

Utilizing Dichloromethane as an Extremely Proficient Substitute for Phenol/Chloroform in Extracting RNA with Exceptional Purity from Woody Tissues of Coconut

Abstract
:1. Introduction
2. Experimental Design
2.1. Materials
 Cautions!
Cautions!2.2. Equipment
- Vortex (Vortex-BE1, Kylin-Bell Lab Instruments, Nantong, China);
- Bench centrifuge (Eppendorf Centrifuge 5804 R);
- PCR machine/thermocycler (TaKaRa thermocycler dice touch TP350, TaKaRa Bio Incharge, Le Pecq, France);
- Nanodrop (Nanodrop 2000 spectrophotometer, Thermo Scientific, Waltham, MA, USA);
- Agrose gel electrophoresis system;
- Gel Doc system (Syngene G: Box F3, Gene Company Limited, Hong Kong, China).
2.3. Plant Materials
2.4. The RNA Extraction Buffer
- Guanidine thiocyanate : 23 g
- 3M sodium acetate (pH 5) : 3 mL
- Glycerol : 4 mL
- ddH2O or DEPC water : up to 100 mL
- PVP-40 : 1.6%
- β-marceptoethanol : 1.9%
3. Procedure
3.1. Tissue Preparation and Maceration
- 1.
- Before homogenizing the tissue, the sample-filled zipper bags from the refrigerator were moved to liquid nitrogen to prevent defrosting.
Critical step - Defrosting could lead to RNA degradation by RNase enzymes.
- 2.
- The sterile mortar and pestle, previously autoclaved, were cooled down by the addition of liquid nitrogen.
- 3.
- Approximately 0.1 g of refrigerated sample was taken from the zipper bag and moved into the liquid nitrogen and PVP-40-holding mortar. The sample was then finely powdered using the pestle, with the mortar being periodically refilled with liquid nitrogen in the course of milling process to avoid defrosting.
Critical step - (a)
- To elude sample loss, it is essential to add liquid nitrogen slowly to the mortar, taking care to prevent splashing during the process.
- (b)
- Preventing sample defrosting is crucial since it can cause the liberation of RNA from disrupted cells, rendering it susceptible to degradation by RNase enzymes.
- (c)
- To ensure the prevention of sample cross-contamination with RNase, it is vital to carry out this step in a fume hood.
3.2. Extraction of RNA
- 4.
- Using a frozen spatula, the ground sample was rapidly shifted into an eppendorf tube holding 700 μL of the extraction buffer. After vortexing the contents of the tube for 15 s, we introduced 500 μL of dichloromethane, vortexed it again for 15 s, and, finally, centrifuged the tube for 5 min at 16,639 rcf and 4 °C.
Critical step 3.3. Precipitation of RNA
- 5.
- We gently moved 500 µL of top aqueous fraction of the solution to a new tube, ensuring that the tube was Ribonuclease-free. Subsequently, we introduced the same amount of chilled isopropanol into the tube, inverted the tubes to mix the solution, and set it aside at ambient temperature for 5 min.
Critical step - 6.
- After the incubation, we centrifuged the contents of the tube at 16,639 rcf and 4 °C for 5 min and subsequently disposed of the supernatant. The nucleic acids aggregated at the base of the tube.
Critical step - 7.
- The pellet was rinsed with 1 mL of 75% ethanol, accompanied by centrifugation at 4 °C and 11,000 rcf for 5 min. The washing step was repeated once more in the same manner.
- 8.
- The mixture was supplemented with 50 μL of DPEC and 1 μL of DNAse (to collect RNA) and then incubated at 37 °C for 30 min.
- 9.
- We added the same amount of chilled isopropanol into the tube, inverted the tubes to mix the solution, and set it aside at ambient temperature for 5 min. After incubation we centrifuged the contents of the tube at 16,639 rcf and 4 °C for 5 min and subsequently disposed of the supernatant. The RNA aggregated at the base of the tube.
- 10.
- The pellet was rinsed with 1 mL of 75% ethanol, accompanied by centrifugation at 4 °C and 11,000 rcf for 5 min. The washing step was repeated once more in the same manner.
- 11.
- After drying the pellet for 5 min, we carefully re-dissolved the RNA in 20 μL of Ribonuclease-free (DEPC) water using a pipette until it was fully dissolved.
3.4. RNA Characterization and Quantification
- 10.
- The RNA abundance and integrity were evaluated using the NanoDrop, whereas the purity was validated through agarose gel electrophoresis.
- (a)
- The amount of RNA was assayed spectrophotometrically using NanoDrop 2000 analysis. A sample (1 μL) was loaded onto the NanoDrop 2000, and the optical density was measured at 260 and 280 nm. The quantity of RNA was calculated using the absorbance at A260, whereas the purity of the RNA was assessed based on the A260/280 ratio.
- (b)
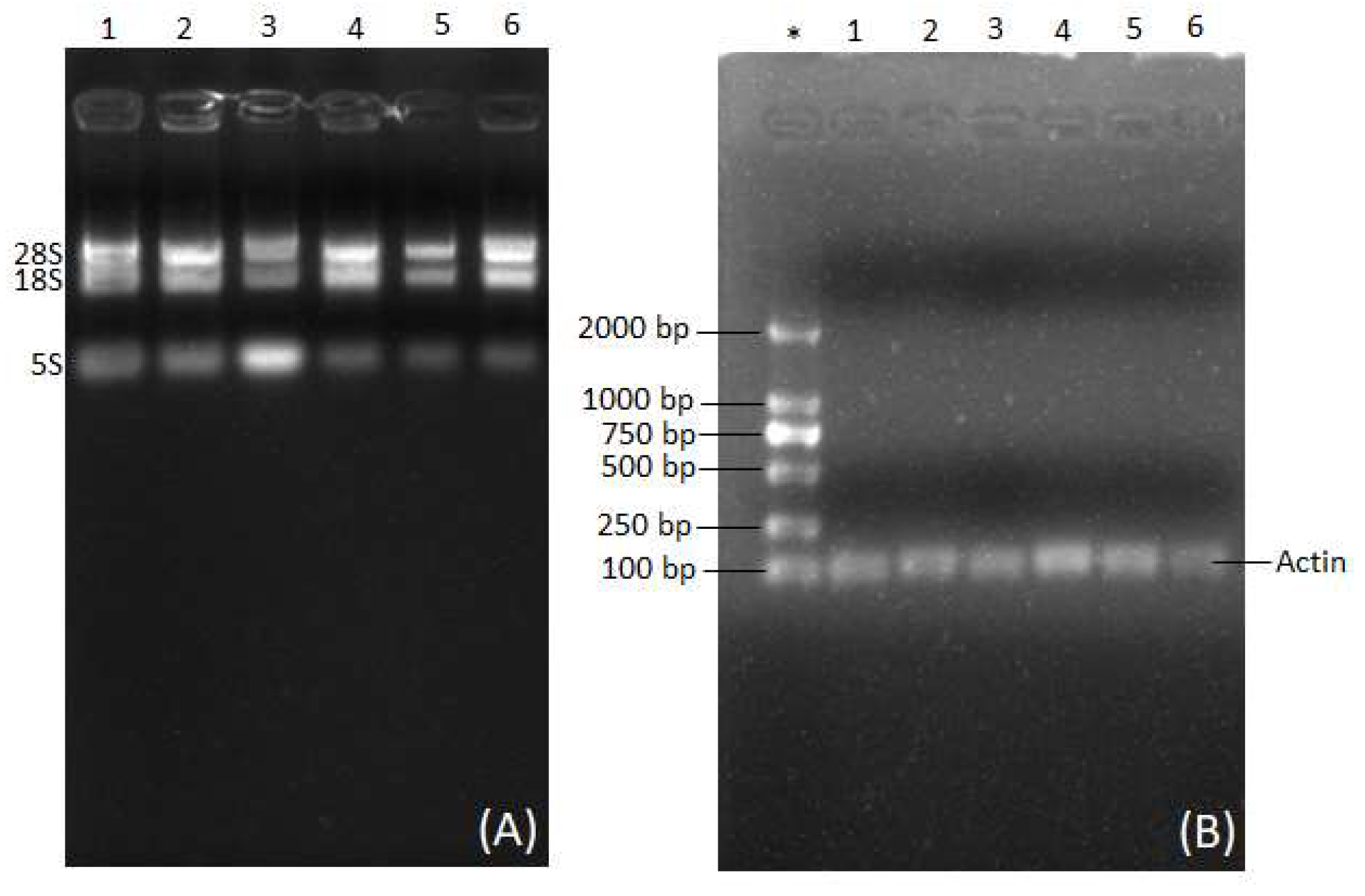
- To set up the agarose gel for electrophoresis, agarose (0.2 g) was dissolved in 25 mL of 0.5x TBE buffer in a Ribonuclease-free conical flask, resulting in a 0.8% (w/v) agarose gel. The conical flask was subjected to microwave heating until the agarose was fully dispersed in TBE buffer. Following that, ethidium bromide (2 μL) was introduced to the contents and stirred to achieve thorough mixing. The mixture was deposited inside the gel tray alongside a comb and allowed to solidify for about 20 min. Once the gel had solidified, RNA-loading buffer (2 μL) was combined with the RNA sample (3 μL), and the resulting mixture was introduced to the gel. The gel was subsequently subjected to electrophoresis at 120 V and 240 mA for 12 min. Once the electrophoresis process was complete, the gel bands were spotted, we recorded the Gel Doc system to capture images, and saved the files. Microsoft Office Picture Manager was utilized for cropping the gel images.
Critical step 3.5. cDNA Generation
3.6. RT-PCR
4. Results
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodrigues, S.M.; Soares, V.L.; De Oliveira, T.M.; Gesteira, A.S.; Otoni, W.C.; Costa, M.G. Isolation and purification of RNA from tissues rich in polyphenols, polysaccharides, and pigments of annatto (Bixa orellana L.). Mol. Biotechnol. 2007, 37, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Stegemann, J.P. Extraction of high quality RNA from polysaccharide matrices using cetlytrimethylammonium bromide. Biomaterials 2010, 31, 1612–1618. [Google Scholar] [CrossRef]
- Rubio-Piña, J.A.; Zapata-Pérez, O. Isolation of total RNA from tissues rich in polyphenols and polysaccharides of mangrove plants. Electron. J. Biotechnol. 2011, 14, 11. [Google Scholar]
- Djami-Tchatchou, A.; Straker, C. The isolation of high quality RNA from the fruit of avocado (Persea americana Mill.). S. Afr. J. Bot. 2012, 78, 44–46. [Google Scholar] [CrossRef]
- Wang, X.; Xiao, H.; Chen, G.; Zhao, X.; Huang, C.; Chen, C.; Wang, F. Isolation of high-quality RNA from Reaumuria soongorica, a desert plant rich in secondary metabolites. Mol. Biotechnol. 2011, 48, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Bilgin, D.D.; DeLucia, E.H.; Clough, S.J. A robust plant RNA isolation method suitable for Affymetrix GeneChip analysis and quantitative real-time RT-PCR. Nat. Protoc. 2009, 4, 333–340. [Google Scholar] [CrossRef]
- Xiao, Y.; Yang, Y.; Cao, H.; Fan, H.; Ma, Z.; Lei, X.; Mason, A.S.; Xia, Z.; Huang, X. Efficient isolation of high quality RNA from tropical palms for RNA-seq analysis. Plant Omics 2012, 5, 584. [Google Scholar]
- Ouyang, K.; Li, J.; Huang, H.; Que, Q.; Li, P.; Chen, X. A simple method for RNA isolation from various tissues of the tree Neolamarckia cadamba. Biotechnol. Biotech. Equip. 2014, 28, 1008–1013. [Google Scholar] [CrossRef]
- Dash, P.K. High quality RNA isolation from ployphenol-, polysaccharide-and protein-rich tissues of lentil (Lens culinaris). 3 Biotechnol. 2013, 3, 109–114. [Google Scholar] [CrossRef]
- Iqbal, A.; Yang, Y.; Qadri, R.; Wu, Y.; Li, J.; Shah, F.; Hamayun, M.; Hussain, A. QRREM method for the isolation of high-quality RNA from the complex matrices of coconut. Biosci. Rep. 2019, 39, BSR20181163. [Google Scholar] [CrossRef]
- Iqbal, A.; Yang, Y.; Shah, F.; Shah, T.; Hamayun, M.; Hussain, A.; Khan, Z.H. Efficient, cheap and reliable method for the extraction of gDNA from the hard and complex tissues of coconut. Fresenius Environ. Bull. 2019, 28, 7951–7954. [Google Scholar]
- Qadri, R.; Iqbal, A.; Wu, Y.; Li, J.; Nisar, N.; Azam, M.; Yang, Y. A modified protocol for total RNA isolation from different oil palm (Elaeis guineensis) tissues using cetyltrimethylammonium bromide. Curr. Sci. 2019, 116, 479–482. [Google Scholar] [CrossRef]
- Iqbal, A.; Yang, Y.; Wu, Y.; Li, J.; Hamayun, M.; Hussain, A.; Shah, F. An easy and robust method for the isolation of high quality RNA from coconut tissues. Electron. J. Biotechnol. 2020, 48, 78–85. [Google Scholar] [CrossRef]
- Pant, B.D.; Musialak-Lange, M.; Nuc, P.; May, P.; Buhtz, A.; Kehr, J.; Walther, D.; Scheible, W.-R. Identification of nutrient-responsive Arabidopsis and rapeseed microRNAs by comprehensive real-time polymerase chain reaction profiling and small RNA sequencing. Plant Physiol. 2009, 150, 1541–1555. [Google Scholar] [CrossRef]
- Box, M.S.; Coustham, V.; Dean, C.; Mylne, J.S. Protocol: A simple phenol-based method for 96-well extraction of high quality RNA from Arabidopsis. Plant Methods 2011, 7, 7. [Google Scholar] [CrossRef]
- Shi, Z.; Wang, J.; Wan, X.; Shen, G.; Wang, X.; Zhang, J. Over-expression of rice OsAGO7 gene induces upward curling of the leaf blade that enhanced erect-leaf habit. Planta 2007, 226, 99–108. [Google Scholar] [CrossRef]
- Zheng, Y.; Chen, M.; Yang, C.; Ming, X.; Liu, X.; Lan, X.; Liao, Z. Cloning and characterization of MECS and HDS genes from Rauvolfia verticillata. Plant Omics 2011, 4, 82. [Google Scholar]
- Westermann, A.J.; Gorski, S.A.; Vogel, J. Dual RNA-seq of pathogen and host. Nat. Rev. Biotechnol. 2012, 10, 618–630. [Google Scholar] [CrossRef]
- Oshlack, A.; Robinson, M.D.; Young, M.D. From RNA-seq reads to differential expression results. Genome Biol. 2010, 11, 220. [Google Scholar] [CrossRef]
- Hirsch, P.R.; Mauchline, T.H.; Clark, I.M. Culture-independent molecular techniques for soil microbial ecology. Soil Biol. Biochem. 2010, 42, 878–887. [Google Scholar] [CrossRef]
- Marchese, D.; de Groot, N.S.; Lorenzo Gotor, N.; Livi, C.M.; Tartaglia, G.G. Advances in the characterization of RNA-binding proteins. Wiley Interdiscip. Rev. RNA 2016, 7, 793–810. [Google Scholar] [CrossRef]
- Bechhofer, D.H.; Deutscher, M.P. Bacterial ribonucleases and their roles in RNA metabolism. Crit. Rev. Biochem. Mol. Biol. 2019, 54, 242–300. [Google Scholar] [CrossRef]
- Aburayyan, W.S. High-throughput RNA extraction method for P. aeruginosa and S. pyogenes biofilms. J. Appl. Pharm. Sci. 2023, 13, 097–105. [Google Scholar] [CrossRef]
- Greco, M.; Saez, C.A.; Brown, M.T.; Bitonti, M.B. A simple and effective method for high quality co-extraction of genomic DNA and total RNA from low biomass Ectocarpus siliculosus, the model brown alga. PLoS ONE 2014, 9, e96470. [Google Scholar] [CrossRef] [PubMed]
- Heerklotz, H. Interactions of surfactants with lipid membranes. Q. Rev. Biophys. 2008, 41, 205–264. [Google Scholar] [CrossRef] [PubMed]
- Schultz, D.J.; Craig, R.; Cox-Foster, D.L.; Mumma, R.O.; Medford, J.I. RNA isolation from recalcitrant plant tissue. Plant Mol. Biol. Rep. 1994, 12, 310–316. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Z.; Zhu, X.; Hu, X.; Zhang, H.; Guo, Q.; Yada, R.Y.; Cui, S.W. Seed coat mucilages: Structural, functional/bioactive properties, and genetic information. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2534–2559. [Google Scholar] [CrossRef]
- Wang, J.; Ali, Z.; Wang, N.; Liang, W.; Liu, H.; Li, F.; Yang, H.; He, L.; Nie, L.; He, N. Simultaneous extraction of DNA and RNA from Escherichia coli BL 21 based on silica-coated magnetic nanoparticles. Sci. China Chem. 2015, 58, 1774–1778. [Google Scholar] [CrossRef]
- Ky, H.; Yeap, S.; Napis, S. The best method for isolated total RNA from durian tissues. Int. Food Res. J. 2012, 19, 1181. [Google Scholar]
- Japelaghi, R.H.; Haddad, R.; Garoosi, G.-A. Rapid and efficient isolation of high quality nucleic acids from plant tissues rich in polyphenols and polysaccharides. Mol. Biotechnol. 2011, 49, 129–137. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Method | Sample Tissue | Conc. (µg/g FW) | A260/280 Ratio |
|---|---|---|---|
| Dichloromethane | Mature coconut tree roots | 120.7 ± 3.18 | 1.95 ± 0.012 |
| Mature coconut mesocarp | 134.6 ± 0.61 | 1.98 ± 0.028 | |
| RNA plant | Mature coconut tree roots | 5.1 ± 1.86 | 0.76 ± 0.165 |
| Mature coconut mesocarp | 47.8 ± 15.78 | 1.02 ± 0.002 | |
| TRIZOL | Mature coconut tree roots | 140.3 ± 6.79 | 1.42 ± 0.015 |
| Mature coconut mesocarp | 346.7 ± 43.18 | 1.26 ± 0.166 | |
| CTAB | Mature coconut tree roots | 18.9 ± 1.15 | 0.83 ± 0.052 |
| Mature coconut mesocarp | 11.4 ± 1.33 | 0.92 ± 0.094 |
| Sample | Conc. (µg/g FW) | A260/280 Ratio |
|---|---|---|
| Coconut endosperm | 120.4 ± 2.30 | 2.01 ± 0.060 |
| Coconut leaf | 172.9 ± 0.95 | 1.97 ± 0.017 |
| Coconut apple | 298.2 ± 2.83 | 1.91 ± 0.050 |
| Coconut leaf bud | 175.7 ± 1.64 | 2.05 ± 0.033 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iqbal, A.; Yang, Y. Utilizing Dichloromethane as an Extremely Proficient Substitute for Phenol/Chloroform in Extracting RNA with Exceptional Purity from Woody Tissues of Coconut. Methods Protoc. 2023, 6, 75. https://0-doi-org.brum.beds.ac.uk/10.3390/mps6050075
Iqbal A, Yang Y. Utilizing Dichloromethane as an Extremely Proficient Substitute for Phenol/Chloroform in Extracting RNA with Exceptional Purity from Woody Tissues of Coconut. Methods and Protocols. 2023; 6(5):75. https://0-doi-org.brum.beds.ac.uk/10.3390/mps6050075
Chicago/Turabian StyleIqbal, Amjad, and Yaodong Yang. 2023. "Utilizing Dichloromethane as an Extremely Proficient Substitute for Phenol/Chloroform in Extracting RNA with Exceptional Purity from Woody Tissues of Coconut" Methods and Protocols 6, no. 5: 75. https://0-doi-org.brum.beds.ac.uk/10.3390/mps6050075