

The Involvement of Cell-Type-Specific Glycans in Hydra Temporary Adhesion Revealed by a Lectin Screen


Abstract
:1. Introduction
2. Methods
2.1. Hydra Culture
2.2. Whole-Mount Lectin Labeling
2.3. Footprint Lectin Labeling
2.4. Hydra and Corresponding Footprint Labeling after Voluntary and Forced Detachment
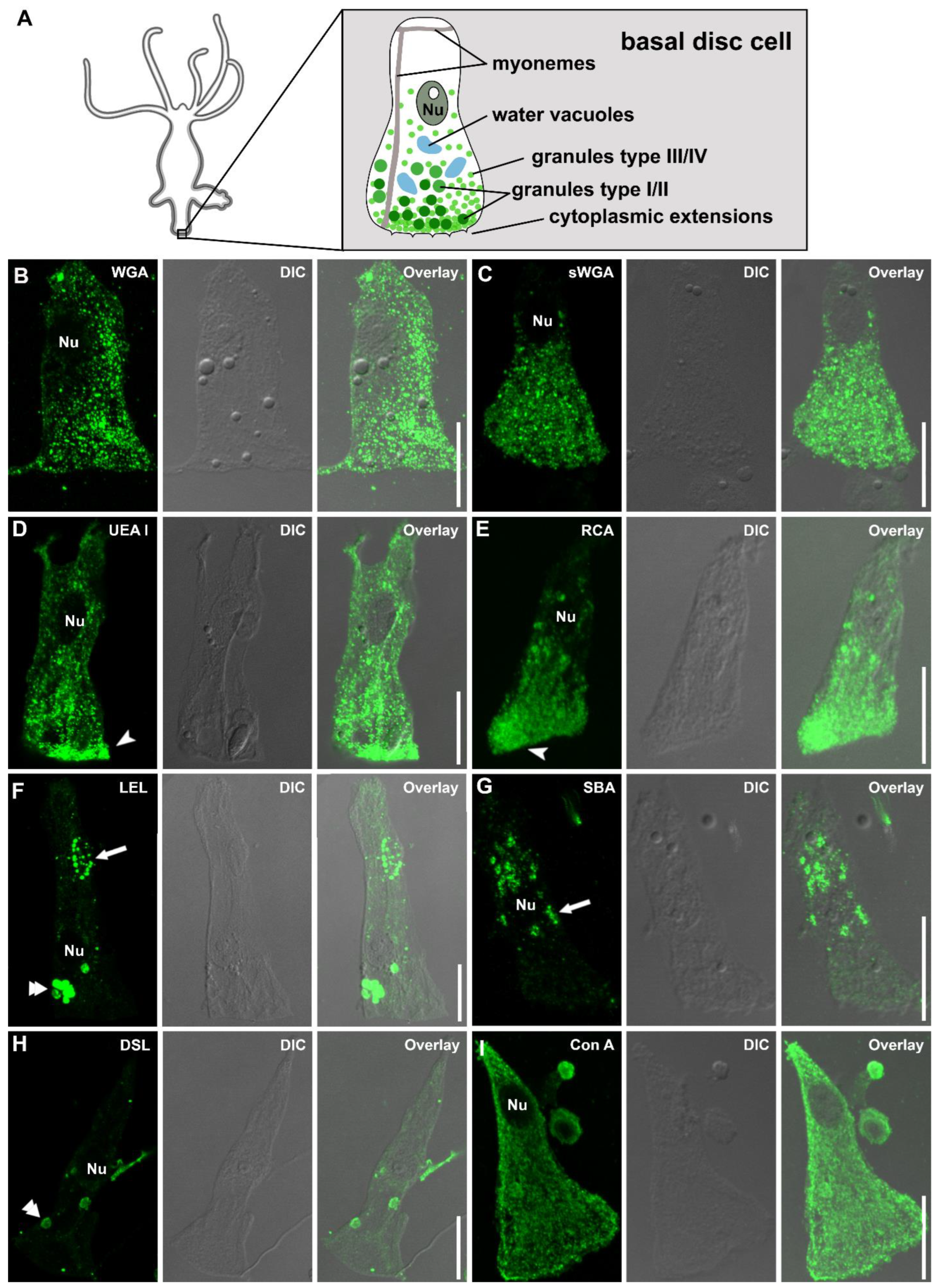
2.5. Single Basal Disc Cells Lectin Labeling
3. Results
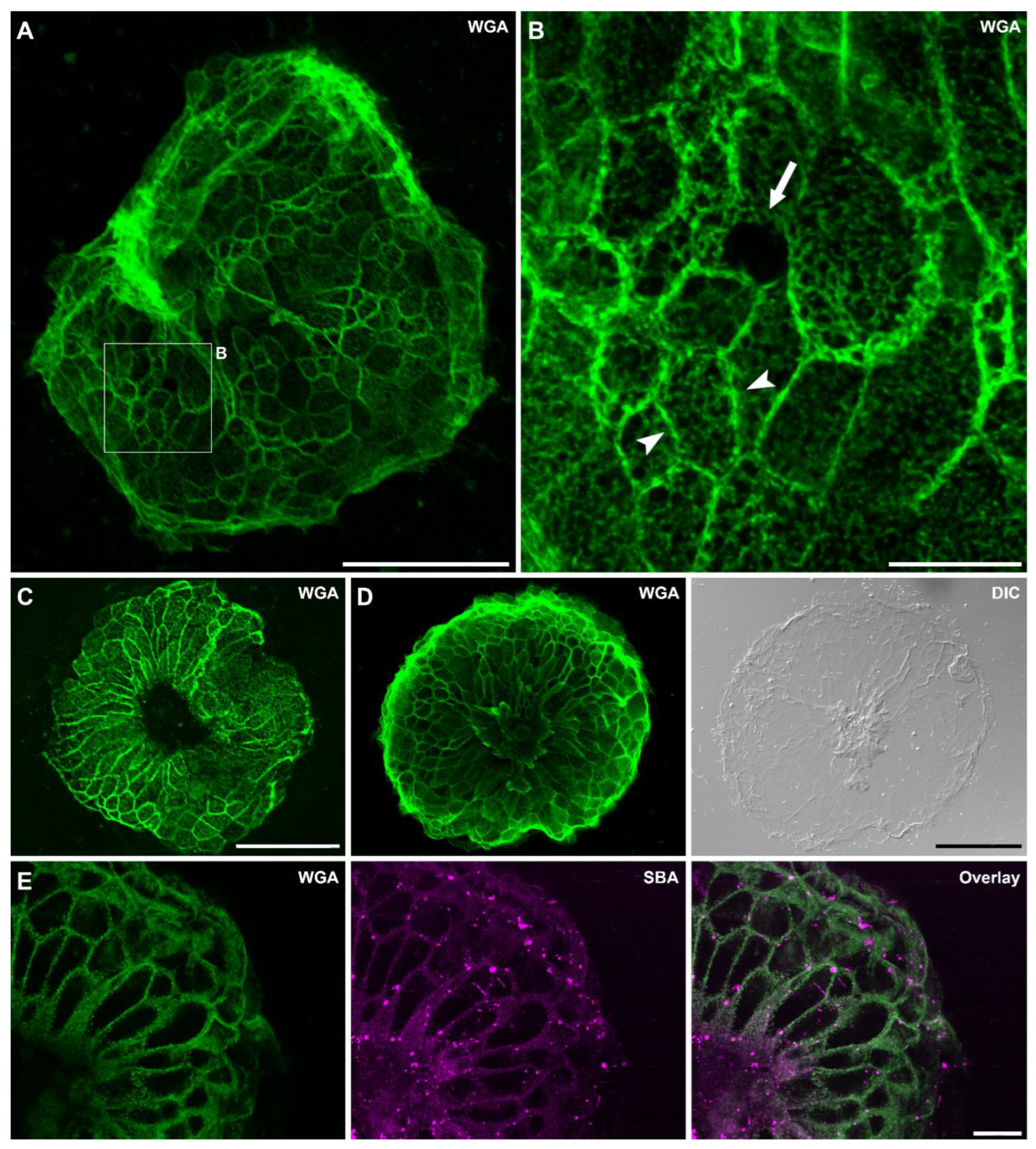
3.1. Lectin Labeling of Hydra Tissues and Footprint Secretions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gastric Region | Tentacles | Foot | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lectin | Acronym | Fixative | Endoderm | Overall Ectoderm Surface | Nematocytes, Nematoblasts or Vacuoles | Tentacle Surface | Nematocytes | Basal Disc Surface | Basal Disc Cells | Footprints | ||
| Capsules | Opercolum | Cnidocil | ||||||||||
| Wheat germ agglutinin | WGA | o.n PFA | +++ | +++ g | ++ | ++ | ++ | ++ | ++ h | +++ | ||
| 1 h PFA | +++ | +++ g | ++ | ++ | ++ | ++ | ||||||
| 4 h Lavdwosky | +++ | +++ g | ++ | ++ | ++ | ++ | ||||||
| Succinylated wheat germ agglutinin | sWGA | o.n PFA | +++ | ++ g | +++ | ++ | ++ | ++ h | +++ | |||
| 1 h PFA | +++ | ++ g | +++ | ++ | ++ | |||||||
| 4 h Lavdwosky | +++ | ++ g | +++ | ++ | ++ | |||||||
| Datura Stramonium lectin | DSL | o.n PFA | ++ g | ++ | ++ | ++ | + i | |||||
| 1 h PFA | + | ++ g | ++ | ++ | ||||||||
| 4 h Lavdwosky | ++ g | ++ g | ++ | ++ | ||||||||
| Lycopersicon esculentum (tomato) lectin | LEL | o.n PFA | +++ | + | +++ | |||||||
| 1 h PFA | +++ g | +++ g | +++ | ++ | ||||||||
| 4 h Lavdwosky | +++ b | +++ | ||||||||||
| Soybean agglutinin | SBA | o.n PFA | ++ g | ++ g | +++ | +++ | +++ | +++ | ||||
| 1 h PFA | ++ g | ++ g | +++ | +++ | ||||||||
| 4 h Lavdwosky | ++ g | ++ g | +++ | +++ | ||||||||
| Ricinus communis agglutinin | RCA | o.n PFA | + | + | +++ h | + i | ||||||
| 1 h PFA | + | + | ||||||||||
| 4 h Lavdwosky | + | ++ | ||||||||||
| Ulex europaeus agglutinin I | UEA I | o.n PFA | +++ | +++ h | + i | |||||||
| 1 h PFA | +++ | |||||||||||
| 4 h Lavdwosky | +++ | ++ f | ++ | |||||||||
| Concanavaline A | Con A | o.n PFA | + | +++ | + | ++ | +++ | ++ | +++ | ++ | ||
| 1 h PFA | + | +++ | + | ++ | +++ | ++ | ||||||
| 4 h Lavdwosky | + | +++ | + | ++ | +++ | ++ | ||||||
| Erythrina cristagalli lectin | ECL | o.n PFA | +++ g | x | ||||||||
| 1 h PFA | ++ g | ++ g | +++ g | |||||||||
| 4 h Lavdwosky | ++ g | +++ | ++ g | +++ g | ||||||||
| Pisum sativum agglutinin | PSA | o.n PFA | + | ++ | +++ | ++ | x | |||||
| 1 h PFA | ++ | ++ | ++ | |||||||||
| 4 h Lavdwosky | +++ f | ++ | ||||||||||
| Griffonia (Bandeiraea) simplicifolia lectin I | GSL I | o.n PFA | + | +++ | +++ | +++ | x | |||||
| 1 h PFA | + | +++ | +++ | +++ | ||||||||
| 4 h Lavdwosky | + | ++ | +++ | +++ | ||||||||
| Dolichos bilforus agglutinin | DBA | o.n PFA | ++ | +++ | +++ | x | ||||||
| 1 h PFA | ++ | +++ | +++ | ++ | ||||||||
| 4 h Lavdwosky | ++ | +++ | +++ | + | ||||||||
| Phaseolus vulgaris erythro agglutinin | PHA-E | o.n PFA | +++ a | ++ | x | |||||||
| 1 h PFA | +++ a | ++ | ||||||||||
| 4 h Lavdwosky | +++ b | +++ | ||||||||||
| Phaseolus vulgaris leuco agglutinin | PHA-L | o.n PFA | +++ a,b | ++ | x | |||||||
| 1 h PFA | +++ a | ++ | ||||||||||
| 4 h Lavdwosky | +++ b | +++ b | ||||||||||
| Griffonia (Bandeiraea) simplicifolia lectin II | GSL II | o.n PFA | ++ c | x | ||||||||
| 1 h PFA | ++ c | |||||||||||
| 4 h Lavdwosky | ++ d | ++ | ||||||||||
| Lens culinaris agglutinin | LCA | o.n PFA | +++ e | x | ||||||||
| 1 h PFA | +++ e | |||||||||||
| 4 h Lavdwosky | ++ | ++ | ++ f | |||||||||
| Vicia villosa agglutinin | VVL | o.n PFA | + | +++ g | ++ | ++ | +++ | x | ||||
| 1 h PFA | + | +++ g | ++ | ++ | +++ | |||||||
| 4 h Lavdwosky | ++ | +++ g | ++ | ++ | ||||||||
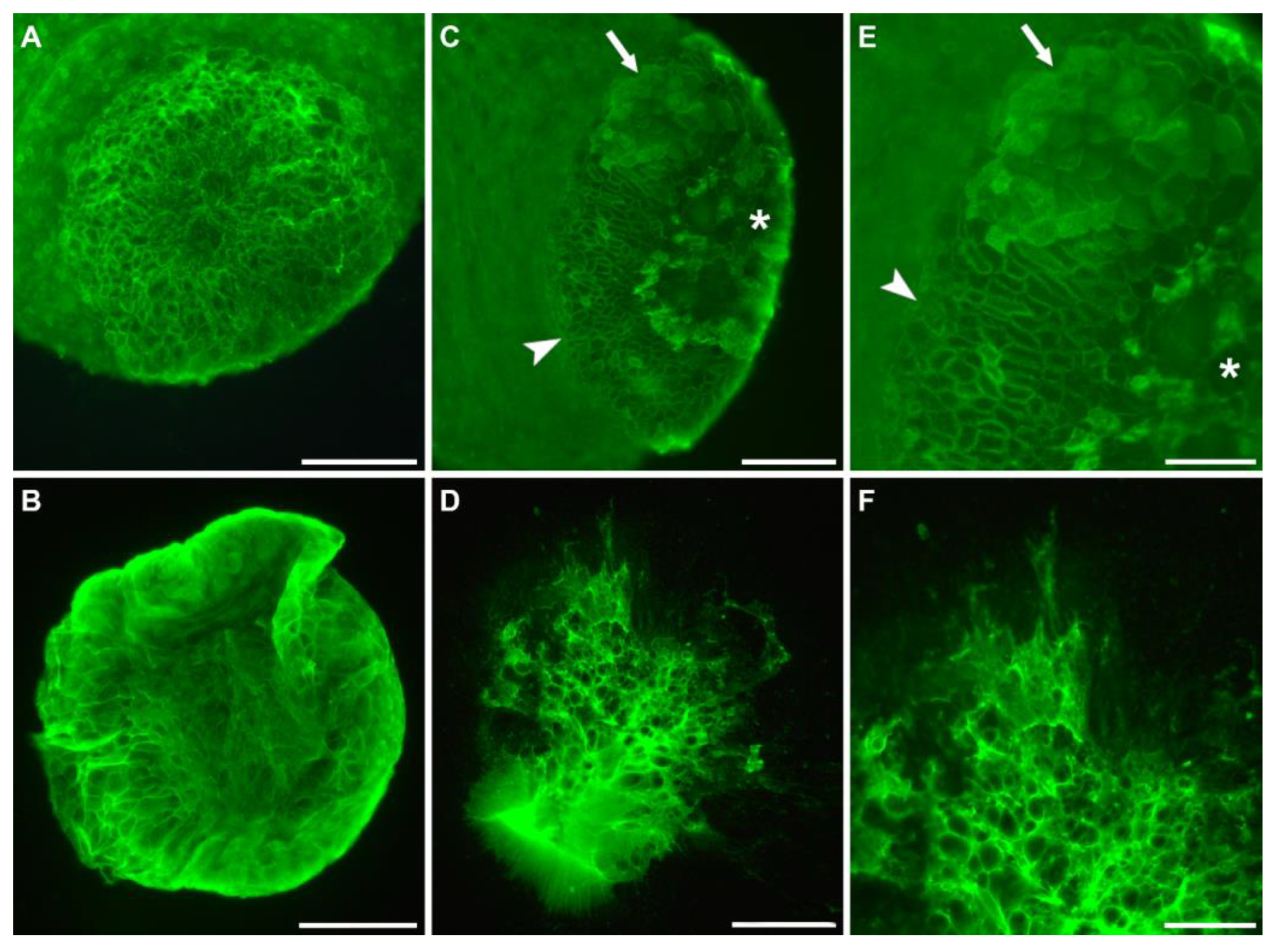
3.2. Lectins Detecting the Secreted Footprints
3.3. Lectin Labeling of Universal and Positional Distinct Hydra Cell Types and Associated Structures
4. Discussion
4.1. Glycan Distribution in Whole-Mount Hydra
4.2. Hydra Footprints Are Built up by a Fibrillar Material
4.3. Glycans Detected in the Secreted Footprint
4.4. Lectins as Markers for Hydra Nematocytes
4.5. Biomimetic Approaches and Their Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ditsche, P.; Summers, A.P. Aquatic versus terrestrial attachment: Water makes a difference. Beilstein J. Nanotechnol. 2014, 5, 2424–2439. [Google Scholar] [CrossRef]
- Davey, P.A.; Power, A.M.; Santos, R.; Bertemes, P.; Ladurner, P.; Palmowski, P.; Clarke, J.; Flammang, P.; Lengerer, B.; Hennebert, E.; et al. Omics-based molecular analyses of adhesion by aquatic invertebrates. Biol. Rev. 2021, 96, 1051–1075. [Google Scholar] [CrossRef]
- Hennebert, E.; Maldonado, B.; Ladurner, P.; Flammang, P.; Santos, R. Experimental strategies for the identification and characterization of adhesive proteins in animals: A review. Interface Focus 2015, 5, 20140064. [Google Scholar] [CrossRef] [Green Version]
- Lengerer, B.; Ladurner, P. Properties of temporary adhesion systems of marine and freshwater organisms. J. Exp. Biol. 2018, 221, jeb182717. [Google Scholar] [CrossRef] [Green Version]
- Simão, M.; Moço, M.; Marques, L.; Santos, R. Characterization of the glycans involved in sea urchin Paracentrotus lividus reversible adhesion. Mar. Biol. 2020, 167, 125. [Google Scholar] [CrossRef]
- Gaspar, L.; Flammang, P.; José, R.; Luis, R.; Ramalhosa, P.; Monteiro, J.; Nogueira, N.; Canning-Clode, J.; Santos, R. Interspecific Analysis of Sea Urchin Adhesive Composition Emphasizes Variability of Glycans Conjugated With Putative Adhesive Proteins. Front. Mar. Sci. 2021, 8, 1–16. [Google Scholar] [CrossRef]
- Hennebert, E.; Wattiez, R.; Flammang, P. Characterisation of the carbohydrate fraction of the temporary adhesive secreted by the tube feet of the sea star Asterias rubens. Mar. Biotechnol. 2011, 13, 484–495. [Google Scholar] [CrossRef]
- Lengerer, B.; Bonneel, M.; Lefevre, M.; Hennebert, E.; Leclère, P.; Gosselin, E.; Ladurner, P.; Flammang, P. The structural and chemical basis of temporary adhesion in the sea star Asterina gibbosa. Beilstein J. Nanotechnol. 2018, 9, 2071–2086. [Google Scholar] [CrossRef] [Green Version]
- Lengerer, B.; Hennebert, E.; Flammang, P.; Salvenmoser, W.; Ladurner, P. Adhesive organ regeneration in Macrostomum lignano. BMC Dev. Biol. 2016, 16, 20. [Google Scholar] [CrossRef] [Green Version]
- Bertemes, P.; Pjeta, R.; Wunderer, J.; Grosbusch, A.L.; Lengerer, B.; Gruner, K.; Knapp, M.; Mertens, B.; Andresen, N.; Hess, M.W.; et al. (Un)expected Similarity of the Temporary Adhesive Systems of Marine, Brackish, and Freshwater Flatworms. Int. J. Mol. Sci. 2021, 22, 12228. [Google Scholar] [CrossRef]
- Bertemes, P.; Grosbusch, A.L.; Geschwindt, A.; Kauffmann, B.; Salvenmoser, W.; Mertens, B.; Pjeta, R.; Egger, B.; Ladurner, P. Sticking Together an Updated Model for Temporary Adhesion. Mar. Drugs 2022, 20, 359. [Google Scholar] [CrossRef]
- Kang, V.; Lengerer, B.; Wattiez, R.; Flammang, P. Molecular insights into the powerful mucus-based adhesion of limpets (Patella vulgata L.). Open Biol. 2020, 10, 200019. [Google Scholar] [CrossRef]
- Wunderer, J.; Lengerer, B.; Pjeta, R.; Bertemes, P.; Kremser, L.; Lindner, H.; Ederth, T.; Hess, M.W.; Stock, D.; Salvenmoser, W.; et al. A mechanism for temporary bioadhesion. Proc. Natl. Acad. Sci. USA 2019, 116, 4297–4306. [Google Scholar] [CrossRef] [Green Version]
- Dreanno, C.; Matsumura, K.; Dohmae, N.; Takio, K.; Hirota, H.; Kirby, R.R.; Clare, A.S. An alpha2-macroglobulin-like protein is the cue to gregarious settlement of the barnacle Balanus amphitrite. Proc. Natl. Acad. Sci. USA 2006, 103, 14396–14401. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.; Leclère, P.; Flammang, P.; Hess, M.W.; Salvenmoser, W.; Hobmayer, B.; Ladurner, P. The cellular basis of bioadhesion of the freshwater polyp Hydra. BMC Zool. 2016, 1, 3. [Google Scholar] [CrossRef] [Green Version]
- Hobmayer, B.; Jenewein, M.; Eder, D.; Eder, M.K.; Glasauer, S.; Gufler, S.; Hartl, M.; Salvenmoser, W. Stemness in Hydra–A current perspective. Int. J. Dev. Biol. 2012, 56, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Bosch, T.C.; Anton-Erxleben, F.; Hemmrich, G.; Khalturin, K. The Hydra polyp: Nothing but an active stem cell community. Dev. Growth Differ. 2010, 52, 15–25. [Google Scholar] [CrossRef]
- Holstein, T.W. The role of cnidarian developmental biology in unraveling axis formation and Wnt signaling. Dev. Biol. 2022, 487, 74–98. [Google Scholar] [CrossRef]
- Vogg, M.C.; Galliot, B.; Tsiairis, C.D. Model systems for regeneration: Hydra. Development 2019, 146, dev177212. [Google Scholar] [CrossRef] [Green Version]
- Siebert, S.; Farrell, J.A.; Cazet, J.F.; Abeykoon, Y.; Primack, A.S.; Schnitzler, C.E.; Juliano, C.E. Stem cell differentiation trajectories in. Science 2019, 365, 1–8. [Google Scholar] [CrossRef]
- Rodrigues, M.; Ostermann, T.; Kremeser, L.; Lindner, H.; Beisel, C.; Berezikov, E.; Hobmayer, B.; Ladurner, P. Profiling of adhesive-related genes in the freshwater cnidarian Hydra magnipapillata by transcriptomics and proteomics. Biofouling 2016, 32, 1115–1129. [Google Scholar] [CrossRef] [Green Version]
- Philpott, D.E.; Chaet, A.B.; Burnett, A.L. A study of the secretory granules of the basal disk of hydra. J. Ultrastruct. Res. 1966, 14, 74–84. [Google Scholar] [CrossRef]
- David, C.N. Quantitative Method for Maceration of Hydra Tissue. Wilhelm Roux Arch. Fur Entwickl. Der. Org. 1973, 171, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Sahadevan, S.; Antonopoulos, A.; Haslam, S.M.; Dell, A.; Ramaswamy, S.; Babu, P. Unique, polyfucosylated glycan-receptor interactions are essential for regeneration of Hydra magnipapillata. ACS Chem. Biol. 2014, 9, 147–155. [Google Scholar] [CrossRef]
- Bottger, A.; Doxey, A.C.; Hess, M.W.; Pfaller, K.; Salvenmoser, W.; Deutzmann, R.; Geissner, A.; Pauly, B.; Altstatter, J.; Munder, S.; et al. Horizontal gene transfer contributed to the evolution of extracellular surface structures: The freshwater polyp Hydra is covered by a complex fibrous cuticle containing glycosaminoglycans and proteins of the PPOD and SWT (sweet tooth) families. PLoS ONE 2012, 7, e52278. [Google Scholar] [CrossRef] [Green Version]
- Holstein, T.W.; Hess, M.W.; Salvenmoser, W. Preparation techniques for transmission electron microscopy of Hydra. Methods Cell Biol. 2010, 96, 285–306. [Google Scholar] [CrossRef]
- Hennebert, E.; Viville, P.; Lazzaroni, R.; Flammang, P. Micro- and nanostructure of the adhesive material secreted by the tube feet of the sea star Asterias rubens. J. Struct. Biol. 2008, 164, 108–118. [Google Scholar] [CrossRef]
- Algrain, M.; Hennebert, E.; Bertemes, P.; Wattiez, R.; Flammang, P.; Lengerer, B. In the footsteps of sea stars: Deciphering the catalogue of proteins involved in underwater temporary adhesion. Open Biol. 2022, 12, 220103. [Google Scholar] [CrossRef]
- Shimizu, H.; Takaku, Y.; Zhang, X.M.; Fujisawa, T. The aboral pore of hydra: Evidence that the digestive tract of hydra is a tube not a sac. Dev. Genes Evol. 2007, 217, 563–568. [Google Scholar] [CrossRef]
- Pjeta, R.; Lindner, H.; Kremser, L.; Salvenmoser, W.; Sobral, D.; Ladurner, P.; Santos, R. Integrative Transcriptome and Proteome Analysis of the Tube Foot and Adhesive Secretions of the Sea Urchin. Int. J. Mol. Sci. 2020, 21, 946. [Google Scholar] [CrossRef]
- Hennebert, E.; Leroy, B.; Wattiez, R.; Ladurner, P. An integrated transcriptomic and proteomic analysis of sea star epidermal secretions identifies proteins involved in defense and adhesion. J. Proteom. 2015, 128, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Pjeta, R.; Wunderer, J.; Bertemes, P.; Hofer, T.; Salvenmoser, W.; Lengerer, B.; Coassin, S.; Erhart, G.; Beisel, C.; Sobral, D.; et al. Temporary adhesion of the proseriate flatworm Minona ileanae. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20190194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, F.; Wunderer, J.; Salvenmoser, W.; Hess, M.W.; Ladurner, P.; Rothbächer, U. Papillae revisited and the nature of the adhesive secreting collocytes. Dev. Biol. 2019, 448, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.L.; Davey, P.A.; Aldred, N. Sea anemones (Exaiptasia pallida) use a secreted adhesive and complex pedal disc morphology for surface attachment. BMC Zool. 2020, 5, 5. [Google Scholar] [CrossRef]
- Ozbek, S. The cnidarian nematocyst: A miniature extracellular matrix within a secretory vesicle. Protoplasma 2011, 248, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Beckmann, A.; Ozbek, S. The nematocyst: A molecular map of the cnidarian stinging organelle. Int. J. Dev. Biol. 2012, 56, 577–582. [Google Scholar] [CrossRef]
- Balasubramanian, P.G.; Beckmann, A.; Warnken, U.; Schnolzer, M.; Schuler, A.; Bornberg-Bauer, E.; Holstein, T.W.; Ozbek, S. Proteome of Hydra nematocyst. J. Biol. Chem. 2012, 287, 9672–9681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, S.; Morimoto, H.; Fujisawa, T.; Sugahara, K. Glycosaminoglycans in Hydra magnipapillata (Hydrozoa, Cnidaria): Demonstration of chondroitin in the developing nematocyst, the sting organelle, and structural characterization of glycosaminoglycans. Glycobiology 2007, 17, 886–894. [Google Scholar] [CrossRef] [Green Version]
- Adamczyk, P.; Zenkert, C.; Balasubramanian, P.G.; Yamada, S.; Murakoshi, S.; Sugahara, K.; Hwang, J.S.; Gojobori, T.; Holstein, T.W.; Ozbek, S. A non-sulfated chondroitin stabilizes membrane tubulation in cnidarian organelles. J. Biol. Chem. 2010, 285, 25613–25623. [Google Scholar] [CrossRef] [Green Version]
- Almeida, M.; Reis, R.L.; Silva, T.H. Marine invertebrates are a source of bioadhesives with biomimetic interest. Mater. Sci. Eng. C 2019, 108, 110467. [Google Scholar] [CrossRef]
- Ahn, B.K. Perspectives on Mussel-Inspired Wet Adhesion. J. Am. Chem. Soc. 2017, 139, 10166–10171. [Google Scholar] [CrossRef] [Green Version]
- Hennebert, E.; Wattiez, R.; Demeuldre, M.; Ladurner, P.; Hwang, D.S.; Waite, J.H.; Flammang, P. Sea star tenacity mediated by a protein that fragments, then aggregates. Proc. Natl. Acad. Sci. USA 2014, 111, 6317–6322. [Google Scholar] [CrossRef] [Green Version]
- Lefevre, M.; Flammang, P.; Aranko, A.S.; Linder, M.B.; Scheibel, T.; Humenik, M.; Leclercq, M.; Surin, M.; Tafforeau, L.; Wattiez, R.; et al. Sea star-inspired recombinant adhesive proteins self-assemble and adsorb on surfaces in aqueous environments to form cytocompatible coatings. Acta Biomater. 2020, 112, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Lefevre, M.; Ederth, T.; Masai, T.; Wattiez, R.; Leclere, P.; Flammang, P.; Hennebert, E. Disentangling the Roles of Functional Domains in the Aggregation and Adsorption of the Multimodular Sea Star Adhesive Protein Sfp1. Mar. Biotechnol. 2021, 23, 724–735. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seabra, S.; Zenleser, T.; Grosbusch, A.L.; Hobmayer, B.; Lengerer, B. The Involvement of Cell-Type-Specific Glycans in Hydra Temporary Adhesion Revealed by a Lectin Screen. Biomimetics 2022, 7, 166. https://0-doi-org.brum.beds.ac.uk/10.3390/biomimetics7040166
Seabra S, Zenleser T, Grosbusch AL, Hobmayer B, Lengerer B. The Involvement of Cell-Type-Specific Glycans in Hydra Temporary Adhesion Revealed by a Lectin Screen. Biomimetics. 2022; 7(4):166. https://0-doi-org.brum.beds.ac.uk/10.3390/biomimetics7040166
Chicago/Turabian StyleSeabra, Sofia, Theresa Zenleser, Alexandra L. Grosbusch, Bert Hobmayer, and Birgit Lengerer. 2022. "The Involvement of Cell-Type-Specific Glycans in Hydra Temporary Adhesion Revealed by a Lectin Screen" Biomimetics 7, no. 4: 166. https://0-doi-org.brum.beds.ac.uk/10.3390/biomimetics7040166