
Modulating Vapor Pressure Deficit in the Plant Micro-Environment May Enhance the Bioactive Value of Lettuce


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design, Lettuce Genotypes, and Controlled Growing Conditions
2.2. Plant Growth Parameters, Biomass Production, and Leaf Colorimetry
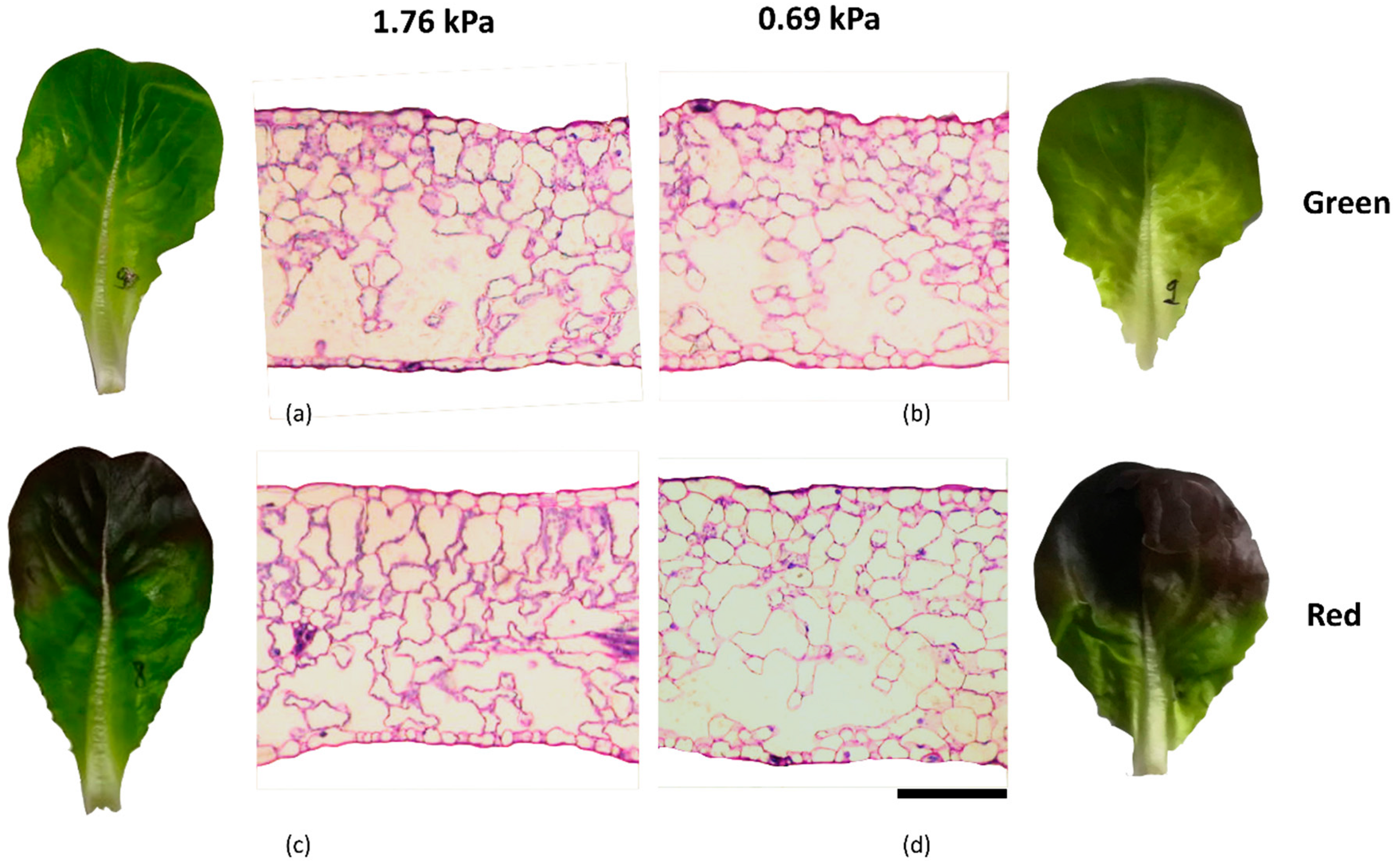
2.3. Anatomical Analyses of Leaves
2.4. Mineral Composition in Leaf Tissue
2.5. Extraction and Quantification of Total Ascorbic Acid, Polyphenols, Lipophylic, and Hydrophilic Antioxidant Activities
2.6. Soil Plant Analysis Development Index and Chlorophyll a Fluorescence Emission
2.7. Statistics
3. Results
3.1. Plant Growth Parameters, Biomass Production, and Leaf Colorimetry
3.2. Morpho-Anatomical Analyses
3.3. Mineral Composition
3.4. Antioxidant Activities and Phytochemicals
3.5. Soil Plant Analysis Development Index and Chlorophyll a Fluorescence Emission
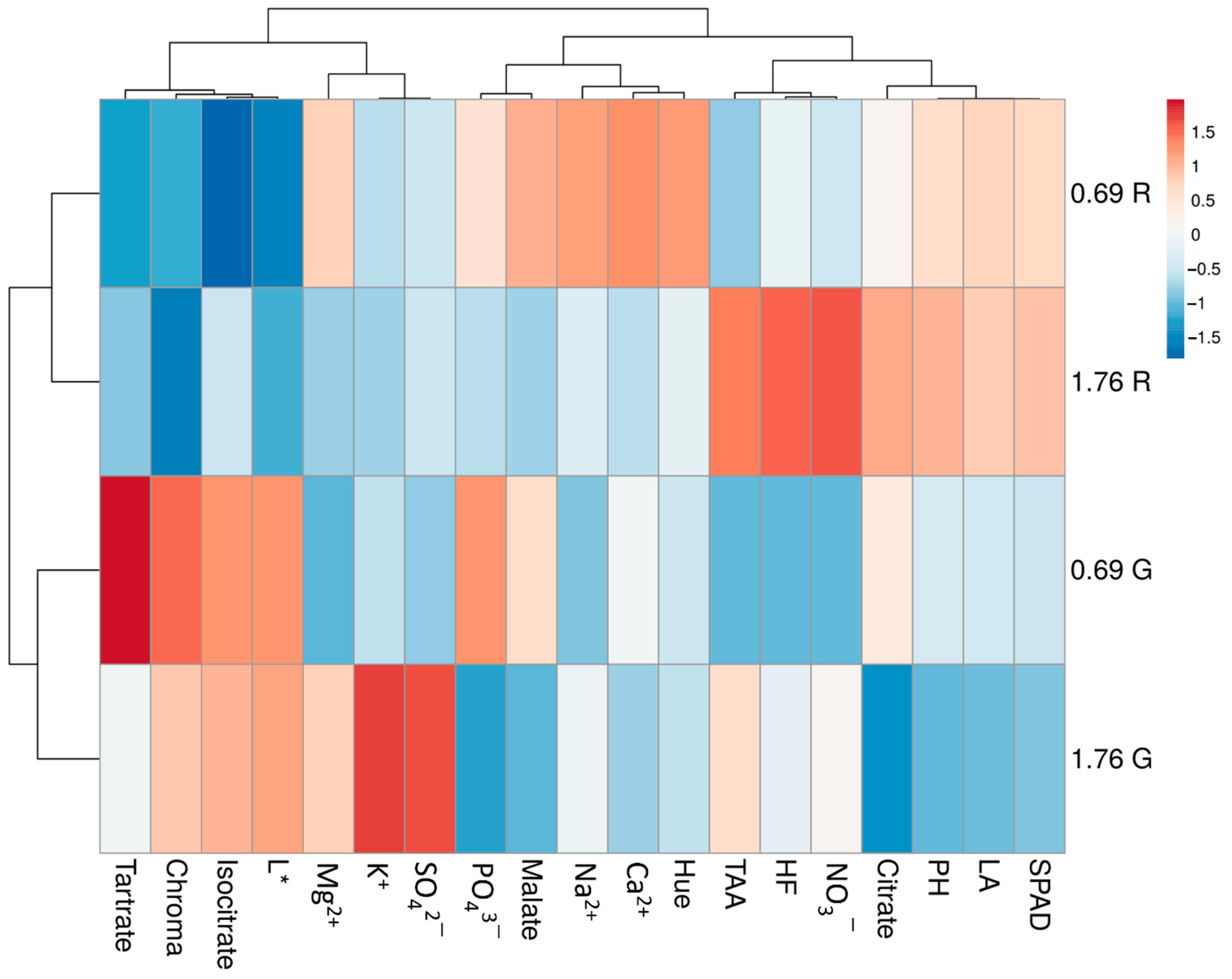
3.6. Hierarchical Clustering of Functional and Nutritional Aspects of Green and Red Salanova
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gruda, N. Impact of Environmental Factors on Product Quality of Greenhouse Vegetables for Fresh Consumption. Crit. Rev. Plant Sci. 2005, 24, 227–247. [Google Scholar] [CrossRef]
- Bakker, J. Physiological disorders in cucumber under high humidity conditions and low ventilation rates in greenhouses. ISHS Acta Hortic. 1984, 257–264. [Google Scholar] [CrossRef]
- Amitrano, C.; Arena, C.; Rouphael, Y.; De Pascale, S.; De Micco, V. Vapour pressure deficit: The hidden driver behind plant morphofunctional traits in controlled environments. Ann. Appl. Biol. 2019, 175, 313–325. [Google Scholar] [CrossRef]
- Krug, H. Gemuseproduktion: Ein Lehr-und Nachschlagewerk fur Studium und Praxis; Verlag Paul Parey: Singhofen, Germany, 1986. [Google Scholar]
- Collier, G.F. Tipburn of Littuce. In Horticultural Reviews; Wiley Blackwell: Hoboken, NJ, USA, 1982; Volume 4, pp. 49–65. [Google Scholar]
- Leonardi, C.; Guichard, S.; Bertin, N. High vapour pressure deficit influences growth, transpiration and quality of tomato fruits. Sci. Hortic. 2000, 84, 285–296. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Devi, J.; Shekoofa, A.; Choudhary, S.; Sadok, W.; Vadez, V.; Riar, M.; Rufty, T. Limited-transpiration response to high vapor pressure deficit in crop species. Plant Sci. 2017, 260, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Dorai, M.; Papadopoulos, A.; Gosselin, A. Influence of electric conductivity management on greenhouse tomato yield and fruit quality. Agronomie 2001, 21, 367–383. [Google Scholar] [CrossRef] [Green Version]
- Rosales, M.A.; Cervilla, L.M.; Sanchez-Rodriguez, E.; Rubio-Wilhelmi, M.M.; Blasco, B.; Rios, J.J.; Soriano, T.; Castilla, N.; Romero, L.; Ruiz, J.M. The effect of environmental conditions on nutritional quality of cherry tomato fruits: Evaluation of two experimental Mediterranean greenhouses. J. Sci. Food Agric. 2011, 91, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.W.; Montagu, M.V.; Inze, D.; Sanmartin, M.; Kanellis, A.; Smirnoff, N.; Benzie, I.J.J.; Strain, J.J.; Favell, D.; Fletcher, J. Plant L-ascorbic acid: Chemistry, function, metabolism, bioavailability and effects of processing. J. Sci. Food Agric. 2000, 80, 825–860. [Google Scholar] [CrossRef]
- Rosales, M.A.; Ruiz, J.M.; Hernández, J.; Soriano, T.; Castilla, N.; Romero, L. Antioxidant content and ascorbate metabolism in cherry tomato exocarp in relation to temperature and solar radiation. J. Sci. Food Agric. 2006, 86, 1545–1551. [Google Scholar] [CrossRef]
- Wang, Y.; Frei, M. Stressed food—The impact of abiotic environmental stresses on crop quality. Agric. Ecosyst. Environ. 2011, 141, 271–286. [Google Scholar] [CrossRef]
- Bisbis, M.B.; Gruda, N.; Blanke, M. Potential impacts of climate change on vegetable production and product quality—A review. J. Clean. Prod. 2018, 170, 1602–1620. [Google Scholar] [CrossRef]
- Favati, F.; Lovelli, S.; Galgano, F.; Miccolis, V.; Di Tommaso, T.; Candido, V. Processing tomato quality as affected by irrigation scheduling. Sci. Hortic. 2009, 122, 562–571. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y. Towards a new definition of quality for fresh fruits and vegetables. Sci. Hortic. 2018, 234, 463–469. [Google Scholar] [CrossRef]
- Amitrano, C.; Arena, C.; De Pascale, S.; De Micco, V. Light and Low Relative Humidity Increase Antioxidants Content in Mung Bean (Vigna radiata L.) Sprouts. Plants 2020, 9, 1093. [Google Scholar] [CrossRef] [PubMed]
- Llorach, R.; Martinez-Sanchez, A.; Tomas-Barberan, F.A.; Gil, M.I.; Ferreres, F. Characterisation of polyphenols and antioxidant properties of five lettuce varieties and escarole. Food Chem. 2008, 108, 1028–1038. [Google Scholar] [CrossRef] [PubMed]
- El-Nakhel, C.; Pannico, A.; Graziani, G.; Kyriacou, M.C.; Giordano, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Variation in Macronutrient Content, Phytochemical Constitution and In Vitro Antioxidant Capacity of Green and Red Butterhead Lettuce Dictated by Different Developmental Stages of Harvest Maturity. Antioxidants 2020, 9, 300. [Google Scholar] [CrossRef] [Green Version]
- Slavin, J.L.; Lloyd, B. Health benefits of fruits and vegetables. Adv. Nutr. 2012, 3, 506–516. [Google Scholar] [CrossRef] [Green Version]
- Gil, M.I. Preharvest factors and fresh-cut quality of leafy vegetables. Acta Hortic. 2016, 57–64. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Feder, N.; O’Brien, T. Plant microtechnique: Some principles and new methods. Am. J. Bot. 1968, 55, 123–142. [Google Scholar] [CrossRef]
- De Micco, V.; Amitrano, C.; Stinca, A.; Izzo, L.G.; Zalloni, E.; Balzano, A.; Barile, R.; Conti, P.; Arena, C. Dust accumulation due to anthropogenic impact induces anatomical and photochemical changes in leaves of Centranthus ruber growing on the slope of the Vesuvius volcano. Plant Biol. 2019, 22, 93–102. [Google Scholar] [CrossRef]
- Kampfenkel, K.; Van Montagu, M.; Inzé, D. Effects of iron excess on Nicotiana plumbaginifolia plants (implications to oxidative stress). Plant Physiol. 1995, 107, 725–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Fogliano, V.; Verde, V.; Randazzo, G.; Ritieni, A. Method for measuring antioxidant activity and its application to monitoring the antioxidant capacity of wines. J. Agric. Food Chem. 1999, 47, 1035–1040. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Gruda, N. Do soilless culture systems have an influence on product quality of vegetables? J. Appl. Bot. Food Qual. 2009, 82, 141–147. [Google Scholar]
- Zhang, D.; Du, Q.; Zhang, Z.; Jiao, X.; Song, X.; Li, J. Vapour pressure deficit control in relation to water transport and water productivity in greenhouse tomato production during summer. Sci. Rep. 2017, 7, 43461. [Google Scholar] [CrossRef]
- Carillo, P.; Cirillo, C.; De Micco, V.; Arena, C.; De Pascale, S.; Rouphael, Y. Morpho-anatomical, physiological and biochemical adaptive responses to saline water of Bougainvillea spectabilis Willd. trained to different canopy shapes. Agric. Water Manag. 2019, 212, 12–22. [Google Scholar] [CrossRef]
- Parry, M.A.; Reynolds, M.; Salvucci, M.E.; Raines, C.; Andralojc, P.J.; Zhu, X.-G.; Price, G.D.; Condon, A.G.; Furbank, R.T. Raising yield potential of wheat. II. Increasing photosynthetic capacity and efficiency. J. Exp. Bot. 2011, 62, 453–467. [Google Scholar] [CrossRef]
- Zhu, X.-G.; Long, S.P.; Ort, D.R. Improving photosynthetic efficiency for greater yield. Annu. Rev. Plant Biol. 2010, 61, 235–261. [Google Scholar] [CrossRef] [Green Version]
- Ogaya, R.; Peñuelas, J.; Asensio, D.; Llusià, J. Chlorophyll fluorescence responses to temperature and water availability in two co-dominant Mediterranean shrub and tree species in a long-term field experiment simulating climate change. Environ. Exp. Bot. 2011, 71, 123–127. [Google Scholar] [CrossRef]
- Sharma, D.K.; Andersen, S.B.; Ottosen, C.O.; Rosenqvist, E. Wheat cultivars selected for high Fv/Fm under heat stress maintain high photosynthesis, total chlorophyll, stomatal conductance, transpiration and dry matter. Physiol. Plant. 2015, 153, 284–298. [Google Scholar] [CrossRef]
- Baker, N.R.; Rosenqvist, E. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poudyal, D.; Rosenqvist, E.; Ottosen, C.-O. Phenotyping from lab to field—Tomato lines screened for heat stress using Fv/Fm maintain high fruit yield during thermal stress in the field. Funct. Plant Biol. 2019, 46, 44–55. [Google Scholar] [CrossRef]
- Nankishore, A.; Farrell, A.D. The response of contrasting tomato genotypes to combined heat and drought stress. J. Plant Physiol. 2016, 202, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Liu, T.; Jiao, X.; Song, X.; Zhang, J.; Li, J. Leaf anatomical adaptations have central roles in photosynthetic acclimation to humidity. J. Exp. Bot. 2019, 70, 4949–4962. [Google Scholar] [CrossRef]
- Tosens, T.; Niinemets, U.; Vislap, V.; Eichelmann, H.; Diez, P.C. Developmental changes in mesophyll diffusion conductance and photosynthetic capacity under different light and water availabilities in Populus tremula: How structure constrains function. Plant Cell Env. 2012, 35, 839–856. [Google Scholar] [CrossRef]
- Murphy, M.R.C.; Jordan, G.J.; Brodribb, T.J. Acclimation to humidity modifies the link between leaf size and the density of veins and stomata. Plant Cell Environ. 2014, 37, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Lihavainen, J.; Keinänen, M.; Keski-Saari, S.; Kontunen-Soppela, S.; Sõber, A.; Oksanen, E. Artificially decreased vapour pressure deficit in field conditions modifies foliar metabolite profiles in birch and aspen. J. Exp. Bot. 2016, 67, 4367–4378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Ge, T.; Sui, F.G.; Nie, S.; Sun, N.B.; Xiao, H.; Tong, C.L. Differential responses of yield and selected nutritional compositions to drought stress in summer maize grains. J. Plant Nutr. 2010, 33, 1811–1818. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Giordano, M.; Pannico, A.; Carillo, P.; Fusco, G.M.; De Pascale, S.; Rouphael, Y. Cultivar-specific performance and qualitative descriptors for butterhead Salanova lettuce produced in closed soilless cultivation as a candidate salad crop for human life support in space. Life 2019, 9, 61. [Google Scholar] [CrossRef] [Green Version]
- Soetan, K.; Olaiya, C.; Oyewole, O. The importance of mineral elements for humans, domestic animals and plants—A review. Afr. J. Food Sci. 2010, 4, 200–222. [Google Scholar]
- Gupta, K.; Wagle, D. Nutritional and antinutritional factors of green leafy vegetables. J. Agric. Food Chem. 1988, 36, 472–474. [Google Scholar] [CrossRef]
- Peuke, A.D.; Jeschke, W.D.; Hartung, W. Flows of elements, ions and abscisic acid in Ricinus communis and site of nitrate reduction under potassium limitation. J. Exp. Bot. 2002, 53, 241–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Statement on possible public health risks for infants and young children from the presence of nitrates in leafy vegetables. EFSA J. 2010, 8, 1935. [CrossRef] [Green Version]
- Colla, G.; Kim, H.-J.; Kyriacou, M.C.; Rouphael, Y. Nitrate in fruits and vegetables. Sci. Hortic. 2018, 237, 221–238. [Google Scholar] [CrossRef]
- Navarro, A.; Elia, A.; Conversa, G.; Campi, P.; Mastrorilli, M. Potted mycorrhizal carnation plants and saline stress: Growth, quality and nutritional plant responses. Sci. Hortic. 2012, 140, 131–139. [Google Scholar] [CrossRef]
- Jha, S.N. Colour Measurements and Modeling. In Nondestructive Evaluation of Food Quality; Springer: Berlin/Heidelberg, Germany, 2010; pp. 17–40. [Google Scholar]
- Tuncay, O. Relationships between nitrate, chlorophyll and chromaticity values in rocket salad and parsley. Afr. J. Biotechnol. 2011, 10, 17152–17159. [Google Scholar]
- Kim, D.E.; Shang, X.; Assefa, A.D.; Keum, Y.S.; Saini, R.K. Metabolite profiling of green, green/red, and red lettuce cultivars: Variation in health beneficial compounds and antioxidant potential. Food Res. Int. 2018, 105, 361–370. [Google Scholar] [CrossRef]
- Baslam, M.; Morales, F.; Garmendia, I.; Goicoechea, N. Nutritional quality of outer and inner leaves of green and red pigmented lettuces (Lactuca sativa L.) consumed as salads. Sci. Hortic. 2013, 151, 103–111. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Petropoulos, S.A.; Pannico, A.; Kyriacou, M.C.; Giordano, M.; Colla, G.; Troise, A.D.; Vitaglione, P.; De Pascale, S.; Rouphael, Y. The bioactive profile of lettuce produced in a closed soilless system as configured by combinatorial effects of genotype and macrocation supply composition. Food Chem. 2020, 309, 125713. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Schellhorn, H.E. New developments and novel therapeutic perspectives for vitamin C. J. Nutr. 2007, 137, 2171–2184. [Google Scholar] [CrossRef] [Green Version]
- Oh, M.M.; Carey, E.E.; Rajashekar, C.B. Environmental stresses induce health-promoting phytochemicals in lettuce. Plant Physiol. Biochem. 2009, 47, 578–583. [Google Scholar] [CrossRef]
- Grange, R.; Hand, D. A review of the effects of atmospheric humidity on the growth of horticultural crops. J. Hortic. Sci. 1987, 62, 125–134. [Google Scholar] [CrossRef]
- Tack, J.; Singh, R.K.; Nalley, L.L.; Viraktamath, B.C.; Krishnamurthy, S.L.; Lyman, N.; Jagadish, K.S. High vapor pressure deficit drives salt-stress-induced rice yield losses in India. Glob. Chang. Biol. 2015, 21, 1668–1678. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.; Grange, R.; Picken, A. An analysis of the accumulation of water and dry matter in tomato fruit. Plant Cell Environ. 1987, 10, 157–162. [Google Scholar]
- Boyer, J.S. Plant productivity and environment. Science 1982, 218, 443–448. [Google Scholar] [CrossRef]
- Bertin, N.; Guichard, S.; Leonardi, C.; Longuenesse, J.; Langlois, D.; Navez, B. Seasonal evolution of the quality of fresh glasshouse tomatoes under Mediterranean conditions, as affected by air vapour pressure deficit and plant fruit load. Ann. Bot. 2000, 85, 741–750. [Google Scholar] [CrossRef]
- Rosales, M.A.; Ríos, J.J.; Cervilla, L.M.; Rubio-Wilhelmi, M.M.; Blasco, B.; Ruiz, J.M.; Romero, L. Environmental conditions in relation to stress in cherry tomato fruits in two experimental Mediterranean greenhouses. J. Sci. Food Agric. 2009, 89, 735–742. [Google Scholar] [CrossRef]
- Burritt, D.J.; Mackenzie, S. Antioxidant metabolism during acclimation of Begonia × erythrophylla to high light levels. Ann. Bot. 2003, 91, 783–794. [Google Scholar] [CrossRef] [Green Version]
- Dixon, R.A.; Paiva, N.L. Stress-Induced Phenylpropanoid Metabolism. Plant Cell 1995, 7, 1085. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| PA (cm2 Plant−1) | LN (No. Plant−1) | FB (g Plant−1) | DB (g Plant−1) | b * | L * | Chroma | Hue | |
|---|---|---|---|---|---|---|---|---|
| Cultivar | ||||||||
| G | 196 ± 1.02 b | 51.9 ± 0.72 a | 33.2 ± 0.50 b | 3.60 ± 0.03 a | 40.3 ± 1.21a | 49.4 ± 0.46 a | 28.1 ± 0.44 a | 107 ± 4.46 b |
| R | 214 ± 0.37 a | 49.3 ± 0.81 b | 36.8 ± 0.29 a | 3.75 ± 0.02 a | 3.62 ± 0.78 b | 22.7 ± 0.47 b | 21.5 ± 0.45 b | 195 ± 3.34 a |
| VPD | ||||||||
| 0.69 kPa | 224 ± 1.01 a | 53.9 ± 0.76 a | 37.9 ± 0.48 a | 4.80 ± 0.06 a | 28.8 ± 1.16 a | 34.0 ± 0.33 b | 42.3 ± 0.62 a | 171 ± 3.15 a |
| 1.76 kPa | 186 ± 0.33 b | 47.3 ± 0.77 b | 32.1 ± 0.31 b | 2.55 ± 0.03 b | 17.1 ± 0.88 b | 38.1 ± 0.34 a | 7.24 ± 0.66 b | 132 ± 2.02 b |
| Int. | ||||||||
| 0.69 G | 211 ± 0.17 b | 53.4 ± 0.24 a | 35.6 ± 0.17 b | 4.75 ± 0.02 a | 44.5 ± 0.81 a | 48.5 ± 0.42 b | 46.2 ± 0.74 a | 106 ± 0.44 c |
| 0.69 R | 237 ± 0.03 a | 54.4 ± 0.40 a | 40.4 ± 0.22 a | 4.85 ± 0.03 a | 9.25 ± 0.80 c | 19.5 ± 0.31 d | 9.84 ± 0.73 c | 236 ± 0.88 a |
| 1.76 G | 180 ± 0.20 d | 50.4 ± 0.25 b | 30.8± 0.23 d | 2.46 ± 0.01 b | 36.2 ± 0.70 b | 50.3 ± 0.77 a | 38.3 ± 0.68 b | 109 ± 0.28 c |
| 1.76 R | 192 ± 0.21 c | 44.2 ± 0.49 c | 33.4 ± 0.07 c | 2.66 ± 0.01 b | -2.00 ± 0.16 d | 25.9 ± 0.13 c | 4.60 ± 0.11 d | 154 ± 2.18 b |
| Sig. | ||||||||
| C | *** | * | *** | NS | *** | *** | *** | *** |
| VPD | *** | *** | *** | ** | *** | *** | *** | *** |
| C × VPD | *** | *** | *** | *** | * | *** | * | *** |
| UET (µm) | LET (µm) | PT (µm) | ST (µm) | LT (µm) | IS (%) | |
|---|---|---|---|---|---|---|
| Cultivar | ||||||
| G | 23.4 ± 0.66 a | 16.7 ± 1.03 a | 97.3 ± 4.69 a | 148 ± 6.29 a | 287 ± 8.86 a | 46.2 ± 2.87 a |
| R | 22.4 ± 1.05 a | 14.8 ± 0.65 b | 94.2 ± 3.71 a | 149 ± 11.8 a | 282 ± 11.3 a | 45.3 ± 2.05 a |
| VPD | ||||||
| 0.69 kPa | 22.9 ± 0.64 a | 15.4 ± 0.79 a | 95.5 ± 5.06 a | 146 ± 7.57 a | 281 ± 8.76 a | 45.6 ± 1.89 a |
| 1.76 kPa | 22.8 ± 1.11 a | 16.1 ± 0.74 a | 95.9 ± 5.31 a | 151 ± 9.15 a | 287 ± 13.4 a | 46.8 ± 2.50 a |
| Int. | ||||||
| 0.69 G | 24.2 ± 0.63 a | 16.2 ± 0.79 ab | 94.5 ± 3.10 a | 144 ± 3.56 a | 280 ± 4.85 a | 44.2 ± 1.96 a |
| 0.69 R | 22.7 ± 0.64 ab | 17.2 ± 0.79 a | 99.8 ± 3.93 a | 152 ± 8.02 a | 293 ± 11.73 a | 44.9 ± 1.33 a |
| 1.76 G | 21.7 ± 0.70 b | 14.6 ± 0.48 b | 96.5 ± 3.19 a | 147 ± 5.47 a | 281 ± 8.02 a | 48.1 ± 1.83 a |
| 1.76 R | 23.1 ± 0.82 ab | 15.1 ± 0.52 b | 92.1 ± 4.24 a | 151 ± 7.56 a | 282 ± 10.82 a | 45.6 ± 1.35 a |
| Sig. | ||||||
| C | NS | * | NS | NS | NS | NS |
| VPD | NS | NS | NS | NS | NS | NS |
| C × VPD | * | * | NS | NS | NS | NS |
| NO3− (mg/kg FW) | PO43− (g/kg DW) | SO42− (g/kg DW) | K+ (g/kg DW) | Ca2+ (g/kg DW) | Mg2+ (g/kg DW) | Na2+ (g/kg DW) | Malate (g/kg DW) | Tartrate (g/kg DW) | Citrate (g/kg DW) | Isocitrate (g/kg DW) | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cultivar | |||||||||||
| G | 4013 ± 711 b | 6.92 ± 0.73 a | 2.01 ± 0.25 a | 58.2 ± 2.36 a | 14.7 ± 0.86 b | 3.46 ± 0.74 a | 3.36 ± 0.40 a | 56.5 ± 4.82 b | 3.64 ± 0.28 a | 12.2 ± 1.86 a | 0.23 ± 0.03 a |
| R | 4911 ± 625 a | 7.44 ± 0.67a | 1.78 ± 0.29 a | 46.6 ± 5.54 b | 18.3 ± 2.44 a | 3.74 ± 0.64 a | 4.11 ± 0.98 a | 84.8 ± 8.60 a | 1.98 ± 0.25 b | 14.5 ± 1.31 a | 0.17 ± 0.03 b |
| VPD | |||||||||||
| 0.69 kPa | 4513 ± 746 b | 8.19 ± 0.36 a | 1.53 ± 0.21b | 42.6 ± 1.56 b | 18.3 ± 1.15 a | 3.56 ± 0.76 a | 3.80 ± 0.56 a | 92.6 ± 8.19 a | 3.21 ± 0.26 a | 13.7 ± 1.41 a | 0.18 ± 0.03 a |
| 1.76 kPa | 4911 ± 553 a | 6. 24 ±1.39 b | 2.25 ± 0.37a | 62.2 ± 7.15 a | 14.7 ± 1.87 b | 3.64 ± 0.59 a | 3.65 ± 0.66 a | 43.3 ± 1.86 b | 2.41 ± 0.28 b | 12.9 ± 2.45 a | 0.21 ± 0.03 a |
| Int. | |||||||||||
| 0.69 G | 3246 ± 556 d | 8.18 ± 0.27 a | 1.26 ± 0.13 b | 37.5 ± 0.48 b | 15.6 ± 0.27 b | 3.04 ± 0.64 c | 3.09 ± 0.24 a | 76.5 ± 4.28 b | 4.65 ± 0.22 a | 13.4 ± 0.74 a | 0.23 ± 0.02 a |
| 0.69 R | 3980 ± 380 c | 8.21 ± 0.19 a | 1.80 ± 0.15 b | 47.7 ± 2.16 b | 21.1 ± 1.76 a | 4.08 ± 0.25 a | 4.52 ± 0.64 a | 108.6 ± 7.83 a | 1.77 ± 0.08 c | 14.2 ± 1.34 a | 0.15 ± 0.01 b |
| 1.76 G | 4080 ± 310 b | 5.65 ± 0.91 b | 2.75 ± 0.23 a | 78.9 ± 3.77 a | 13.9 ± 1.19 b | 3.87 ± 0.21 ab | 3.62 ± 0.32 a | 31.5 ± 1.09 c | 2.62 ± 0.12 b | 10.9 ± 1.49 a | 0.23 ± 0.01 a |
| 1.76 R | 4942 ± 490 a | 6.84 ± 0.96 ab | 1.75 ± 0.28 b | 45.5 ± 6.77 b | 15.6 ± 1.36 b | 3.41 ± 0.77 bc | 3.69 ± 0.68 a | 55.1 ± 1.54 b | 2.19 ± 0.33 bc | 14.9 ± 1.93 a | 0.19 ± 0.03 ab |
| Sig. | |||||||||||
| C | *** | NS | NS | * | * | NS | NS | ** | *** | NS | * |
| VPD | * | * | ** | *** | * | NS | NS | *** | ** | NS | NS |
| C × VPD | *** | * | ** | *** | * | ** | NS | NS | *** | NS | * |
| TAA mg100 g−1 FW | PH mg GA eq. 100 g−1 DW | HAA mmol AA eq. 100 g−1 DW | LAA mmol trolox eq. 100 g−1 DW | |
|---|---|---|---|---|
| Cultivar | ||||
| G | 76.6 ± 5.22 b | 9.21 ± 0.24 b | 14.9 ± 0.52 a | 30.2 ± 1.73 b |
| R | 97.7 ± 3.74 a | 10.3 ± 0.48 a | 16.6 ± 0.36 a | 42.3 ± 0.70 a |
| VPD | ||||
| 0.69 kPa | 63.7 ± 3.65 b | 8.08 ± 0.21 b | 10.3 ± 0.57 b | 36.1 ± 1.20 a |
| 1.76 kPa | 111 ± 6.86 a | 11.9 ± 0.54 a | 11.2 ± 0.25 a | 36.5 ± 1.77 a |
| Int. | ||||
| 0.69 G | 52.5 ± 2.93 d | 5.83 ± 0.14 c | 9.71 ± 0.41 c | 28.9 ± 1.01 c |
| 0.69 R | 75.1 ± 1.44 c | 10.3 ± 0.13 a | 10.8 ± 0.32 ab | 43.3 ± 0.38 a |
| 1.76 G | 101 ± 4.57 b | 6.77 ± 0.19 b | 10.4 ± 0.21 b | 31.5 ± 1.45 b |
| 1.76 R | 120 ± 4.59 a | 10.3 ± 0.71 a | 11.6 ± 0.09 a | 41.4 ± 0.64 a |
| Sig. | ||||
| C | ** | *** | NS | *** |
| VPD | *** | * | * | * |
| C × VPD | * | * | * | * |
| SPAD Index | Fv/Fm | |||
|---|---|---|---|---|
| 12 DAT | 23 DAT | 12 DAT | 23 DAT | |
| Cultivar | ||||
| G | 28.07 ± 0.74 b | 27.9 ± 1.46 b | 0.77 ± 0.10 a | 0.80 ± 0.07a |
| R | 41.85 ± 0.91 a | 41.1 ± 1.17 a | 0.77 ± 0.30 a | 0.80 ± 0.12a |
| VPD | ||||
| 0.69 kPa | 36.70 ± 0.85 a | 35.2 ± 1.00 a | 0.78 ± 0.11 a | 0.82 ± 0.12a |
| 1.76 kPa | 30.51 ± 0.69 b | 33.8 ± 1.63 b | 0.76 ± 0.20 b | 0.78 ± 0.13b |
| Int. | ||||
| 0.69 G | 28.5 ± 0.57 c | 28.5 ± 0.54 c | 0.79 ± 0.06 a | 0.82 ± 0.02 a |
| 0.69 R | 44.9 ± 0.57 a | 41.8 ± 0.46 a | 0.78 ± 0.06 a | 0.82 ± 0.01 a |
| 1.76 G | 27.6 ± 0.35 c | 27.3 ± 0.92 c | 0.75 ± 0.13 b | 0.79 ± 0.06 b |
| 1.76 R | 38.2 ± 0.69 b | 33.4 ± 0.71 b | 0.77 ± 0.07 ab | 0.78 ± 0.07 b |
| Sig. | ||||
| C | *** | * | NS | NS |
| VPD | *** | *** | * | *** |
| C × VPD | *** | * | * | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amitrano, C.; Rouphael, Y.; De Pascale, S.; De Micco, V. Modulating Vapor Pressure Deficit in the Plant Micro-Environment May Enhance the Bioactive Value of Lettuce. Horticulturae 2021, 7, 32. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7020032
Amitrano C, Rouphael Y, De Pascale S, De Micco V. Modulating Vapor Pressure Deficit in the Plant Micro-Environment May Enhance the Bioactive Value of Lettuce. Horticulturae. 2021; 7(2):32. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7020032
Chicago/Turabian StyleAmitrano, Chiara, Youssef Rouphael, Stefania De Pascale, and Veronica De Micco. 2021. "Modulating Vapor Pressure Deficit in the Plant Micro-Environment May Enhance the Bioactive Value of Lettuce" Horticulturae 7, no. 2: 32. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7020032