified a decreased expression of LINC00092 and C2orf71 as associated with poor prognosis, and identified a putative network of coexpression of LINC00092 with mRNAs RGMA and SFRP1 that were regulated by miR-449a and miR-452-5p [296]. However, neither of these miRNAs were dysregulated in breast tumors [61]. Further, C2orf71 has been identified as a protein-coding gene PCARE in GeneCards.
The lncRNA SRA1, which is a steroid receptor RNA activator, was first identified in a complex with the coactivator SRC-1 (NCOA1) as an RNA coactivator that increased the transcriptional activity of NR, including ERα [297]. SRA1 expression was higher in breast tumors compared with adjacent normal breast tissue [298]. SRA1 is unique in that not only does it encode lncSRA1s of different length [299], but SRA1 also encodes a protein: steroid receptor co-activator protein (SRAP) (reviewed in [300]). SRAP does not interact with the lncRNA SRA1, but it interacts with NRs: ERα, androgen receptor (AR), and glucocorticoid receptor (GR), and is involved in splicing and cell cycle regulation [301]. The lncRNA SRA1 functions as a scaffold, and interacts with miRNA processing (DICER and TRBP) and RISC components, i.e., AGO2 and PACT; with other transcription factors, e.g., OCT4, NANOG, and FOXO1; and with chromatin modifiers and binding proteins, e.g., KMT2A (MLL1), KMT2D (MLL2), EZH2, and CTCF [301].
MALAT1 (metastasis-associated lung adenocarcinoma transcript 1) is one of the best characterized lncRNAs with roles in neural development and function, retina, myogenesis, and vascular cell proliferation; it is dysregulated in cancers, including upregulation in breast tumors (reviewed in [302]). Serum levels of MALAT1, as examined by quantitative real-time PCR (qPCR), were higher in breast cancer patients (n = 157) than normal women (n = 107) [303]. MALAT1 expression is associated with ERα+/PR+ breast tumors and with lower relapse-free survival (RFS) [282]. MALAT1 staining was also higher in formalin-fixed paraffin embedded (FFPE) breast tumors than normal tissue [304]. MALAT1 is oncogenic in breast cancer, and it upregulates the WNT/β-catenin (CTNNB1) pathway [305]. MALAT1 mutations are frequent in breast tumors [306,307]. MALAT1 acts as a ceRNA for miR-9, miR-133, miR-145, miR-195, miR-200s, miR-205, miR-206, and miR-503 (reviewed in [302]). MALAT1 is targeted by interaction with miR-101, miR-125b, and miR-217. MALAT1 acts as scaffold to position nuclear speckles at active gene loci. Capture hybridization analysis of RNA targets (CHART) revealed binding of MALAT1 at actively transcribed loci (reviewed in [302]. Proteins interacting with MALAT1 were identified by SILAC (stable isotope labeling with amino acids) labeling and LC-MS/MS proteomics in HEPG2 human hepatoma cells, and included proteins involved in RNA processing, splicing and gene transcription, and HNRNPAB [308]. Despite its lower expression in HER2+ and TNBC tumors, MALAT1 expression was associated with decreased disease-specific survival in these patients [309]. Despite these studies implicating MALAT1 as oncogenic, a recent study in MMTV-PyMT;Malat1−/− mice demonstrated that MALAT1 is a suppressor of lung metastasis in this model [251]. Notably, there was no difference in overall survival or tumor weight in the MMTV-PyMT;Malat1+/+ versus MMTV-PyMT;Malat1−/− mice and no difference in histological metrics of the tumors. The authors also reported that in TCGA RNA seq data, MALAT1 was “underexpressed in human breast tumors compared with normal breast tissue”, and that lower MALAT1 levels correlated with shorter distant metastasis-free survival in all breast cancer, as well as in luminal A and basal breast cancer [251]. Using chromatin isolation by RNA purification coupled to mass spectrometry (ChIRP-MS) studies, they demonstrated that Malat1 sequestered the transcription factor TEAD, thus inhibiting its activity in mouse mammary tumors [251].
The expression of the lncRNA CCAT2 (Colon Cancer Associated Transcript 2) is increased in breast tumors and breast cancer cells relative to normal breast tissue/cells [310,311]. Knockdown of CCAT2 inhibited MCF-7 and MDA-MB-231 breast cancer cell proliferation and invasion in transwell migration assays by inhibiting WNT/β-catenin signaling [311]. CCAT2 expression was higher in MDA-MB-231 and LCC9 TAM-resistant breast cancer cells derived from MCF-7 cells than in parental MCF-7 cells, and knockdown of CCAT2 inhibited the activation of TGFβ signaling in LCC9 and MCF-7 cells [312]. CCAT2 was reported to bind EZH2 and increased H3K27me3 in chromatin, thus repressing CDKN2B transcript and protein expression in MDA-MB-231 cells [313]. CDKN2B encodes a cyclin-dependent kinase inhibitor that interacts with CDK4 or CDK6, thus preventing interaction with cyclin D and inhibiting G1-S cell cycle progression [314]. Thus, the repression of CDKN2B would allow cell cycle progression. These results are as of yet unconfirmed in human breast tumors. The authors suggest the potential for therapeutic agents targeting cellular pathways linked to MALAT1.
Levels of the lncRNA CRNDE (colorectal neoplasia differentially expressed) were higher in breast tumors than normal breast and correlated with reduced OS [315]. CRNDE is a ceRNA for miR-136, resulting in the activation of WNT/β-catenin signaling in MDA-MB-231 cells [315]. Wnt signaling is a key driver of stem cells in embryonic and adult tissues and CSCs [316,317].
Analysis of TCGA breast tumor data identified higher expression of the lncRNA MIAT (myocardial infarction associated transcript) in breast tumors than normal breast tissue [184]. MIAT is a ceRNA for miR-155-5p, and knockdown of MIAT increased expression of the miR-155-5p target DUSP7 and inhibited MDA-MB-231 cell proliferation and xenograft tumor growth [184].
The lncRNA LINC-ROR was discovered in pluripotent stem cells, where it functions as a ceRNA for miR-145 to increase key pluripotency transcription factors OCT4, NANOG, and SOX2 [318]. LINC-ROR expression was higher in breast tumors and cell lines relative to normal breast tissue, and MCF-10A cells and the overexpression of LINC-ROR induced markers of EMT in MCF-10A cells [319]. High LINC-ROR expression in breast tumors was associated with reduced OS, and knockdown of LINC-ROR inhibited TGFβ signaling in MCF-7 and MDA-MB-231 cells [320]. Whether LINC-ROR is involved in the stimulation of CSC or metastasis remains to be examined.
Conversely, the expression of MEG3 (Maternally Expressed 3) is lower in breast tumors than in normal breast tissues, and the expression level of MEG3 was negatively correlated with histological tumor grade [282], and MEG3 downregulation correlated with poor OS [321]. MEG3 is a ceRNA for miR-421 in MDA-MB-231 TNBC and increased E-cadherin, while decreasing cell invasion in vitro [322]. The overexpression of MEG3 in MDA-MB-231 cells suppressed xenograft tumor growth and angiogenesis, and reduced P-AKT, PCNA, and MMP-9 protein expression in the tumors formed in the mice [323]. Whether MEG3 could be a potential therapy in breast cancer is not yet known, but is of interest in nervous system cancers [324].
lncRNAs continue to be discovered through the bioinformatic analysis of breast tumor data. For example, a recently published analysis of TCGA breast tumor identified putative driver lncRNAs: the amplification of AC084809.2, RP11-108P20.3, ACOO5076.5, RP11-385J1.2, RP11-567G11.1, LINC00909, ATP1B3-AS1, MCCC1-AS1, YEATS2-AS1, TP53TG1, and SOX2-OT [325]. As suggested previously, much more research is needed to understand the roles of lncRNAs in breast cancer initiation, progression, and metastasis.
7. Micro RNAs and Long Non-Coding RNAs in Extracellular Vesicles and Exosomes
Exosomes are a type of extracellular vesicle (EV) that are ~50–140 nm in size, endosome-derived, and secreted by most cells under normal and disease states [326]. Surface markers on exosomes include CD9, CD63, CD81, LAMP1, and TSG101 [327]. Microvesicles (MVs, also called ‘ectosomes’) are EVs that bud from the plasma membrane surface, but many people refer to MVs and exosomes interchangeably [328]. Exosomes contain ncRNA (circRNA, miRNA, lncRNA) in addition to dsDNA, mRNA, proteins, lipids, DICER, and TRBP, as well as AGO2, and process pre-miRNAs to mature miRNAs [327]. A web-based database exoRBase is a repository of 15,501 lncRNA, 58,330 circRNA, and 18,333 mRNA [329]. Exosome levels are higher in the serum of breast cancer patients compared to normal subjects [330]. EVs deliver cargo, including ncRNA, lipids, mtDNA, and proteins, between cells and play roles in the context of tumor growth and the stimulation of metastasis while suppressing immune detection [328,331]. EVs, and exosomes in particular, are considered of great promise as ‘liquid biopsy’ and biomarkers in cancer detection and monitoring therapeutic response [332]. Examples of exosomal miRNAs that are released by breast cancer cells and found in higher amount in the serum of breast cancer patients than controls include miR-21, miR-195, miR-484, and miR 1246 [332]. However, there is concern about the manner of sample collection and variability in content of EVs, and exosomes due to different methods of isolation that are yet to be standardized for clinical lab testing [328].
The role of exosomal miRNAs in breast cancer has been reviewed [60,333,334]. Exosomes are secreted by cancer cells and by cancer-associated fibroblasts, and can be taken up by neighboring cells in the tumor environment, including NK cells and T lymphocytes, as well as distant cells after traveling in blood [60]. Examples of exosomal miRNAs and their targets in breast cancer are shown in Figure 4. Exosomes secreted from TAM-resistant MCF-7 cells in vitro were shown to confer TAM resistance to parental MCF-7 cells, in part by delivering miR-222 and miR-221 that repress ESR1 (ERα) translation [335]. Exosomes from TAM-resistant LCC2 cells, derived from MCF-7 cells, had higher levels of the lncRNA UCA1 than parental MCF-7 cells, and the incubation of MCF-7 cells with exosomes from LCC2 cells conferred TAM-resistance [336]. We reported that miR-29b-1/a directly targets ATP synthase subunit genes ATP5G1 and ATPIF1 and inhibit OCR and decrease ATP in MCF-7 and LCC9 breast cancer cells [337]. UCA1 activates AKT-mTOR signaling [338] and is a ceRNA for miR-18a, thus derepressing HIF-1α [137]. The lncRNA MALAT1 is found in exosomes from breast cancer patients [339]. MALAT1 is known to activate WNT–β-catenin signaling, and is oncogenic in breast cancer (reviewed in [28]). Most studies have characterized the activities of miRNAs and lncRNAs in cancer cell exosomes in vitro; thus, research is needed to validate the role of these miRNAs and lncRNAs in breast cancer metastasis in vivo.
8. Concluding Considerations
ncRNAs are dysregulated in breast cancer and miRNAs, noted for their stability, and lncRNAs are being investigated as biomarkers of disease pathology, prognostic indicators, and potential therapeutic targets, as well as modalities to block cancer progression and metastasis. We know much more about miRNAs than lncRNAs in human breast tumors. Our understanding of the networks between miRNAs, lncRNAs, their interacting partners, and targets is expanding. However, individual and combinations of miRNAs and lncRNAs have cell-specific activities that are not fully understood, nor is their interaction with the immune system, microbiome, microenvironment, hormonal milieu, or metabolome elucidated. Further investigation is needed to bring studies on miRNAs and lncRNAs into clinical practice for diagnosis, prognosis, and therapeutics in breast cancer patients.
Funding
The writing of this review of existing published research did not receive any specific grant from any funding agency in the public, commercial or not-for-profit sector.
Conflicts of Interest
I declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the research reviewed here.
References
- Hammond, M.E.; Hayes, D.F.; Dowsett, M.; Allred, D.C.; Hagerty, K.L.; Badve, S.; Fitzgibbons, P.L.; Francis, G.; Goldstein, N.S.; Hayes, M.; et al. American Society of Clinical Oncology/College of American Pathologists guideline recommendations for immunohistochemical testing of estrogen and progesterone receptors in breast cancer (unabridged version). Arch. Pathol. Lab. Med. 2010, 134, e48–e72. [Google Scholar] [PubMed]
- Burstein, H.J.; Temin, S.; Anderson, H.; Buchholz, T.A.; Davidson, N.E.; Gelmon, K.E.; Giordano, S.H.; Hudis, C.A.; Rowden, D.; Solky, A.J.; et al. Adjuvant Endocrine Therapy for Women With Hormone Receptor–Positive Breast Cancer: American Society of Clinical Oncology Clinical Practice Guideline Focused Update. J. Clin. Oncol. 2014, 32, 3959–3966. [Google Scholar] [CrossRef] [PubMed]
- Ring, A.; Dowsett, M. Mechanisms of tamoxifen resistance. Endocr Rel. Cancer 2004, 11, 643–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piggott, L.; da Silva, A.M.; Robinson, T.; Santiago–Gómez, A.; Simões, B.M.; Becker, M.; Fichtner, I.; Andera, L.; Piva, M.; Vivanco, M.d.; et al. Acquired resistance of ER-positive breast cancer to endocrine treatment confers an adaptive sensitivity to TRAIL through post–translational downregulation of c–FLIP. Clin. Cancer Res. 2018, 24, 2452–2463. [Google Scholar] [CrossRef] [PubMed]
- Clarke, R.; Tyson, J.J.; Dixon, J.M. Endocrine resistance in breast cancer—An overview and update. Mol. Cell. Endocrinol. 2015, 418, 220–234. [Google Scholar] [CrossRef] [PubMed]
- Jeselsohn, R.; De Angelis, C.; Brown, M.; Schiff, R. The Evolving Role of the Estrogen Receptor Mutations in Endocrine Therapy-Resistant Breast Cancer. Curr. Oncol. Rep. 2017, 19, 35. [Google Scholar] [CrossRef]
- Toy, W.; Weir, H.; Razavi, P.; Lawson, M.; Goeppert, A.U.; Mazzola, A.M.; Smith, A.; Wilson, J.; Morrow, C.; Wong, W.L.; et al. Activating ESR1 Mutations Differentially Affect the Efficacy of ER Antagonists. Cancer Discov. 2017, 7, 277–287. [Google Scholar] [CrossRef]
- Chandarlapaty, S.; Chen, D.; He, W.; Sung, P.; Samoila, A.; You, D.; Bhatt, T.; Patel, P.; Voi, M.; Gnant, M.; et al. Prevalence of ESR1 mutations in cell-free DNA and outcomes in metastatic breast cancer: A secondary analysis of the bolero-2 clinical trial. JAMA Oncol. 2016, 2, 1310–1315. [Google Scholar] [CrossRef]
- Finn, R.S.; Martin, M.; Rugo, H.S.; Jones, S.; Im, S.A.; Gelmon, K.; Harbeck, N.; Lipatov, O.N.; Walshe, J.M.; Moulder, S.; et al. Palbociclib and Letrozole in Advanced Breast Cancer. N. Engl. J. Med. 2016, 375, 1925–1936. [Google Scholar] [CrossRef]
- Ayyagari, R.; Tang, D.; Patterson-Lomba, O.; Zhou, Z.; Xie, J.; Chandiwana, D.; Dalal, A.A.; Niravath, P.A. Progression-free Survival With First-line Endocrine-based Therapies Among Postmenopausal Women With HR+/HER2− Metastatic Breast Cancer: A Network Meta-analysis. Clin. Ther. 2018, 40, 628–639.e623. [Google Scholar] [CrossRef]
- Kornblum, N.; Zhao, F.; Manola, J.; Klein, P.; Ramaswamy, B.; Brufsky, A.; Stella, P.J.; Burnette, B.; Telli, M.; Makower, D.F.; et al. Randomized Phase II Trial of Fulvestrant Plus Everolimus or Placebo in Postmenopausal Women With Hormone Receptor-Positive, Human Epidermal Growth Factor Receptor 2-Negative Metastatic Breast Cancer Resistant to Aromatase Inhibitor Therapy: Results of PrE0102. J. Clin. Oncol. 2018, 36, 1556–1563. [Google Scholar] [PubMed]
- Rugo, H.S.; Turner, N.C.; Finn, R.S.; Joy, A.A.; Verma, S.; Harbeck, N.; Masuda, N.; Im, S.-A.; Huang, X.; Kim, S.; et al. Palbociclib plus endocrine therapy in older women with HR+/HER2− advanced breast cancer: A pooled analysis of randomised PALOMA clinical studies. Eur. J. Cancer 2018, 101, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Cejalvo, J.M.; Pascual, T.; Fernández–Martínez, A.; Brasó–Maristany, F.; Gomis, R.R.; Perou, C.M.; Muñoz, M.; Prat, A. Clinical implications of the non-luminal intrinsic subtypes in hormone receptor-positive breast cancer. Cancer Treat. Rev. 2018, 67, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Huo, D.; Clayton, W.M.; Yoshimatsu, T.F.; Chen, J.; Olopade, O.I. Identification of a circulating microRNA signature to distinguish recurrence in breast cancer patients. Oncotarget 2016, 7, 55231–55248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleivi Sahlberg, K.; Bottai, G.; Naume, B.; Burwinkel, B.; Calin, G.A.; Børresen-Dale, A.-L.; Santarpia, L. A Serum MicroRNA Signature Predicts Tumor Relapse and Survival in Triple-Negative Breast Cancer Patients. Clin. Cancer Res. 2015, 21, 1207–1214. [Google Scholar] [CrossRef] [PubMed]
- Egeland, N.G.; Lunde, S.; Jonsdottir, K.; Lende, T.H.; Cronin-Fenton, D.; Gilje, B.; Janssen, E.A.; Soiland, H. The Role of MicroRNAs as Predictors of Response to Tamoxifen Treatment in Breast Cancer Patients. Int. J. Mol. Sci. 2015, 16, 24243–24275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, Y.; Fan, R.; Chen, L.; Qian, H. Clinical Significance of Long Non-coding RNA MALAT1 Expression in Tissue and Serum of Breast Cancer. Ann. Clin. Lab. Sci. 2016, 46, 418–424. [Google Scholar]
- Yu, G.; Zhang, W.; Zhu, L.; Xia, L. Upregulated long non-coding RNAs demonstrate promising efficacy for breast cancer detection: A meta-analysis. OncoTargets Ther. 2018, 11, 1491–1499. [Google Scholar] [CrossRef]
- Zhao, T.; Wu, L.; Li, X.; Dai, H.; Zhang, Z. Large intergenic non-coding RNA-ROR as a potential biomarker for the diagnosis and dynamic monitoring of breast cancer. Cancer Biomark. Sect. A Dis. Mark. 2017, 20, 165–173. [Google Scholar] [CrossRef]
- Romano, G.; Veneziano, D.; Acunzo, M.; Croce, C.M. Small non-coding RNA and cancer. Carcinogenesis 2017, 38, 485–491. [Google Scholar] [CrossRef]
- Cech Thomas, R.; Steitz Joan, A. The Noncoding RNA Revolution—Trashing Old Rules to Forge New Ones. Cell 2014, 157, 77–94. [Google Scholar] [CrossRef]
- Meldolesi, J. Exosomes and Ectosomes in Intercellular Communication. Curr. Biol. 2018, 28, R435–R444. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Yang, S.; Zhou, Q.; Wang, G.; Song, J.; Li, Z.; Zhang, Z.; Xu, J.; Xia, K.; Chang, Y.; et al. Emerging role of exosome-derived long non-coding RNAs in tumor microenvironment. Mol. Cancer 2018, 17, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palazzo, A.F.; Lee, E.S. Non-coding RNA: What is functional and what is junk? Front. Genet. 2015, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Liz, J.; Esteller, M. lncRNAs and microRNAs with a role in cancer development. Biochim. Biophys. Acta Gene Regul. Mech. 2016, 1859, 169–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Xiong, X.; Yi, C. Epitranscriptome sequencing technologies: Decoding RNA modifications. Nat. Methods 2016, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Salzman, D.W.; Nakamura, K.; Nallur, S.; Dookwah, M.T.; Metheetrairut, C.; Slack, F.J.; Weidhaas, J.B. miR-34 activity is modulated through 5′-end phosphorylation in response to DNA damage. Nat. Commun. 2016, 7, 10954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klinge, C.M. Non-coding RNAs: Long non-coding RNAs and microRNAs in endocrine-related cancers. Endocr.–Relat. Cancer 2018, 25, R259–R282. [Google Scholar] [CrossRef]
- Meng, S.; Zhou, H.; Feng, Z.; Xu, Z.; Tang, Y.; Li, P.; Wu, M. CircRNA: Functions and properties of a novel potential biomarker for cancer. Mol. Cancer 2017, 16, 94. [Google Scholar] [CrossRef]
- Wan, Y.; Hopper, A.K. Size matters: Conserved proteins function in length-dependent nuclear export of circular RNAs. Genes Dev. 2018, 32, 600–601. [Google Scholar] [CrossRef]
- Barrett, S.P.; Salzman, J. Circular RNAs: Analysis, expression and potential functions. Development 2016, 143, 1838. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Fan, X.; Mao, M.; Song, X.; Wu, P.; Zhang, Y.; Jin, Y.; Yang, Y.; Chen, L.-L.; Wang, Y.; et al. Extensive translation of circular RNAs driven by N6-methyladenosine. Cell Res. 2017, 27, 626–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, A.A.; Niu, N.; Tang, X.; Thompson, K.J.; Wang, L.; Kocher, J.P.; Subramanian, S.; Kalari, K.R. Circular RNAs and their associations with breast cancer subtypes. Oncotarget 2016, 7, 80967–80979. [Google Scholar] [CrossRef]
- Liang, H.F.; Zhang, X.Z.; Liu, B.G.; Jia, G.T.; Li, W.L. Circular RNA circ-ABCB10 promotes breast cancer proliferation and progression through sponging miR-1271. Am. J. Cancer Res. 2017, 7, 1566–1576. [Google Scholar] [PubMed]
- Tang, Y.Y.; Zhao, P.; Zou, T.N.; Duan, J.J.; Zhi, R.; Yang, S.Y.; Yang, D.C.; Wang, X.L. Circular RNA hsa_circ_0001982 Promotes Breast Cancer Cell Carcinogenesis Through Decreasing miR-143. DNA Cell Biol. 2017, 36, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Coscujuela Tarrero, L.; Ferrero, G.; Miano, V.; De Intinis, C.; Ricci, L.; Arigoni, M.; Riccardo, F.; Annaratone, L.; Castellano, I.; Calogero, R.A.; et al. Luminal breast cancer-specific circular RNAs uncovered by a novel tool for data analysis. Oncotarget 2018, 9, 14580–14596. [Google Scholar] [PubMed]
- Wu, J.; Jiang, Z.; Chen, C.; Hu, Q.; Fu, Z.; Chen, J.; Wang, Z.; Wang, Q.; Li, A.; Marks, J.R.; et al. CircIRAK3 sponges miR-3607 to facilitate breast cancer metastasis. Cancer Lett. 2018, 430, 179–192. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, C.; Zhou, Y.; Zhang, Z.; Sun, L. Circular RNA hsa_circ_0008039 promotes breast cancer cell proliferation and migration by regulating miR-432-5p/E2F3 axis. Biochem. Biophys. Res. Commun. 2018, 502, 358–363. [Google Scholar] [CrossRef]
- Wery, M.; Kwapisz, M.; Morillon, A. Noncoding RNAs in gene regulation. Wiley Interdiscip. Rev. Syst. Biol. Med. 2011, 3, 728–738. [Google Scholar] [CrossRef]
- Luteijn, M.J.; Ketting, R.F. PIWI-interacting RNAs: From generation to transgenerational epigenetics. Nat. Rev. Genet. 2013, 14, 523. [Google Scholar] [CrossRef]
- Hashim, A.; Rizzo, F.; Marchese, G.; Ravo, M.; Tarallo, R.; Nassa, G.; Giurato, G.; Santamaria, G.; Cordella, A.; Cantarella, C.; et al. RNA sequencing identifies specific PIWI-interacting small non-coding RNA expression patterns in breast cancer. Oncotarget 2014, 5, 9901–9910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, A.; Jacobs, D.I.; Hoffman, A.E.; Zheng, T.; Zhu, Y. PIWI-interacting RNA 021285 is involved in breast tumorigenesis possibly by remodeling the cancer epigenome. Carcinogenesis 2015, 36, 1094–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iorio, M.V.; Croce, C.M. microRNA involvement in human cancer. Carcinogenesis 2012, 33, 1126–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulrane, L.; McGee, S.F.; Gallagher, W.M.; O’Connor, D.P. miRNA Dysregulation in Breast Cancer. Cancer Res. 2013, 73, 6554–6562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Previati, M.; Manfrini, M.; Galasso, M.; Zerbinati, C.; Palatini, J.; Gasparini, P.; Volinia, S. Next generation analysis of breast cancer genomes for precision medicine. Cancer Lett. 2013, 339, 1–7. [Google Scholar] [CrossRef]
- Singh, R.; Mo, Y.Y. Role of microRNAs in breast cancer. Cancer Biol. Ther. 2013, 14, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Teng, R.; Wang, Q.; Xu, C.; Guo, J.; Yuan, C.; Shen, J.; Hu, W.; Wang, L.; Xie, S. Endocrine resistance in breast cancer: Current status and a perspective on the roles of miRNAs (Review). Oncol. Lett. 2013, 6, 295–305. [Google Scholar] [CrossRef] [Green Version]
- Piatek Monica, J.; Werner, A. Endogenous siRNAs: Regulators of internal affairs. Biochem. Soc. Trans. 2014, 42, 1174. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, J.; Wang, G. The role of microRNAs in human breast cancer progression. Tumour. Biol. 2014, 35, 6235–6244. [Google Scholar] [CrossRef]
- Cathcart, P.; Lucchesi, W.; Ottaviani, S.; De Giorgio, A.; Krell, J.; Stebbing, J.; Castellano, L. Noncoding RNAs and the control of signalling via nuclear receptor regulation in health and disease. Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 529–543. [Google Scholar] [CrossRef]
- Green, T.M.; Alpaugh, M.L.; Barsky, S.H.; Rappa, G.; Lorico, A. Breast Cancer-Derived Extracellular Vesicles: Characterization and Contribution to the Metastatic Phenotype. BioMed. Res. Int. 2015, 2015, 634865. [Google Scholar] [CrossRef] [PubMed]
- Hayes, E.L.; Lewis-Wambi, J.S. Mechanisms of endocrine resistance in breast cancer: An overview of the proposed roles of noncoding RNA. Breast Cancer Res. 2015, 17, 542. [Google Scholar] [CrossRef] [PubMed]
- Klinge, C.M. miRNAs regulated by estrogens, tamoxifen, and endocrine disruptors and their downstream gene targets. Mol. Cell Endocrinol. 2015, 418, 273–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Schooneveld, E.; Wildiers, H.; Vergote, I.; Vermeulen, P.B.; Dirix, L.Y.; Van Laere, S.J. Dysregulation of microRNAs in breast cancer and their potential role as prognostic and predictive biomarkers in patient management. Breast Cancer Res. 2015, 17, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwan, J.Y.Y.; Psarianos, P.; Bruce, J.P.; Yip, K.W.; Liu, F.-F. The complexity of microRNAs in human cancer. J. Radiat. Res. (Tokyo) 2016, 57, i106–i111. [Google Scholar] [CrossRef]
- Adhami, M.; Haghdoost, A.A.; Sadeghi, B.; Malekpour Afshar, R. Candidate miRNAs in human breast cancer biomarkers: A. systematic review. Breast Cancer 2018, 25, 198–205. [Google Scholar] [CrossRef] [PubMed]
- O’Bryan, S.; Dong, S.; Mathis, J.M.; Alahari, S.K. The roles of oncogenic miRNAs and their therapeutic importance in breast cancer. Eur. J. Cancer 2017, 72, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Piasecka, D.; Braun, M.; Kordek, R.; Sadej, R.; Romanska, H. MicroRNAs in regulation of triple-negative breast cancer progression. J. Cancer Res. Clin. Oncol. 2018, 144, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Muluhngwi, P.; Klinge, C.M. Roles for miRNAs in endocrine resistance in breast cancer. Endocr.-Relat. Cancer 2015, 22, R279–R300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muluhngwi, P.; Klinge, C.M. Identification of miRNAs as biomarkers for acquired endocrine resistance in breast cancer. Mol. Cell Endocrinol. 2017, 456, 76–86. [Google Scholar] [CrossRef]
- Hannafon, B.N.; Ding, W.-Q. miRNAs as Biomarkers for Predicting the Progression of Ductal Carcinoma in situ. Am. J. Pathol. 2018, 188, 542–549. [Google Scholar] [CrossRef] [PubMed]
- Hannafon, B.N.; Ding, W.-Q. Functional Role of microRNAs in the Progression of Breast Ductal Carcinoma in situ. Am. J. Pathol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Guttman, M.; Donaghey, J.; Carey, B.W.; Garber, M.; Grenier, J.K.; Munson, G.; Young, G.; Lucas, A.B.; Ach, R.; Bruhn, L.; et al. lincRNAs act in the circuitry controlling pluripotency and differentiation. Nature 2011, 477, 295–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, R.; Gebhard, C.; Miguel-Escalada, I.; Hoof, I.; Bornholdt, J.; Boyd, M.; Chen, Y.; Zhao, X.; Schmidl, C.; Suzuki, T.; et al. An atlas of active enhancers across human cell types and tissues. Nature 2014, 507, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Peng, W.X.; Koirala, P.; Mo, Y.Y. LncRNA-mediated regulation of cell signaling in cancer. Oncogene 2017, 36, 5661–5667. [Google Scholar] [CrossRef] [PubMed]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef]
- Karijolich, J.; Yu, Y.T. Spliceosomal snRNA modifications and their function. RNA Biol. 2010, 7, 192–204. [Google Scholar] [CrossRef] [Green Version]
- Su, H.; Xu, T.; Ganapathy, S.; Shadfan, M.; Long, M.; Huang, T.H.M.; Thompson, I.; Yuan, Z.M. Elevated snoRNA biogenesis is essential in breast cancer. Oncogene 2014, 33, 1348–1358. [Google Scholar] [CrossRef] [PubMed]
- Jorjani, H.; Kehr, S.; Jedlinski, D.J.; Gumienny, R.; Hertel, J.; Stadler, P.F.; Zavolan, M.; Gruber, A.R. An updated human snoRNAome. Nucleic Acids Res. 2016, 44, 5068–5082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, P.; Ghosh, S.; Wang, B.; Heyns, M.; Graham, K.; Mackey, J.R.; Kovalchuk, O.; Damaraju, S. Profiling of Small Nucleolar RNAs by Next Generation Sequencing: Potential New Players for Breast Cancer Prognosis. PLoS ONE 2016, 11, e0162622. [Google Scholar] [CrossRef]
- Bastide, A.; David, A. The ribosome, (slow) beating heart of cancer (stem) cell. Oncogenesis 2018, 7, 34. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.S.; Ono, M. From snoRNA to miRNA: Dual function regulatory non-coding RNAs. Biochimie 2011, 93, 1987–1992. [Google Scholar] [CrossRef] [PubMed]
- Patterson, D.G.; Roberts, J.T.; King, V.M.; Houserova, D.; Barnhill, E.C.; Crucello, A.; Polska, C.J.; Brantley, L.W.; Kaufman, G.C.; Nguyen, M.; et al. Human snoRNA-93 is processed into a microRNA–like RNA that promotes breast cancer cell invasion. NPJ Breast Cancer 2017, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Tatomer, D.C.; Luo, Z.; Wu, H.; Yang, L.; Chen, L.-L.; Cherry, S.; Wilusz, J.E. The Output of Protein–Coding Genes Shifts to Circular RNAs When the Pre-mRNA Processing Machinery Is Limiting. Mol. Cell 2017, 68, 940–954.e943. [Google Scholar] [CrossRef]
- Qu, S.; Yang, X.; Li, X.; Wang, J.; Gao, Y.; Shang, R.; Sun, W.; Dou, K.; Li, H. Circular RNA: A new star of noncoding RNAs. Cancer Lett. 2015, 365, 141–148. [Google Scholar] [CrossRef]
- Begum, S.; Yiu, A.; Stebbing, J.; Castellano, L. Novel tumour suppressive protein encoded by circular RNA, circ-SHPRH, in glioblastomas. Oncogene 2018, 37, 4055–4057. [Google Scholar] [CrossRef]
- Zhang, M.; Huang, N.; Yang, X.; Luo, J.; Yan, S.; Xiao, F.; Chen, W.; Gao, X.; Zhao, K.; Zhou, H.; et al. A novel protein encoded by the circular form of the SHPRH gene suppresses glioma tumorigenesis. Oncogene 2018, 37, 1805–1814. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, W.W.; Peng, F.; Sun, J.Y.; He, Z.Y.; Wu, S.G. Downregulation of hsa_circ_0011946 suppresses the migration and invasion of the breast cancer cell line MCF-7 by targeting RFC3. Cancer Manag. Res. 2018, 10, 535–544. [Google Scholar] [CrossRef]
- Zhang, H.-d.; Jiang, L.-h.; Hou, J.-c.; Zhong, S.-l.; Zhou, S.-y.; Zhu, L.-p.; Li, J.; Wang, D.-d.; Sun, D.-w.; Ji, Z.-l.; et al. Circular RNA hsa_circ_0052112 promotes cell migration and invasion by acting as sponge for miR-125a-5p in breast cancer. Biomed. Pharmacother. 2018, 107, 1342–1353. [Google Scholar] [CrossRef]
- Zeng, K.; He, B.; Yang, B.B.; Xu, T.; Chen, X.; Xu, M.; Liu, X.; Sun, H.; Pan, Y.; Wang, S. The pro-metastasis effect of circANKS1B in breast cancer. Mol. Cancer 2018, 17, 160. [Google Scholar] [CrossRef]
- Toth, K.F.; Pezic, D.; Stuwe, E.; Webster, A. The piRNA Pathway Guards the Germline Genome Against Transposable Elements. Adv. Exp. Med. Biol. 2016, 886, 51–77. [Google Scholar] [PubMed]
- Ye, Y.; Yin, D.T.; Chen, L.; Zhou, Q.; Shen, R.; He, G.; Yan, Q.; Tong, Z.; Issekutz, A.C.; Shapiro, C.L.; et al. Identification of Piwil2-like (PL2L) proteins that promote tumorigenesis. PLoS ONE 2010, 5, e13406. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, N.; Shi, S.; Liu, S.; Lin, H. The Role of PIWIL4, an Argonaute Family Protein, in Breast Cancer. J. Biol. Chem. 2016, 291, 10646–10658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurozumi, S.; Yamaguchi, Y.; Kurosumi, M.; Ohira, M.; Matsumoto, H.; Horiguchi, J. Recent trends in microRNA research into breast cancer with particular focus on the associations between microRNAs and intrinsic subtypes. J. Hum. Genet. 2016, 62, 15. [Google Scholar] [CrossRef] [PubMed]
- Pinzón, N.; Li, B.; Martinez, L.; Sergeeva, A.; Presumey, J.; Apparailly, F.; Seitz, H. microRNA target prediction programs predict many false positives. Genome Res. 2017, 27, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Lin, H. Posttranscriptional Regulation of Gene Expression by Piwi Proteins and piRNAs. Mol. Cell. 2014, 56, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef]
- Telonis, A.G.; Loher, P.; Jing, Y.; Londin, E.; Rigoutsos, I. Beyond the one-locus-one-miRNA paradigm: MicroRNA isoforms enable deeper insights into breast cancer heterogeneity. Nucleic Acids Res. 2015, 43, 9158–9175. [Google Scholar] [CrossRef]
- Drusco, A.; Croce, C.M. Chapter One-MicroRNAs and Cancer: A Long Story for Short RNAs. In Advances in Cancer Research; Croce, C.M., Fisher, P.B., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 135, pp. 1–24. [Google Scholar]
- Saini, H.K.; Griffiths-Jones, S.; Enright, A.J. Genomic analysis of human microRNA transcripts. Proc. Natl. Acad. Sci. USA 2007, 104, 17719–17724. [Google Scholar] [CrossRef]
- Daugaard, I.; Hansen, T.B. Biogenesis and Function of Ago-Associated RNAs. Trends Genet. 2017, 33, 208–219. [Google Scholar] [CrossRef]
- Sheng, P.; Fields, C.; Aadland, K.; Wei, T.; Kolaczkowski, O.; Gu, T.; Kolaczkowski, B.; Xie, M. Dicer cleaves 5′-extended microRNA precursors originating from RNA polymerase II transcription start sites. Nucleic Acids Res. 2018, 46, 5737–5752. [Google Scholar] [CrossRef] [PubMed]
- Leaderer, D.; Hoffman, A.E.; Zheng, T.; Fu, A.; Weidhaas, J.; Paranjape, T.; Zhu, Y. Genetic and epigenetic association studies suggest a role of microRNA biogenesis gene exportin-5 (XPO5) in breast tumorigenesis. Int. J. Mol. Epidemiol. Genet. 2011, 2, 9–18. [Google Scholar] [PubMed]
- Joyce, B.T.; Zheng, Y.; Zhang, Z.; Liu, L.; Kocherginsky, M.; Murphy, R.; Achenbach, C.J.; Musa, J.; Wehbe, F.; Just, A.; et al. miRNA-Processing Gene Methylation and Cancer Risk. Cancer Epidemiol. Prev. Biomark. 2018, 27, 550. [Google Scholar] [CrossRef] [PubMed]
- Hata, A.; Kashima, R. Dysregulation of microRNA biogenesis machinery in cancer. Crit. Rev. Biochem. Mol. Biol. 2016, 51, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Greco, D.; Michailidou, K.; Milne, R.L.; Muranen, T.A.; Heikkinen, T.; Aaltonen, K.; Dennis, J.; Bolla, M.K.; Liu, J.; et al. MicroRNA Related Polymorphisms and Breast Cancer Risk. PLoS ONE 2014, 9, e109973. [Google Scholar] [CrossRef] [PubMed]
- Hock, J.; Meister, G. The Argonaute protein family. Genome Biol. 2008, 9, 210. [Google Scholar] [CrossRef] [Green Version]
- Pitchiaya, S.; Heinicke, L.A.; Park, J.I.; Cameron, E.L.; Walter, N.G. Resolving Subcellular miRNA Trafficking and Turnover at Single-Molecule Resolution. Cell Rep. 2017, 19, 630–642. [Google Scholar] [CrossRef]
- Selever, J.; Gu, G.; Lewis, M.T.; Beyer, A.; Herynk, M.H.; Covington, K.R.; Tsimelzon, A.; Dontu, G.; Provost, P.; Di Pietro, A.; et al. Dicer-Mediated Upregulation of BCRP Confers Tamoxifen Resistance in Human Breast Cancer Cells. Clin. Cancer Res. 2011, 17, 6510–6521. [Google Scholar] [CrossRef] [Green Version]
- Conger, A.K.; Martin, E.C.; Yan, T.J.; Rhodes, L.V.; Hoang, V.T.; La, J.; Anbalagan, M.; Burks, H.E.; Rowan, B.G.; Nephew, K.P.; et al. Argonaute 2 Expression Correlates with a Luminal B Breast Cancer Subtype and Induces Estrogen Receptor Alpha Isoform Variation. Non-Codin. RNA 2016, 2, 8. [Google Scholar] [CrossRef]
- Wang, Z.-Y.; Yin, L. Estrogen receptor alpha-36 (ER-α36): A new player in human breast cancer. Mol. Cell Endocrinol. 2015, 418, 193–206. [Google Scholar] [CrossRef] [Green Version]
- Vendramin, R.; Marine, J.C.; Leucci, E. Non-coding RNAs: The dark side of nuclear-mitochondrial communication. EMBO J. 2017, 36, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Feeley, K.P.; Bray, A.W.; Westbrook, D.G.; Johnson, L.W.; Kesterson, R.A.; Ballinger, S.W.; Welch, D.R. Mitochondrial Genetics Regulate Breast Cancer Tumorigenicity and Metastatic Potential. Cancer Res. 2015, 75, 4429–4436. [Google Scholar] [CrossRef] [PubMed]
- Brinker, A.E.; Vivian, C.J.; Koestler, D.C.; Tsue, T.T.; Jensen, R.A.; Welch, D.R. Mitochondrial Haplotype Alters Mammary Cancer Tumorigenicity and Metastasis in an Oncogenic Driver-Dependent Manner. Cancer Res. 2017, 77, 6941–6949. [Google Scholar] [CrossRef] [PubMed]
- Mattie, M.D.; Benz, C.C.; Bowers, J.; Sensinger, K.; Wong, L.; Scott, G.K.; Fedele, V.; Ginzinger, D.; Getts, R.; Haqq, C. Optimized high-throughput microRNA expression profiling provides novel biomarker assessment of clinical prostate and breast cancer biopsies. Mol. Cancer 2006, 5, 24. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Liu, M.; Stribinskis, V.; Klinge, C.M.; Ramos, K.S.; Colburn, N.H.; Li, Y. MicroRNA-21 Promotes Cell Transformation by Targeting the Programmed Cell Death 4 Gene. Oncogene 2008, 27, 4373–4379. [Google Scholar] [CrossRef]
- Yan, L.-X.; Huang, X.-F.; Shao, Q.; Huang, M.-Y.; Deng, L.; Wu, Q.-L.; Zeng, Y.-X.; Shao, J.-Y. MicroRNA miR-21 overexpression in human breast cancer is associated with advanced clinical stage, lymph node metastasis and patient poor prognosis. RNA 2008, 14, 2348–2360. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Wu, H.; Wu, F.; Nie, D.; Sheng, S.; Mo, Y.Y. MicroRNA-21 targets tumor suppressor genes in invasion and metastasis. Cell Res. 2008, 18, 350–359. [Google Scholar] [CrossRef] [Green Version]
- Gong, C.; Yao, Y.; Wang, Y.; Liu, B.; Wu, W.; Chen, J.; Su, F.; Yao, H.; Song, E. Up-regulation of miR-21 mediates resistance to trastuzumab therapy for breast cancer. J. Biol. Chem. 2011, 286, 19127–19137. [Google Scholar] [CrossRef]
- Rask, L.; Balslev, E.V.A.; JØRgensen, S.; Eriksen, J.; Flyger, H.; MØLler, S.; HØGdall, E.; Litman, T.; Schnack Nielsen, B. High expression of miR-21 in tumor stroma correlates with increased cancer cell proliferation in human breast cancer. APMIS 2011, 119, 663–673. [Google Scholar] [CrossRef]
- Yan, L.X.; Wu, Q.N.; Zhang, Y.; Li, Y.Y.; Liao, D.Z.; Hou, J.H.; Fu, J.; Zeng, M.S.; Yun, J.P.; Wu, Q.L.; et al. Knockdown of miR-21 in human breast cancer cell lines inhibits proliferation, in vitro migration and in vivo tumor growth. Breast Cancer Res. 2011, 13, R2. [Google Scholar] [CrossRef]
- Han, M.; Wang, Y.; Liu, M.; Bi, X.; Bao, J.; Zeng, N.; Zhu, Z.; Mo, Z.; Wu, C.; Chen, X. MiR-21 regulates EMT phenotype and HIF-1α expression in third–sphereforming breast cancer stem cell-like cells. Cancer Sci. 2012, 103, 1058–1064. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liang, Z.; Xu, L.; Zou, F. MicroRNA-21: A ubiquitously expressed pro-survival factor in cancer and other diseases. Mol. Cell Biochem. 2012, 360, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Petrović, N.; Mandušić, V.; Stanojević, B.; Lukić, S.; Todorović, L.; Roganović, J.; Dimitrijević, B. The difference in miR-21 expression levels between invasive and non-invasive breast cancers emphasizes its role in breast cancer invasion. Med. Oncol. 2014, 31, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, N. miR-21 Might be Involved in Breast Cancer Promotion and Invasion Rather than in Initial Events of Breast Cancer Development. Mol. Diagn. Ther. 2016, 20, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or Tumor Suppressor? The Duplicity of MicroRNAs in Cancer. Cancer Res. 2016, 76, 3666. [Google Scholar] [CrossRef]
- Klinge, C.M. miRNAs and estrogen action. Trends Endocrinol. Metab. 2012, 23, 223–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Xu, J.; Shi, Y.; Sun, Q.; Zhang, Q.; Guan, X. The novel role of miRNAs for tamoxifen resistance in human breast cancer. Cell Mol. Life Sci. 2015, 72, 2575–2584. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.-D.; Tseng, Y.-T.; Shrestha, S.; Lin, Y.-L.; Khaleel, A.; Chou, C.-H.; Chu, C.-F.; Huang, H.-Y.; Lin, C.-M.; Ho, S.-Y.; et al. miRTarBase update 2014: An information resource for experimentally validated miRNA-target interactions. Nucleic Acids Res. 2014, 42, D78–D85. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, P.; Chen, Z.; Liu, M.; Li, X.; Tang, H. MicroRNA-7 downregulates XIAP expression to suppress cell growth and promote apoptosis in cervical cancer cells. FEBS Lett. 2013, 587, 2247–2253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhang, X.; Hu, S.; Zheng, M.; Zhang, J.; Zhao, J.; Zhang, X.; Yan, B.; Jia, L.; Zhao, J.; et al. Identification of miRNA-7 by genome-wide analysis as a critical sensitizer for TRAIL-induced apoptosis in glioblastoma cells. Nucleic Acids Res. 2017, 45, 5930–5944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannafon, B.; Sebastiani, P.; de las Morenas, A.; Lu, J.; Rosenberg, C. Expression of microRNAs and their gene targets are dysregulated in pre-invasive breast cancer. Breast Cancer Res. 2011, 13, R24. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.-X.; Bradbury, R.; Flamini, V.; Wu, B.; Jordan, N.; Jiang, W.G. MicroRNA-7 suppresses the homing and migration potential of human endothelial cells to highly metastatic human breast cancer cells. Br. J. Cancer 2017, 117, 89. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, Y.; Wu, H.; Li, Y.; Zhang, Y.; Liu, M.; Li, X.; Tang, H. miR-10a suppresses colorectal cancer metastasis by modulating the epithelial-to-mesenchymal transition and anoikis. Cell Death Dis. 2017, 8, e2739. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.; Meng, X.; Liang, H.; Zhang, H.; Liu, X.; Li, L.; Li, W.; Sun, W.; Zhang, H.; Zen, K.; et al. miR-10a inhibits cell proliferation and promotes cell apoptosis by targeting BCL6 in diffuse large B-cell lymphoma. Protein Cell 2016, 7, 899–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, M.-J.; Wu, F.-X.; Li, P.; Liu, M.; Li, X.; Tang, H. MicroRNA-10a targets CHL1 and promotes cell growth, migration and invasion in human cervical cancer cells. Cancer Lett. 2012, 324, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Foley, N.H.; Bray, I.; Watters, K.M.; Das, S.; Bryan, K.; Bernas, T.; Prehn, J.H.M.; Stallings, R.L. MicroRNAs 10a and 10b are potent inducers of neuroblastoma cell differentiation through targeting of nuclear receptor corepressor 2. Cell Death Differ. 2011, 18, 1089–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arai, T.; Okato, A.; Kojima, S.; Idichi, T.; Koshizuka, K.; Kurozumi, A.; Kato, M.; Yamazaki, K.; Ishida, Y.; Naya, Y.; et al. Regulation of spindle and kinetochore-associated protein 1 by antitumor miR-10a-5p in renal cell carcinoma. Cancer Sci. 2017, 108, 2088–2101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, J.; Duncavage, E.; Tamburrino, A.; Salerno, P.; Xi, L.; Raffeld, M.; Moley, J.; Chernock, R.D. Overexpression of miR-10a and miR-375 and downregulation of YAP1 in medullary thyroid carcinoma. Exp. Mol. Pathol. 2013, 95, 62–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, Z.; Yao, B.; Dou, C.; Xu, M.; Xue, Y.; Ding, L.; Jia, Y.; Zhang, H.; Li, Q.; et al. Long non–coding RNA TUSC7 acts a molecular sponge for miR-10a and suppresses EMT in hepatocellular carcinoma. Tumour Biol. 2016, 37, 11429–11441. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, N.; Liu, B.; Huang, Y.; Zeng, H.; Yang, Z.; He, Z.; Guo, H. Long noncoding RNA RP11-838N2.4 enhances the cytotoxic effects of temozolomide by inhibiting the functions of miR-10a in glioblastoma cell lines. Oncotarget 2016, 7, 43835–43851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, K.; Lou, T. MicroRNA-10a suppresses breast cancer progression via PI3K/Akt/mTOR pathway. Oncol. Lett. 2017, 14, 5994–6000. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, R.; Achinger-Kawecka, J.; Winter, S.; Fritz, P.; Lo, W.-Y.; Schroth, W.; Brauch, H. Increased expression of miR-126 and miR-10a predict prolonged relapse-free time of primary oestrogen receptor-positive breast cancer following tamoxifen treatment. Eur. J. Cancer 2013, 49, 3598–3608. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.; Go, E.J.; Kim, A. Recurrence prediction using microRNA expression in hormone receptor positive breast cancer during tamoxifen treatment. Biomarkers 2018, 23, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Castellano, L.; Giamas, G.; Jacob, J.; Coombes, R.C.; Lucchesi, W.; Thiruchelvam, P.; Barton, G.; Jiao, L.R.; Wait, R.; Waxman, J.; et al. The estrogen receptor-alpha induced microRNA signature regulates itself and its transcriptional response. Proc. Natl. Acad. Sci. USA 2009, 106, 15732–15737. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhang, H.; Liu, Y.; Su, P.; Zhang, J.; Wang, X.; Sun, M.; Chen, B.; Zhao, W.; Wang, L.; et al. SREBP1, targeted by miR-18a-5p, modulates epithelial–mesenchymal transition in breast cancer via forming a co-repressor complex with Snail and HDAC1/2. Cell Death Differ. 2018. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, Y.; Liu, A.; Tang, X. Long non-coding RNA UCA1 enhances tamoxifen resistance in breast cancer cells through a miR-18a-HIF1alpha feedback regulatory loop. Tumor Biol. 2016, 37, 14733–14743. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, N.; Toyama, T.; Takahashi, S.; Sugiura, H.; Endo, Y.; Iwasa, M.; Fujii, Y.; Yamashita, H. Distinct expressions of microRNAs that directly target estrogen receptor α in human breast cancer. Breast Cancer Res. Treat. 2011, 130, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Wickramasinghe, N.; Manavalan, T.; Dougherty, S.; Riggs, K.; Li, Y.; Klinge, C. Estradiol downregulates miR-21 expression and increases miR-21 target gene expression in MCF-7 breast cancer cells. Nucleic Acids Res. 2009, 37, 2584–2595. [Google Scholar] [CrossRef] [Green Version]
- Queiros, A.M.; Eschen, C.; Fliegner, D.; Kararigas, G.; Dworatzek, E.; Westphal, C.; Sanchez Ruderisch, H.; Regitz-Zagrosek, V. Sex- and estrogen-dependent regulation of a miRNA network in the healthy and hypertrophied heart. Int. J. Cardiol. 2013, 169, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Kanwal, R.; Plaga, A.R.; Liu, X.; Shukla, G.C.; Gupta, S. MicroRNAs in prostate cancer: Functional role as biomarkers. Cancer Lett. 2017, 407, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, X.; Yang, J.; Zhen, J.; Zhang, D. Serum microRNA-21 as a potential diagnostic biomarker for breast cancer: A systematic review and meta-analysis. Clin. Exp. Med. 2016, 16, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zheng, J.; Zhang, Y.; Yang, L.; Wang, J.; Ni, J.; Cui, D.; Yu, C.; Cai, Z. Tumor-specific Expression of MicroRNA-26a Suppresses Human Hepatocellular Carcinoma Growth via Cyclin-dependent and-independent Pathways. Mol. Ther. 2011, 19, 1521–1528. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Ding, K.; Li, R.; Zhang, W.; Li, G.; Kong, X.; Qian, P.; Lobie, P.E.; Zhu, T. Identification of miR-26 as a key mediator of estrogen stimulated cell proliferation by targeting CHD1, GREB1 and KPNA2. Breast Cancer Res. 2014, 16, R40. [Google Scholar] [CrossRef] [PubMed]
- Jansen, M.P.; Reijm, E.A.; Sieuwerts, A.M.; Ruigrok-Ritstier, K.; Look, M.P.; Rodriguez-Gonzalez, F.G.; Heine, A.A.; Martens, J.W.; Sleijfer, S.; Foekens, J.A.; et al. High miR-26a and low CDC2 levels associate with decreased EZH2 expression and with favorable outcome on tamoxifen in metastatic breast cancer. Breast Cancer Res. Treat. 2012, 133, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Tanno, T.; Zhang, P.; Lazarski, C.A.; Liu, Y.; Zheng, P. An aptamer–based targeted delivery of miR-26a protects mice against chemotherapy toxicity while suppressing tumor growth. Blood Adv. 2017, 1, 1107–1119. [Google Scholar] [CrossRef] [PubMed]
- Muluhngwi, P.; Krishna, A.; Vittitow, S.L.; Napier, J.T.; Richardson, K.M.; Ellis, M.; Mott, J.L.; Klinge, C.M. Tamoxifen differentially regulates miR-29b-1 and miR-29a expression depending on endocrine–sensitivity in breast cancer cells. Cancer Lett. 2017, 388, 230–238. [Google Scholar] [CrossRef]
- Ding, D.; Li, C.; Zhao, T.; Li, D.; Yang, L.; Zhang, B. LncRNA H19/miR-29b-3p/PGRN Axis Promoted Epithelial-Mesenchymal Transition of Colorectal Cancer Cells by Acting on Wnt Signaling. Mol. Cells 2018, 41, 423–435. [Google Scholar]
- Zhang, X.; Cheng, L.; Xu, L.; Zhang, Y.; Yang, Y.; Fu, Q.; Mi, W.; Li, H. The lncRNA H19 mediates the protective eEffect of hypoxia postconditioning against hypoxia-Rreoxygenation injury to senescent cardiomyocytes by targeting microRNA-29b-3p. Shock 2018. [Google Scholar] [CrossRef]
- Zhang, H.; Li, H.; Ge, A.; Guo, E.; Liu, S.; Zhang, L. Long non-coding RNA TUG1 inhibits apoptosis and inflammatory response in LPS-treated H9c2 cells by down-regulation of miR–29b. Biomed. Pharmacother. 2018, 101, 663–669. [Google Scholar] [CrossRef]
- Du, P.; Zhao, H.; Peng, R.; Liu, Q.; Yuan, J.; Peng, G.; Liao, Y. LncRNA-XIST interacts with miR-29c to modulate the chemoresistance of glioma cell to TMZ through DNA mismatch repair pathway. Biosci. Rep. 2017, 37, BSR20170696. [Google Scholar] [CrossRef]
- Han, Q.; Li, L.; Liang, H.; Li, Y.; Xie, J.; Wang, Z. Downregulation of lncRNA X Inactive Specific Transcript (XIST) Suppresses Cell Proliferation and Enhances Radiosensitivity by Upregulating mir-29c in Nasopharyngeal Carcinoma Cells. Med. Sci. Monit. 2017, 23, 4798–4807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alipoor, F.J.; Asadi, M.H.; Torkzadeh-Mahani, M. MIAT lncRNA is overexpressed in breast cancer and its inhibition triggers senescence and G1 arrest in MCF7 cell line. J. Cell Biochem. 2018, 119, 6470–6481. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Zhang, G.; Wu, J.H.; Jiang, C.P. Diverse roles of miR-29 in cancer (review). Oncol. Rep. 2014, 31, 1509–1516. [Google Scholar] [CrossRef] [PubMed]
- Bhat-Nakshatri, P.; Wang, G.; Collins, N.R.; Thomson, M.J.; Geistlinger, T.R.; Carroll, J.S.; Brown, M.; Hammond, S.; Srour, E.F.; Liu, Y.; et al. Estradiol-regulated microRNAs control estradiol response in breast cancer cells. Nucleic Acids Res. 2009, 37, 4850–4861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, X.H.; Chen, Z.Y.; Luo, H.W.; Liu, Z.Z.; Liang, Y.K.; Chen, G.X.; Jiang, F.N.; Zhong, W.D. BCL9, a coactivator for Wnt/beta-catenin transcription, is targeted by miR-30c and is associated with prostate cancer progression. Oncol. Lett. 2016, 11, 2001–2008. [Google Scholar] [CrossRef] [PubMed]
- Ni, L.Y.; Zhao, J.D.; Lu, Y.H.; Li, W.; Li, B.L.; Wang, X.C.; Meng, Q.G. MicroRNA-30c suppressed giant-cell tumor of bone cell metastasis and growth via targeting HOXA1. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 4819–4827. [Google Scholar] [PubMed]
- Huang, Y.Q.; Ling, X.H.; Yuan, R.Q.; Chen, Z.Y.; Yang, S.B.; Huang, H.X.; Zhong, W.D.; Qiu, S.P. miR-30c suppresses prostate cancer survival by targeting the ASF/SF2 splicing factor oncoprotein. Mol. Med. Rep. 2017, 16, 2431–2438. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, X.; Wang, Y.; Peng, R.; Lin, Z.; Wang, Y.; Hu, B.; Wang, J.; Shi, G. Low expression of microRNA-30c promotes prostate cancer cells invasion involved in downregulation of KRAS protein. Oncol. Lett. 2017, 14, 363–368. [Google Scholar] [CrossRef] [Green Version]
- Su, W.; Hong, L.; Xu, X.; Huang, S.; Herpai, D.; Li, L.; Xu, Y.; Truong, L.; Hu, W.-Y.; Wu, X.; et al. miR-30 disrupts senescence and promotes cancer by targeting both p16INK4A and DNA damage pathways. Oncogene 2018, 37, 5618–5632. [Google Scholar] [CrossRef]
- Liang, T.; Zhou, B.; Shi, L.; Wang, H.; Chu, Q.; Xu, F.; Li, Y.; Chen, R.; Shen, C.; Schinckel, AP. lncRNA AK017368 promotes proliferation and suppresses differentiation of myoblasts in skeletal muscle development by attenuating the function of miR-30c. FASEB J. 2017, 32, 377–389. [Google Scholar] [CrossRef]
- Liu, X.; Li, M.; Peng, Y.; Hu, X.; Xu, J.; Zhu, S.; Yu, Z.; Han, S. miR-30c regulates proliferation, apoptosis and differentiation via the Shh signaling pathway in P19 cells. Exp. Mol. Med. 2016, 48, e248. [Google Scholar] [CrossRef] [PubMed]
- Croset, M.; Pantano, F.; Kan, C.W.S.; Bonnelye, E.; Descotes, F.; Alix-Panabières, C.; Lecellier, C.-H.; Bachelier, R.; Allioli, N.; Hong, S.-S.; et al. MicroRNA-30 family members inhibit breast cancer invasion, osteomimicry, and bone destruction by directly targeting multiple bone metastasis-associated genes. Cancer Res. 2018, 78, 5259–5273. [Google Scholar] [CrossRef] [PubMed]
- Razaviyan, J.; Hadavi, R.; Tavakoli, R.; Kamani, F.; Paknejad, M.; Mohammadi-Yeganeh, S. Expression of miRNAs Targeting mTOR and S6K1 Genes of mTOR Signaling Pathway Including miR-96, miR-557, and miR-3182 in Triple-Negative Breast Cancer. Appl. Biochem. Biotechnol. 2018, 186, 1074–1089. [Google Scholar] [CrossRef] [PubMed]
- Cai, T.; Long, J.; Wang, H.; Liu, W.; Zhang, Y. Identification and characterization of miR-96, a potential biomarker of NSCLC, through bioinformatic analysis. Oncol. Rep. 2017, 38, 1213–1223. [Google Scholar] [CrossRef] [PubMed]
- Xiong, D.D.; Lv, J.; Wei, K.L.; Feng, Z.B.; Chen, J.T.; Liu, K.C.; Chen, G.; Luo, D.Z. A nine–miRNA signature as a potential diagnostic marker for breast carcinoma: An integrated study of 1110 cases. Oncol. Rep. 2017, 37, 3297–3304. [Google Scholar] [CrossRef]
- Scott, G.K.; Goga, A.; Bhaumik, D.; Berger, C.E.; Sullivan, C.S.; Benz, C.C. Coordinate Suppression of ERBB2 and ERBB3 by Enforced Expression of Micro-RNA miR-125a or miR-125b. J. Biol. Chem. 2007, 282, 1479–1486. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, L.-X.; Wu, Q.-N.; Du, Z.-M.; Chen, J.; Liao, D.-Z.; Huang, M.-Y.; Hou, J.-H.; Wu, Q.-L.; Zeng, M.-S.; et al. miR-125b Is Methylated and Functions as a Tumor Suppressor by Regulating the ETS1 Proto–oncogene in Human Invasive Breast Cancer. Cancer Res. 2011, 71, 3552–3562. [Google Scholar] [CrossRef] [PubMed]
- Vilquin, P.; Donini, C.F.; Villedieu, M.; Grisard, E.; Corbo, L.; Bachelot, T.; Vendrell, J.A.; Cohen, P.A. MicroRNA-125b upregulation confers aromatase inhibitor resistance and is a novel marker of poor prognosis in breast cancer. Breast Cancer Res. 2015, 17, 13. [Google Scholar] [CrossRef] [PubMed]
- Matamala, N.; Vargas, M.T.; González-Cámpora, R.; Miñambres, R.; Arias, J.I.; Menéndez, P.; Andrés-León, E.; Gómez-López, G.; Yanowsky, K.; Calvete-Candenas, J.; et al. Tumor MicroRNA Expression Profiling Identifies Circulating MicroRNAs for Early Breast Cancer Detection. Clin. Chem. 2015, 61, 1098. [Google Scholar] [CrossRef] [PubMed]
- Shivapurkar, N.; Vietsch, E.E.; Carney, E.; Isaacs, C.; Wellstein, A. Circulating microRNAs in patients with hormone receptor-positive, metastatic breast cancer treated with dovitinib. Clin. Trans. Med. 2017, 6, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoppe, R.; Fan, P.; Buttner, F.; Winter, S.; Tyagi, A.K.; Cunliffe, H.; Jordan, V.C.; Brauch, H. Profiles of miRNAs matched to biology in aromatase inhibitor resistant breast cancer. Oncotarget 2016, 7, 71235–71254. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.-Y.; Zhang, F.; Sun, C.-C.; Li, S.-J.; Li, G.; Gong, F.-Y.; Bo, T.; He, J.; Hua, R.-X.; Hu, W.-D.; et al. miR-134: A Human Cancer Suppressor? Mol. Ther. Nucleic Acids 2017, 6, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Bergamaschi, A.; Madak-Erdogan, Z.; Kim, Y.J.; Choi, Y.L.; Lu, H.; Katzenellenbogen, B.S. The forkhead transcription factor FOXM1 promotes endocrine resistance and invasiveness in estrogen receptor-positive breast cancer by expansion of stem-like cancer cells. Breast Cancer Res. 2014, 16, 436. [Google Scholar] [CrossRef] [PubMed]
- Hamam, R.; Ali, A.M.; Alsaleh, K.A.; Kassem, M.; Alfayez, M.; Aldahmash, A.; Alajez, N.M. microRNA expression profiling on individual breast cancer patients identifies novel panel of circulating microRNA for early detection. Sci. Rep. 2016, 6, 25997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benetatos, L.; Hatzimichael, E.; Londin, E.; Vartholomatos, G.; Loher, P.; Rigoutsos, I.; Briasoulis, E. The microRNAs within the DLK1-DIO3 genomic region: Involvement in disease pathogenesis. Cell Mol. Life Sci. 2013, 70, 795–814. [Google Scholar] [CrossRef] [PubMed]
- Millour, J.; Constantinidou, D.; Stavropoulou, A.V.; Wilson, M.S.C.; Myatt, S.S.; Kwok, J.M.M.; Sivanandan, K.; Coombes, R.C.; Medema, R.H.; Hartman, J.; et al. FOXM1 is a transcriptional target of ER[alpha] and has a critical role in breast cancer endocrine sensitivity and resistance. Oncogene 2010, 29, 2983–2995. [Google Scholar] [CrossRef] [PubMed]
- Dinami, R.; Ercolani, C.; Petti, E.; Piazza, S.; Ciani, Y.; Sestito, R.; Sacconi, A.; Biagioni, F.; le Sage, C.; Agami, R.; et al. miR-155 drives telomere fragility in human breast cancer by targeting TRF1. Cancer Res. 2014, 74, 4145–4156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.M.; Zhao, J.; Deng, H.Y. miR-155 promotes proliferation of human breast cancer MCF-7 cells through targeting tumor protein 53–induced nuclear protein 1. J. Biomed. Sci. 2013, 20, 79. [Google Scholar] [CrossRef]
- Brown, C.Y.; Dayan, S.; Wong, S.W.; Kaczmarek, A.; Hope, C.M.; Pederson, S.M.; Arnet, V.; Goodall, G.J.; Russell, D.; Sadlon, T.J. FOXP3 and miR-155 cooperate to control the invasive potential of human breast cancer cells by down regulating ZEB2 independently of ZEB1. Oncotarget 2018, 9, 27708–27727. [Google Scholar] [CrossRef]
- Zhou, X.; Ye, F.; Yin, C.; Zhuang, Y.; Yue, G.; Zhang, G. The Interaction Between miR-141 and lncRNA-H19 in Regulating Cell Proliferation and Migration in Gastric Cancer. Cell Physiol. Biochem. 2015, 36, 1440–1452. [Google Scholar] [CrossRef]
- Chen, L.; Wang, W.; Cao, L.; Li, Z.; Wang, X. Long Non-Coding RNA CCAT1 Acts as a Competing Endogenous RNA to Regulate Cell Growth and Differentiation in Acute Myeloid Leukemia. Mol. Cells 2016, 39, 330–336. [Google Scholar] [Green Version]
- Cao, S.; Wang, Y.; Li, J.; Lv, M.; Niu, H.; Tian, Y. Tumor-suppressive function of long noncoding RNA MALAT1 in glioma cells by suppressing miR-155 expression and activating FBXW7 function. Am. J. Cancer Res. 2016, 6, 2561–2574. [Google Scholar] [PubMed]
- Luan, T.; Zhang, X.; Wang, S.; Song, Y.; Zhou, S.; Lin, J.; An, W.; Yuan, W.; Yang, Y.; Cai, H.; et al. Long non–coding RNA MIAT promotes breast cancer progression and functions as ceRNA to regulate DUSP7 expression by sponging miR-155-5p. Oncotarget 2017, 8, 76153–76164. [Google Scholar] [CrossRef] [PubMed]
- Dou, L.; Lin, H.; Wang, K.; Zhu, G.; Zou, X.; Chang, E.; Zhu, Y. Long non-coding RNA CCAT1 modulates neuropathic pain progression through sponging miR-155. Oncotarget 2017, 8, 89949–89957. [Google Scholar] [CrossRef] [PubMed]
- Eichelser, C.; Flesch-Janys, D.; Chang-Claude, J.; Pantel, K.; Schwarzenbach, H. Deregulated Serum Concentrations of Circulating Cell–Free MicroRNAs miR-17, miR-34a, miR-155, and miR-373 in Human Breast Cancer Development and Progression. Clin. Chem. 2013, 59, 1489–1496. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Shi, W.; Wu, C.; Ju, J.; Jiang, J. miR-181b as a key regulator of the oncogenic process and its clinical implications in cancer (Review). Biomed. Rep. 2014, 2, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Cui, B.; Li, B.; Liu, Q.; Cui, Y. lncRNA CCAT1 Promotes Glioma Tumorigenesis by Sponging miR-181b. J. Cell Biochem. 2017, 118, 4548–4557. [Google Scholar] [CrossRef]
- Sochor, M.; Basova, P.; Pesta, M.; Dusilkova, N.; Bartos, J.; Burda, P.; Pospisil, V.; Stopka, T. Oncogenic microRNAs: MiR-155, miR-19a, miR-181b, and miR-24 enable monitoring of early breast cancer in serum. BMC Cancer 2014, 14, 448. [Google Scholar] [CrossRef]
- Yoo, J.-O.; Kwak, S.-Y.; An, H.-J.; Bae, I.-H.; Park, M.-J.; Han, Y.-H. miR-181b-3p promotes epithelial–mesenchymal transition in breast cancer cells through Snail stabilization by directly targeting YWHAG. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 1601–1611. [Google Scholar] [CrossRef]
- Wang, R.; Tian, S.; Wang, H.B.; Chu, D.P.; Cao, J.L.; Xia, H.F.; Ma, X. MiR-185 is involved in human breast carcinogenesis by targeting Vegfa. FEBS Lett. 2014, 588, 4438–4447. [Google Scholar] [CrossRef]
- Tang, H.; Liu, P.; Yang, L.; Xie, X.; Ye, F.; Wu, M.; Liu, X.; Chen, B.; Zhang, L.; Xie, X. miR-185 Suppresses Tumor Proliferation by Directly Targeting E2F6 and DNMT1 and Indirectly Upregulating BRCA1 in Triple–Negative Breast Cancer. Mol. Cancer Ther. 2014, 13, 3185. [Google Scholar] [CrossRef] [PubMed]
- Zou, Q.; Wu, H.; Fu, F.; Yi, W.; Pei, L.; Zhou, M. RKIP suppresses the proliferation and metastasis of breast cancer cell lines through up–regulation of miR-185 targeting HMGA2. Arch. Biochem. Biophys. 2016, 610, 25–32. [Google Scholar] [CrossRef]
- Imam, J.S.; Buddavarapu, K.; Lee-Chang, J.S.; Ganapathy, S.; Camosy, C.; Chen, Y.; Rao, M.K. MicroRNA-185 suppresses tumor growth and progression by targeting the Six1 oncogene in human cancers. Oncogene 2010, 29, 4971–4979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulin, J.-A.; Tommasi, S.; Elliot, D.; Hu, D.G.; Lewis, B.C.; Mangoni, A.A. MiR-193b regulates breast cancer cell migration and vasculogenic mimicry by targeting dimethylarginine dimethylaminohydrolase 1. Sci. Rep. 2017, 7, 13996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, F.; Hosany, S.; Zhong, S.; Jiang, Y.; Zhang, F.; Lin, L.; Wang, X.; Gao, S.; Hu, X. MicroRNA-193a inhibits breast cancer proliferation and metastasis by downregulating WT1. PLoS ONE 2017, 12, e0185565. [Google Scholar] [CrossRef]
- Tsai, K.-W.; Leung, C.-M.; Lo, Y.-H.; Chen, T.-W.; Chan, W.-C.; Yu, S.-Y.; Tu, Y.-T.; Lam, H.-C.; Li, S.-C.; Ger, L.-P.; et al. Arm Selection Preference of MicroRNA–193a Varies in Breast Cancer. Sci. Rep. 2016, 6, 28176. [Google Scholar] [CrossRef] [PubMed]
- Dvinge, H.; Git, A.; Graf, S.; Salmon-Divon, M.; Curtis, C.; Sottoriva, A.; Zhao, Y.; Hirst, M.; Armisen, J.; Miska, E.A.; et al. The shaping and functional consequences of the microRNA landscape in breast cancer. Nature 2013, 497, 378–382. [Google Scholar] [CrossRef]
- Chen, Z.; Li, Y.; Zhang, H.; Huang, P.; Luthra, R. Hypoxia–regulated microRNA-210 modulates mitochondrial function and decreases ISCU and COX10 expression. Oncogene 2010, 29, 4362–4368. [Google Scholar] [CrossRef]
- Fasanaro, P.; D’Alessandra, Y.; Di Stefano, V.; Melchionna, R.; Romani, S.; Pompilio, G.; Capogrossi, M.C.; Martelli, F. MicroRNA-210 Modulates Endothelial Cell Response to Hypoxia and Inhibits the Receptor Tyrosine Kinase Ligand Ephrin–A3. J. Biol. Chem. 2008, 283, 15878–15883. [Google Scholar] [CrossRef]
- Rothé, F.; Ignatiadis, M.; Chaboteaux, C.; Haibe-Kains, B.; Kheddoumi, N.; Majjaj, S.; Badran, B.; Fayyad-Kazan, H.; Desmedt, C.; Harris, A.L.; et al. Global MicroRNA Expression Profiling Identifies MiR-210 Associated with Tumor Proliferation, Invasion and Poor Clinical Outcome in Breast Cancer. PLoS ONE 2011, 6, e20980. [Google Scholar] [CrossRef]
- Camps, C.; Buffa, F.M.; Colella, S.; Moore, J.; Sotiriou, C.; Sheldon, H.; Harris, A.L.; Gleadle, J.M.; Ragoussis, J. has-miR-210 Is Induced by Hypoxia and Is an Independent Prognostic Factor in Breast Cancer. Clin. Cancer Res. 2008, 14, 1340–1348. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.-J.; Lin, J.; Yang, H.; Kong, W.; He, L.; Ma, X.; Coppola, D.; Cheng, J.Q. MicroRNA-221/222 negatively regulates ERalpha and associates with tamoxifen resistance in breast cancer. J. Biol. Chem. 2008, 283, 31079–31086. [Google Scholar] [CrossRef] [PubMed]
- Di Leva, G.; Gasparini, P.; Piovan, C.; Ngankeu, A.; Garofalo, M.; Taccioli, C.; Iorio, M.V.; Li, M.; Volinia, S.; Alder, H.; et al. MicroRNA Cluster 221–222 and Estrogen Receptor {alpha} Interactions in Breast Cancer. J. Natl. Cancer Inst. 2010, 102, 706–721. [Google Scholar] [CrossRef] [PubMed]
- Di Leva, G.; Piovan, C.; Gasparini, P.; Ngankeu, A.; Taccioli, C.; Briskin, D.; Cheung, D.G.; Bolon, B.; Anderlucci, L.; Alder, H.; et al. Estrogen Mediated–Activation of miR-191/425 Cluster Modulates Tumorigenicity of Breast Cancer Cells Depending on Estrogen Receptor Status. PLoS Genet. 2013, 9, e1003311. [Google Scholar] [CrossRef]
- Rao, X.; Di Leva, G.; Li, M.; Fang, F.; Devlin, C.; Hartman-Frey, C.; Burow, M.E.; Ivan, M.; Croce, C.M.; Nephew, K.P. MicroRNA-221/222 confers breast cancer fulvestrant resistance by regulating multiple signaling pathways. Oncogene 2011, 30, 1082–1097. [Google Scholar] [CrossRef]
- Miller, T.E.; Ghoshal, K.; Ramaswamy, B.; Roy, S.; Datta, J.; Shapiro, C.L.; Jacob, S.; Majumder, S. MicroRNA-221/222 confers tamoxifen resistance in breast cancer by targeting p27(Kip1). J. Biol. Chem. 2008, 283, 29897–29903. [Google Scholar] [CrossRef]
- Lu, Y.; Roy, S.; Nuovo, G.; Ramaswamy, B.; Miller, T.; Shapiro, C.; Jacob, S.T.; Majumder, S. Anti-microRNA-222 (Anti-miR-222) and –181B Suppress Growth of Tamoxifen–resistant Xenografts in Mouse by Targeting TIMP3 Protein and Modulating Mitogenic Signal. J. Biol. Chem. 2011, 286, 42292–42302. [Google Scholar] [CrossRef]
- Dentelli, P.; Rosso, A.; Orso, F.; Olgasi, C.; Taverna, D.; Brizzi, M.F. microRNA-222 controls neovascularization by regulating signal transducer and activator of transcription 5A expression. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1562–1568. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, X.; Zhang, L.; Hong, W.; Zeng, K. MicroRNA-222 regulates the viability of fibroblasts in hypertrophic scars via matrix metalloproteinase 1. Exp. Ther. Med. 2018, 15, 1803–1808. [Google Scholar] [CrossRef]
- Li, Y.; Liang, C.; Ma, H.; Zhao, Q.; Lu, Y.; Xiang, Z.; Li, L.; Qin, J.; Chen, Y.; Cho, W.C.; et al. miR-221/222 promotes S–phase entry and cellular migration in control of basal–like breast cancer. Molecules 2014, 19, 7122–7137. [Google Scholar] [CrossRef]
- Wu, Q.; Lu, Z.; Li, H.; Lu, J.; Guo, L.; Ge, Q. Next–generation sequencing of microRNAs for breast cancer detection. J. Biomed. Biotechnol. 2011, 2011, 597145. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, E.; De Smaele, E.; Miele, E.; Laneve, P.; Po, A.; Pelloni, M.; Paganelli, A.; Di Marcotullio, L.; Caffarelli, E.; Screpanti, I.; et al. Concerted microRNA control of Hedgehog signalling in cerebellar neuronal progenitor and tumour cells. EMBO J. 2008, 27, 2616–2627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, W.-T.; Yu, S.-Y.; Li, S.-C.; Lam, H.-C.; Chang, H.-T.; Chen, W.-S.; Yeh, C.-Y.; Hung, S.-F.; Liu, T.-C.; Wu, T.; et al. MicroRNA-324 in Human Cancer: miR-324-5p and miR-324-3p Have Distinct Biological Functions in Human Cancer. Anticancer. Res. 2016, 36, 5189–5196. [Google Scholar] [CrossRef] [PubMed]
- Nie, J.; Liu, L.; Zheng, W.; Chen, L.; Wu, X.; Xu, Y.; Du, X.; Han, W. microRNA-365, down-regulated in colon cancer, inhibits cell cycle progression and promotes apoptosis of colon cancer cells by probably targeting Cyclin D1 and Bcl-2. Carcinogenesis 2012, 33, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Hamada, S.; Masamune, A.; Miura, S.; Satoh, K.; Shimosegawa, T. MiR-365 induces gemcitabine resistance in pancreatic cancer cells by targeting the adaptor protein SHC1 and pro–apoptotic regulator BAX. Cell Signal. 2014, 26, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Madhavan, D.; Peng, C.; Wallwiener, M.; Zucknick, M.; Nees, J.; Schott, S.; Rudolph, A.; Riethdorf, S.; Trumpp, A.; Pantel, K.; et al. Circulating miRNAs with prognostic value in metastatic breast cancer and for early detection of metastasis. Carcinogenesis 2016, 37, 461–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, A.; Shukla, K.; Balwierz, A.; Soons, Z.; König, R.; Sahin, Ö.; Wiemann, S. microRNA-519a is a novel oncomir conferring tamoxifen resistance by targeting a network of tumor-suppressor genes in ER+ breast cancer. J. Pathol. 2014, 233, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Breunig, C.; Pahl, J.; Kublbeck, M.; Miller, M.; Antonelli, D.; Erdem, N.; Wirth, C.; Will, R.; Bott, A.; Cerwenka, A.; et al. MicroRNA-519a-3p mediates apoptosis resistance in breast cancer cells and their escape from recognition by natural killer cells. Cell Death Dis. 2017, 8, e2973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yahya, S.M.M.; Elsayed, G.H. A summary for molecular regulations of miRNAs in breast cancer. Clin. Biochem. 2015, 48, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Zare, M.; Bastami, M.; Solali, S.; Alivand, M.R. Aberrant miRNA promoter methylation and EMT–involving miRNAs in breast cancer metastasis: Diagnosis and therapeutic implications. J. Cell. Physiol. 2017, 233, 3729–3744. [Google Scholar] [CrossRef] [PubMed]
- Weidle, U.H.; Dickopf, S.; Hintermair, C.; Kollmorgen, G.; Birzele, F.; Brinkmann, U. The Role of micro RNAs in Breast Cancer Metastasis: Preclinical Validation and Potential Therapeutic Targets. Cancer Genom. Proteom. 2018, 15, 17–39. [Google Scholar] [Green Version]
- Yang, F.; Ning, Z.; Ma, L.; Liu, W.; Shao, C.; Shu, Y.; Shen, H. Exosomal miRNAs and miRNA dysregulation in cancer-associated fibroblasts. Mol. Cancer 2017, 16, 148. [Google Scholar] [CrossRef] [PubMed]
- Yau, C.; Meyer, L.; Benz, S.; Vaske, C.; Scott, G.; Egan, B.; Labhart, P.; Pourmand, N.; Benz, C.C. FOXM1 cistrome predicts breast cancer metastatic outcome better than FOXM1 expression levels or tumor proliferation index. Breast Cancer Res. Treat. 2015, 154, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Liu, Y.; Yu, X.; Yin, L.; Peng, Y.; Gao, Y.; Zhu, Q.; Cao, T.; Yang, Y.; Fan, X.; et al. FOXM1 contributes to taxane resistance by regulating UHRF1-controlled cancer cell stemness. Cell Death Dis. 2018, 9, 562. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Tymen, S.D.; Chen, D.; Fang, Z.J.; Zhao, Y.; Dragas, D.; Dai, Y.; Marucha, P.T.; Zhou, X. MicroRNA-99 family targets AKT/mTOR signaling pathway in dermal wound healing. PLoS ONE 2013, 8, e64434. [Google Scholar] [CrossRef]
- Wang, X.; Li, Y.; Qi, W.; Zhang, N.; Sun, M.; Huo, Q.; Cai, C.; Lv, S.; Yang, Q. MicroRNA-99a inhibits tumor aggressive phenotypes through regulating HOXA1 in breast cancer cells. Oncotarget 2015, 6, 32737–32747. [Google Scholar] [CrossRef] [Green Version]
- Turcatel, G.; Rubin, N.; El-Hashash, A.; Warburton, D. MIR-99a and MIR-99b Modulate TGF-β Induced Epithelial to Mesenchymal Plasticity in Normal Murine Mammary Gland Cells. PLoS ONE 2012, 7, e31032. [Google Scholar] [CrossRef]
- Yang, Z.; Han, Y.; Cheng, K.; Zhang, G.; Wang, X. miR-99a directly targets the mTOR signalling pathway in breast cancer side population cells. Cell Prolif. 2014, 47, 587–595. [Google Scholar] [CrossRef]
- Haakensen, V.D.; Nygaard, V.; Greger, L.; Aure, M.R.; Fromm, B.; Bukholm, I.R.; Luders, T.; Chin, S.F.; Git, A.; Caldas, C.; et al. Subtype-specific microRNA expression signatures in breast cancer progression. Int. J. Cancer 2016, 139, 1117–1128. [Google Scholar] [CrossRef]
- Li, J.; Song, Z.J.; Wang, Y.Y.; Yin, Y.; Liu, Y.; Nan, X. Low levels of serum miR-99a is a predictor of poor prognosis in breast cancer. Genet. Mol. Res. 2016, 15, BSR20170696. [Google Scholar] [CrossRef]
- Kim, S.-W.; Ramasamy, K.; Bouamar, H.; Lin, A.-P.; Jiang, D.; Aguiar, R.C.T. MicroRNAs miR-125a and miR-125b constitutively activate the NF-κB pathway by targeting the tumor necrosis factor alpha-induced protein 3 (TNFAIP3, A20). Proc. Natl. Acad. Sci. USA 2012, 109, 7865–7870. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.V.; Ferracin, M.; Liu, C.-G.; Veronese, A.; Spizzo, R.; Sabbioni, S.; Magri, E.; Pedriali, M.; Fabbri, M.; Campiglio, M.; et al. MicroRNA gene expression deregulation in human breast cancer. Cancer Res. 2005, 65, 7065–7070. [Google Scholar] [CrossRef] [PubMed]
- Leucci, E.; Onnis, A.; Cocco, M.; De Falco, G.; Imperatore, F.; Giuseppina, A.; Costanzo, V.; Cerino, G.; Mannucci, S.; Cantisani, R.; et al. B-cell differentiation in EBV-positive Burkitt lymphoma is impaired at posttranscriptional level by miRNA–altered expression. Int. J. Cancer 2010, 126, 1316–1326. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kong, D. Knockdown of lncRNA MEG3 inhibits viability, migration, and invasion and promotes apoptosis by sponging miR-127 in osteosarcoma cell. J. Cell Biochem. 2018, 119, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhang, Y.; Wang, L. A feedback inhibition between miRNA-127 and TGFbeta/c-Jun cascade in HCC cell migration via MMP13. PLoS ONE 2013, 8, e65256. [Google Scholar]
- Pajic, M.; Froio, D.; Daly, S.; Doculara, L.; Millar, E.; Graham, P.H.; Drury, A.; Steinmann, A.; de Bock, C.E.; Boulghourjian, A.; et al. miR-139-5p modulates radiotherapy resistance in breast cancer by repressing multiple gene networks of DNA repair and ROS defense. Cancer Res. 2018, 78, 501–515. [Google Scholar] [CrossRef] [PubMed]
- Teng, H.; Wang, P.; Xue, Y.; Liu, X.; Ma, J.; Cai, H.; Xi, Z.; Li, Z.; Liu, Y. Role of HCP5-miR-139-RUNX1 Feedback Loop in Regulating Malignant Behavior of Glioma Cells. Mol. Ther. 2016, 24, 1806–1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.; Tong, Z.; Fang, W.-L.; Fu, Q.-B.; Gu, Y.-J.; Lv, T.-T.; Liu, D.-M.; Xue, W.; Lv, J.-W. Microrna-139-5p inhibits epithelial–mesenchymal transition and fibrosis in post-menopausal women with interstitial cystitis by targeting LPAR4 via the PI3K/Akt signaling pathway. J. Cell Biochem. 2018, 119, 6429–6441. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Zhang, D.; Shi, H.; Bian, Y.; Guo, R. MiR-143 inhibits endometrial cancer cell proliferation and metastasis by targeting MAPK1. Oncotarget 2017, 8, 84384–84395. [Google Scholar] [CrossRef]
- Zhang, Q.; Feng, Y.; Liu, P.; Yang, J. MiR-143 inhibits cell proliferation and invasion by targeting DNMT3A in gastric cancer. Tumour. Biol. 2017, 39. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Wang, X.W.; Qu, D.; Sun, J.W.; Guo, F.X.; Lu, D. Upregulation of microRNA-143 reverses drug resistance in human breast cancer cells via inhibition of cytokine–induced apoptosis inhibitor 1. Oncol. Lett. 2017, 13, 4695–4700. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Luo, Y.; Qiu, M. miR-143 Induces the Apoptosis of Prostate Cancer LNCap Cells by Suppressing Bcl-2 Expression. Med. Sci. Monit. 2017, 23, 359–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Liu, J.C.; Ju, Y.; Pellecchia, G.; Voisin, V.; Wang, D.Y.; Leha, L.R.; Ben-David, Y.; Bader, G.D.; Zacksenhaus, E. MicroRNA-143/145 loss induces Ras signaling to promote aggressive Pten-deficient basal-like breast cancer. JCI Insight 2017, 2, 93313. [Google Scholar] [CrossRef] [PubMed]
- Kent, O.A.; McCall, M.N.; Cornish, T.C.; Halushka, M.K. Lessons from miR-143/145: The importance of cell-type localization of miRNAs. Nucleic Acids Res. 2014, 42, 7528–7538. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Zhang, Y.; Tang, Y.; Li, Q. MALAT1 Modulates TGF-beta1-Induced Endothelial–to–Mesenchymal Transition through Downregulation of miR-145. Cell Physiol. Biochem. 2017, 42, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Ren, K.; Li, Z.; Li, Y.; Zhang, W.; Han, X. Long Noncoding RNA Taurine–Upregulated Gene 1 Promotes Cell Proliferation and Invasion in Gastric Cancer via Negatively Modulating miRNA-145-5p. Oncol. Res. 2017, 25, 789–798. [Google Scholar] [CrossRef]
- Hu, C.E.; Du, P.Z.; Zhang, H.D.; Huang, G.J. Long Noncoding RNA CRNDE Promotes Proliferation of Gastric Cancer Cells by Targeting miR-145. Cell Physiol. Biochem. 2017, 42, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Chi, H.; Chen, J.; Chen, C.; Huang, Y.; Xi, H.; Xue, J.; Si, Y. Curcumin suppresses proliferation and in vitro invasion of human prostate cancer stem cells by ceRNA effect of miR-145 and lncRNA-ROR. Gene 2017, 631, 29–38. [Google Scholar] [CrossRef]
- Xu, W.; Chang, J.; Du, X.; Hou, J. Long non-coding RNA PCAT-1 contributes to tumorigenesis by regulating FSCN1 via miR-145-5p in prostate cancer. Biomed. Pharmacother. 2017, 95, 1112–1118. [Google Scholar] [CrossRef]
- Kim, J.; Piao, H.-L.; Kim, B.-J.; Yao, F.; Han, Z.; Wang, Y.; Xiao, Z.; Siverly, A.N.; Lawhon, S.E.; Ton, B.N.; et al. Long noncoding RNA MALAT1 suppresses breast cancer metastasis. Nat. Genet. 2018, 50, 1705–1715. [Google Scholar] [CrossRef]
- Leivonen, S.-K.; Sahlberg, K.K.; Mäkelä, R.; Due, E.U.; Kallioniemi, O.; Børresen-Dale, A.-L.; Perälä, M. High–throughput screens identify microRNAs essential for HER2 positive breast cancer cell growth. Mol. Oncol. 2014, 8, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Hyun, J.; Wang, S.; Kim, J.; Rao, K.M.; Park, S.Y.; Chung, I.; Ha, C.-S.; Kim, S.-W.; Yun, Y.H.; Jung, Y. MicroRNA-378 limits activation of hepatic stellate cells and liver fibrosis by suppressing Gli3 expression. Nat. Commun. 2016, 7, 10993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, K.; Horie-Inoue, K.; Ueno, T.; Suzuki, T.; Sato, W.; Shigekawa, T.; Osaki, A.; Saeki, T.; Berezikov, E.; Mano, H.; et al. miR-378a-3p modulates tamoxifen sensitivity in breast cancer MCF-7 cells through targeting GOLT1A. Sci. Rep. 2015, 5, 13170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, S.; Pandey, A.; Shukla, A.; Talwelkar, S.S.; Kumar, A.; Pant, A.B.; Parmar, D. miR-497 and miR-302b Regulate Ethanol–induced Neuronal Cell Death through BCL2 Protein and Cyclin D2. J. Biol. Chem. 2011, 286, 37347–37357. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Kuang, D.; Zhao, X.; Chen, D.; Wang, X.; Yang, Q.; Wan, J.; Zhu, Y.; Wang, Y.; Zhang, S.; et al. miR-497-5p inhibits cell proliferation and invasion by targeting KCa3.1 in angiosarcoma. Oncotarget 2016, 7, 58148–58161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Zhao, Y.; Liu, C.; Chen, X.; Qi, Y.; Jiang, Y.; Zou, C.; Zhang, X.; Liu, S.; Wang, X.; et al. Analysis of miR-195 and miR-497 Expression, Regulation and Role in Breast Cancer. Clin. Cancer Res. 2011, 17, 1722–1730. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Li, X.; Niu, Y.; Zhu, S.; Jin, Y.; Deng, S.; Chen, J.; Liu, Y.; He, C.; Yin, T.; et al. miR-652 inhibits acidic microenvironment-induced epithelial-mesenchymal transition of pancreatic cancer cells by targeting ZEB1. Oncotarget 2015, 6, 39661–39675. [Google Scholar] [CrossRef] [PubMed]
- Schmalhofer, O.; Brabletz, S.; Brabletz, T. E-cadherin, β-catenin, and ZEB1 in malignant progression of cancer. Cancer Metast. Rev. 2009, 28, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Mangolini, A.; Ferracin, M.; Zanzi, M.V.; Saccenti, E.; Ebnaof, S.O.; Poma, V.V.; Sanz, J.M.; Passaro, A.; Pedriali, M.; Frassoldati, A.; et al. Diagnostic and prognostic microRNAs in the serum of breast cancer patients measured by droplet digital PCR. Biomarker Res. 2015, 3, 12. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Tan, W.; Chen, K.; You, N.; Zhu, S.; Liang, G.; Xie, X.; Li, Q.; Zeng, Y.; Ouyang, N.; et al. Prognostic Value of a BCSC-associated MicroRNA Signature in Hormone Receptor-Positive HER2-Negative Breast Cancer. EBioMedicine 2016, 11, 199–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singla, H.; Ludhiadch, A.; Kaur, R.P.; Chander, H.; Kumar, V.; Munshi, A. Recent advances in HER2 positive breast cancer epigenetics: Susceptibility and therapeutic strategies. Eur. J. Med. Chem. 2017, 142, 316–327. [Google Scholar] [CrossRef] [PubMed]
- D’Ippolito, E.; Iorio, M.V. MicroRNAs and triple negative breast cancer. Int. J. Mol. Sci. 2013, 14, 22202–22220. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Wang, X.; Han, W.; Li, D.; Xu, Y.; Lou, F.; Zhou, J.; Gu, X.; Zhu, J.; Zhang, C.; et al. MicroRNAs-mediated cell fate in triple negative breast cancers. Cancer Lett. 2015, 361, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Mao, X.; Shi, P.; He, B.; Xu, K.; Zhang, S.; Wang, J. MicroRNAs in the prognosis of triple-negative breast cancer: A systematic review and meta-analysis. Medicine (Baltimore) 2017, 96, e7085. [Google Scholar] [CrossRef]
- Dykes, I.M.; Emanueli, C. Transcriptional and Post-transcriptional Gene Regulation by Long Non-coding RNA. Genom. Proteom. Bioinform. 2017, 15, 177–186. [Google Scholar] [CrossRef]
- Assefa, A.T.; De Paepe, K.; Everaert, C.; Mestdagh, P.; Thas, O.; Vandesompele, J. Differential gene expression analysis tools exhibit substandard performance for long non-coding RNA-sequencing data. Genome Biol. 2018, 19, 96. [Google Scholar] [CrossRef]
- De Paepe, B.; Lefever, S.; Mestdagh, P. How long noncoding RNAs enforce their will on mitochondrial activity: Regulation of mitochondrial respiration, reactive oxygen species production, apoptosis, and metabolic reprogramming in cancer. Curr. Genet. 2018, 64, 163–172. [Google Scholar] [CrossRef]
- Gao, S.; Tian, X.; Chang, H.; Sun, Y.; Wu, Z.; Cheng, Z.; Dong, P.; Zhao, Q.; Ruan, J.; Bu, W. Two novel lncRNAs discovered in human mitochondrial DNA using PacBio full-length transcriptome data. Mitochondrion 2018, 38, 41–47. [Google Scholar] [CrossRef]
- Ørom Ulf, A.; Shiekhattar, R. Long Noncoding RNAs Usher in a New Era in the Biology of Enhancers. Cell 2013, 154, 1190–1193. [Google Scholar] [CrossRef]
- Marchese, F.P.; Raimondi, I.; Huarte, M. The multidimensional mechanisms of long noncoding RNA function. Genome Biol. 2017, 18, 206. [Google Scholar] [CrossRef]
- Corrà, F.; Agnoletto, C.; Minotti, L.; Baldassari, F.; Volinia, S. The Network of Non-coding RNAs in Cancer Drug Resistance. Front. Oncol. 2018, 8, 327. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, L.; Wang, Y.; Li, H.; Ren, X.; Wei, F.; Yu, W.; Wang, X.; Zhang, L.; Yu, J.; et al. Long noncoding RNA HOTAIR involvement in cancer. Tumor Biol. 2014, 35, 9531–9538. [Google Scholar] [CrossRef] [PubMed]
- Malih, S.; Saidijam, M.; Malih, N. A brief review on long noncoding RNAs: A new paradigm in breast cancer pathogenesis, diagnosis and therapy. Tumor Biol. 2016, 37, 1479–1485. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Li, S.; Song, E.; Liu, S. The roles of ncRNAs and histone-modifiers in regulating breast cancer stem cells. Protein Cell 2016, 7, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, A.; Jain, M.; Prakash, H.; Vasquez, K.M.; Jain, A. The regulatory roles of long non-coding RNAs in the development of chemoresistance in breast cancer. Oncotarget 2017, 8, 110671–110684. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ye, C.; Xiong, H.; Shen, Y.; Lu, Y.; Zhou, J.; Wang, L. Dysregulation of long non-coding RNA in breast cancer: An overview of mechanism and clinical implication. Oncotarget 2017, 8, 5508–5522. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Gao, S.; Li, H.; Lv, M.; Lu, C. Long Non-coding RNAs inTriple Negative Breast Cancer. J. Cell Physiol. 2017, 232, 3226–3233. [Google Scholar] [CrossRef] [PubMed]
- Warburton, A.J.; Boone, D.N. Insights from Global Analyses of Long Noncoding RNAs in Breast Cancer. Curr. Pathobiol. Rep. 2017, 5, 23–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koleckova, M.; Janikova, M.; Kolar, Z. MicroRNAs in triple-negative breast cancer. Neoplasma 2018, 65, 1–13. [Google Scholar] [CrossRef]
- Richard, J.L.C.; Eichhorn, P.J.A. Deciphering the roles of lncRNAs in breast development and disease. Oncotarget 2018, 9, 20179–20212. [Google Scholar] [CrossRef]
- Tian, T.; Wang, M.; Lin, S.; Guo, Y.; Dai, Z.; Liu, K.; Yang, P.; Dai, C.; Zhu, Y.; Zheng, Y.; et al. The Impact of lncRNA Dysregulation on Clinicopathology and Survival of Breast Cancer: A Systematic Review and Meta-analysis. Mol. Ther. Nucleic Acids 2018, 12, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Bradford, J.R.; Cox, A.; Bernard, P.; Camp, N.J. Consensus Analysis of Whole Transcriptome Profiles from Two Breast Cancer Patient Cohorts Reveals Long Non-Coding RNAs Associated with Intrinsic Subtype and the Tumour Microenvironment. PLoS ONE 2016, 11, e0163238. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Gadad Shrikanth, S.; Kim, D.-S.; Kraus, W.L. Discovery, Annotation, and Functional Analysis of Long Noncoding RNAs Controlling Cell-Cycle Gene Expression and Proliferation in Breast Cancer Cells. Mol. Cell 2015, 59, 698–711. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Zhong, L.; Xu, W.; Sun, Y.; Zhang, Z.; Zhao, H.; Yang, L.; Sun, J. Discovery of potential prognostic long non-coding RNA biomarkers for predicting the risk of tumor recurrence of breast cancer patients. Sci. Rep. 2016, 6, 31038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.; Wang, Q.; Zhan, Y.; Chen, X.; Yu, Q.; Zhang, J.; Wang, Y.; Xu, X.-j.; Zhu, L. Transcriptome sequencing uncovers a three-long noncoding RNA signature in predicting breast cancer survival. Sci. Rep. 2016, 6, 27931. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Yoshitomi, T.; Hu, J.F.; Cui, J. Long noncoding RNAs coordinate functions between mitochondria and the nucleus. Epigenetics Chromatin 2017, 10, 41. [Google Scholar] [CrossRef] [PubMed]
- Rheinbay, E.; Parasuraman, P.; Grimsby, J.; Tiao, G.; Engreitz, J.M.; Kim, J.; Lawrence, M.S.; Taylor-Weiner, A.; Rodriguez-Cuevas, S.; Rosenberg, M.; et al. Recurrent and functional regulatory mutations in breast cancer. Nature 2017, 547, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Niknafs, Y.S.; Han, S.; Ma, T.; Speers, C.; Zhang, C.; Wilder-Romans, K.; Iyer, M.K.; Pitchiaya, S.; Malik, R.; Hosono, Y.; et al. The lncRNA landscape of breast cancer reveals a role for DSCAM-AS1 in breast cancer progression. Nat. Commun. 2016, 7, 12791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spector, D.L.; Lamond, A.I. Nuclear Speckles. Cold Spring Harbor Perspect. Biol. 2011, 3, A000646. [Google Scholar] [CrossRef] [PubMed]
- Galganski, L.; Urbanek, M.O.; Krzyzosiak, W.J. Nuclear speckles: Molecular organization, biological function and role in disease. Nucleic Acids Res. 2017, 45, 10350–10368. [Google Scholar] [CrossRef]
- Li, W.; Zhang, Z.; Liu, X.; Cheng, X.; Zhang, Y.; Han, X.; Zhang, Y.; Liu, S.; Yang, J.; Xu, B.; et al. The FOXN3-NEAT1-SIN3A repressor complex promotes progression of hormonally responsive breast cancer. J. Clin. Investig. 2017, 127, 3421–3440. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Wang, L.; Yang, Q.; Liu, H.; Meng, Q.; Jiang, L.; Wang, S.; Jiang, W. Systematical analysis of lncRNA-mRNA competing endogenous RNA network in breast cancer subtypes. Breast Cancer Res. Treat. 2018, 169, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.C.; Korkut, A.; Kanchi, R.S.; Hegde, A.M.; Lenoir, W.; Liu, W.; Liu, Y.; Fan, H.; Shen, H.; Ravikumar, V.; et al. A Comprehensive Pan-Cancer Molecular Study of Gynecologic and Breast Cancers. Cancer Cell 2018, 33, 690–705.e699. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wagner, E.K.; Guo, X.; May, I.; Cai, Q.; Zheng, W.; He, C.; Long, J. Long intergenic non–coding RNA expression signature in human breast cancer. Sci. Rep. 2016, 6, 37821. [Google Scholar] [CrossRef] [PubMed]
- Xiao, B.; Zhang, W.; Chen, L.; Hang, J.; Wang, L.; Zhang, R.; Liao, Y.; Chen, J.; Ma, Q.; Sun, Z.; et al. Analysis of the miRNA-mRNA-lncRNA network in human estrogen receptor–positive and estrogen receptor-negative breast cancer based on TCGA data. Gene 2018, 658, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Lanz, R.B.; McKenna, N.J.; Onate, S.A.; Albrecht, U.; Wong, J.; Tsai, S.Y.; Tsai, M.J.; O’Malley, B.W. A steroid receptor coactivator, SRA, functions as an RNA and is present in an SRC-1 complex. Cell 1999, 97, 17–27. [Google Scholar] [CrossRef]
- Murphy, L.C.; Simon, S.L.; Parkes, A.; Leygue, E.; Dotzlaw, H.; Snell, L.; Troup, S.; Adeyinka, A.; Watson, P.H. Altered expression of estrogen receptor coregulators during human breast tumorigenesis. Cancer Res. 2000, 60, 6266–6271. [Google Scholar] [PubMed]
- Novikova, I.V.; Hennelly, S.P.; Sanbonmatsu, K.Y. Structural architecture of the human long non-coding RNA, steroid receptor RNA activator. Nucleic Acids Res. 2012, 40, 5034–5051. [Google Scholar] [CrossRef] [PubMed]
- Leygue, E. Steroid receptor RNA activator (SRA1): Unusual bifaceted gene products with suspected relevance to breast cancer. Nucl. Recept. Signal. 2007, 5, e006. [Google Scholar] [CrossRef] [PubMed]
- Sheng, L.; Ye, L.; Zhang, D.; Cawthorn, W.P.; Xu, B. New Insights Into the Long Non–coding RNA SRA: Physiological Functions and Mechanisms of Action. Front. Med. 2018, 5, 244. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hamblin, M.H.; Yin, K.J. The long noncoding RNA Malat1: Its physiological and pathophysiological functions. RNA Biol. 2017, 14, 1705–1714. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.K.; Luo, Q.; Peng, H.; Li, J.; Zhao, M.; Wang, J.; Gu, Y.Y.; Li, Y.; Yuan, P.; Zhao, G.H. A Panel of Serum Noncoding RNAs for the Diagnosis and Monitoring of Response to Therapy in Patients with Breast Cancer. Med. Sci. Monit. 2018, 24, 2476–2488. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, M.; Zacheaus, C.; Doxtater, K.; Keramatnia, F.; Gao, C.; Yallapu, M.; Jaggi, M.; Chauhan, S.; Probe, Z. An Efficient Tool for Characterizing Long Non-Coding RNA in FFPE Tissues. Non-Cod. RNA 2018, 4, 20. [Google Scholar] [CrossRef] [PubMed]
- Ong, M.S.; Cai, W.; Yuan, Y.; Leong, H.C.; Tan, T.Z.; Mohammad, A.; You, M.L.; Arfuso, F.; Goh, B.C.; Warrier, S.; et al. ‘Lnc’-ing Wnt in female reproductive cancers: Therapeutic potential of long non-coding RNAs in Wnt signalling. Br. J. Pharmacol. 2017, 174, 4684–4700. [Google Scholar] [CrossRef] [PubMed]
- Arun, G.; Diermeier, S.; Akerman, M.; Chang, K.C.; Wilkinson, J.E.; Hearn, S.; Kim, Y.; MacLeod, A.R.; Krainer, A.R.; Norton, L.; et al. Differentiation of mammary tumors and reduction in metastasis upon Malat1 lncRNA loss. Genes Dev. 2016, 30, 34–51. [Google Scholar] [CrossRef] [PubMed]
- Nik–Zainal, S.; Davies, H.; Staaf, J.; Ramakrishna, M.; Glodzik, D.; Zou, X.; Martincorena, I.; Alexandrov, L.B.; Martin, S.; Wedge, D.C.; et al. Landscape of somatic mutations in 560 breast cancer whole-genome sequences. Nature 2016, 534, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Liu, Y.; Zhuang, H.; Yang, B.; Hei, K.; Xiao, M.; Hou, C.; Gao, H.; Zhang, X.; Jia, C.; et al. Quantitative proteomics reveals that long non-coding RNA MALAT1 interacts with DBC1 to regulate p53 acetylation. Nucleic Acids Res. 2017, 45, 9947–9959. [Google Scholar] [CrossRef] [Green Version]
- Jadaliha, M.; Zong, X.; Malakar, P.; Ray, T.; Singh, D.K.; Freier, S.M.; Jensen, T.; Prasanth, S.G.; Karni, R.; Ray, P.S.; et al. Functional and prognostic significance of long non-coding RNA MALAT1 as a metastasis driver in ER negative lymph node negative breast cancer. Oncotarget 2016, 7, 40418–40436. [Google Scholar] [CrossRef] [Green Version]
- Redis, R.S.; Sieuwerts, A.M.; Look, M.P.; Tudoran, O.; Ivan, C.; Spizzo, R.; Zhang, X.; de Weerd, V.; Shimizu, M.; Ling, H.; et al. CCAT2, a novel long non-coding RNA in breast cancer: Expression study and clinical correlations. Oncotarget 2013, 4, 1748–1762. [Google Scholar] [CrossRef]
- Cai, Y.; He, J.; Zhang, D. Long noncoding RNA CCAT2 promotes breast tumor growth by regulating the Wnt signaling pathway. OncoTargets Ther. 2015, 8, 2657–2664. [Google Scholar]
- Wu, Z.J.; Li, Y.; Wu, Y.Z.; Wang, Y.; Nian, W.Q.; Wang, L.L.; Li, L.C.; Luo, H.L.; Wang, D.L. Long non–coding RNA CCAT2 promotes the breast cancer growth and metastasis by regulating TGF-beta signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 706–714. [Google Scholar] [PubMed]
- Deng, X.; Zhao, Y.; Wu, X.; Song, G. Upregulation of CCAT2 promotes cell proliferation by repressing the P15 in breast cancer. Biomed. Pharmacother. 2017, 91, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, C.; Garbe, J.; Bhattacharya, N.; Daksis, J.; Pan, C.H.; Yaswen, P.; Koh, J.; Slingerland, J.M.; Stampfer, M.R. Transforming growth factor beta stabilizes p15INK4B protein, increases p15INK4B-cdk4 complexes, and inhibits cyclin D1-cdk4 association in human mammary epithelial cells. Mol. Cell Biol. 1997, 17, 2458. [Google Scholar] [CrossRef] [PubMed]
- Huan, J.; Xing, L.; Lin, Q.; Xui, H.; Qin, X. Long noncoding RNA CRNDE activates Wnt/beta–catenin signaling pathway through acting as a molecular sponge of microRNA-136 in human breast cancer. Am. J. Trans. Res. 2017, 9, 1977–1989. [Google Scholar]
- Nusse, R.; Clevers, H. Wnt/β-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Holland, J.D.; Klaus, A.; Garratt, A.N.; Birchmeier, W. Wnt signaling in stem and cancer stem cells. Curr. Opin. Cell Biol. 2013, 25, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Z.; Jiang, J.; Xu, C.; Kang, J.; Xiao, L.; Wu, M.; Xiong, J.; Guo, X.; Liu, H. Endogenous miRNA Sponge lincRNA-RoR Regulates Oct4, Nanog, and Sox2 in Human Embryonic Stem Cell Self–Renewal. Dev. Cell 2013, 25, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Hou, P.; Zhao, Y.; Li, Z.; Yao, R.; Ma, M.; Gao, Y.; Zhao, L.; Zhang, Y.; Huang, B.; Lu, J. LincRNA–ROR induces epithelial-to-mesenchymal transition and contributes to breast cancer tumorigenesis and metastasis. Cell Death Dis. 2014, 5, e1287. [Google Scholar] [CrossRef]
- Hou, L.; Tu, J.; Cheng, F.; Yang, H.; Yu, F.; Wang, M.; Liu, J.; Fan, J.; Zhou, G. Long noncoding RNA ROR promotes breast cancer by regulating the TGF-β pathway. Cancer Cell Int. 2018, 18, 142. [Google Scholar] [CrossRef]
- Zhang, J.J.; Guo, S.H.; Jia, B.Q. Down-regulation of long non–coding RNA MEG3 serves as an unfavorable risk factor for survival of patients with breast cancer. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 5143–5147. [Google Scholar]
- Zhang, W.; Shi, S.; Jiang, J.; Li, X.; Lu, H.; Ren, F. LncRNA MEG3 inhibits cell epithelial-mesenchymal transition by sponging miR-421 targeting E-cadherin in breast cancer. Biomed. Pharmacother. 2017, 91, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Y.; Yu, M.S.; Li, X.; Zhang, Z.; Han, C.R.; Yan, B. Overexpression of long non-coding RNA MEG3 suppresses breast cancer cell proliferation, invasion, and angiogenesis through AKT pathway. Tumor Biol. 2017, 39. [Google Scholar] [CrossRef] [PubMed]
- Malissovas, N.; Ninou, E.; Michail, A.; Politis, P.K. Targeting Long Non-Coding RNAs in Nervous System Cancers: New Insights in Prognosis, Diagnosis and Therapy. Curr. Med. Chem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Luo, S.; Zhang, X.; Zou, C.; Yuan, H.; Liao, G.; Xu, L.; Deng, C.; Lan, Y.; Zhao, T.; et al. A pan–cancer atlas of cancer hallmark–associated candidate driver lncRNAs. Mol. Oncol. 2018, 12, 1980–2005. [Google Scholar] [CrossRef]
- Colombo, M.; Raposo, G.; Thery, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef] [PubMed]
- Melo Sonia, A.; Sugimoto, H.; O’Connell Joyce, T.; Kato, N.; Villanueva, A.; Vidal, A.; Qiu, L.; Vitkin, E.; Perelman Lev, T.; Melo Carlos, A.; et al. Cancer Exosomes Perform Cell-Independent MicroRNA Biogenesis and Promote Tumorigenesis. Cancer Cell 2014, 26, 707–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, R.E.; Korbie, D.; Hill, M.M.; Trau, M. Extracellular vesicles as circulating cancer biomarkers: Opportunities and challenges. Clin. Transl. Med. 2018, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, Y.; Chen, B.; Zhao, J.; Yu, S.; Tang, Y.; Zheng, Q.; Li, Y.; Wang, P.; He, X.; et al. exoRBase: A database of circRNA, lncRNA and mRNA in human blood exosomes. Nucleic Acids Res. 2018, 46, D106–D112. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.; Rani, S.; Corcoran, C.; Wallace, R.; Hughes, L.; Friel, A.M.; McDonnell, S.; Crown, J.; Radomski, M.W.; O’Driscoll, L. Exosomes from triple-negative breast cancer cells can transfer phenotypic traits representing their cells of origin to secondary cells. Eur. J. Cancer 2013, 49, 1845–1859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sansone, P.; Savini, C.; Kurelac, I.; Chang, Q.; Amato, L.B.; Strillacci, A.; Stepanova, A.; Iommarini, L.; Mastroleo, C.; Daly, L.; et al. Packaging and transfer of mitochondrial DNA via exosomes regulate escape from dormancy in hormonal therapy–resistant breast cancer. Proc. Natl. Acad. Sci. USA 2017, 114, E9066–E9075. [Google Scholar] [CrossRef]
- Halvaei, S.; Daryani, S.; Eslami-S, Z.; Samadi, T.; Jafarbeik-Iravani, N.; Bakhshayesh, T.O.; Majidzadeh-A, K.; Esmaeili, R. Exosomes in Cancer Liquid Biopsy: A Focus on Breast Cancer. Mol. Ther. Nucleic Acids 2018, 10, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Hannafon, B.N.; Trigoso, Y.D.; Calloway, C.L.; Zhao, Y.D.; Lum, D.H.; Welm, A.L.; Zhao, Z.J.; Blick, K.E.; Dooley, W.C.; Ding, W.Q. Plasma exosome microRNAs are indicative of breast cancer. Breast Cancer Res. 2016, 18, 90. [Google Scholar] [CrossRef] [PubMed]
- Sempere, L.F.; Keto, J.; Fabbri, M. Exosomal MicroRNAs in Breast Cancer towards Diagnostic and Therapeutic Applications. Cancers 2017, 9, 71. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Lai, X.; Yu, S.; Chen, S.; Ma, Y.; Zhang, Y.; Li, H.; Zhu, X.; Yao, L.; Zhang, J. Exosomal miR-221/222 enhances tamoxifen resistance in recipient ER-positive breast cancer cells. Breast Cancer Res. Treat. 2014, 147, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.G.; Yang, M.F.; Ren, Y.Q.; Wu, C.H.; Wang, L.Q. Exosomes mediated transfer of lncRNA UCA1 results in increased tamoxifen resistance in breast cancer cells. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4362–4368. [Google Scholar] [PubMed]
- Muluhngwi, P.; Alizadeh-Rad, N.; Vittitow, S.L.; Kalbfleisch, T.S.; Klinge, C.M. The miR-29 transcriptome in endocrine-sensitive and resistant breast cancer cells. Sci. Rep. 2017, 7, 5205. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Luo, J. Long Non–Coding RNA (lncRNA) Urothelial Carcinoma-Associated 1 (UCA1) Enhances Tamoxifen Resistance in Breast Cancer Cells via Inhibiting mTOR Signaling Pathway. Med. Sci. Monit. 2016, 22, 3860–3867. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhou, H.; Lu, K.; Lu, Y.; Wang, Y.; Feng, T. Exosome-mediated delivery of MALAT1 induces cell proliferation in breast cancer. OncoTargets Ther. 2018, 11, 291–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
miRNAs regulating ERα transcriptional activity. (A) ERα is directly targeted by the indicated miRNAs that are increased in breast tumors (Table 2). E2-liganded ERα recruits coactivators and chromatin remodeling complexes to increase RNA pol II transcription at target genes. E2–ERα increases the transcription of FOXM1, which, in turn as a transcription factor, increases the transcription of ERα, including a number of genes for cell cycle progression [224], and UHRF1, which is a key regulator of DNA methylation that is involved in the self-renewal and differentiation of cancer stem cells [225]. (B) The selective ER modulator (SERM) tamoxifen is metabolized to 4-hydroxytamoxifen, which binds ERα and alters its conformation, thus inhibiting coactivator recruitment, and instead allowing interaction of the 4-OHT-bound-ERα with corepressors, including NCOR2, which recruits histone deacetylase complex (HDAC) complexes to inhibit target gene transcription in breast tumors. NCOR2 is a target of miR-10a-5p (Table 2).
Figure 1.
miRNAs regulating ERα transcriptional activity. (A) ERα is directly targeted by the indicated miRNAs that are increased in breast tumors (Table 2). E2-liganded ERα recruits coactivators and chromatin remodeling complexes to increase RNA pol II transcription at target genes. E2–ERα increases the transcription of FOXM1, which, in turn as a transcription factor, increases the transcription of ERα, including a number of genes for cell cycle progression [224], and UHRF1, which is a key regulator of DNA methylation that is involved in the self-renewal and differentiation of cancer stem cells [225]. (B) The selective ER modulator (SERM) tamoxifen is metabolized to 4-hydroxytamoxifen, which binds ERα and alters its conformation, thus inhibiting coactivator recruitment, and instead allowing interaction of the 4-OHT-bound-ERα with corepressors, including NCOR2, which recruits histone deacetylase complex (HDAC) complexes to inhibit target gene transcription in breast tumors. NCOR2 is a target of miR-10a-5p (Table 2).

Figure 2.
Breast cancer dysregulated miRNAs and lncRNAs as ceRNAs in cell signaling, cell cycle, and EMT. Shown are validated targets of some miRNAs dysregulated in human tumors and lncRNAs that at as ceRNAs for the indicated miRNAs.
Figure 2.
Breast cancer dysregulated miRNAs and lncRNAs as ceRNAs in cell signaling, cell cycle, and EMT. Shown are validated targets of some miRNAs dysregulated in human tumors and lncRNAs that at as ceRNAs for the indicated miRNAs.

Figure 3.
Breast cancer dysregulated miRNAs in apoptosis. Shown in abbreviated form are key regulators in the intrinsic and extrinsic pathways of apoptosis and their regulation by miRNAs that are dysregulated in breast tumors (Table 2 and Table 3). The lncRNA MIAT is a ceRNA for miR-155-5p (Table 2).
Figure 3.
Breast cancer dysregulated miRNAs in apoptosis. Shown in abbreviated form are key regulators in the intrinsic and extrinsic pathways of apoptosis and their regulation by miRNAs that are dysregulated in breast tumors (Table 2 and Table 3). The lncRNA MIAT is a ceRNA for miR-155-5p (Table 2).

Figure 4.
Exosomal transfer of miRNAs and lncRNAs in breast cancer. Exosomes released from breast cancer cells and cancer-associated fibroblasts into the extracellular compartment contain ncRNAs, mRNAs, mtDNA, proteins, and lipids. Exosomes can deliver their contents to adjacent cells or cells at a distance. Examples of miRNAs and lncRNAs in breast cancer exosomes and their known roles in breast cancer are shown.
Figure 4.
Exosomal transfer of miRNAs and lncRNAs in breast cancer. Exosomes released from breast cancer cells and cancer-associated fibroblasts into the extracellular compartment contain ncRNAs, mRNAs, mtDNA, proteins, and lipids. Exosomes can deliver their contents to adjacent cells or cells at a distance. Examples of miRNAs and lncRNAs in breast cancer exosomes and their known roles in breast cancer are shown.


{kind=link}
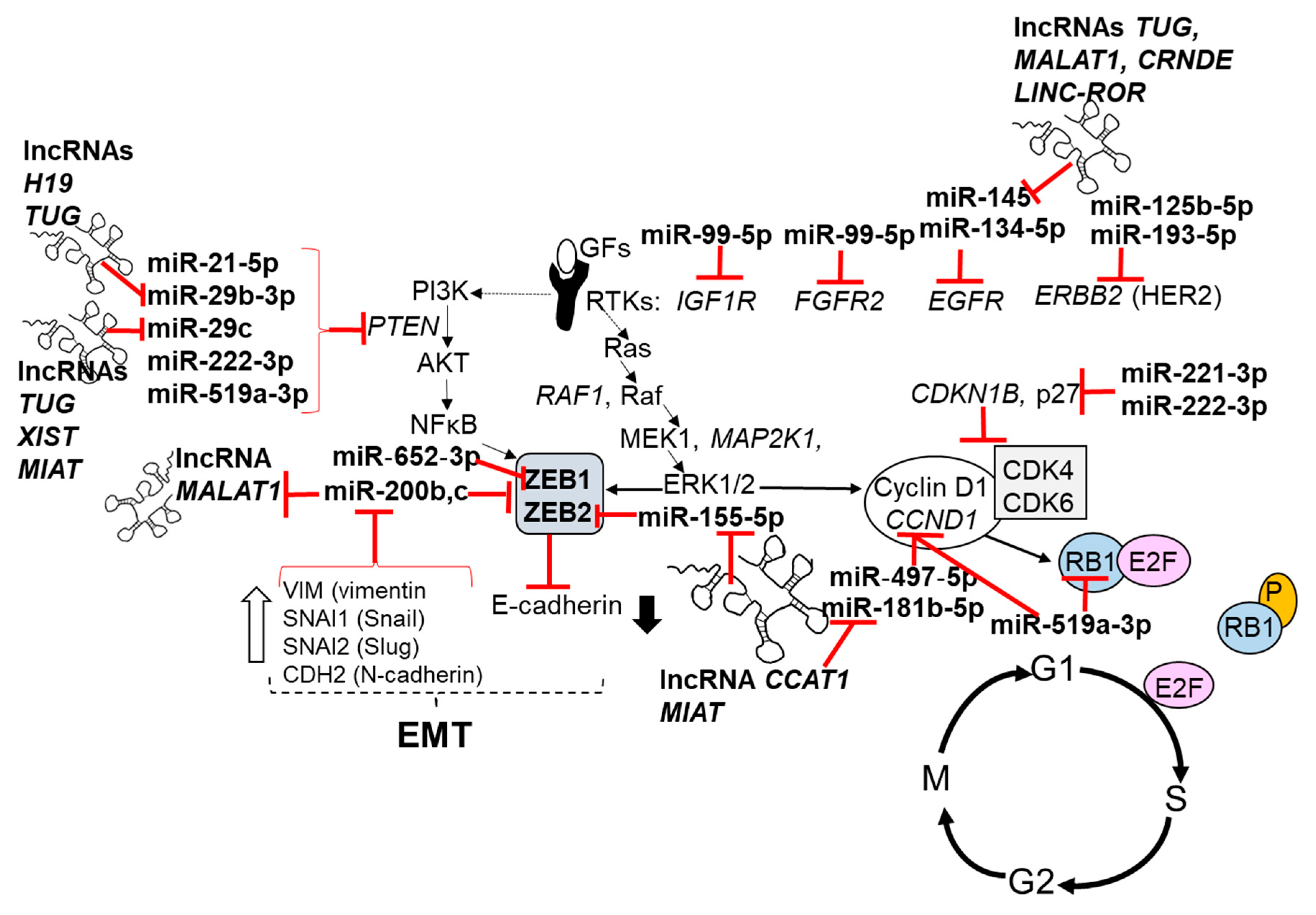{kind=link}
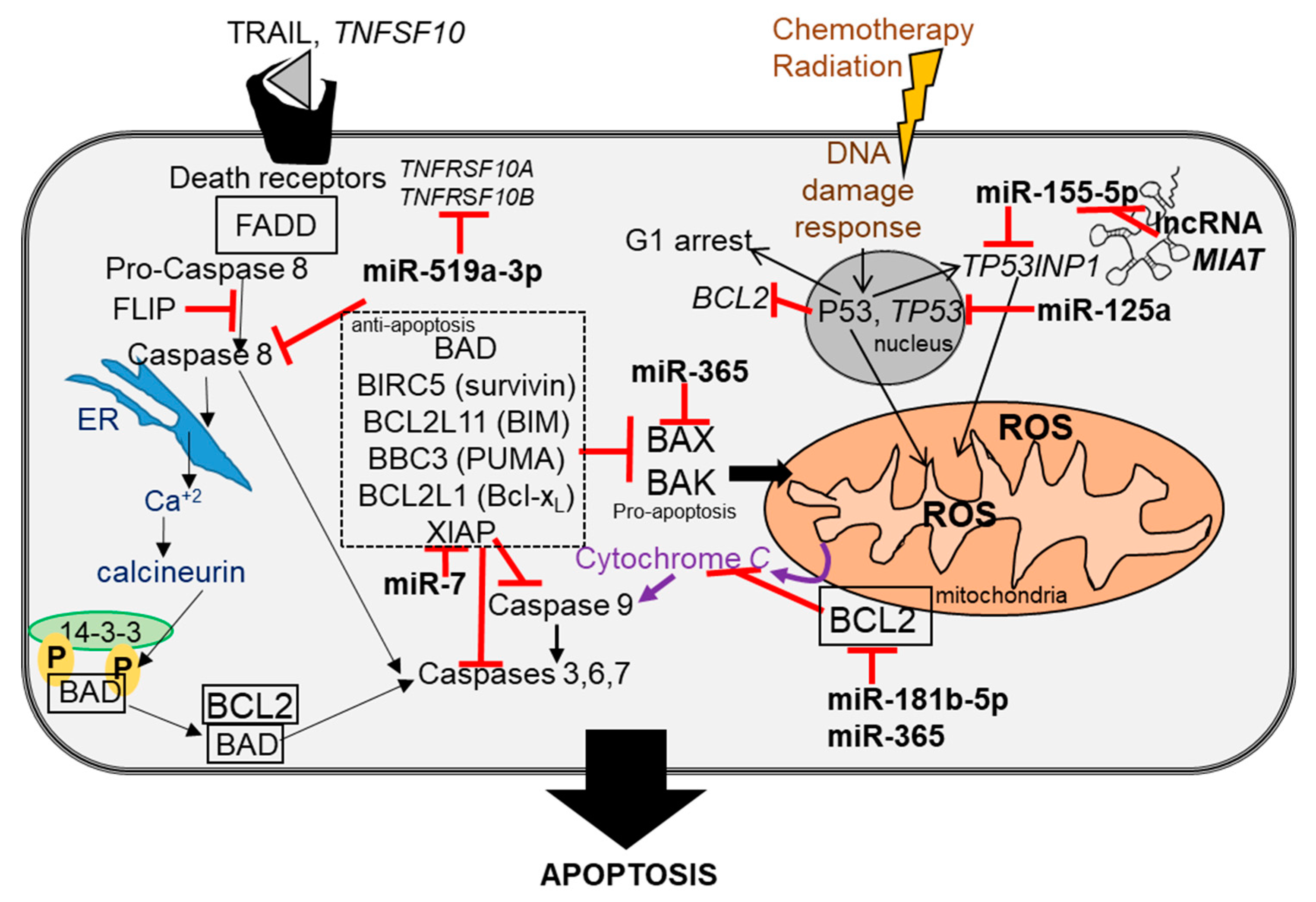{kind=link}
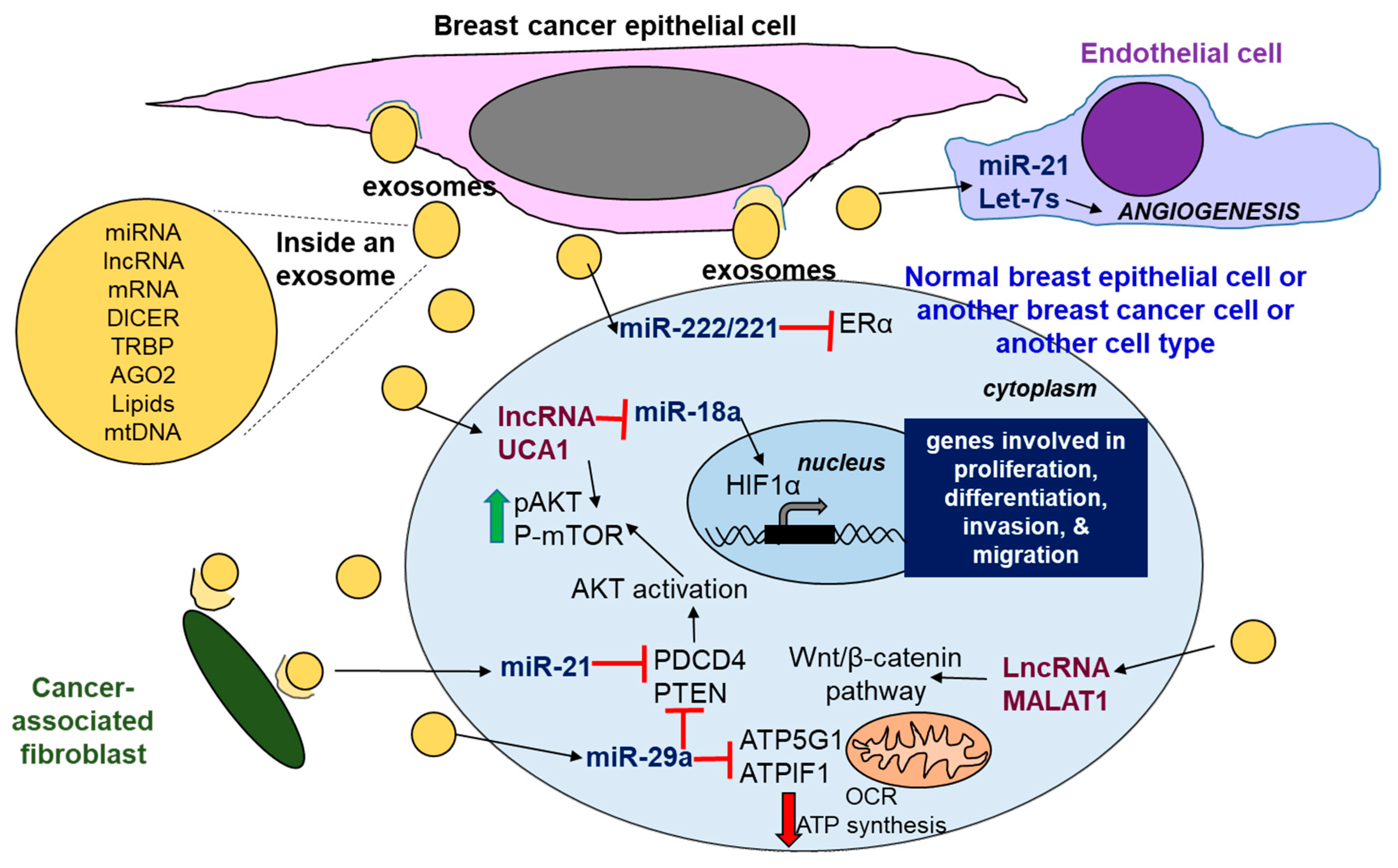{kind=link}
Table 1.
Regulatory non-coding RNAs (ncRNAs) in breast cancer. Abbreviations: BCa (breast cancer), ceRNA: competing endogenous RNA; TNBC: triple negative breast cancer, ssRNA: single-stranded RNA.
Table 1.
Regulatory non-coding RNAs (ncRNAs) in breast cancer. Abbreviations: BCa (breast cancer), ceRNA: competing endogenous RNA; TNBC: triple negative breast cancer, ssRNA: single-stranded RNA.
| ncRNA | Description, Size, Cellular Location, Function | Examples in Breast Cancer |
|---|---|---|
| Circular RNAs (circRNAs) | Circular RNAs: Four types: circular RNAs from introns, exonic circRNAs, exon-intron circRNAs (EIciRNA), and intergenic circRNAs [29]
| Tumor-specific circRNAs were identified in human breast tumors and BCa cells (BT-20, BT-474, MCF-7, MDA-MB-231, MDA-MB-468, T-47D, and ZR-75-1) [33] circ-ABCB10 was upregulated in BCa tumors and acted as a ceRNA for miR-1271 [34] hsa_circ_0001982 was overexpressed in BCa tissues and cell lines, and acted as a ceRNA for miR-143 [35]. circGFRA1 was upregulated in TNBC and acted as a ceRNA for miR-34a Analysis using CircHunter and HashCirc recently identified circRNAs in MCF-7 BCa cells [36] circIRAK3, was increased in TNBC cells (MDA-MB-231, MDA-MB-157, HCC70, HCC1806, HCC1937) and acts as a ceRNA for miR-3607 [37] circ_0008039 was upregulated in BCa tissues, and acted as a ceRNA for miR-432-5 [38] |
| PIWI-interacting RNAs (piRNA) | piRNAs: DQ596670, DQ598183, DQ597341, DQ598252, and DQ596311 were underexpressed; DQ598677, DQ597960, and DQ570994 overexpressed in BCa tissues compared to normal breast tissue [41]. piR-021285 was identified as a potential modulator of BCa invasiveness, which is a function linked to piR-021285-dependent 5′ UTR/first exon methylation of the pro-invasive ARHGAP11A gene [42]. | |
| MicroRNAs (miRNA) | ~22 nt ssRNA
| Reviewed in [43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62]. |
| lncRNA |
| Reviewed in [63,64,65,66] |
Table 2.
Examples of miRNAs upregulated in breast tumors, patient plasma, and breast cancer cell lines with their authenticated targets. miRTarBase was used to identify validated targets [119], i.e., those not identified by reference number. Abbreviations: AI = aromatase inhibitors, BCa = breast cancer; ceRNA = competing endogenous RNA (‘miRNA sponge’), CSCs = cancer stem cells, DCIS = ductal carcinoma in situ, EMT = epithelial-to-mesenchymal transformation, ERα = estrogen receptor α, HN = healthy normal, PR = progesterone receptor, TAM = tamoxifen, TNBC = triple negative breast cancer, TAM = tamoxifen.
Table 2.
Examples of miRNAs upregulated in breast tumors, patient plasma, and breast cancer cell lines with their authenticated targets. miRTarBase was used to identify validated targets [119], i.e., those not identified by reference number. Abbreviations: AI = aromatase inhibitors, BCa = breast cancer; ceRNA = competing endogenous RNA (‘miRNA sponge’), CSCs = cancer stem cells, DCIS = ductal carcinoma in situ, EMT = epithelial-to-mesenchymal transformation, ERα = estrogen receptor α, HN = healthy normal, PR = progesterone receptor, TAM = tamoxifen, TNBC = triple negative breast cancer, TAM = tamoxifen.
| miRNA Increased in BrCa | Examples of Validated Target (s) | Pathway (s) | Comments |
|---|---|---|---|
| miR-7 | XIAP [120] | TRAIL-induced apoptosis [121] | Higher in DCIS than HN [122]; High in BCa tissues [56]; miR-7 expression was negatively correlated with the stage, grade, and survival of BCa patients [123]. |
| miR-10a-5p | ACTG1 [124]; BCL6 [125]; CHL1 [126]; NCOR2 [127]; MMP14 and SKA1 [128]; YAP1 [129]; lncRNAs TUSC7 and RP11-838N2.4 act as ceRNAs for miR-10a in HCC [130] and glioblastoma [131] cells | PI3K/AKT/mTOR pathway [132] | High expression was predictive of tumor relapse in TAM-treated ER+ postmenopausal BCa patients [133,134]; Low miR-10a-5p correlated with reduced relapse-free survival in BCa [16]. |
| miR-18a-5p | ESR1/ERα [135] SREBP1 [136] lncRNA UCA1 is a ceRNA miR-18a in BCa cells [137] | Higher in ERα-breast tumors [138]; higher in DCIS than HN [122]; Higher in metastatic BCa cells where it decreased ECAD (E-cadherrin) and increased EMT and metastasis of xenografted MDA-MB-231 TNBC cells [136]; | |
| miR-21-5p | PTEN, PDCD4 [106,139]; NFIB [122]; RASA1 and RASA2 [140]; BTG2, FBXO11, MARCKS, RECK, and TPM1 [141]; TIMP3 [115] miR-21 negatively regulates lncRNAs GAS5 and CASC2 [28] | PI3K-AKT signaling, apoptosis | Consistently increased in breast tumors and in plasma from BCa patients [56]. High in breast tumors; high miR-21 correlates with lymph node status and tumor stage [107]. A meta-analysis of serum/plasma miR-21 in 438 BCa patients and 228 healthy controls concluded that increased miR-21 is a potential biomarker for BCa with a sensitivity of 0.79 [142]. |
| miR-26a-1-3p | ESR1 [143]; CHD1, GREB1, and KPNA2 [144] CDC2, CCNE1 [145]; EZH2 [146] | Apoptosis in cancer cells [146] | Higher expression in primary breast tumors was associated with clinical benefit of tamoxifen [16]. |
| miR-29b-3p, miR-29c | DICER, TTP, PTEN, ARP1B1, KLF4, MYP, ANGPTL4, LOX, MMP, PDFGC, VEGFA, ADAM12, SERPINH1 (reviewed in [60,147]). LncRNAs: H19 targets miR-29b-3p [148,149]; TUG targets miR-29b and miR-29c [150]; XIST targets miR-29c [151,152]; MIAT targets miR-29c [153]. | Have both tumor suppressor and oncomiR roles [154] | Upregulated in BCa tissues [56]. |
| miR-30c-5p | CTGF [155]; BCL9 [156]; HOXA1 [157]; SRSF1 [158]; KRAS [159]; CHD7 and TNRC6A [160] lncRNA AK017368 acts as a ceRNA for miR-30c in skeletal muscle cells [161] | Proliferation, apoptosis, differentiation [162]. Oncogene-induced senescence: a key tumor-suppressing mechanism [160] | High miR-30c-5p in primary tumors associated with clinical benefit of tamoxifen treatment [16]. Low miR-30 family expression in breast tumors was associated with poor relapse-free survival and bone metastasis [163]. |
| miR-96-5p | MTOR and RPS6KB1 (also called S6K1) in TNBC [164] | Insulin signaling in non-small cell lung cander (NSCLC) [165] | Upregulated in BCa samples [166]. |
| miR-125b-5p | ERBB2, ERBB3 [167]; ETS1 [168] | Epidermal growth factor receptor (EGFR) signaling Resistance to TAM and AIs [59] | In ER+/PR+ patients, high miR-125b-5p correlated with earlier relapse [134,169]. Also increased in blood plasma from BCa patients [170,171]. |
| miR-134-5p | EGFR [172], FOXM1, KRAS, STAT4B, ERBB2 | Cell proliferation, apoptosis, invasion, metastasis, drug resistance; however, it also acts as a tumor suppressor miRNA by targeting STAT4B, KRAS, and the Notch signaling pathway [173]. FOXM1 increases transcription of genes for G1/S transition, promotes CSCs and endocrine resistance [174]. | Increased in circulating plasma from BCa patients [175]. Encoded in the DLK1-DIO3 genomic region, located on 14q32 that contains the paternally expressed imprinted genes DLK1, RTL1, and DIO3, and the maternally expressed imprinted genes MEG3, MEG8, and as RTL1, two lncRNAs, and 53 miRNAs [176]. FOXM1 transcriptionally increased E2–ERα in BCa cells, and regulation is reciprocal [177]. |
| miR-155-5p | TERF1 [178]. TP53INP1 [179]; ZEB2 [180]; lncRNA H19 [181] lncRNAs acting as ceRNA for miR-155 include CCAT1 [182]; MALAT1 [183], MIAT [184], and CCAT1 [185] in cancer cells | EMT and metastasis [180] | Upregulated in BCa tissues [56]; higher in the serum of BCa patients than healthy women [186]. |
| miR-181b-5p | CCND1, CBX7, BCL2, HMGA2, TP53 [187]; DAX1 [141] lncRNA CCAT1 acts as a ceRNA for miR-181b in glioma cells [188] | Growth factor signaling [59] | Higher in DCIS than HN [122]; levels of miR-181b decline in serum after surgical removal of breast tumors [189]. |
| miR-181b-3p | YWHAG [190] | EMT [190] | Higher in metastatic BCa lines versus MCF-7 and T47D [190] |
| miR-185-5p | VEGFA [191]; E2F6 and DNMT1 [192] | Upregulated by tumor and metastasis suppressor PEBP1 (also called RKIP) in BCa cells [193]; apoptosis [194]; | Reduced in BCa tissues [193]. |
| miR-193a-3p | DDAH1 [195]; WT1 [196] | Cell growth [196] miR-193a-5p was decreased, but no significant differences in miR-193a-3p in BCa [197]. | Highly expressed in breast tumors [198]; downregulated in BCa tumors [196] |
| miR-210-3p | ISCU and COX10 [199]; FGFRL1, RAD52; BDNF, PTPN1, ISCU, NCAM1, and the lncRNA XIST [200] | Cell proliferation, migration, and invasion | High miR-210 was associated with lower relapse-free survival [201]. Induced by hypoxia [202] Increased in cell line models of aromatase resistance [172]. Consistently increased in breast tumors and plasma from BCa patients [56]. |
| miR-221-3p | ESR1/ERα [203], CDKN1B, FOXO3, KIT, TIMP3, BRAP, ARIH2, FOS, ICAM1 | ERα regulation of gene transcription in BCa [204,205]. Both miR-221 and miR-222 are involved in regulating adherens junction, PI3K and MAPK signaling, transforming growth factor β (TGFβ)signaling, apoptosis, and cell cycle [206]. | Increased in tumors of patients who develop tamoxifen resistance [207]. |
| miR-222-3p | ESR1/ERα [203], TIMP3 [208], STAT5A [209], MMP1 [210], FOXO3, FOX, PTEN, KIT, SOCS1 and CDKN1B [211] | ERα regulation of gene transcription in BCa [204]. Promotes S-phase entry, EMT, and TAM resistance [172]. | Increased in breast tumors [212]. High expression of miR-222 was associated with short relapse-free time in ER+/PR+ BCa patients [134]. |
| miR-324-5p | SMO [213] | Suppressed invasion of MDA-MB-231 cells [214] | Higher in DCIS than HN [122]; |
| miR-365 | BCL2 [215]; SHC1 and BAX [216] | Higher circulating levels in plasma predicted decreased OS in metastatic BCa patients [217]. | Higher in DCIS than HN [122]; |
| miR-519a-3p | CDKN1A, RB1, and PTEN [218]; TNFRSF10B (TRAIL-R2) and CASP8 [219] | TRAIL-induced apoptosis [219]. | High expression correlated with lower disease free survival in ER+ patients, not ER- patients [218]. TAM resistance in MCF-7 cells: transient transfection of MCF-7 cells with a miR-519a mimic resulted in TAM resistance; conversely, the transfection of TAM-resistant MCF-7 cells with a miR-519a inhibitor restored TAM growth inhibition [218]. |
Table 3.
Examples of miRNAs downregulated in breast tumors, patient plasma, and breast cancer cell lines with their authenticated targets. miRTarBase was used to identify validated targets not identified by reference number.
Table 3.
Examples of miRNAs downregulated in breast tumors, patient plasma, and breast cancer cell lines with their authenticated targets. miRTarBase was used to identify validated targets not identified by reference number.
| miRNA Decreased in BrCa | Examples of Validated Target (s) | Pathway (s) | Comments |
|---|---|---|---|
| miR-99a-5p | IGF1R, AKT1 [226]; HOXA1 [227]; RAVER2, FGFR2, IGF1R, MTOR, AGO2 | TGFβ pathway [228]; mTOR signaling pathway [229] | Downregulated in BCa tissues [56]; downregulated in DCIS [230]; low serum miR-99a is a poor prognostic indicator in BCa correlating with lymph node metastasis, and distant metastasis [231]. |
| miR-125a-5p | CDKN1, NTRK3, TP53, VEGFA, ERBB2, ERBB3, BAK1, KLF13, ARID3B, ELAV1 lncRNAs that act as ceRNAs for miR-125a-5p include HOTAIR, ANRIL, and HOXA11-AS (reviewed in [28]) | NFκB pathway [232] | Lower in breast tumors [233]. |
| miR-127 | PRDM1 [234] lncRNA MEG3 acts as a ceRNA for miR-127 in osteosarcoma cells [235] | p53 transactivates miR-127 leading to the inhibition of MMP13 translation, whereas c-Jun (activated by TGFβ) inhibits miR-127 transcription [236] | Lower in DCIS than histologically normal tissue [122]. |
| miR-139-5p | POLQ, TOP1, TOP2A, RAD54L, and XRCC5 [237]; RUNX1 [238] | EMT [239] | miR-139-5p was downregulated in BCa tissues [56]. |
| miR-143 | MAPK1 [240]; DNMT3A [241]; CIAPIN1 [242]; BCL2 [243]; | RAS signaling in basal-like BCa [244]. | Downregulated in BCa tissues [56]. |
| miR-145 | RASA1, MEKK, EGFR [245]; TGFNR2 and SMAD3 [246] lncRNAs act as ceRNAs for miR-145-5p: TUG1 and MALAT1 [246,247]; CRNDE in gastric cancer cells [248], LINC-ROR in gastric cancer cells [249], and PCAT1 in prostate cancer cells [250] | RAS signaling in basal-like BCa [244]. TEAD-YAP transcriptional pathway, including VEGFA and ITGB4 [251]. | Lower in breast tumors [233]; downregulated in BCa tissues [56]. |
| miR-193a-5p | ERBB2 (HER2) [252]; WT1, SRSF2, HIC2, HOXC9, PSEN1, LOXL4, ING5, c-KIT, PLAU, and MCL1 [196] | Downregulated in DCIS [230]. | |
| miR-378a-3p | GLI3 [253]; GOLT1A [254] | Downregulated in DCIS [230]. | |
| miR-497-5p | BCL2 and CCND1 [255]; KCNN4 [256] | Downregulated by methylation in breast tumors [257]; Downregulated in DCIS [230]. | |
| miR-652-3p | ZEB1 [258] | EMT [259] | Downregulated in DCIS [230] Lower in serum of BCa patients [260]. |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Klinge, C.M. Non-Coding RNAs in Breast Cancer: Intracellular and Intercellular Communication. Non-Coding RNA 2018, 4, 40. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna4040040
AMA Style
Klinge CM. Non-Coding RNAs in Breast Cancer: Intracellular and Intercellular Communication. Non-Coding RNA. 2018; 4(4):40. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna4040040
Chicago/Turabian StyleKlinge, Carolyn M. 2018. "Non-Coding RNAs in Breast Cancer: Intracellular and Intercellular Communication" Non-Coding RNA 4, no. 4: 40. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna4040040
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.