
Seed Maturity and Its In Vitro Initiation of Chilean Endemic Geophyte Alstroemeriapelegrina L.


Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Characterization
2.2. Morphological Characterization of Fruits and Seeds
2.2.1. Characterization of Fruits
2.2.2. Characterization of Seeds
2.3. Seed Experiment
2.3.1. Establishment In Vitro
2.3.2. Histological Study of Seeds
3. Results
3.1. Characterization of Phenological Stages
3.2. Morphological Characterization of Fruits and Seeds
3.2.1. Fruits Characterization
3.2.2. Seed Characterization
3.3. Seed Testing
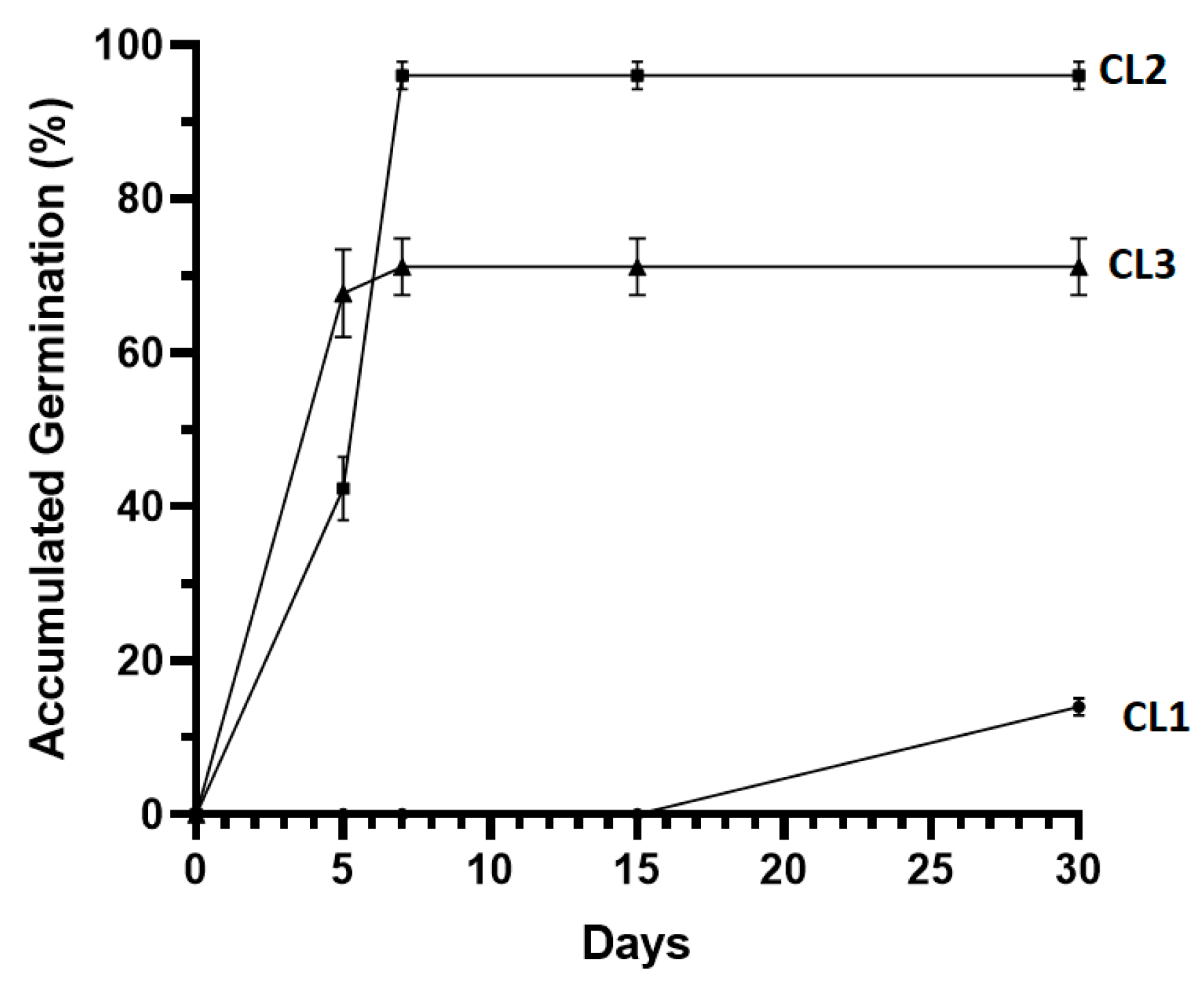
3.3.1. In Vitro Establishment
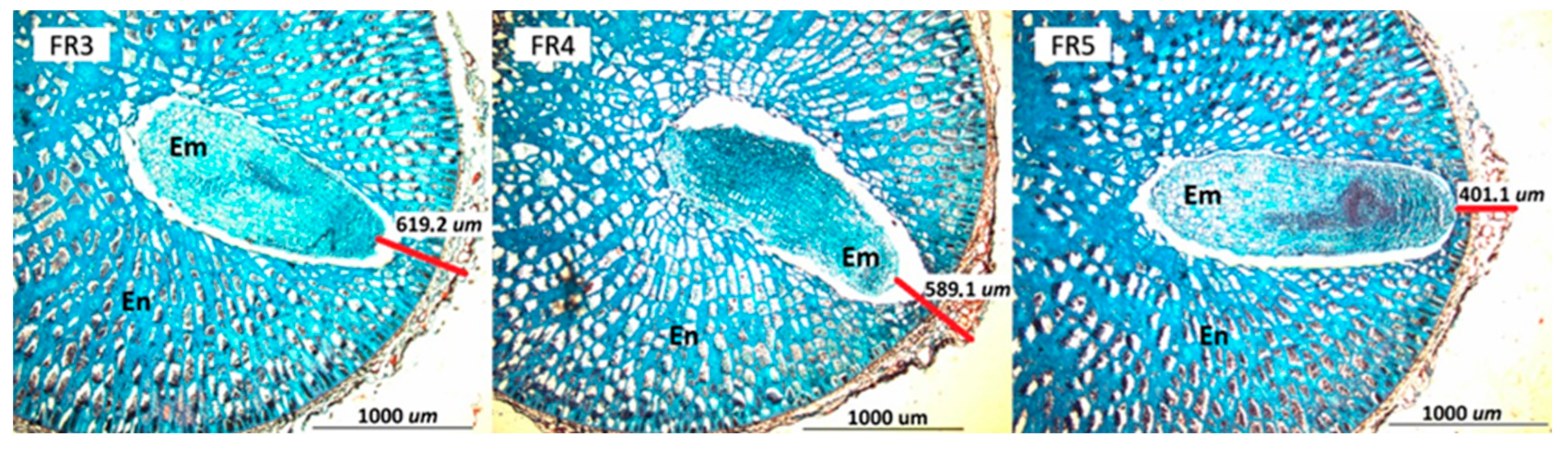
3.3.2. Histological Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodríguez, R.; Marticorena, C.; Alarcón, D.; Baeza, C.; Cavieres, L.; Finot, V.; Fuentes, N.; Kiessling, A.; Mihoc, M.; Pauchard, A.; et al. Catálogo de las plantas vasculares de Chile. Gayana Bot. 2018, 75, 1–430. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Schick, M.; Moreira-Muñoz, A. Consideraciones taxonómicas y de distribución geográfica de especies chilenas del género Tropaeolum L. (Tropaeolaceae): T. reicheanum Buchenau ex Reiche; T. looseri Sparre; T. leptophyllum G. Don y T. myriophyllum (Poepp. & Endl.) Sparre. Gayana Bot. 2013, 70, 294–306. [Google Scholar]
- Aagesen, L.; Sanso, M. The phylogeny of Alstroemeriaceae; based on Morphology; rps16 Intron; and rbcL sequence data. Syst. Bot. 2003, 28, 47–69. [Google Scholar]
- Chacón, J.; Sousa, A.; Baeza, C.M.; Renner, S.S. Ribosomal DNA distribution and a genus-wide phylogeny reveal patterns of chromosomal evolution in Alstroemeria (Alstroemeriaceae). Am.J. Bot. 2012, 99, 1501–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, M.; Moreira, A. Alstroemerias de Chile: Diversidad; Distribución y Conservación; Taller La Era: Santiago, Chile, 2003; pp. 16–25. [Google Scholar]
- Sanso, A.M. El Género Alstroemeria (Alstroemeriaceae) En Argentina. Darwiniana 1996, 34, 349–382. [Google Scholar]
- Buitendijk, J.; Pinsonneaux, N.; Van Donk, A.; Ramanna, M.; Van Lammeren, A. Embryo rescue by half-ovule culture for the production of interspecific hybrids in Alstroemeria. Sci. Hortic. 1995, 64, 65–75. [Google Scholar] [CrossRef]
- Ministerio del Medio Ambiente (MMA). Inventario Nacional de especies de Chile. Available online: http://especies.mma.gob.cl/ (accessed on 15 March 2018).
- Finot, V.; Baeza, C.; Muñoz-Schick, M.; Ruiz, E.; Espejo, J.; Alarcón, D.; Carrasco, P.; Novoa, P.; Eyzaguirre, M. Guía de Campo Alstroemerias Chilenas, 1st ed.; Corporación Chilena de la Madera: Concepción, Chile, 2018; pp. 176–179. [Google Scholar]
- Ravenna, P. New or noteworthy species of Alstroemeria. Phytologia 1988, 64, 281–288. [Google Scholar] [CrossRef]
- Bayer, E. Die Gattung Alstroemeria in Chile. Mitt. Bot. Staatssamml. Münch. 1987, 24, 241–362. [Google Scholar]
- Riedemann, P.; Aldunate, G. Flora Nativa de Valor Ornamental; Identificación y Propagación: Chile; Zona Centro, 2nd ed.; Ediciones Jardín Botánico Chagual: Santiago, Chile, 2016; pp. 310–311. [Google Scholar]
- Sanso, A.; Xifreda, C. Generic delimitation between Alstroemeria and Bomarea (Alstroemeriaceae). Ann. Bot. 2001, 88, 1057–1069. [Google Scholar] [CrossRef] [Green Version]
- King, J.; Bridgen, M. Environmental and Genotypic Regulation of Alstroemeria Seed Germination. HortScience 1990, 25, 1607–1609. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, H.; Kester, D. Propagación de Plantas: Principios y Prácticas, 2nd ed.; Compañía Editorial Continental: México, Mexico, 1999; pp. 75–204. [Google Scholar]
- Latencia de Yemas y Semillas. Available online: http://www.euita.upv.es/varios/biologia/Temas/tema_16.htm#Latenciadeyemas (accessed on 5 March 2021).
- Lu, C.; Bridgen, M.P. Effects of genotype; culture medium and embryo developmental stage on the in vitro responses from ovule cultures of interspecific hybrids of Alstroemeria. Plant Sci. 1996, 116, 205–212. [Google Scholar] [CrossRef]
- Burchi, G.; Mercuri, A.; Bianchini, C.; Bregliano, R.; Schiva, T. New interspecific hybrids of alstroemeria obtained through in vitro embryo-rescue. Acta Hortic. 2000, 508, 233–236. [Google Scholar] [CrossRef]
- Kamstra, S.A.; Ramanna, M.S.; de Jeu, M.J.; Kuipers, A.G.J.; Jacobsen, E. Homoeologous chromosome pairing in the distant hybrid Alstroemeria aurea × A. inodora and the genome composition of its backcross derivatives determined by fluorescence in situ hybridization with species-specific probes. Heredity 1999, 82, 69–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Benedetti, L.; Burchi, G.; Mercuri, A.; Pecchioni, N.; Faccioli, P.; Schiva, T. Random amplified polymorphic DNA (RAPD) analysis for the verification of hybridity in interspecific crosses of Alstroemeria. Plant Breed. 2000, 119, 443–445. [Google Scholar] [CrossRef]
- Bridgen, M.; Kollman, E.; Chunsheng, L. Interspecific hybridization of alstroemeria for the development of new; ornamental plants. Acta Hortic. 2009, 836, 73–78. [Google Scholar] [CrossRef]
- Grigoriadau, K.; Krigas, N.; Sarropoulou, V.; Papanastasi, K.; Tsoktouridis, G.; Maloupa, E. In vitro propagation of medicinal and aromatic plants: The case of selected Greek species with conservation priority. In Vitro Cell. Dev. Biol. Plant. 2019, 55, 635–646. [Google Scholar] [CrossRef]
- Hartmann, H.; Kester, D.; Davies, T.; Geneve, R. Principios y Prácticas de Propagación de Plantas, 7th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2002; pp. 367–374. [Google Scholar]
- Maloupa, E.; Grigoriadou, K.; Papanastassi, K.; Krigas, N. Conservation; Propagation; Development and Utilization of Xe-rophytic Species of the Native Greek Flora towards Commercial Floriculture. Acta Hortic. 2008, 766, 205–214. [Google Scholar] [CrossRef]
- Bridgen, M.; Olate, E.; Schiappacasse, F. Flowering Geophytes from Chile. Acta Hortic. 2002, 570, 75–78. [Google Scholar] [CrossRef]
- Issac, M.; Kuriakose, P.; Leung, S.; Costa, A.B.; Johnson, S.; Bucalo, K.; Stober, J.M.; Determann, R.O.; Rogers, W.L.; Cruse-Sanders, J.M.; et al. Seed Cryopreservation; Germination; and Micropropagation of Eastern Turkeybeard; Xerophyllum asphodeloides (L.) Nutt.: A Threatened Species from the Southeastern United States. Plants 2021, 10, 1462. [Google Scholar] [CrossRef]
- Pullman, G.S.; Bai, K.; Hane, M.; Ruland, D.; Cruse-Sanders, J.M.; Boyd, R.S.; Johnson, S. Seed cryopreservation and mi-cropropagation of the federally threatened species; Price’s potato-bean (Apios priceana B.L. Robins.). In Vitro Cell. Dev. Biol. Plant 2019, 55, 558–568. [Google Scholar] [CrossRef]
- Santibáñez, F. Atlas Agroclimático de Chile. Estado Actual y Tendencias del Clima. Tomo III: Regiones de Valparaíso, Metropolitana, O’Higgins y Maule; Fundación para la Innovación Agraria: Santiago, Chile, 2017; p. 37. [Google Scholar]
- Munsell Color. Munsell Color Chart for Plant Tissues with Genuine Munsell Color Chips; Munsell Color Company: Grand Rapids, MI, USA, 2012; p. 9. [Google Scholar]
- ISTA. International Rules for Seed Testing. Determination of Moisture Content; Rules ISTA: Bassersdorf, Switzerland, 2016; Volume 9, p. 9-1. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Johansen, D.A. Plant Microtechnique; McGraw-Hill Book Company: New York, NY, USA, 1940. [Google Scholar]
- D’Ambrogio de Argüeso, A. Manual de Técnicas en Histología Vegetal; Hemisferio Sur: Buenos Aires, Argentina, 1986. [Google Scholar]
- Singh, D.; Mathur, S.B. Histopathology of Seed-Borne Infections, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2004; p. 296. [Google Scholar]
- Wagstaff, C.; Chanasut, U.; Harren, F.; Laarhoven, L.; Thomas, B.; Rogers, H.; Stead, A. Ethylene and flower longevity in Alstroemeria: Relationship between tepal senescence; abscission and ethylene biosynthesis. J. Exp. Bot. 2005, 56, 1007–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillaspy, G.; Ben-David, H.; Gruissem, W. Fruits: A developmental perspective. Plant Cell 1993, 5, 1439–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, R. Growth analysis; individual plants. In Encyclopaedia of Applied Plant Sciences; Thomas, B., Murphy, D.J., Murray, B.G., Eds.; Academic Press: London, UK, 2003; pp. 579–588. [Google Scholar]
- Araújo, J.; Ramalho, M.; Bruzi, A.; Abreu, N. Dry matter accumulation in genotypes of bean submitted to different levels of nitrogen. Ciênc. Agrotec. 2009, 33, 985–992. [Google Scholar] [CrossRef] [Green Version]
- Machuca, A.P. Efecto de la Estratificación de Embriones y Escarificación de Semillas Sobre el Crecimiento de Alstroemeria spp. in vitro. deTìtulo, M., Ed.; Universidad de Chile: Santiago, Chile, 2006. [Google Scholar]
- Vargas, M.; Jofré, E.; Navarrete, C.; Bravo, J.; Jamett, F.; Inostroza-Blancheteau, C.; Ibáñez, C. Sexual and asexual reproductive aspects of Leontochir ovallei; a rare and endangered geophyte of the Atacama Desert. Rev. Chil. Hist. Nat. 2018, 91, 5. [Google Scholar] [CrossRef] [Green Version]
- Geneve, R.L.; Baskin, C.C.; Baskin, J.M.; Gehan Jayasuriya, K.; Gama-Arachchige, N.S. Functional morpho-anatomy of water-gap complexes in physically dormant seed. Seed Sci. Res. 2018, 28, 186–191. [Google Scholar] [CrossRef]
- Martínez-Fernández, V.; Martínez-García, F.; Pérez-García, F. Census; reproductive biology; and germination of Astragalus gines-lopezii (Fabaceae); a narrow and endangered endemic species of SW Spain. Turk. J. Bot. 2014, 38, 686–695. [Google Scholar] [CrossRef] [Green Version]
- Statwick, J.M. Germination pretreatments to break hard-seed dormancy in Astragalus cicer L. (Fabaceae). Peer J. 2016, 4, e2621. [Google Scholar] [CrossRef] [Green Version]
- Mira, S.; Arnal, A.; Pérez-García, F. Seed germination of Phillyrea angustifolia L.; a species of difficult propagation. For. Syst. 2016, 26, e002. [Google Scholar] [CrossRef] [Green Version]
- Galíndez, G.; Lopez-Spahr, D.; Gomez, C.A.; Pastrana-Ignes, V.; Diaz, R.; Bertuzzi, T.; Ortega-Baes, P. Morpho-anatomical structure and physical dormancy breaking of Malvastrum coromandelianum ssp. Coromandelianum (Malvaceae) seeds. B. Soc. Argent. Bot. 2019, 54, 509–517. [Google Scholar] [CrossRef]
- Singh, N.; Gore, P.G.; Aravind, J. Breaking seed coat impermeability to aid conservation and utilization of wild Vigna species. Genet. Resour. Crop Evol. 2020, 67, 523–529. [Google Scholar] [CrossRef]
- Jaya Rathi, J.; Sasirekha, R.; Ranjith Kumar, R. Effect of physical and chemical treatments on breaking the seed dormancy of Caesalpinia bonduc (L.) Roxb. Plant Sci. Today 2021, 8, 572–577. [Google Scholar] [CrossRef]
- Illescas-Gallegos, E.; Rodríguez-Trejo, D.A.; Villanueva-Morales, A.; Borja-de La Rosa, M.A.; Ordóñez-Candelaria, V.R.; Ortega-Aragón, L.A. Factors influencing physical dormancy and its elimination in two legumes genus. Rev. Chapingo Ser. Cie. 2021, 27, 413–429. [Google Scholar]
- Schiappacasse, F.; Peñailillo, P.; Yáñez, P.; Bridgen, M. Propagation Studies on Chilean Geophytes. Acta Hortic. 2005, 673, 121–126. [Google Scholar] [CrossRef]
- Aros, D.; Rivas, C.; Toledo, M.; Céspedes, C.; Cortés, C.; Santander, M.; Vásquez, M.; Álvarez, N.; Donoso, A.; Prat, M. Chilean native species from the Andes Mountains: An attractive source of germplasm for ornamental plant breeding. Acta Hortic. 2017, 1171, 375–380. [Google Scholar] [CrossRef]
- Jansen, L.; Gebauer, J.; Wichern, F. Effect of different mechanical seed scarification methods on germination and emergence dynamics of baobab (Adansonia digitata L.). J. Agric. Rural Dev. Trop. 2021, 122, 183–192. [Google Scholar] [CrossRef]
- Filho, A.R.; Vesco, L.D.; Nodari, R.; Lischka, R.; Müller, C.; Guerra, M. Tissue culture for the conservation and mass propa-gation of Vriesea reitzii Leme and Costa; a bromeliad threatened of extinction from the Brazilian Atlantic Forest. Biodivers. Conserv. 2005, 14, 1799–1808. [Google Scholar] [CrossRef]
- Sarasan, V.; Cripps, R.; Ramsay, M.M.; Atherton, C.; McMichen, M.; Prendergast, G.; Rowntree, J.K. Conservation In vitro of threatened plants—Progress in the past decade. In Vitro Cell. Dev. Biol. Plant 2006, 42, 206–214. [Google Scholar] [CrossRef]
- Tyagi, R.K.; Agrawal, A.; Mahalakshmi, C.; Hussain, Z.; Tyagi, H. Low-cost media for in vitro conservation of turmeric (Curcuma longa L.) and genetic stability assessment using RAPD markers. In Vitro Cell. Dev. Biol. Plant 2007, 43, 51–58. [Google Scholar] [CrossRef]
- Engelmann, F. Use of biotechnologies for the conservation of plant biodiversity. In Vitro Cell. Dev. Biol. Plant 2010, 47, 5–16. [Google Scholar] [CrossRef]
- Reed, B.M.; Sarasan, V.; Kane, M.; Bunn, E.; Pence, V.C. Biodiversity conservation and conservation biotechnology tools. In Vitro Cell. Dev. Biol. Plant 2011, 47, 1–4. [Google Scholar] [CrossRef]
- Pence, V.C.; Ballesteros, D.; Walters, C.; Reed, B.M.; Philpott, M.; Dixon, K.W.; Pritchard, H.W.; Culley, T.M.; Vanhove, A.C. Cryobiotechnologies: Tools for expanding long-term ex situ conservation to all plant species. Biol. Conserv. 2020, 250, 108736. [Google Scholar] [CrossRef]
- Hussain, A.; Qarshi, I.A.; Nazir, H.; Ullah, I. Plant tissue culture: Current status and opportunities. In Recent Advances in Plant In Vitro Culture; Leva, A., Rinaldi, L., Eds.; IntechOpen: London, UK, 2012. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reproductive Stage | Duration | Characteristics (Mean ± Standard Deviation) |
|---|---|---|
| B1: Initiation of flower bud development | 11 days | Length of flower bud 10.92 ± 1.24 mm; width 7.67 ± 1.30 mm |
| B2: Flower bud | 2 days | Length of flower bud 27.76 ± 2.13 mm; width 16.48 ± 1.32 mm |
| B3: Flower bud opening | 3 days | Length of flower bud 45.92 ± 1.85 mm; width 20.89 ± 1.28 mm |
| B4: Flowering-Anthesis | 9 days | Length 97.23 ± 4.85 mm, upper outer tepal to lower outer tepal; width 45.36 ± 2.18 mm |
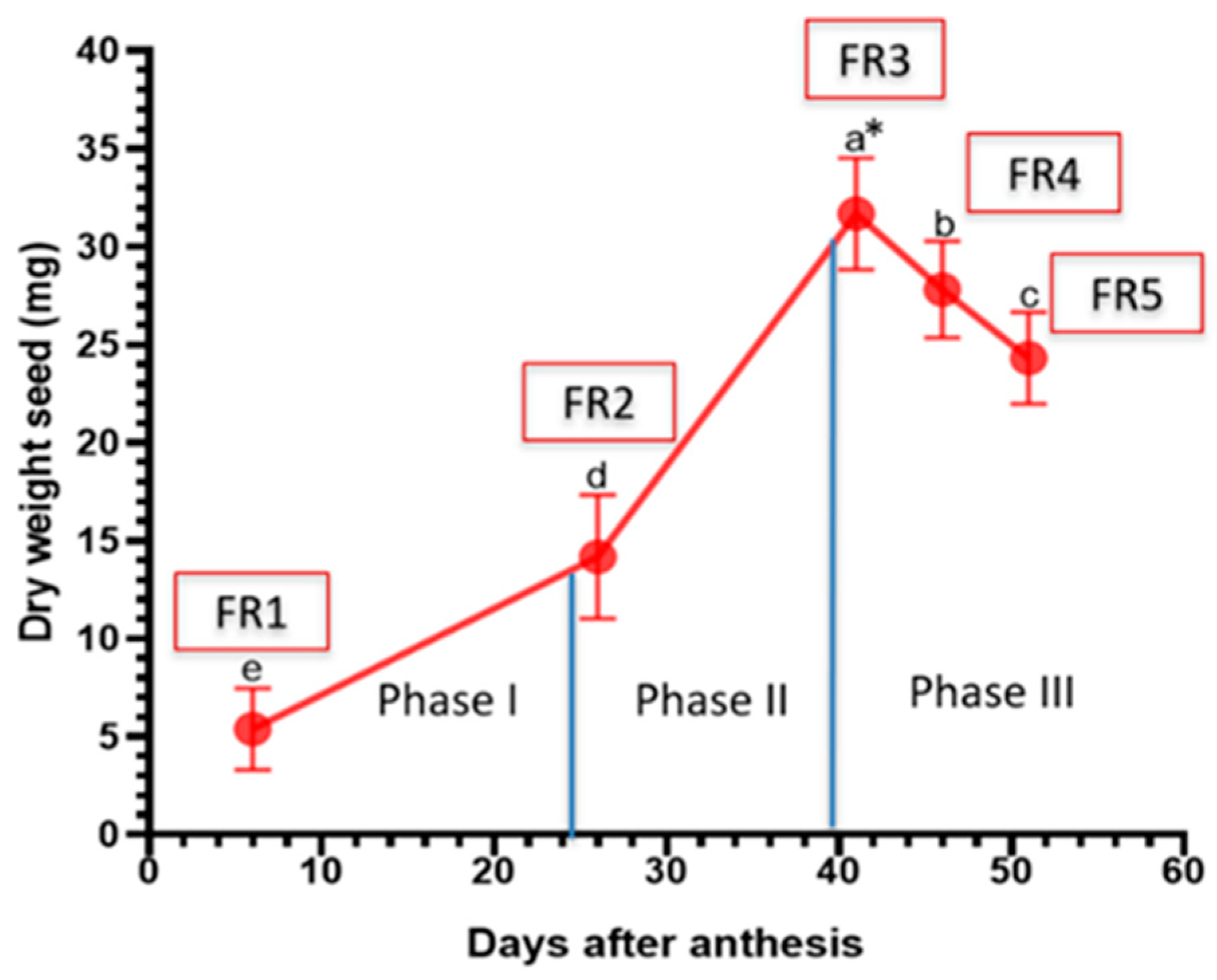
| FR1: Fruit stage 1 | 6 days | Fruit length 17.65± 2.21 mm; width 14.65 ± 1.12 mm |
| FR2: Fruit stage 2 | 20 days | Fruit length 21.01 ± 1.68 mm; width 16.24 ± 1.03 mm |
| FR3: Fruit stage 3 | 15 days | Fruit length 23.88 ± 1.36 mm; width 16.86 ± 1.45 mm |
| FR4: Fruit stage 4 | 5 days | Fruit length 20.25 ± 1.96 mm; width 16.62 ± 1.33 mm |
| FR5: Fruit stage 5 | 5 days | Fruit length 19.28 mm ± 1.78 mm; width 16.13 ± 0.73 mm |
| Phenological Stages of A. pelegrina Fruit Development | Fruit Color |
|---|---|
| Fruit stage 1 (FR1) | Green (5GY 7/10) with white spots |
| Fruit stage 2 (FR2) | Green (5GY 7/8) with white spots and 5 burgundy veins (2.5R 5/10) |
| Fruit stage 3 (FR3) | Yellow-green (2.5GY 8/8) with 5 burgundy veins (5R 4/10) |
| Fruit stage 4 (FR4) | Yellow (5Y 8/8) with 5 burgundy veins (10R 4/8) extending to the middle of the fruit |
| Fruit stage 5 (FR5) | Brown (2.5YR 7/6) with 5 brown veins (7.5YR 5/2) |
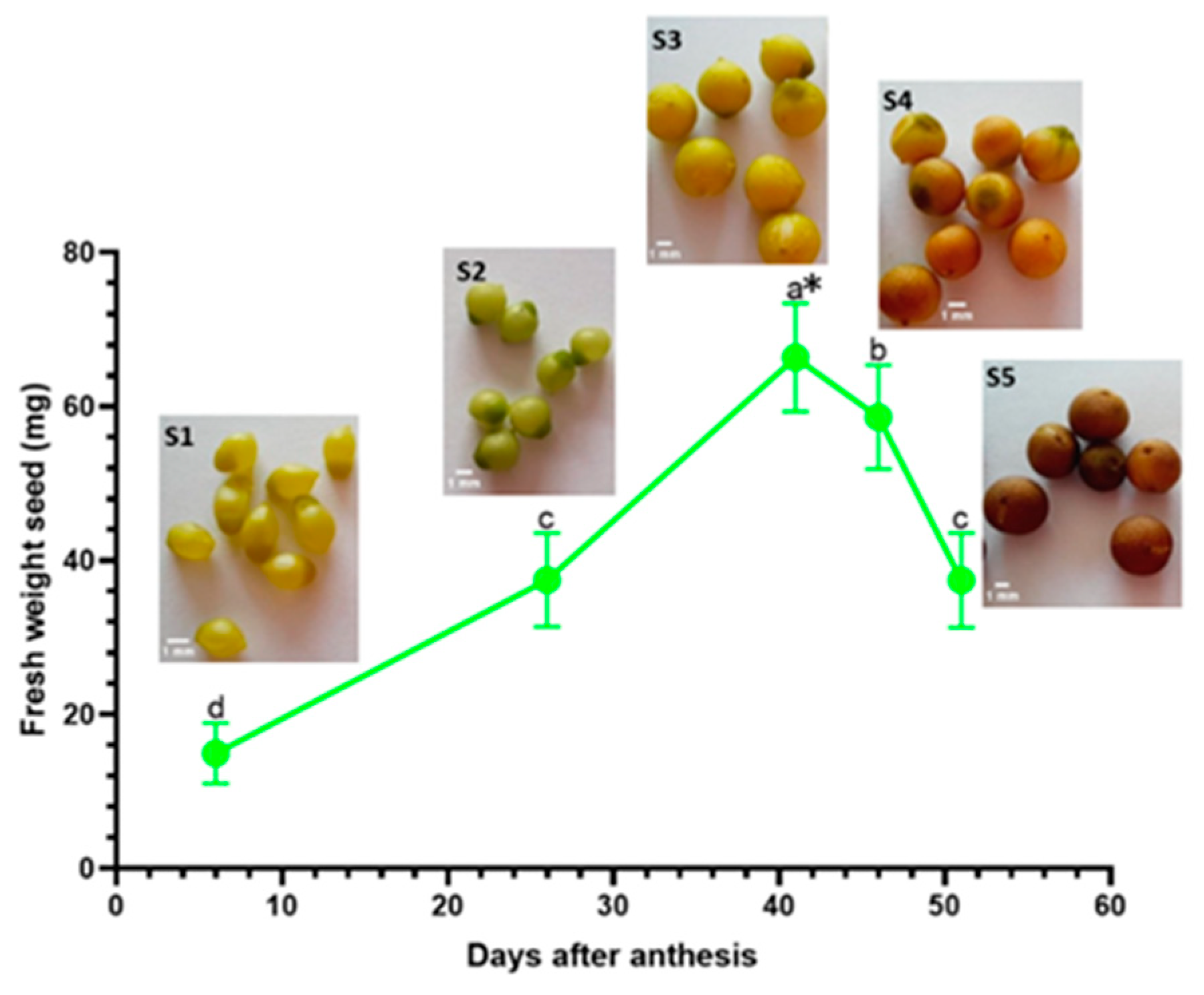
| Developmental Stage | Seed Color | Mean Fresh Weight | Mean Dry Weight |
|---|---|---|---|
| Seed stage 1 (S1) | Lime-green seed color (2.5GY 8/12) | 14.97 ± 3.9 mg | 5.38 ± 2.1 mg |
| Seed stage 2 (S2) | Green seed color (2.5GY 7/10) | 37.48 ± 6.1 mg | 14.18 ± 3.2 mg |
| Seed stage 3 (S3) | Green seed color (2.5GY 8/10) | 66.37 ± 7.0 mg | 31.70 ± 2.8 mg |
| Seed stage 4 (S4) | Brown seed color (2.5Y 6/8) | 58.61 ± 6.8 mg | 27.83 ± 2.5 mg |
| Seed stage 5 (S5) | Dark brown seed color (7.5YR 4/4) | 37.44 ± 6.1 mg | 24.38 ± 2.4 mg |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerra, F.; Peñaloza, P.; Vidal, A.; Cautín, R.; Castro, M. Seed Maturity and Its In Vitro Initiation of Chilean Endemic Geophyte Alstroemeriapelegrina L. Horticulturae 2022, 8, 464. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8050464
Guerra F, Peñaloza P, Vidal A, Cautín R, Castro M. Seed Maturity and Its In Vitro Initiation of Chilean Endemic Geophyte Alstroemeriapelegrina L. Horticulturae. 2022; 8(5):464. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8050464
Chicago/Turabian StyleGuerra, Francesca, Patricia Peñaloza, Alexis Vidal, Ricardo Cautín, and Mónica Castro. 2022. "Seed Maturity and Its In Vitro Initiation of Chilean Endemic Geophyte Alstroemeriapelegrina L." Horticulturae 8, no. 5: 464. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8050464