
Effects of Preharvest Aminoethoxyvinylglycine (AVG) Treatment on Fruit Ripening, Core Browning and Related Gene Expression in ‘Huangguan’ Pear (Pyrus bretschneideri Rehd.)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Treatments
2.2. Determination of Fruit Quality
2.3. Estimation of the Core Browning Index
2.4. Determination of Ethylene Production Rate
2.5. Determination of CGA Content
2.6. Determination of Polyphenol Oxidase (PPO) Activity and Malondialdehyde (MDA) Content
2.7. RNA Extraction and Quantitative RT-PCR (qRT-PCR) Analysis
2.8. Statistical Analysis
3. Results
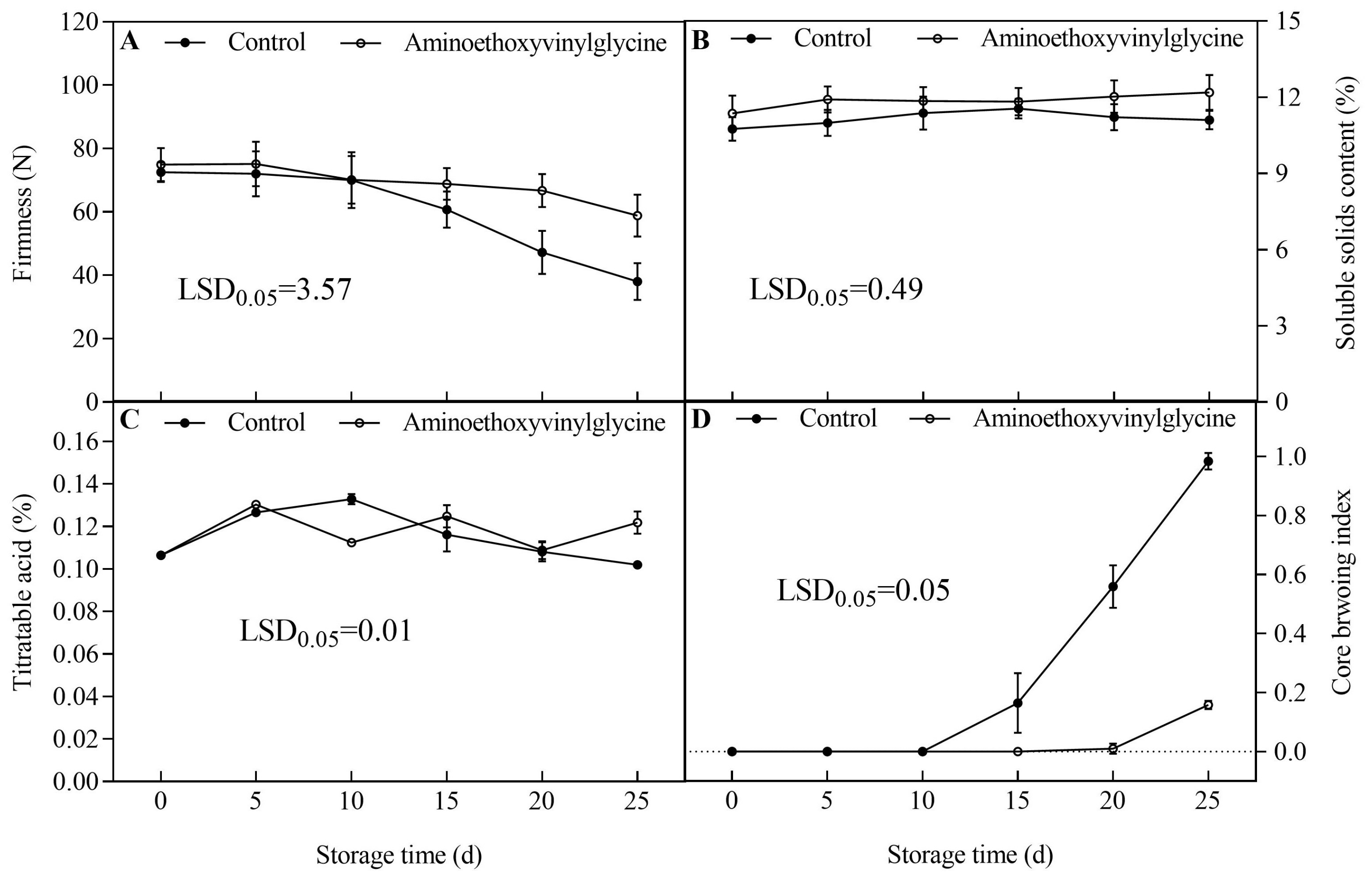
3.1. Effects of AVG on the Fruit Quality and Core Browning
3.2. Effect of AVG on Ethylene Production Rate
3.3. Effect of AVG on CGA and MDA Contents and PPO Activity in the Core Tissue
3.4. Effect of AVG on the Expression of Genes Associated with Ethylene Biosynthesis and Signal Transduction in Core Tissue
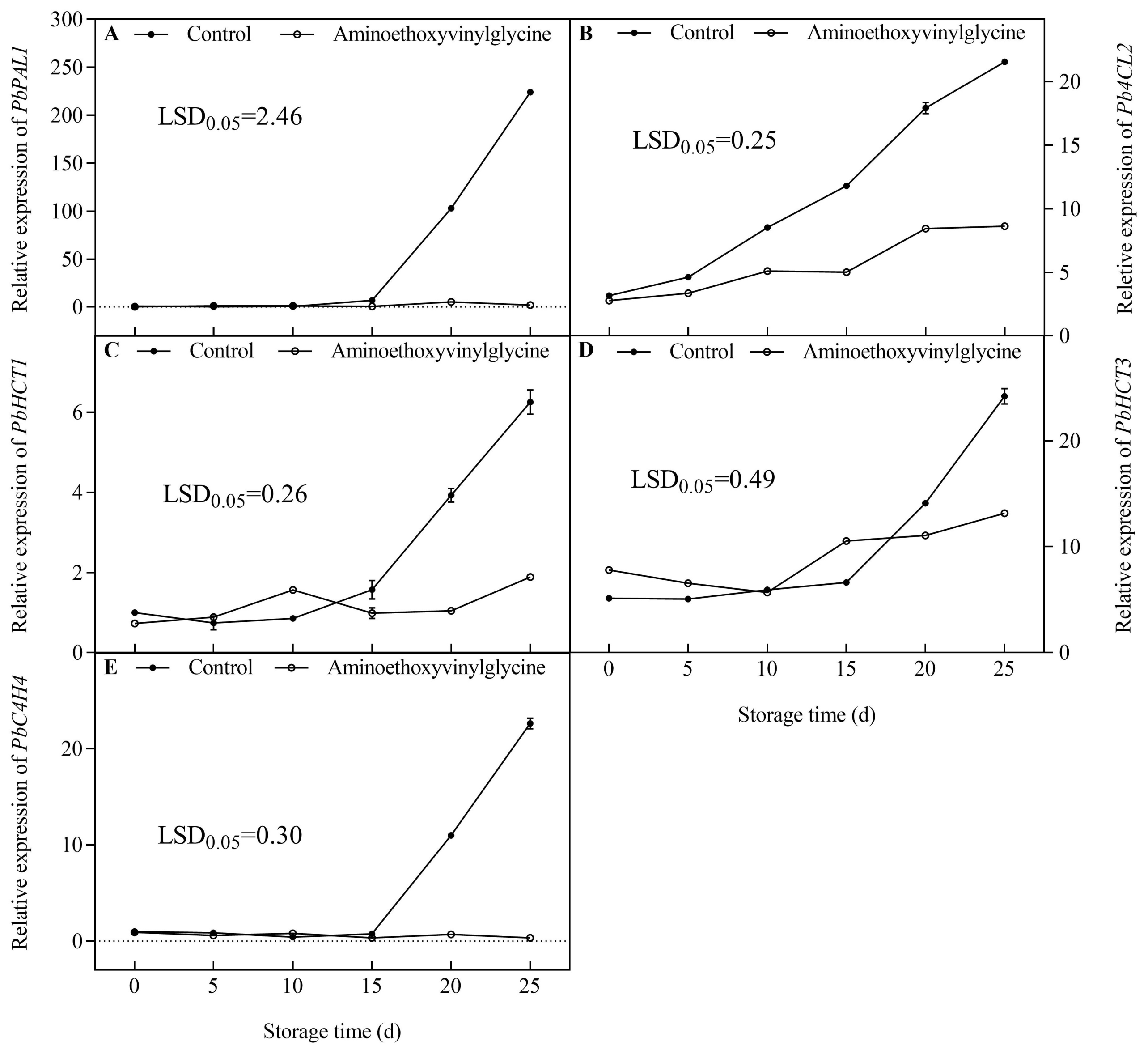
3.5. Effect of AVG on the Expression of Genes Associated with CGA Biosynthesis in the Core Tissue
3.6. Effect of AVG on the Expression of the PbPPO1 and PbPPO5 Genes in the Core Tissue
3.7. Effect of AVG on the Expression of PbLOX1, PbLOX5 and PbPLD1 in the Core Tissue
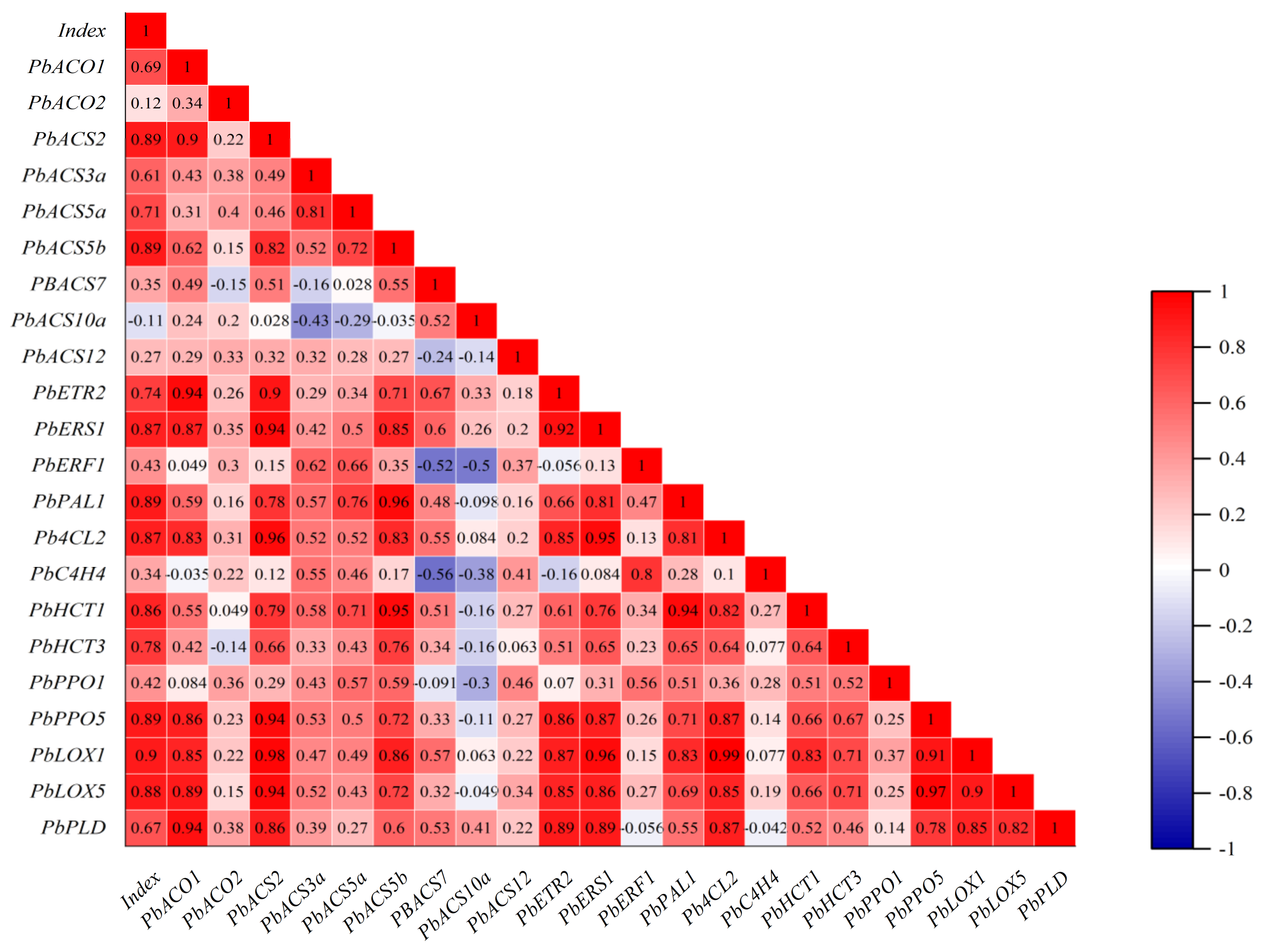
3.8. Correlation Analysis of Core Browning Index with Gene Expression Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kende, H. Ethylene Biosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1993, 44, 283–307. [Google Scholar] [CrossRef]
- Yu, Y.B.; Yang, S.F. Auxin-induced ethylene production and its inhibition by aminoethoxyvinylglycine and cobalt ion. Plant Physiol. 1979, 64, 1074–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Autio, W.R.; Bramlage, W.J. Effects of AVG on maturation, ripening, and storage of apples. J. Am. Soc. Hortic. Sci. 1982, 107, 1074–1077. [Google Scholar] [CrossRef]
- Romani, R.; Puschmann, R.; Finch, J.; Beutel, J. Effects of preharvest applications of AVG on ripening of ‘Bartlett’ pears with and without cold storage. HortScience 1982, 17, 214–215. [Google Scholar] [CrossRef]
- Romani, R.; Labavitch, J.; Yamashita, T.; Hess, B.; Rae, H. Preharvest AVG treatment of ‘Bartlett’ pear fruits: Effects on ripening, color change, and volatiles. J. Am. Soc. Hortic. Sci. 1983, 108, 1046–1049. [Google Scholar] [CrossRef]
- Halder-Doll, H.; Bangerth, F. Inhibition of autocatalytic C2H4-biosynthesis by AVG applications and consequences on the physiological behavior and quality of apple fruits in cool storage. Sci. Hortic. 1987, 33, 87–96. [Google Scholar] [CrossRef]
- Bregoli, A.M.; Scaramagli, S.; Costa, G.; Sabatini, E.; Ziosi, V.; Biondi, S.; Torrigiani, P. Peach (Prunus persica) fruit ripening: Aminoethoxyvinylglycine (AVG) and exogenous polyamines affect ethylene emission and flesh firmness. Physiol. Plant. 2002, 114, 472–481. [Google Scholar] [CrossRef]
- Torrigiani, P.; Bregoli, A.M.; Ziosi, V.; Scaramagli, S.; Ciriaci, T.; Rasori, A.; Biondi, S.; Costa, G. Pre-harvest polyamine and aminoethoxyvinylglycine (AVG) applications modulate fruit ripening in stark red gold nectarines (Prunus persica l. batsch). Postharvest Biol. Technol. 2004, 33, 293–308. [Google Scholar] [CrossRef]
- Schupp, J.R.; Greene, D.W. Effect of aminoethoxyvinylglycine (AVG) on preharvest drop, fruit quality, and maturation of ‘McIntosh’ apples. I. Concentration and timing of dilute applications of AVG. HortScience 2004, 39, 1030–1035. [Google Scholar] [CrossRef] [Green Version]
- Rath, A.C.; Kang, I.K.; Park, C.H.; Yoo, W.J.; Byun, J.K. Foliar application of aminoethoxyvinylglycine (AVG) delays fruit ripening and reduces pre-harvest fruit drop and ethylene production of bagged “Kogetsu” apples. Plant Growth Regul. 2006, 50, 91–100. [Google Scholar] [CrossRef]
- Ziosi, V.; Bregoli, A.M.; Bonghi, C.; Fossati, T.; Biondi, S.; Costa, G.; Torrigiani, P. Transcription of ethylene perception and biosynthesis genes is altered by putrescine, spermidine and aminoethoxyvinylglycine (AVG)during ripening in peach fruit (Prunus persica). New Phytol. 2006, 172, 229–238. [Google Scholar] [CrossRef]
- Clayton, M.; Biasi, W.V.; Southwick, S.M.; Mitcham, E.J. ReTain™ affects maturity and ripening of ‘Bartlett’ Pear. HortScience 2006, 35, 1294–1299. [Google Scholar] [CrossRef] [Green Version]
- D’Aquino, S.; Schirra, M.; Molinu, M.G.; Tedde, M.; Palma, A. Preharvest aminoethoxyvinylglycine treatments reduce internal browning and prolong the shelf-life of early ripening pears. Sci. Hortic. 2010, 125, 353–360. [Google Scholar] [CrossRef]
- Lee, U.Y.; Oh, K.S.; Choi, J.H.; Ahn, Y.J.; Chun, J.P. Changes of fruit quality and reduction of physiological disorders during shelf-life in early season pear (Pyrus pyrifolia Nakai) fruits treated with aminoethoxyvinylglycine. Korean J. Hortic. Sci. 2014, 32, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xie, X.B.; Song, J.K. Preharvest aminoethoxyvinylglycine spray efficacy in improving storability of ‘Bartlett’ pears is affected by application rate, timing, and fruit harvest maturity. Postharvest Biol. Technol. 2016, 119, 69–76. [Google Scholar] [CrossRef]
- Xie, X.B.; Einhorn, T.; Wang, Y. Inhibition of ethylene biosynthesis and associated gene expression by aminoethoxyvinylglycine and 1-methylcyclopropene and their consequences on eating quality and internal browning of ‘Starkrimson’ pears. J. Am. Soc. Hortic. Sci. 2015, 140, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Xylia, P.; Chrysargyris, A.; Ahmed, Z.F.R.; Tzortzakis, N. Application of rosemary and eucalyptus essential oils and their main component on the preservation of apple and pear fruits. Horticulturae 2021, 7, 479. [Google Scholar] [CrossRef]
- Wang, L.C.; Li, H.; Ecker, J.R. Ethylene biosynthesis and signaling networks. Plant Cell 2002, 14 (Suppl. 1), S51–S131. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Ecker, J.R. The ethylene signaling pathway: New insights. Curr. Opin. Plant Biol. 2004, 7, 40–49. [Google Scholar] [CrossRef]
- Alonso, J.M.; Stepanova, A.N. The ethylene signaling pathway. Science 2004, 306, 1513–1515. [Google Scholar] [CrossRef]
- Zdarska, M.; Cuyacot, A.R.; Tarr, P.T.; Yamoune, A.; Szmitkowska, A.; Hrdinová, V.; Gelová, Z.; Meyerowitz, E.M.; Hejátko, J. ETR1 integrates response to ethylene and cytokinins into a single multistep phosphorelay pathway to control root growth. Mol. Plant 2019, 12, 1338–1352. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.; Yu, J.P.; Chen, F.; Zhao, T.J.; Fang, X.H.; Li, Y.Q.; Sui, S.F. TINY, a dehydration-responsive element (DRE)-binding protein-like transcription factor connecting the DRE- and ethylene-responsive element-mediated signaling pathways in Arabidopsis. J. Biol. Chem. 2008, 283, 6261–6271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.C.; Kuang, J.F.; Chen, J.Y.; Liu, X.C.; Xiao, Y.Y.; Fu, C.C.; Wang, J.N.; Wu, K.Q.; Lu, W.J. Banana transcription factor MaERF11 recruits histone deacetylase MaHDA1 and represses the expression of MaACO1 and expansins during fruit ripening. Plant Physiol. 2016, 171, 1070–1084. [Google Scholar] [CrossRef] [Green Version]
- Yao, G.; Ming, M.; Allan, A.C.; Gu, C.; Li, L.; Wu, X.; Wang, R.; Chang, Y.; Qi, K.; Zhang, S.; et al. Map-based cloning of the pear gene MYB114 identifies an interaction with other transcription factors to coordinately regulate fruit anthocyanin biosynthesis. Plant J. 2017, 92, 437–451. [Google Scholar] [CrossRef] [Green Version]
- Ni, J.; Bai, S.; Zhao, Y.; Qian, M.; Tao, R.; Yin, L.; Gao, L.; Teng, W. Ethylene response factors Pp4ERF24 and Pp12ERF96 regulate blue light-induced anthocyanin biosynthesis in ‘Red Zaosu’ pear fruits by interacting with MYB114. Plant Mol. Biol. 2019, 99, 67–78. [Google Scholar] [CrossRef]
- Ahmed, Z.F.R.; Palta, J.P. A Postharvest dip treatment with lysophosphatidylethanolamine, a natural phospholipid, may retard senescence and improve the shelf life of banana fruit. HortScience 2015, 50, 1035–1040. [Google Scholar] [CrossRef] [Green Version]
- Yue, P.; Wang, Y.; Bu, H.; Li, X.; Yuan, H.; Wang, A. Ethylene promotes IAA reduction through PuERFs-activated PuGH3.1 during fruit ripening in pear (Pyrus ussuriensis). Postharvest Biol. Technol. 2019, 157, 110955. [Google Scholar] [CrossRef]
- Lindo-García, V.; Giné-Bordonaba, J.; Vall-Llaura, N.; Duaigües, E.; Larrigaudière, C. Unravelling the cold-induced regulation of ethylene and α-farnesene and its involvement with the development of scald-like disorders in different pear cultivars. Postharvest Biol. Technol. 2020, 174, 111441. [Google Scholar] [CrossRef]
- Amiot, M.J.; Tacchini, M.; Aubert, S.; Nicolas, J. Phenolic composition and browning susceptibility of various apple cultivars at maturity. J. Food Sci. 1992, 57, 958–962. [Google Scholar] [CrossRef]
- Franck, C.; Lammertyn, J.; Ho, Q.T.; Verboven, P.; Verlinden, B.; NicolaIï, B.M. Browning disorders in pear fruit. Postharvest Biol. Technol. 2007, 43, 1–13. [Google Scholar] [CrossRef]
- Cheng, Y.D.; Liu, L.Q.; Zhao, G.Q.; Shen, C.G.; Yan, H.B.; Guan, J.F.; Yang, K. The effects of modified atmosphere packaging on core browning and the expression patterns of PPO and PAL genes in ‘Yali’ pears during cold storage. LWT Food Sci. Technol. 2015, 60, 1243–1248. [Google Scholar] [CrossRef]
- Dong, Y.; Liu, L.Q.; Zhao, Z.; Zhi, H.H.; Guan, J.F. Effects of 1-MCP on reactive oxygen species, polyphenol oxidase activity, and cellular ultra-structure of core tissue in ‘Yali’ pear (Pyrus bretschneideri Rehd.) during storage. Hortic. Environ. Biotechnol. 2015, 56, 207–215. [Google Scholar] [CrossRef]
- Zhou, H.W.; Feng, X. Polyphenol oxidase from Yali pear Pyrus bretschneider. J. Sci. Food Agric. 1991, 57, 307–313. [Google Scholar] [CrossRef]
- He, J.G.; Cheng, Y.D.; Guan, J.F.; Ge, W.Y.; Zhao, Z. Changes of chlorogenic acid content and its synthesis-associated genes expression in Xuehua pear fruit during development. J. Integr. Agric. 2017, 16, 471–477. [Google Scholar] [CrossRef] [Green Version]
- Feys, M.; Naesens, W.; Tobback, P.; Maes, E. Lipoxygenase activity in apples in relation to storage and physiological disorders. Phytochemistry 1980, 19, 1009–1011. [Google Scholar] [CrossRef]
- Sun, J.; You, X.R.; Li, L.; Peng, H.X.; Su, W.Q.; Li, C.B.; He, Q.G.; Liao, F. Effects of a phospholipase D inhibitor on postharvest enzymatic browning and oxidative stress of litchi fruit. Postharvest Biol. Technol. 2011, 62, 288–294. [Google Scholar] [CrossRef]
- Sun, J.; Li, C.B.; Prasad, K.N.; You, X.R.; Li, L.; Liao, F.; Peng, H.X.; He, X.M.; Li, Z.C.; Zhang, Y.Y. Membrane deterioration, enzymatic browning and oxidative stress in fresh fruits of three litchi cultivars during six-day storage. Sci. Hortic. 2012, 148, 97–103. [Google Scholar] [CrossRef]
- Sheng, L.; Zhou, X.; Liu, Z.Y.; Wang, J.W.; Zhou, Q.; Wang, L.; Zhang, Q.; Ji, S.J. Changed activities of enzymes crucial to membrane lipid metabolism accompany pericarp browning in ‘Nanguo’ pears during refrigeration and subsequent shelf life at room temperature. Postharvest Biol. Technol. 2016, 117, 1–8. [Google Scholar] [CrossRef]
- Hong, K.; Zhang, L.B.; Zhan, R.L.; Huang, B.Y.; Song, K.H.; Jia, Z.W. Identification and characterization of phospholipase D genes putatively involved in internal browning of pineapple during postharvest storage. Front. Plant Sci. 2017, 8, 913. [Google Scholar] [CrossRef]
- Wang, J.W.; Lv, M.; Li, G.D.; Jiang, Y.G.; Fu, W.W.; Zhang, L.; Ji, S.J. Effect of intermittent warming on alleviation of peel browning of ‘Nanguo’ pears by regulation energy and lipid metabolisms after cold storage. Postharvest Biol. Technol. 2018, 142, 99–106. [Google Scholar] [CrossRef]
- Feng, Y.X.; Cheng, Y.D.; He, J.G.; Li, L.M.; Guan, Y.Q.; Guan, J.F. Effect of MAP combined with 1-MCP on quality of ‘Huangguan’ pear fruit under normal temperature storage. Storage Process 2017, 17, 1–5. (In Chinese) [Google Scholar] [CrossRef]
- Cheng, Y.D.; Liu, L.Q.; Feng, Y.X.; Dong, Y.; Guan, J.F. Effects of 1-MCP on fruit quality and core browning in ‘Yali’pear during cold storage. Sci. Hortic. 2019, 243, 350–356. [Google Scholar] [CrossRef]
- Yuan, G.F.; Sun, B.; Yuan, J.; Wang, Q.M. Effect of 1-methylcyclopropene on shelf life, visual quality, antioxidant enzymes and health-promoting compounds in broccoli florets. Food Chem. 2010, 118, 774–781. [Google Scholar] [CrossRef]
- Malnoy, M.; Reynoird, J.P.; Mourgues, F.; Chevreau, E.; Simoneau, P. A method for isolating total RNA from pear leaves. Plant Mol. Biol. Rep. 2001, 19, 69–74. [Google Scholar] [CrossRef]
- Yuan, H.; Yue, P.T.; Bu, H.D.; Han, D.G.; Wang, A.D. Genome-wide analysis of ACO and ACS genes in pear (Pyrus ussuriensis). In Vitro Cell. Dev. Biol. Plant 2020, 56, 193–199. [Google Scholar] [CrossRef]
- Cin, D.V.; Danesin, M.; Botton, A.; Boschetti ADorigoni, A.; Ramina, A. Ethylene and preharvest drop: The effect of AVG and NAA on fruit abscission in apple (Malus domestica L. Borkh). Plant Growth Regul. 2008, 56, 317–325. [Google Scholar] [CrossRef]
- Whitaker, B.D. Membrane lipid metabolism and oxidative stress involved in postharvest ripening, senescence, and storage disorders of fruits. Acta Hortic. 2012, 945, 269–282. [Google Scholar] [CrossRef]
- Pasquariello, M.S.; Di Patre, D.; Mastrobuoni, F.; Zampella, L.; Scortichini, M.; Petriccione, M. Influence of postharvest chitosan treatment on enzymatic browning and antioxidant enzyme activity in sweet cherry fruit. Postharvest Biol. Technol. 2015, 109, 45–56. [Google Scholar] [CrossRef]
- Jannatizadeh, A. Exogenous melatonin applying confers chilling tolerance in pomegranate fruit during cold storage. Sci. Hortic. 2019, 246, 544–549. [Google Scholar] [CrossRef]
- Gao, H.; Zhang, Z.K.; Lv, X.G.; Cheng, N.; Peng, B.Z.; Cao, W. Effect of 24-epibrassinolide on chilling injury of peach fruit in relation to phenolic and proline metabolisms. Postharvest Biol. Technol. 2016, 111, 390–397. [Google Scholar] [CrossRef]
- Lin, Y.F.; Lin, H.T.; Zhang, S.; Chen, Y.H.; Jiang, X.J. The roles of metabolism of membrane lipids and phenolics in hydrogen peroxide-induced pericarp browning of harvested longan fruit. Postharvest Biol. Technol. 2016, 111, 53–61. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, J.; Feng, Y.; Cheng, Y.; Guan, J. Effects of Preharvest Aminoethoxyvinylglycine (AVG) Treatment on Fruit Ripening, Core Browning and Related Gene Expression in ‘Huangguan’ Pear (Pyrus bretschneideri Rehd.). Horticulturae 2023, 9, 179. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae9020179
He J, Feng Y, Cheng Y, Guan J. Effects of Preharvest Aminoethoxyvinylglycine (AVG) Treatment on Fruit Ripening, Core Browning and Related Gene Expression in ‘Huangguan’ Pear (Pyrus bretschneideri Rehd.). Horticulturae. 2023; 9(2):179. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae9020179
Chicago/Turabian StyleHe, Jingang, Yunxiao Feng, Yudou Cheng, and Junfeng Guan. 2023. "Effects of Preharvest Aminoethoxyvinylglycine (AVG) Treatment on Fruit Ripening, Core Browning and Related Gene Expression in ‘Huangguan’ Pear (Pyrus bretschneideri Rehd.)" Horticulturae 9, no. 2: 179. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae9020179