
The Functional Roles and Regulation of Circular RNAs during Cellular Stresses
by
 , , and
, , and
Yueh-Chun Lee
1,2,† ,
,
Wei-Yu Wang
3,†,
Hui-Hsuan Lin
4,
Yi-Ren Huang
5,
Ya-Chi Lin
6,7 and
Kuei-Yang Hsiao
4,5,8,9,10,*
1
Department of Radiation Oncology, Chung Shan Medical University Hospital, Taichung 40201, Taiwan
2
School of Medicine, Chung Shan Medical University, Taichung 40201, Taiwan
3
Division of Hemato-Oncology, Department of Internal Medicine, Ditmanson Medical Foundation Chia-Yi Christian Hospital, Chia-Yi City 60002, Taiwan
4
Ph.D. Program in Tissue Engineering and Regenerative Medicine, National Chung Hsing University, Taichung 40227, Taiwan
5
Institute of Biochemistry, College of Life Sciences, National Chung Hsing University, Taichung 40227, Taiwan
6
Department of Plant Pathology, College of Agriculture and Natural Resources, National Chung Hsing University, Taichung 40227, Taiwan
7
Department of Medical Laboratory Science and Biotechnology, Asia University, Taichung 41354, Taiwan
8
Ph.D. Program in Translational Medicine, College of Life Sciences, National Chung Hsing University, Taichung 40227, Taiwan
9
Rong Hsing Research Center for Translational Medicine, College of Life Sciences, National Chung Hsing University, Taichung 40227, Taiwan
10
Bachelor Program of Biotechnology, College of Agriculture and Natural Resources, National Chung Hsing University, Taichung 40227, Taiwan
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Non-Coding RNA 2022, 8(3), 38; https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna8030038
Submission received: 11 April 2022
/
Revised: 19 May 2022
/
Accepted: 26 May 2022
/
Published: 27 May 2022
(This article belongs to the Special Issue circRNAs in Cell and Organ Development)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Circular RNAs (circRNAs) are a novel class of regulatory RNA involved in many biological, physiological and pathological processes by functioning as a molecular sponge, transcriptional/epigenetic/splicing regulator, modulator of protein–protein interactions, and a template for encoding proteins. Cells are constantly dealing with stimuli from the microenvironment, and proper responses rely on both the precise control of gene expression networks and protein–protein interactions at the molecular level. The critical roles of circRNAs in the regulation of these processes have been heavily studied in the past decades. However, how the microenvironmental stimulation controls the circRNA biogenesis, cellular shuttling, translation efficiency and degradation globally and/or individually remains largely uncharacterized. In this review, how the impact of major microenvironmental stresses on the known transcription factors, splicing modulators and epitranscriptomic regulators, and thereby how they may contribute to the regulation of circRNAs, is discussed. These lines of evidence will provide new insight into how the biogenesis and functions of circRNA can be precisely controlled and targeted for treating human diseases.
Keywords:
circRNA; backsplicing; DNA damage response; genotoxic stress; chemoresistance; hypoxia; heat shock; m6A1. Introduction
Circular RNA is a class of regulatory RNA with a circular configuration, and is produced by backsplicing, in which the downstream splice donor reacts with the upstream splice acceptor by canonical spliceosome machinery. Since its first discovery in mammalian cells in 1979 [1] and the rediscovery of its molecular function and wide distribution in various tissues in 2012 [2,3,4,5], great attention has been paid to the molecular functions of circRNA, such as the microRNA (miRNA)/RNA-binding protein (RBP) sponge [4,6,7,8], transcriptional/epigenetic regulator [9,10], splicing regulator [11,12], modulator for protein–protein interactions [13,14,15] and templates for encoding proteins [16,17,18].
It is known that circRNAs are transcribed from both the exonic and intronic regions of coding or noncoding genes [19,20,21,22]. Several genome-wide analyses reported that circRNAs originate from virtually any genomic locus, such as intergenic, intronic, coding region, 5′- and 3′-untranslational regions (UTRs) [23,24]. Despite the diverse loci/regions of origin, the majority of circRNAs are produced by the unique process termed ‘backsplicing’, in which a downstream splice donor joins an upstream splice acceptor with no preference for U2 or U12 spliceosome [5,25]. However, there are a few molecular properties/events favoring the occurrence of backsplicing. First, the exons flanked by large introns tend to be circularized [2,5,26,27] (Figure 1, top). Second, the repetitive sequences or any complementary sequences in the flanking introns would bring the downstream splice donor and upstream splice acceptor to a proximity, allowing the backsplicing to take place [5,9,28,29] (Figure 1, left). In an analogy, RBPs antagonizing complementary pairing hinder the efficiency of backsplicing, while RBPs favoring the interaction between flanking introns enhance backsplicing [30] (Figure 1, middle). Lastly, the lost/masking of the splice donor of the upstream flanking intron or splice acceptor of the downstream flanking intron also increases the frequency of backsplicing (Figure 1, right). The cells should respond to the external stimulation properly, and deregulated responses to cellular stresses typically cause the development of human diseases. In this mini-review, we would like to focus on the potential mechanisms underlying how cells modulate the process of backsplicing in response to or during the cellular stresses. The presentation of these lines of clues would help to bridge the external stimuli, backsplicing, and finally the biogenesis and functions of circRNAs, shedding light on the identification of potential targets for treating human diseases.
2. Roles of CircRNAs in Cellular Stress
2.1. Roles and Regulation of CircRNAs in Response to Genotoxic Stress
DNA damage poses a significant threat to the cells, and thus it is critical for cells to properly respond to the DNA damage. One of the early molecular events of DNA damage is mediated by phosphatidylinositol 3-kinase-related kinase (PI3KK) activation. In addition to DNA repair, the activation of PI3KK coordinates global transcriptional repression [31,32,33,34]. DExH-Box helicase 9 (DHX9), also known as RNA helicase A, is associated with active transcription machinery and is involved in resolving the secondary structure during active transcription [35,36] (Figure 2A, left), while its helicase activity negatively regulates the formation of circRNA [36]. Interestingly, DHX9 also plays important roles during DNA damage response and is associated with factors related to DNA damage [37,38] (Figure 2A, top). It was shown that the depletion of DHX9 using small interfering RNA decreases the levels of a few circRNAs [36], and thus, factors inhibiting DHX9 activity may potentially facilitate circRNA backsplicing. Intriguingly, DHX9 was post-translationally regulated by DNA damage-induced ATM/ATR activation [39,40] (Figure 2A, top and right), and the phosphorylation sites located adjacent to the RNA binding motif. This suppressive phosphorylation promotes the level of an oncogenic circRNA, CCDC66, which governs a subset of oncogenes contributing to the development of chemoresistance [7,40]. The aberrant induction of circRNAs is a feature of chemo-/radio-resistant colorectal cancer (CRC). It has been reported that a group of circRNAs was upregulated in fluorouracil/radio-resistant CRC consistent with the observation in oxaliplatin-resistant CRC [40,41]. In both platinum-based chemo-resistant gastrointestinal tract-derived tumors, the patients with higher levels of these circRNAs had a poorer prognosis, and the induction of the chemoresistance-regulated circRNAs contributed to the metastatic features of the tumors [7,40,42]. In agreement with the role of DHX9 in DNA damage response, DHX9 interacting partners, splicing factor proline/glutamine rich (SFPQ) and non-POU domain-containing octamer binding protein (NONO), were also recruited to the DNA damage site in an ATM-dependent manner [43] (Figure 2B). Functionally, SFPQ hinders the activity of the cryptic splicing signal, regulating the availability of the splicing signal for backsplicing [44]. The change of the subnuclear localization of SFPQ and NONO may alter their availability for regulating splicing. It has been reported that the absence/unavailability of SFPQ in the upstream intron of circularizable exons promoted the backsplicing efficiency of a particular circRNA with a long intron, but no proximal inverted Alu elements [44], implying that SFPQ may be involved in the regulation of genotoxic stress-induced circRNA biogenesis.
The exact roles of circRNA have not been thoroughly explored. However, some pioneer studies of noncoding RNA may point out the direction. For example, a group of small non-coding RNAs, termed DNA damage-response RNA (DDRNA), were produced by DICER and DROSHA in response to DNA damage, and contributed to the accumulation of MDC1 and 53BP1 in the late stage of the DNA damage response [45]. lncRNA in nonhomologous end joining (NHEJ) pathway 1 (LINP1) served as a platform for Ku80 and DNA-dependent protein kinase catalytic subunits (DNA-PKcs), promoting the activity of NHEJ [46]. In contrast, small Cajal body-specific RNA 2 (scaRNA2) constrained the activity of DNA-PK through binding to the catalytic subunit, and thus, scaRNA2 weakens its interaction with the Ku70/80 subunits, as well as with the LINP1 lncRNA [47]. Whether circRNA is involved in this fine regulation warrants further investigation.
2.2. Modulation of CircRNA under Hypoxic Stress
2.2.1. Hypoxia-Regulated CircRNAs
Hypoxia, a condition in which cells are deprived of an adequate oxygen supply, is one of the most challenging stresses to tissues, and plays vital roles in both physiological and pathological processes such as tumorigenesis and diseases in various tissues/organs [48,49,50,51,52]. The post-translational and transcriptional regulation of hypoxia-inducible factor (HIF) have been extensively investigated [53], but the roles of circRNA in response to hypoxia is yet to be explored. Hypoxia-regulated circRNAs were identified through transcriptomic analyses in a handful pioneer studies [54,55]. Several hypoxia-induced circR-
NAs such as cireZNF292, circAFF1 and circDENND4C have been identified (Figure 3, left). The upregulation of these genes is prone to result from the transcriptional activation, but not the alteration of backsplicing efficiency. Among these circRNAs, circZNF292 was shown to promote endothelial cell proliferation and sprouting through a non-miRNA sponging activity. An independent study using models of breast cancer also reported that the level of circDENND4C was upregulated by hypoxic stress, and was positively correlated to the HIF-1α mRNA level and tumor size [28], implying the importance of transcriptional activation by HIF-1α under hypoxia for circDENND4C induction. In spite of the uncharacterized molecular functions of these hypoxia-regulated circRNAs, a cardiac- necroptosis-associated circRNA (CNEACR) originating from exon 2 to 5 of Fbxw4 was identified from a mouse model of ischemia/reperfusion [56] (Figure 3, right). The level of CNEACR was downregulated by ischemia/reperfusion in mice, and the same pattern was confirmed in the cell culture system. The expression of CNEACR was specific to myocytes compared to fibroblasts, and resided more in cytoplasm than nuclei. The overexpression of CNEAR attenuated hypoxia/reoxygenation-induced cell death. It was observed that CNEAR interacts with histone deacetylase 7 (HDAC7) by using biotin pulldown assay and trapping HDAC7 in cytoplasm. Mechanically, the trapping of HDAC7, a transcriptional co-repressor, in cytoplasm relieved the suppression of Foxa2 and facilitated the induction of Foxa2-regulated receptor-interacting protein kinase 3 (RIPK3). This particularly widened the horizon regarding how circRNA may function in addition to miRNA sponges in physiological and pathological conditions. Nevertheless, it has been reported that circRNA may be used for inter-cellular communication by incorporation into exosomes [57,58]. Among these exosomal circRNAs, the level of circZNF91 was elevated by HIF-1α-mediated transcriptional activation, and was shown to antagonize the activity of miR-23b-3p, protecting SIRT1 mRNA from degradation and contributing the development of gemcitabine resistance [59].
2.2.2. Players Regulate Function and Abundance of CircRNAs in Hypoxia
Although the mechanisms underlying hypoxia-regulated circRNA biogenesis and functions have not yet been fully explored, there are a few studies that have attempted to demonstrate potential mechanisms. A comprehensive transcriptomic study indicated that the binding motifs of heterogeneous nuclear ribonucleoprotein C (hnRNPC), human antigen R (HuR) and poly(A)-binding protein 4 are enriched in the flanking introns of hypoxia-regulated circRNAs [55]. Although none of these RBPs are readily linked to circRNA biogenesis, there are some clues linking the regulation of backsplicing and hypoxia. For example, the interaction of hnRNPC with DHX9 and competition with splicing factor U2AF 65 kDa subunit (U2AF65) in Alu element make hnRNPC a potential regulator for circRNA biogenesis under hypoxia [36,60]. The role of hnRNPC in controlling intron pairing and backsplicing is supported by a study showing that the knockdown of hnRNPC increased the abundance of double-stranded RNA regions [61]. This suppressive role of hnRNPC on double-stranded RNA formation may be partially contributed to by the ability of hnRNPC to interact with DHX9 [36]. In addition to resolving the RNA pairing, completion between hnRNPC and U2AF65 for U tract within Alu elements suppresses the splicing activity [60], and thus potentially inhibits backsplicing in the context of circRNA biogenesis. In addition, it has been reported that the level of HuR is modulated by hypoxic stress [62], and recognizes RNA motifs via the formation of homodimer [63], implying that the backsplicing efficiency may be potentially modulated by the hypoxia-regulated availability of HuR.
N6-methyladenosine (m6A), one of the most prevalent, abundant and conserved modifications identified in eukaryotic RNAs, was recently identified on circRNA [64,65] and serves as another link to hypoxia-regulated circRNA functions and homeostasis. The abundance of m6A is dynamically regulated by a dozen m6A methyltransferases, such as methyltransferase-like 3/14/16 [66,67,68], zinc finger CCCH-type containing 3 [69], RNA binding motif protein 15 [70], WT1-associated protein [71,72] and vir-like m6A methyltransferase-associated [72], and demethylases, fat mass and obesity-associated (FTO) and AlkB homolog 5 (ALKBH5) [73,74]. Different effectors/readers include YTH domain-containing family 1/2/3 (YTHDF1/2/3), YTH domain-containing 1/2 (YTHDC1/2), insulin-like growth factor 2 mRNA-binding protein 1/2/3 (IGF2BP1/2/3) and hnRNPA2B1 grant m6A multiple cellular/molecular functions. The deposition of m6A on circRNA differentially controls various molecular events such as nuclear export, degradation, biogenesis and translation by a distinct m6A reader [64,75,76,77,78]. For example, the deposition of m6A in exon followed by the recognition of YTHDC1 promotes the backsplicing of circZNF609 and nuclear export of circNSUN2 in nuclei, while recognition by YTHDF3 in cytoplasm increases translation [75,77] (Figure 4, top). Through recognition by a distinct m6A reader, YTHDF2, which works with heat-responsive protein 12 and RNase P, a subset of m6A-containg circRNAs, was destabilized [76]. Intriguingly, the expression of many m6A regulators was regulated by hypoxic stress. The expression of m6A demethylase, FTO, was suppressed by hypoxia [79], while the levels of ALKBH5, YTHDC1, YTHDF1 and YTHDF2 were upregulated [80,81,82,83,84] (Figure 4, indicated by (+) or (−)). Most of these cases were direct targets of the hypoxia-inducible factor, while YTHDF1 was post-transcription-ally regulated by hypoxia-induced miR-16-5p. These lines of evidence suggest that the hypoxia-regulated m6A dynamics may control functions of a subset of circRNAs. Along with the hypoxia-m6A-circRNA regulatory axis, circRNA also controls the level of HIF-1α. For example, it was reported that circERBIN promotes the expression of HIF-1α through upregulation of the cap-independent translation of HIF-1α by suppressing miRNAs targeting 4EBP-1 [85]. Taken together, this evidence highlights the critical roles of circRNA during hypoxic stress.
3. Heat Shock Stress
The heat shock response is well studied in aspects of both transcriptional and translational regulation. However, whether the functions or expression of circRNA are controlled under such circumstances remains largely unknown. Recently, the coding potential of circRNA has begun to receive more attention. One of the cap-independent translations of circRNA is mediated by m6A [77] (Figure 5). It has been reported that m6A reader—YTHDF2—translocates to the nuclei and protects the m6A within 5′-UTR of mRNA from FTO-mediated demethylation and promotes translation through the recruitment of eukaryotic initiation factor 3 (eIF3) in response to heat shock [86,87]. Consistent with the role of the m6A-stimulating cap-independent translation of mRNA, the m6A-mediated translation of circRNA is enhanced by heat shock [17,64], implying that YTHDF2 and/or eIF3 may be potential players for regulating the coding activity of circRNA under heat shock stress.
4. Closing Remarks
The regulation of circRNA under genotoxic stress, hypoxia and heat shock stress is discussed in the current article. Although many progressions have been made in the last decade since the re-discovery of circRNA regarding its functions and distribution in 2012 [2,3,4,5], the diverse mechanisms underlying the regulation of backsplicing still remain largely uncharacterized. In this mini-review, how the known effectors under three of the major cellular stresses potentially link to backsplicing modulators were outlined and discussed. These materials should prompt researchers to identify the current gaps in circRNA regulation, and hopefully stimulate the investigation for paving new roads for the development of novel strategies to treat human diseases.
Author Contributions
Conceptualization, K.-Y.H.; original draft preparation, Y.-C.L. (Yueh-Chun Lee), W.-Y.W., H.-H.L., Y.-R.H. and Y.-C.L. (Ya-Chi Lin); review and editing, Y.-C.L. (Yueh-Chun Lee), W.-Y.W., H.-H.L. and K.-Y.H.; visualization, Y.-C.L. (Ya-Chi Lin); supervision, K.-Y.H.; funding acquisition, Y.-C.L. (Yueh-Chun Lee), W.-Y.W. and K.-Y.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Ministry of Science and Technology of Taiwan (MOST 109-2320-B-005-008-MY3) to K.-Y.H., ‘Hsing Chung inter-institutional project’ (NCHU-CSMU-10805, NCHU-CSMU-10906 and NCHU-CSMU-11007) to Y.-C.L. (Yueh-Chun Lee) and K.-Y.H., and ‘Chia Hsing bilateral joint research project’ (CYCH-NCHULS110002) to W.-Y.W. and K.-Y.H.
Acknowledgments
H.-H.L. carried out her thesis research under the auspices of the Ph.D. program in Tissue Engineering and Regenerative Medicine, National Chung Hsing University and National Health Research Institutes in Taiwan.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hsu, M.T.; Coca-Prados, M. Electron microscopic evidence for the circular form of RNA in the cytoplasm of eukaryotic cells. Nature 1979, 280, 339–340. [Google Scholar] [CrossRef]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS ONE 2012, 7, e30733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Wang, Y.; Cao, M.; Pantaleo, V.; Burgyan, J.; Li, W.X.; Ding, S.W. Homology-independent discovery of replicating pathogenic circular RNAs by deep sequencing and a new computational algorithm. Proc. Natl. Acad. Sci. USA 2012, 109, 3938–3943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Bao, C.; Guo, W.; Li, S.; Chen, J.; Chen, B.; Luo, Y.; Lyu, D.; Li, Y.; Shi, G.; et al. Circular RNA profiling reveals an abundant circHIPK3 that regulates cell growth by sponging multiple miRNAs. Nat. Commun. 2016, 7, 11215. [Google Scholar] [CrossRef]
- Hsiao, K.Y.; Lin, Y.C.; Gupta, S.K.; Chang, N.; Yen, L.; Sun, H.S.; Tsai, S.J. Noncoding Effects of Circular RNA CCDC66 Promote Colon Cancer Growth and Metastasis. Cancer Res. 2017, 77, 2339–2350. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.C.; Lee, Y.C.; Chang, K.L.; Hsiao, K.Y. Analysis of common targets for circular RNAs. BMC Bioinform. 2019, 20, 372. [Google Scholar] [CrossRef]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Chen, N.; Zhao, G.; Yan, X.; Lv, Z.; Yin, H.; Zhang, S.; Song, W.; Li, X.; Li, L.; Du, Z.; et al. A novel FLI1 exonic circular RNA promotes metastasis in breast cancer by coordinately regulating TET1 and DNMT1. Genome. Biol. 2018, 19, 218. [Google Scholar] [CrossRef] [Green Version]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. circRNA biogenesis competes with pre-mRNA splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conn, V.M.; Hugouvieux, V.; Nayak, A.; Conos, S.A.; Capovilla, G.; Cildir, G.; Jourdain, A.; Tergaonkar, V.; Schmid, M.; Zubieta, C.; et al. A circRNA from SEPALLATA3 regulates splicing of its cognate mRNA through R-loop formation. Nat. Plants 2017, 3, 17053. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Liu, B.; Shen, D.D.; Yan, P.; Zhang, Y.; Tian, Y.; Hou, L.; Jiang, G.; Zhu, Y.; Liang, Y.; et al. A tumor-suppressive circular RNA mediates uncanonical integrin degradation by the proteasome in liver cancer. Sci. Adv. 2021, 7, eabe5043. [Google Scholar] [CrossRef]
- Du, W.W.; Fang, L.; Yang, W.; Wu, N.; Awan, F.M.; Yang, Z.; Yang, B.B. Induction of tumor apoptosis through a circular RNA enhancing Foxo3 activity. Cell Death Differ. 2017, 24, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Du, W.W.; Yang, W.; Liu, E.; Yang, Z.; Dhaliwal, P.; Yang, B.B. Foxo3 circular RNA retards cell cycle progression via forming ternary complexes with p21 and CDK2. Nucleic Acids Res. 2016, 44, 2846–2858. [Google Scholar] [CrossRef] [Green Version]
- Li, X.F.; Lytton, J. A circularized sodium-calcium exchanger exon 2 transcript. J. Biol. Chem. 1999, 274, 8153–8160. [Google Scholar] [CrossRef] [Green Version]
- Legnini, I.; Di Timoteo, G.; Rossi, F.; Morlando, M.; Briganti, F.; Sthandier, O.; Fatica, A.; Santini, T.; Andronache, A.; Wade, M.; et al. Circ-ZNF609 Is a Circular RNA that Can Be Translated and Functions in Myogenesis. Mol. Cell 2017, 66, 22–37.e9. [Google Scholar] [CrossRef] [Green Version]
- Pamudurti, N.R.; Bartok, O.; Jens, M.; Ashwal-Fluss, R.; Stottmeister, C.; Ruhe, L.; Hanan, M.; Wyler, E.; Perez-Hernandez, D.; Ramberger, E.; et al. Translation of CircRNAs. Mol. Cell 2017, 66, 9–21.e7. [Google Scholar] [CrossRef] [Green Version]
- Halbreich, A.; Pajot, P.; Foucher, M.; Grandchamp, C.; Slonimski, P. A pathway of cytochrome b mRNA processing in yeast mitochondria: Specific splicing steps and an intron-derived circular RNA. Cell 1980, 19, 321–329. [Google Scholar] [CrossRef]
- Pasman, Z.; Been, M.D.; Garcia-Blanco, M.A. Exon circularization in mammalian nuclear extracts. RNA 1996, 2, 603–610. [Google Scholar]
- Braun, S.; Domdey, H.; Wiebauer, K. Inverse splicing of a discontinuous pre-mRNA intron generates a circular exon in a HeLa cell nuclear extract. Nucleic Acids Res. 1996, 24, 4152–4157. [Google Scholar] [CrossRef] [PubMed]
- Capel, B.; Swain, A.; Nicolis, S.; Hacker, A.; Walter, M.; Koopman, P.; Goodfellow, P.; Lovell-Badge, R. Circular transcripts of the testis-determining gene Sry in adult mouse testis. Cell 1993, 73, 1019–1030. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.U.; Agarwal, V.; Guo, H.; Bartel, D.P. Expanded identification and characterization of mammalian circular RNAs. Genome Biol. 2014, 15, 409. [Google Scholar] [CrossRef] [PubMed]
- Cocquerelle, C.; Daubersies, P.; Majerus, M.A.; Kerckaert, J.P.; Bailleul, B. Splicing with inverted order of exons occurs proximal to large introns. EMBO J. 1992, 11, 1095–1098. [Google Scholar] [CrossRef] [PubMed]
- Westholm, J.; Miura, P.; Olson, S.; Shenker, S.; Joseph, B.; Sanfilippo, P.; Celniker, S.E.; Graveley, B.; Lai, E.C. Genome-wide Analysis of Drosophila Circular RNAs Reveals Their Structural and Sequence Properties and Age-Dependent Neural Accumulation. Cell Rep. 2014, 9, 1966–1980. [Google Scholar] [CrossRef] [Green Version]
- Liang, D.; Wilusz, J.E. Short intronic repeat sequences facilitate circular RNA production. Genes Dev 2014, 28, 2233–2247. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.O.; Wang, H.B.; Zhang, Y.; Lu, X.; Chen, L.L.; Yang, L. Complementary sequence-mediated exon circularization. Cell 2014, 159, 134–147. [Google Scholar] [CrossRef] [Green Version]
- Das, A.; Sinha, T.; Shyamal, S.; Panda, A.C. Emerging Role of Circular RNA-Protein Interactions. Noncoding RNA 2021, 7, 48. [Google Scholar] [CrossRef]
- Hsiao, K.Y.; Mizzen, C.A. Histone H4 deacetylation facilitates 53BP1 DNA damage signaling and double-strand break repair. J. Mol. Cell Biol. 2013, 5, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Munoz, M.J.; Perez Santangelo, M.S.; Paronetto, M.P.; de la Mata, M.; Pelisch, F.; Boireau, S.; Glover-Cutter, K.; Ben-Dov, C.; Blaustein, M.; Lozano, J.J.; et al. DNA damage regulates alternative splicing through inhibition of RNA polymerase II elongation. Cell 2009, 137, 708–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pankotai, T.; Bonhomme, C.; Chen, D.; Soutoglou, E. DNAPKcs-dependent arrest of RNA polymerase II transcription in the presence of DNA breaks. Nat. Struct. Mol. Biol. 2012, 19, 276–282. [Google Scholar] [CrossRef] [Green Version]
- Shanbhag, N.M.; Rafalska-Metcalf, I.U.; Balane-Bolivar, C.; Janicki, S.M.; Greenberg, R.A. ATM-dependent chromatin changes silence transcription in cis to DNA double-strand breaks. Cell 2010, 141, 970–981. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, T.; Uchida, C.; Anderson, S.F.; Lee, C.G.; Hurwitz, J.; Parvin, J.D.; Montminy, M. RNA helicase A mediates association of CBP with RNA polymerase II. Cell 1997, 90, 1107–1112. [Google Scholar] [CrossRef] [Green Version]
- Aktas, T.; Avsar Ilik, I.; Maticzka, D.; Bhardwaj, V.; Pessoa Rodrigues, C.; Mittler, G.; Manke, T.; Backofen, R.; Akhtar, A. DHX9 suppresses RNA processing defects originating from the Alu invasion of the human genome. Nature 2017, 544, 115–119. [Google Scholar] [CrossRef]
- Zhang, S.; Hemmerich, P.; Grosse, F. Werner syndrome helicase (WRN), nuclear DNA helicase II (NDH II) and histone gammaH2AX are localized to the centrosome. Cell Biol. Int. 2007, 31, 1109–1121. [Google Scholar] [CrossRef]
- Anderson, S.F.; Schlegel, B.P.; Nakajima, T.; Wolpin, E.S.; Parvin, J.D. BRCA1 protein is linked to the RNA polymerase II holoenzyme complex via RNA helicase A. Nat. Genet. 1998, 19, 254–256. [Google Scholar] [CrossRef]
- Matsuoka, S.; Ballif, B.A.; Smogorzewska, A.; McDonald, E.R., 3rd; Hurov, K.E.; Luo, J.; Bakalarski, C.E.; Zhao, Z.; Solimini, N.; Lerenthal, Y.; et al. ATM and ATR substrate analysis reveals extensive protein networks responsive to DNA damage. Science 2007, 316, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.C.; Yu, Y.S.; Lin, H.H.; Hsiao, K.Y. Oxaliplatin-induced DHX9 phosphorylation promotes oncogenic circular RNA CCDC66 expression and development of chemoresistance. Cancers 2020, 12, 697. [Google Scholar] [CrossRef] [Green Version]
- Xiong, W.; Ai, Y.Q.; Li, Y.F.; Ye, Q.; Chen, Z.T.; Qin, J.Y.; Liu, Q.Y.; Wang, H.; Ju, Y.H.; Li, W.H.; et al. Microarray Analysis of Circular RNA Expression Profile Associated with 5-Fluorouracil-Based Chemoradiation Resistance in Colorectal Cancer Cells. Biomed Res. Int. 2017, 2017, 8421614. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Li, Z.; Zhang, Q.; Wang, W.; Li, B.; Wang, L.; Xu, Z.; Zeng, A.; Zhang, X.; Zhang, X.; et al. Circular RNA AKT3 upregulates PIK3R1 to enhance cisplatin resistance in gastric cancer via miR-198 suppression. Mol. Cancer 2019, 18, 71. [Google Scholar] [CrossRef] [PubMed]
- Salton, M.; Lerenthal, Y.; Wang, S.Y.; Chen, D.J.; Shiloh, Y. Involvement of Matrin 3 and SFPQ/NONO in the DNA damage response. Cell Cycle 2010, 9, 1568–1576. [Google Scholar] [CrossRef] [Green Version]
- Stagsted, L.V.W.; O’Leary, E.T.; Ebbesen, K.K.; Hansen, T.B. The RNA-binding protein SFPQ preserves long-intron splicing and regulates circRNA biogenesis in mammals. Elife 2021, 10, e63088. [Google Scholar] [CrossRef] [PubMed]
- Francia, S.; Cabrini, M.; Matti, V.; Oldani, A.; d’Adda di Fagagna, F. DICER, DROSHA and DNA damage response RNAs are necessary for the secondary recruitment of DNA damage response factors. J. Cell Sci. 2016, 129, 1468–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; He, Q.; Hu, Z.; Feng, Y.; Fan, L.; Tang, Z.; Yuan, J.; Shan, W.; Li, C.; Hu, X.; et al. Long noncoding RNA LINP1 regulates repair of DNA double-strand breaks in triple-negative breast cancer. Nat. Struct. Mol. Biol. 2016, 23, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Bergstrand, S.; O’Brien, E.M.; Coucoravas, C.; Hrossova, D.; Peirasmaki, D.; Schmidli, S.; Dhanjal, S.; Pederiva, C.; Siggens, L.; Mortusewicz, O.; et al. Small Cajal body-associated RNA 2 (scaRNA2) regulates DNA repair pathway choice by inhibiting DNA-PK. Nat. Commun. 2022, 13, 1015. [Google Scholar] [CrossRef]
- Marsch, E.; Sluimer, J.C.; Daemen, M.J. Hypoxia in atherosclerosis and inflammation. Curr. Opin. Lipidol. 2013, 24, 393–400. [Google Scholar] [CrossRef]
- Maes, C.; Carmeliet, G.; Schipani, E. Hypoxia-driven pathways in bone development, regeneration and disease. Nat. Rev. Rheumatol. 2012, 8, 358–366. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.H.; Hsiao, K.Y.; Tsai, S.J. Hypoxia: The force of endometriosis. J. Obstet. Gynaecol. Res. 2019, 45, 532–541. [Google Scholar] [CrossRef]
- Lin, S.C.; Hsiao, K.Y.; Chang, N.; Hou, P.C.; Tsai, S.J. Loss of dual-specificity phosphatase-2 promotes angiogenesis and metastasis via up-regulation of interleukin-8 in colon cancer. J. Pathol. 2017, 241, 638–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, K.Y.; Chang, N.; Tsai, J.L.; Lin, S.C.; Tsai, S.J.; Wu, M.H. Hypoxia-inhibited DUSP2 expression promotes IL-6/STAT3 signaling in endometriosis. Am. J. Reprod. Immunol. 2017, 78, e12690. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. The Genomics and Genetics of Oxygen Homeostasis. Annu. Rev. Genom. Hum. Genet. 2020, 21, 183–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boeckel, J.N.; Jae, N.; Heumuller, A.W.; Chen, W.; Boon, R.A.; Stellos, K.; Zeiher, A.M.; John, D.; Uchida, S.; Dimmeler, S. Identification and Characterization of Hypoxia-Regulated Endothelial Circular RNA. Circ. Res. 2015, 117, 884–890. [Google Scholar] [CrossRef] [Green Version]
- Di Liddo, A.; de Oliveira Freitas Machado, C.; Fischer, S.; Ebersberger, S.; Heumuller, A.W.; Weigand, J.E.; Muller-McNicoll, M.; Zarnack, K. A combined computational pipeline to detect circular RNAs in human cancer cells under hypoxic stress. J. Mol. Cell Biol. 2019, 11, 829–844. [Google Scholar] [CrossRef]
- Gao, X.Q.; Liu, C.Y.; Zhang, Y.H.; Wang, Y.H.; Zhou, L.Y.; Li, X.M.; Wang, K.; Chen, X.Z.; Wang, T.; Ju, J.; et al. The circRNA CNEACR regulates necroptosis of cardiomyocytes through Foxa2 suppression. Cell Death Differ. 2022, 29, 527–539. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, Q.; Bao, C.; Li, S.; Guo, W.; Zhao, J.; Chen, D.; Gu, J.; He, X.; Huang, S. Circular RNA is enriched and stable in exosomes: A promising biomarker for cancer diagnosis. Cell Res. 2015, 25, 981–984. [Google Scholar] [CrossRef] [Green Version]
- Fanale, D.; Taverna, S.; Russo, A.; Bazan, V. Circular RNA in Exosomes. Adv. Exp. Med. Biol. 2018, 1087, 109–117. [Google Scholar]
- Zeng, Z.; Zhao, Y.; Chen, Q.; Zhu, S.; Niu, Y.; Ye, Z.; Hu, P.; Chen, D.; Xu, P.; Chen, J.; et al. Hypoxic exosomal HIF-1alpha-stabilizing circZNF91 promotes chemoresistance of normoxic pancreatic cancer cells via enhancing glycolysis. Oncogene 2021, 40, 5505–5517. [Google Scholar] [CrossRef]
- Zarnack, K.; Konig, J.; Tajnik, M.; Martincorena, I.; Eustermann, S.; Stevant, I.; Reyes, A.; Anders, S.; Luscombe, N.M.; Ule, J. Direct competition between hnRNP C and U2AF65 protects the transcriptome from the exonization of Alu elements. Cell 2013, 152, 453–466. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zhao, W.; Liu, Y.; Tan, X.; Li, X.; Zou, Q.; Xiao, Z.; Xu, H.; Wang, Y.; Yang, X. Function of HNRNPC in breast cancer cells by controlling the dsRNA-induced interferon response. EMBO J. 2018, 37, e99017. [Google Scholar] [CrossRef]
- Hsiao, K.Y.; Wu, M.H.; Chang, N.; Yang, S.H.; Wu, C.W.; Sun, H.S.; Tsai, S.J. Coordination of AUF1 and miR-148a destabilizes DNA methyltransferase 1 mRNA under hypoxia in endometriosis. Mol. Hum. Reprod. 2015, 21, 894–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ripin, N.; Boudet, J.; Duszczyk, M.M.; Hinniger, A.; Faller, M.; Krepl, M.; Gadi, A.; Schneider, R.J.; Sponer, J.; Meisner-Kober, N.C.; et al. Molecular basis for AU-rich element recognition and dimerization by the HuR C-terminal RRM. Proc. Natl. Acad. Sci. USA 2019, 116, 2935–2944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Fan, X.; Mao, M.; Song, X.; Wu, P.; Zhang, Y.; Jin, Y.; Yang, Y.; Chen, L.L.; Wang, Y.; et al. Extensive translation of circular RNAs driven by N6-methyladenosine. Cell Res. 2017, 27, 626–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Molinie, B.; Daneshvar, K.; Pondick, J.V.; Wang, J.; Van Wittenberghe, N.; Xing, Y.; Giallourakis, C.C.; Mullen, A.C. Genome-Wide Maps of m6A circRNAs Identify Widespread and Cell-Type-Specific Methylation Patterns that Are Distinct from mRNAs. Cell Rep. 2017, 20, 2262–2276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokar, J.A.; Shambaugh, M.E.; Polayes, D.; Matera, A.G.; Rottman, F.M. Purification and cDNA cloning of the AdoMet-binding subunit of the human mRNA (N6-adenosine)-methyltransferase. RNA 1997, 3, 1233–1247. [Google Scholar]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat. Chem. Biol. 2014, 10, 93–95. [Google Scholar] [CrossRef] [Green Version]
- Pendleton, K.E.; Chen, B.; Liu, K.; Hunter, O.V.; Xie, Y.; Tu, B.P.; Conrad, N.K. The U6 snRNA m(6)A Methyltransferase METTL16 Regulates SAM Synthetase Intron Retention. Cell 2017, 169, 824–835.e14. [Google Scholar] [CrossRef] [Green Version]
- Wen, J.; Lv, R.; Ma, H.; Shen, H.; He, C.; Wang, J.; Jiao, F.; Liu, H.; Yang, P.; Tan, L.; et al. Zc3h13 Regulates Nuclear RNA m(6)A Methylation and Mouse Embryonic Stem Cell Self-Renewal. Mol. Cell 2018, 69, 1028–1038.e6. [Google Scholar] [CrossRef] [Green Version]
- Patil, D.P.; Chen, C.K.; Pickering, B.F.; Chow, A.; Jackson, C.; Guttman, M.; Jaffrey, S.R. m(6)A RNA methylation promotes XIST-mediated transcriptional repression. Nature 2016, 537, 369–373. [Google Scholar] [CrossRef]
- Ping, X.L.; Sun, B.F.; Wang, L.; Xiao, W.; Yang, X.; Wang, W.J.; Adhikari, S.; Shi, Y.; Lv, Y.; Chen, Y.S.; et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res. 2014, 24, 177–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, S.; Mumbach, M.R.; Jovanovic, M.; Wang, T.; Maciag, K.; Bushkin, G.G.; Mertins, P.; Ter-Ovanesyan, D.; Habib, N.; Cacchiarelli, D.; et al. Perturbation of m6A writers reveals two distinct classes of mRNA methylation at internal and 5’ sites. Cell Rep. 2014, 8, 284–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.G.; et al. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat. Chem. Biol. 2011, 7, 885–887. [Google Scholar] [CrossRef]
- Zheng, G.; Dahl, J.A.; Niu, Y.; Fedorcsak, P.; Huang, C.M.; Li, C.J.; Vagbo, C.B.; Shi, Y.; Wang, W.L.; Song, S.H.; et al. ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol. Cell 2013, 49, 18–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.X.; Chen, X.; Xia, L.P.; Zhang, J.X.; Pan, Z.Z.; Ma, X.D.; Han, K.; Chen, J.W.; Judde, J.G.; Deas, O.; et al. N(6)-methyladenosine modification of circNSUN2 facilitates cytoplasmic export and stabilizes HMGA2 to promote colorectal liver metastasis. Nat. Commun. 2019, 10, 4695. [Google Scholar] [CrossRef] [Green Version]
- Park, O.H.; Ha, H.; Lee, Y.; Boo, S.H.; Kwon, D.H.; Song, H.K.; Kim, Y.K. Endoribonucleolytic Cleavage of m(6)A-Containing RNAs by RNase P/MRP Complex. Mol. Cell 2019, 74, 494–507.e8. [Google Scholar] [CrossRef] [PubMed]
- Di Timoteo, G.; Dattilo, D.; Centron-Broco, A.; Colantoni, A.; Guarnacci, M.; Rossi, F.; Incarnato, D.; Oliviero, S.; Fatica, A.; Morlando, M.; et al. Modulation of circRNA Metabolism by m(6)A Modification. Cell Rep. 2020, 31, 107641. [Google Scholar] [CrossRef]
- Li, Y.; Chen, B.; Zhao, J.; Li, Q.; Chen, S.; Guo, T.; Li, Y.; Lai, H.; Chen, Z.; Meng, Z.; et al. HNRNPL Circularizes ARHGAP35 to Produce an Oncogenic Protein. Adv. Sci. 2021, 8, 2001701. [Google Scholar] [CrossRef]
- Ruan, D.Y.; Li, T.; Wang, Y.N.; Meng, Q.; Li, Y.; Yu, K.; Wang, M.; Lin, J.F.; Luo, L.Z.; Wang, D.S.; et al. FTO downregulation mediated by hypoxia facilitates colorectal cancer metastasis. Oncogene 2021, 40, 5168–5181. [Google Scholar] [CrossRef]
- Yao, X.; Li, W.; Li, L.; Li, M.; Zhao, Y.; Fang, Z.X.; Luo, Z. YTHDF1 upregulation mediates hypoxia-dependent breast cancer growth and metastasis through regulating PKM2 to affect glycolysis. Cell Death Dis. 2022, 13, 258. [Google Scholar] [CrossRef]
- Imai, Y.; Matsuo, N.; Ogawa, S.; Tohyama, M.; Takagi, T. Cloning of a gene, YT521, for a novel RNA splicing-related protein induced by hypoxia/reoxygenation. Mol. Brain Res. 1998, 53, 33–40. [Google Scholar] [CrossRef]
- Thalhammer, A.; Bencokova, Z.; Poole, R.; Loenarz, C.; Adam, J.; O’Flaherty, L.; Schodel, J.; Mole, D.; Giaslakiotis, K.; Schofield, C.J.; et al. Human AlkB homologue 5 is a nuclear 2-oxoglutarate dependent oxygenase and a direct target of hypoxia-inducible factor 1alpha (HIF-1alpha). PLoS ONE 2011, 6, e16210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craig, E.A.; Weber, J.D.; Spiegelman, V.S. Involvement of the mRNA binding protein CRD-BP in the regulation of metastatic melanoma cell proliferation and invasion by hypoxia. J. Cell Sci. 2012, 125, 5950–5954. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Shao, Y.L.; Wang, L.L.; Lin, J.; Zhang, J.B.; Ding, Y.; Gao, B.B.; Liu, D.H.; Gao, X.N. YTHDF2 is a potential target of AML1/ETO-HIF1alpha loop-mediated cell proliferation in t(8;21) AML. Oncogene 2021, 40, 3786–3798. [Google Scholar] [CrossRef]
- Chen, L.Y.; Wang, L.; Ren, Y.X.; Pang, Z.; Liu, Y.; Sun, X.D.; Tu, J.; Zhi, Z.; Qin, Y.; Sun, L.N.; et al. The circular RNA circ-ERBIN promotes growth and metastasis of colorectal cancer by miR-125a-5p and miR-138-5p/4EBP-1 mediated cap-independent HIF-1alpha translation. Mol. Cancer 2020, 19, 164. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wan, J.; Gao, X.; Zhang, X.; Jaffrey, S.R.; Qian, S.B. Dynamic m(6)A mRNA methylation directs translational control of heat shock response. Nature 2015, 526, 591–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, K.D.; Patil, D.P.; Zhou, J.; Zinoviev, A.; Skabkin, M.A.; Elemento, O.; Pestova, T.V.; Qian, S.B.; Jaffrey, S.R. 5’ UTR m(6)A Promotes Cap-Independent Translation. Cell 2015, 163, 999–1010. [Google Scholar] [CrossRef] [Green Version]
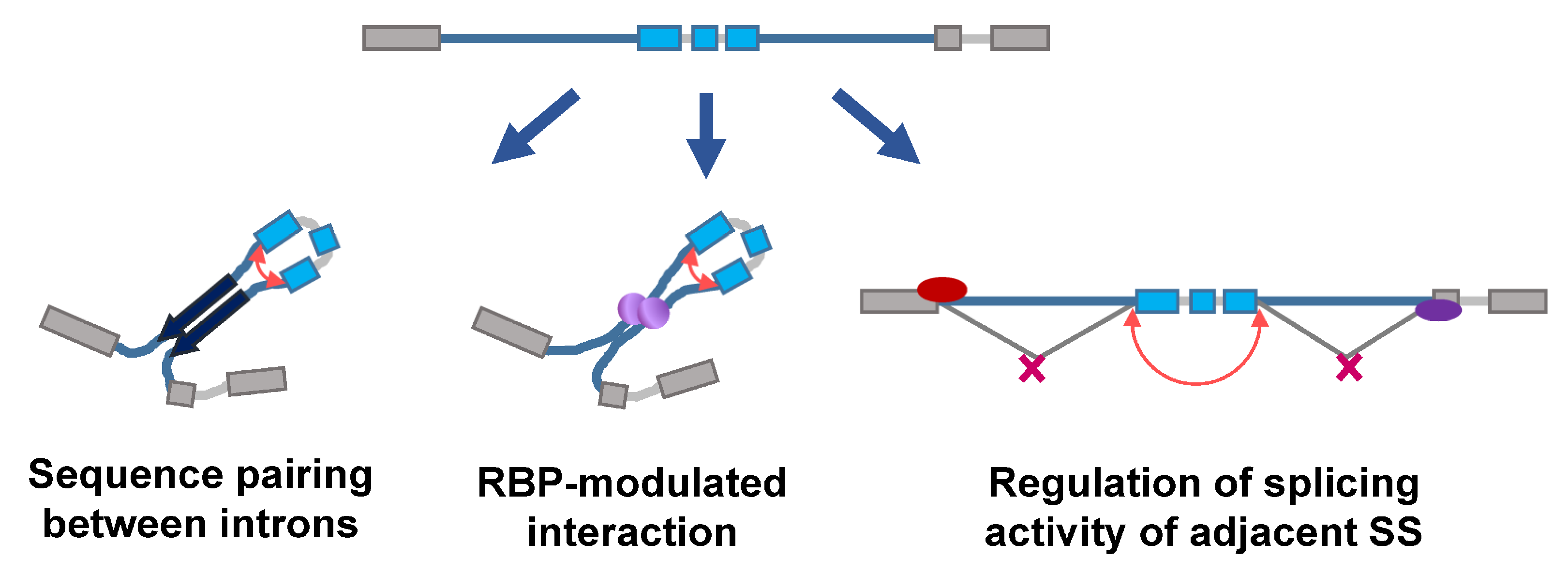
Figure 1.
Different models controlling backsplicing. The given pre-spliced RNA (top) consists of introns (thin lines; blue ones for flanking long introns), circularizable exons (blue boxes) and the exons for linear splicing (grey boxes), and may undergo the following processes for backsplicing. The left panel shows the sequence-pairing dependent mechanism. A pair of arrows indicates the complementary sequences in the flanking long introns for circularizable exons. The red curved arrow indicates where the backsplicing takes place. Similarly, the interaction between flanking introns can be mediated by RBPs (middle panel, purple spheres). Alternatively, the splicing modulators (right panel, red or purple ovals) occupy the splicing signals (SS) and decrease the availability of splice donors in the upstream intron and/or the availability of splice acceptors in the downstream intron, inhibiting linear splicing and thereby favoring backsplicing.
Figure 1.
Different models controlling backsplicing. The given pre-spliced RNA (top) consists of introns (thin lines; blue ones for flanking long introns), circularizable exons (blue boxes) and the exons for linear splicing (grey boxes), and may undergo the following processes for backsplicing. The left panel shows the sequence-pairing dependent mechanism. A pair of arrows indicates the complementary sequences in the flanking long introns for circularizable exons. The red curved arrow indicates where the backsplicing takes place. Similarly, the interaction between flanking introns can be mediated by RBPs (middle panel, purple spheres). Alternatively, the splicing modulators (right panel, red or purple ovals) occupy the splicing signals (SS) and decrease the availability of splice donors in the upstream intron and/or the availability of splice acceptors in the downstream intron, inhibiting linear splicing and thereby favoring backsplicing.

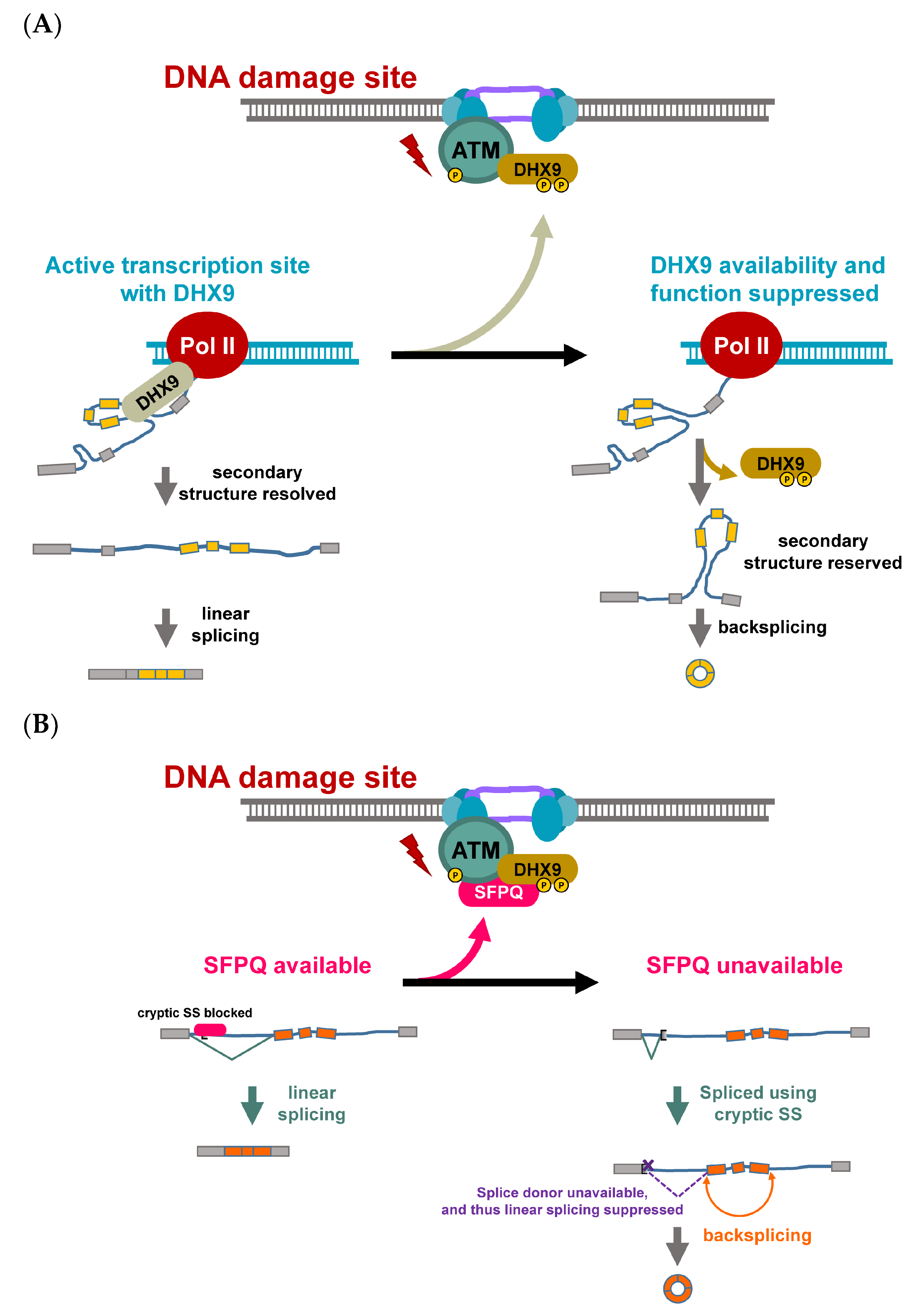
Figure 2.
DNA damage response controls the function and availability of RBP for regulation of backsplicing. (A) In the absence of DNA damage, DHX9 works with transcription machinery for resolving the secondary structure of nascent RNA, and thereby keeps a low frequency of backsplicing (bottom left). Upon double-stranded DNA breaks (top), the PI3KKs (ATM for example) are recruited to the damage sites, accumulated and autophosphorylated for its maximal kinase activity and for recruiting other DDR proteins. DHX9 is recruited to the damage site and becomes phosphorylated near to its double-stranded RNA binding domain, hindering its binding to the double-stranded RNA substrate. The unresolved pairing between upstream and downstream introns allows the backsplicing to take place (bottom right). (B) In a similar fashion, SFPQ forms a complex with DHX9 upon genotoxic stress. When SFPQ sits on the cryptic splice site in the upstream intron of circularizable exons (orange boxes), SFPQ suppresses the linear splicing, which uses the cryptic site (bottom left). However, when SFPQ becomes unavailable due to genotoxic stress-induced relocation to DNA damage sites, the splicing uses up the splice signal, making backsplicing the only option for the rest of the molecule (bottom right).
Figure 2.
DNA damage response controls the function and availability of RBP for regulation of backsplicing. (A) In the absence of DNA damage, DHX9 works with transcription machinery for resolving the secondary structure of nascent RNA, and thereby keeps a low frequency of backsplicing (bottom left). Upon double-stranded DNA breaks (top), the PI3KKs (ATM for example) are recruited to the damage sites, accumulated and autophosphorylated for its maximal kinase activity and for recruiting other DDR proteins. DHX9 is recruited to the damage site and becomes phosphorylated near to its double-stranded RNA binding domain, hindering its binding to the double-stranded RNA substrate. The unresolved pairing between upstream and downstream introns allows the backsplicing to take place (bottom right). (B) In a similar fashion, SFPQ forms a complex with DHX9 upon genotoxic stress. When SFPQ sits on the cryptic splice site in the upstream intron of circularizable exons (orange boxes), SFPQ suppresses the linear splicing, which uses the cryptic site (bottom left). However, when SFPQ becomes unavailable due to genotoxic stress-induced relocation to DNA damage sites, the splicing uses up the splice signal, making backsplicing the only option for the rest of the molecule (bottom right).

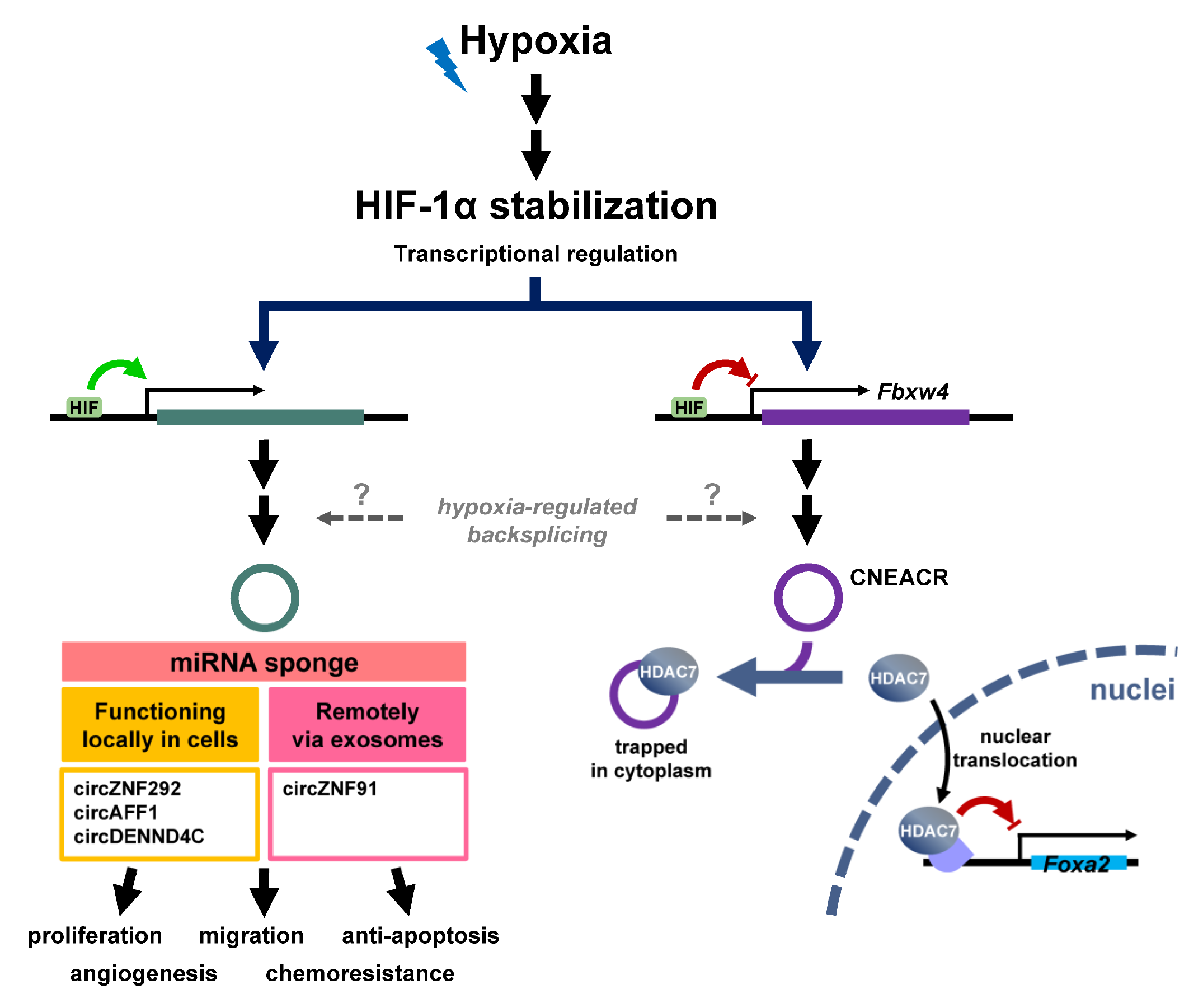
Figure 3.
HIF-mediated transcriptional regulation of circRNAs. The majority of circRNA expression in response to hypoxia is mediated through HIF-1α, which is stabilized by hypoxic stress (top). Through binding to the promoter of target genes, HIF-1α either activates or suppresses the transcription. A group of circRNA is upregulated via transcriptional activation, and mainly functions as a miRNA sponge. CircZNF91 is incorporated into exosomes and delivered to remote sites where it exerts its activity as an miRNA sponge. Nevertheless, CNEACR binds and sequesters HDAC7 in cytoplasm in normoxia, while HIF-1α-inhibited CNEACR expression allows HDAC7 to be released and to translocate to nuclei in hypoxia. HDAC7 represses the expression of Foxa2, relieving the expression of Foxa2-inhibited target genes.
Figure 3.
HIF-mediated transcriptional regulation of circRNAs. The majority of circRNA expression in response to hypoxia is mediated through HIF-1α, which is stabilized by hypoxic stress (top). Through binding to the promoter of target genes, HIF-1α either activates or suppresses the transcription. A group of circRNA is upregulated via transcriptional activation, and mainly functions as a miRNA sponge. CircZNF91 is incorporated into exosomes and delivered to remote sites where it exerts its activity as an miRNA sponge. Nevertheless, CNEACR binds and sequesters HDAC7 in cytoplasm in normoxia, while HIF-1α-inhibited CNEACR expression allows HDAC7 to be released and to translocate to nuclei in hypoxia. HDAC7 represses the expression of Foxa2, relieving the expression of Foxa2-inhibited target genes.

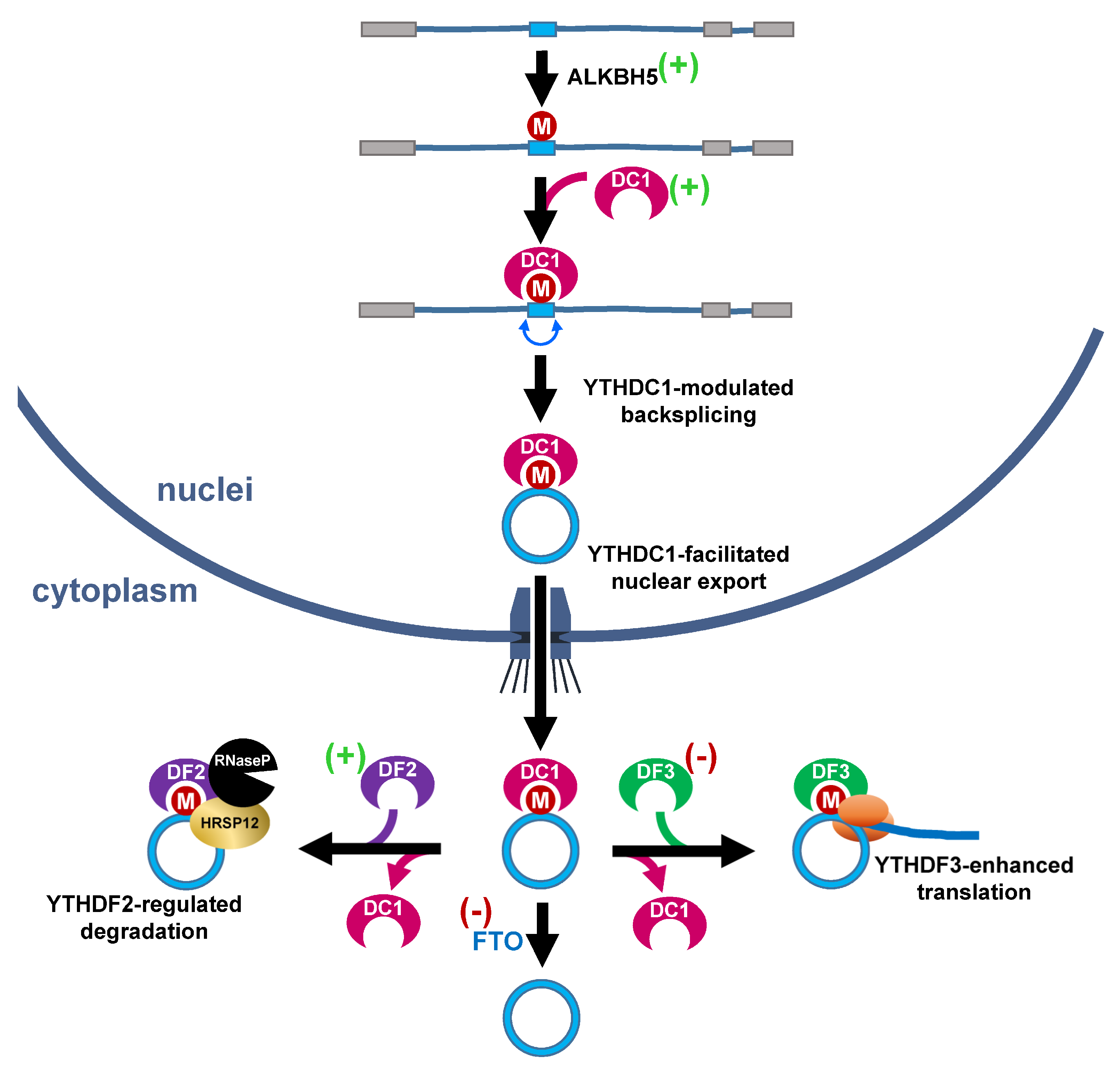
Figure 4.
Potential roles of hypoxia-responsive m6A effectors in regulation of circRNA functions. The deposition of m6A on exons by m6A writers (such as ALKBH5) allows the binding of YTHDC1 (DC1), which promotes the backsplicing of circZNF609. In addition, YTHDC1 also helps the nuclear export of circRNA. In cytoplasm, the m6A-harboring circRNA is recognized by YTHDF3 (DF3) and proceeded to translation, or alternatively bound by YTHDF2 (DF2) and targeted for degradation. The m6A eraser, FTO, eventually removes the modification. (+): reported to be upregulated by hypoxic stress; (−): suppressed by hypoxia.
Figure 4.
Potential roles of hypoxia-responsive m6A effectors in regulation of circRNA functions. The deposition of m6A on exons by m6A writers (such as ALKBH5) allows the binding of YTHDC1 (DC1), which promotes the backsplicing of circZNF609. In addition, YTHDC1 also helps the nuclear export of circRNA. In cytoplasm, the m6A-harboring circRNA is recognized by YTHDF3 (DF3) and proceeded to translation, or alternatively bound by YTHDF2 (DF2) and targeted for degradation. The m6A eraser, FTO, eventually removes the modification. (+): reported to be upregulated by hypoxic stress; (−): suppressed by hypoxia.

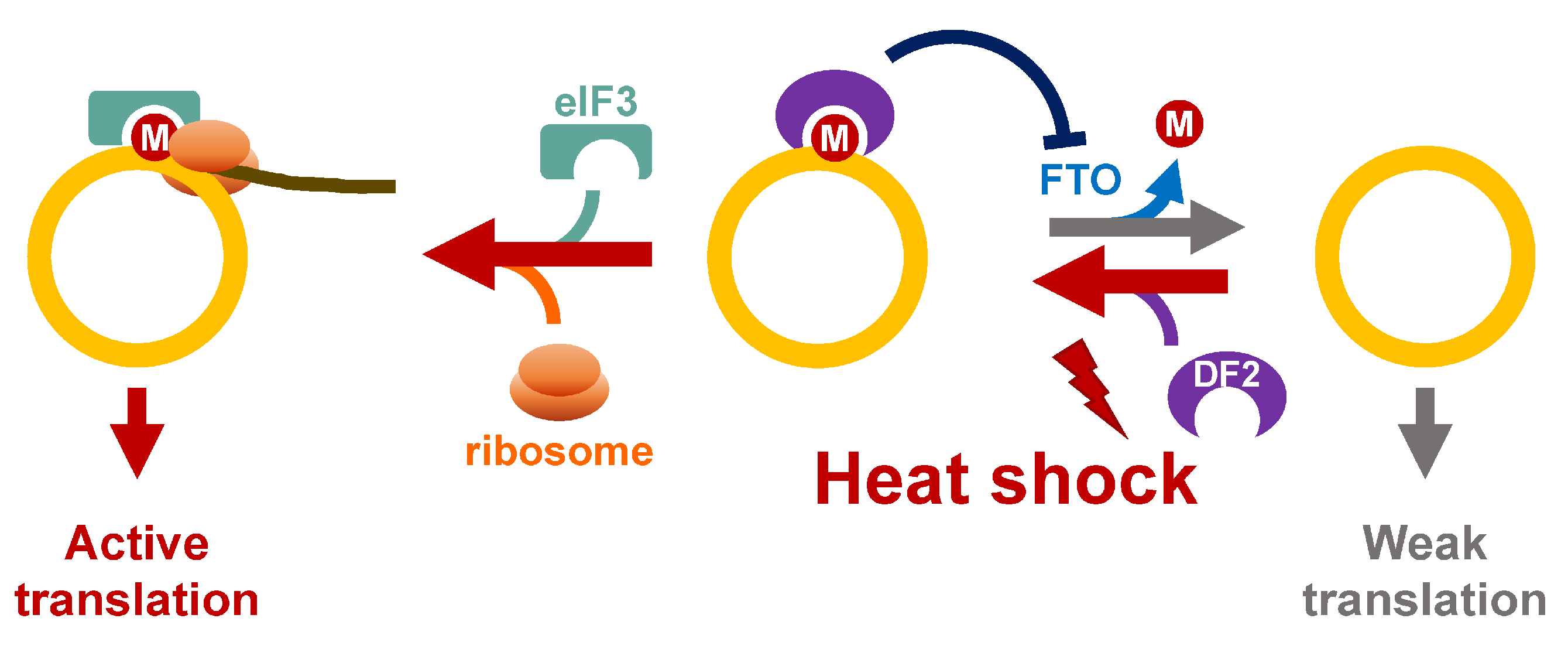
Figure 5.
Potential roles of m6A in regulation of circRNA translation in response to heat shock. Upon heat stress, YTHDF2 (DF2) binds m6A-modified circRNA, and prevents FTO-mediated demethylation. The ribosomes are recruited to circRNA through eIF3/YTHDF2.
Figure 5.
Potential roles of m6A in regulation of circRNA translation in response to heat shock. Upon heat stress, YTHDF2 (DF2) binds m6A-modified circRNA, and prevents FTO-mediated demethylation. The ribosomes are recruited to circRNA through eIF3/YTHDF2.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lee, Y.-C.; Wang, W.-Y.; Lin, H.-H.; Huang, Y.-R.; Lin, Y.-C.; Hsiao, K.-Y. The Functional Roles and Regulation of Circular RNAs during Cellular Stresses. Non-Coding RNA 2022, 8, 38. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna8030038
AMA Style
Lee Y-C, Wang W-Y, Lin H-H, Huang Y-R, Lin Y-C, Hsiao K-Y. The Functional Roles and Regulation of Circular RNAs during Cellular Stresses. Non-Coding RNA. 2022; 8(3):38. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna8030038
Chicago/Turabian StyleLee, Yueh-Chun, Wei-Yu Wang, Hui-Hsuan Lin, Yi-Ren Huang, Ya-Chi Lin, and Kuei-Yang Hsiao. 2022. "The Functional Roles and Regulation of Circular RNAs during Cellular Stresses" Non-Coding RNA 8, no. 3: 38. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna8030038
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.