
Tissue-Specific and Differential Cold Responses in the Domesticated Cold Tolerant Fugu


1
International Research Center for Marine Biosciences, Ministry of Science and Technology, Shanghai Ocean University, Shanghai 201306, China
2
Key Laboratory of Exploration and Utilization of Aquatic Genetic Resources, Ministry of Education, Shanghai Ocean University, Shanghai 201306, China
3
College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 201306, China
*
Author to whom correspondence should be addressed.
Fishes 2022, 7(4), 159; https://0-doi-org.brum.beds.ac.uk/10.3390/fishes7040159
Submission received: 2 June 2022
/
Revised: 26 June 2022
/
Accepted: 28 June 2022
/
Published: 30 June 2022
(This article belongs to the Special Issue Discovery and Utilization of Fish Resistance Genes)
Abstract
:Domestication can be defined as the artificial selection in animals to achieve morphological, physiological, and developmental conformity to human needs, with the aim of improving various limitations in species under a human feeding environment. The future sustainability of aquaculture may rely partly on the availability of numerous domesticated fish species. However, the underlying adaptive mechanisms that result in the domestication of fish are still unclear. Because they are poikilothermic, temperature is a key environmental element that affects the entire life of fish, so studying the association between physiological and behavioral changes in low-temperature domesticated fish can provide a model for understanding the response mechanisms of fish under cold stress. Through 5 generations and 10 years of artificial selection at low temperatures, we used cold-tolerant fugu as a biological model to compare transcriptome changes in brain and liver tissues to study the effects of cold stress on fish. It was found that the expression of genes such as apoptosis, p53, oxidative phosphorylation, and mitochondrial β-oxidation in the brain of cold-tolerant fugu was significantly lower than the wild type due to cold stress, while excessive energy metabolism would lead to the production of reactive oxygen species (ROS) and exacerbate the brain damage, thus causing rollover and coma. Meanwhile, under cold stress, the signaling pathways involved in glycogenolysis and lipid metabolism, such as insulin signaling, adipocytokines, and mTOR signaling pathways, were significantly up-regulated in the liver of cold-tolerant fugu. Although the mitochondrial β-oxidation pathway was increased in cold-tolerant fugu liver tissues, the transcriptome was not enriched in apoptotic. These phenomena predict that in response to low-temperature conditions, cold-tolerant fugu employs a dynamic inter-organ metabolic regulation strategy to cope with cold stress and reduce damage to brain tissues.
1. Introduction
Domestication lies at the heart of human civilization and involves raising animals in an environment that differs substantially from their natural habitat [1]. Aquaculture is considerably inexperienced compared to agriculture, which relies on natural sources to farm numerous species. Similar to mammalian species, the indispensable prerequisite to domestication is a consistent control of reproduction year after year in successive generations of fish maintained and bred in captivity [2]. Domestication gives rise to organisms that perform better under culture conditions than their wild conspecifics and may express additional attributes desired by culturists [3]. Domestication at individual or population levels starts with the transfer of animals from the wild (F0 fish) to a captive place where they are faced with a new living environment [4,5]. Every history of domestication is unique, and the populations during domestication have undergone a range of biological phenomena linked to human practices. These mechanisms contribute to genetic, genomic, and phenotypic changes in animals [4]. However, despite the importance of domestication for the aquaculture industry, there is very little knowledge about the processes being modified along with the adaptation of fish to the culture conditions.
The impact of temperature changes on biological systems is an important topic related to global climate changes and seasonal temperature differences [6,7]. Environmental stressors disrupt homeostasis and are harmful to the physiological function of organisms in fish. With the exacerbation of extreme weather, temperature stress seriously affects the survival of organisms and causes mass mortalities in aquaculture. As poikilothermic vertebrates, aquatic animals can change their body temperature according to the external environment. Thus, cold stress has the greatest impact on reproductive, metabolic, and developmental functions in the teleost fish.
Water temperature is an important environmental factor that affects the physiological and biochemical activities of fish [8]. Information about the genes and biological pathways that affect the acclimation and function of fish with the change in environmental and body temperatures is scarce. Several studies focusing on different fish species used transcriptomic approaches to gain a broad view of genes involved in the response and acclimation to low temperatures. These studies pointed out that tissues involved in the physiological response and acclimation to low-temperature stress could exhibit tissue-specific characteristics sometimes. In addition, certain pathways, such as mitochondrial function, lipid and carbohydrate metabolism, antioxidant response, apoptosis, RNA processing, and protein catabolism, were found to exhibit temperature-dependent regulation [9,10,11]. Investigating the molecular mechanisms between various organs of fish affected by low temperatures can provide a better understanding of how organisms adapt or respond to environmental challenges. To date, few studies have focused on changes in transcriptome and gene expression regulation in cold-tolerant fish.
Takifugu obscurus is a migratory river and sea animal whose optimum growth temperature is between 20~25 °C. When the temperature drops to 18 °C, wild-type fugu will reduce food intake and stop growing, and farmed Takifugu obscurus will suffer from frostbite, with symptoms such as skin congestion and loss of equilibrium. When the temperature drops to 12 °C for more than 12 h, Takifugu obscurus will become comatose and eventually die [9,12]. Generally speaking, farmed Takifugu obscurus need to be warmed with a greenhouse boiler in winter, and it is impossible to avoid death by frostbite caused by the untimely transfer to pools, which may increase farming costs and mortality rates. Therefore, the Zhongyang Group has artificially selected wild Takifugu obscurus under low-temperature stress. The wild population collected from the Yangtze River is named after ’Zhongyang No.1′, which has undergone population selection for 5 consecutive generations from 1997 to 2017, with cold tolerance being the main target trait. This new species provides an excellent model for investigating the extent of qualitative changes in gene expression in response to cold acclimation in fish.
2. Materials and Methods
2.1. Animals and Experimental Conditions
Both wild-type and cold-tolerant fugu (with an average body weight of 343 ± 6.2 g and about 18 months of age) were collected from Zhongyang Group, a fish farm in Nantong (Jiangsu, China), and acclimated for 2 weeks in 500 L cycling-filtered plastic tanks containing continuously circulating aerated water at 24 ± 1 °C (pH 7.2, 7.5 mg/L dissolved oxygen). During the acclimation period, the fish were fed a commercial fish diet twice a day until 24 h before the experimental cooling treatments. Then, the wild-type and domesticated cold-tolerant fugu were kept as 2 groups of 40 samples each in different tanks. The two groups of fugu were exposed to the same continuous, linear cooling scheme, with the water temperature declining from 24 °C to 16 °C and then to 7 °C, finally reaching 4 °C at a rate of 0.5 °C/h, and each step maintained for 12 h. Brain and liver tissues were collected from the wild-type and cold-tolerant fugu at 24 °C, 16 °C, 7 °C, and 4 °C, and the tissues were stored separately at −80 °C until use. The experiments were conducted according to the principles expressed in the Guide for the Care and Use of Laboratory Animals published by the National Research Council of the National Academies.
2.2. Tissue Collection and mRNA-Seq Library Construction for Illumina Sequencing
After cold treatment for 12 h at 4 °C, 12 cold-tolerant and 4 wild-type fish survived (could not swim normally but had the ability to move), and for each temperature range, we chose 3 samples. Two tissues (brain and liver) were sampled, and the tissues were stored separately at −80 °C until use. mRNA was extracted from the tissue samples using TRIzol reagent from Invitrogen, then the content of RNA was measured using NanoDrop 2000, and the quality of RNA samples was assessed by agarose gel electrophoresis. The integrity of RNA samples was confirmed using Agilent 2100 Bioanalyzer, and 5 μg of total RNA was used to isolate mRNA. Purified mRNA samples were fragmented into small pieces, and double-stranded cDNA was synthesized using random hexamer primers. The synthesized cDNA was subjected to end-repair, phosphorylation, 3′ adenylation, adapter ligation, and PCR amplification. Finally, sequencing libraries of 150 to 350 bp were constructed. Three independent biological replicates for cold stress in cold-tolerant and wild-type samples at each temperature were used for library construction, and each RNA sample was obtained from two different tissues. The library was sequenced with the Illumina HiSeq xten platform.
2.3. Quantitative Real-Time PCR
To validate the results of the RNA-Seq analysis, six DEGs were randomly selected for quantitative real-time PCR (qRT-PCR). The β-actin (ACTB) and Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as the internal control gene, as previously described. Total RNA was extracted from the brain and liver, then the samples were tested under the same conditions as the samples of transcriptome analysis and were dissected at the same time. A 1 μg/μL concentration of total RNA was used for cDNA synthesis, in accordance with the steps provided by the reverse transcription kit (PrimeScript RT Master Mix, Takara), and the cDNA was used as the template for the qRT-PCR experiments. TB Green Premix Ex Taq II (Takara) was used to perform a quantitative analysis of genes in the cfx96 fluorescence quantitative PCR instrument (BioRad, Hercules, CA, USA). The relative expression of each sample was calculated by the 2−ΔΔCT. SPSS (Version 23.0, IBM, Armonk, NY, USA) was used for Pearson’s correlation analysis. Detailed information on the primers is given in Table 1.
2.4. Bioinformatic Analyses
The raw paired-end reads were filtered, and the trimming quality was controlled by SeqPrep (https://github.com/jstjohn/SeqPrep (accessed on 25 October 2019)). Then, clean reads were separately aligned to the reference genome (ftp://ftp.ncbi.nlm.nih.gov/genomes/Takifugu rubripes/) with orientation mode using HISAT2 (http://0-ccb-jhu-edu.brum.beds.ac.uk/software/hisat2/index.shtml (accessed on 2 November 2020)) software. The Cufflinks (http://cole-trapnell-lab.github.io/cufflinks/ (accessed on 18 February 2021)) software was used for transcript quantification and differential expression analysis between the cold-tolerant and wild-type at the same temperatures. To identify DEGs (differential expression genes) between two different samples, the expression level of each transcript was calculated according to the transcripts per million reads (TPM) method. Transcripts with at least a 2-fold change in response to temperature (DEGs with |log2FC| > 1 and Q value <= 0.05) were regarded as significantly up-or down-regulated using RSEM (http://deweylab.biostat.wisc.edu/rsem/ (accessed on 14 October 2021)) software. In addition, the functional–enrichment annotation of transcripts with significant temperature-dependent expression was extended using gene ontology (GO) and KEGG biological pathway enrichment (regarded as significantly up- or down-regulated at an adjusted p-value of 0.05) categories assigned to the zebrafish orthologs. GO functional enrichment and KEGG pathway analysis were carried out with KOBAS (http://kobas.cbi.pku.edu.cn/home.do (accessed on 10 February 2022)).
Gene Set Enrichment Analysis (GSEA) has been extensively applied to identify underlying pathways [13]. According to the different expression of cold-tolerant and wild-type genes under cold stress, the C2. symbols.gmt dataset was obtained from the Molecular Signatures Database (NOM p-values < 0.05, |NES| > 1 and FDR q < 0.05 were considered statistically significant). To overcome the limitations of hard clustering, we applied soft clustering using the Mfuzz [14] package to obtain the gene expression patterns when fish were under different low temperatures.
3. Results
3.1. Distinct Cold Responses in Wild-Type and Cold-Tolerant Fugu under Cooling Scheme
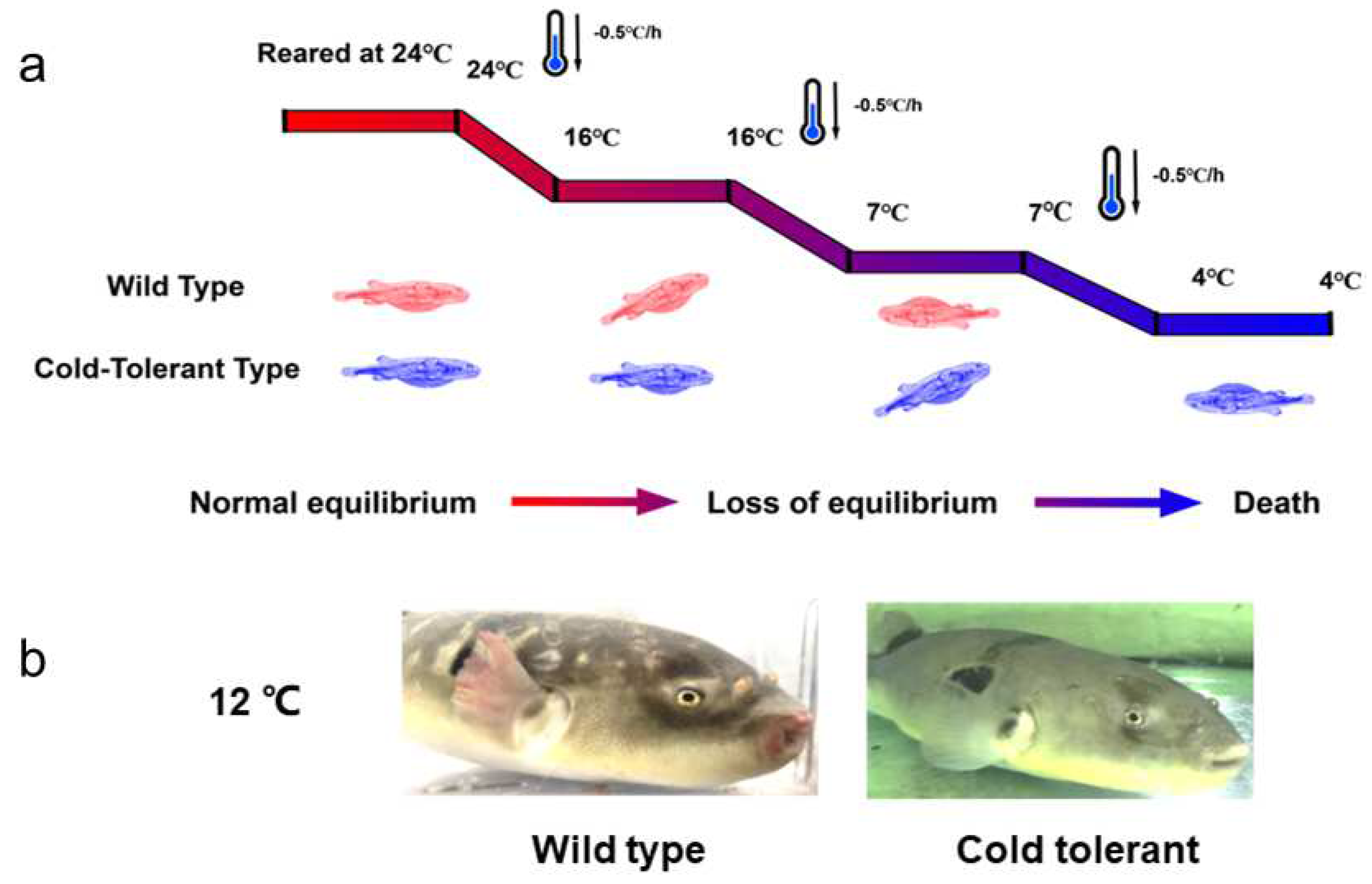
To investigate their resistance to cold stress, fugu were pre-acclimated to the optimal temperature for 7 days. Subsequently, the temperature was gradually lowered from 24 °C, and the behavior of the 2 types of fugu under low-temperature response was recorded. For wild-type fugu, the behavioral responses changed with decreasing temperatures. The fish showed normal swimming behavior at 18 °C for 12 h, began to swim uncoordinatedly (e.g., head down and bumping into tank walls and subcutaneous congestion) at 16–12 °C, had difficulty maintaining equilibrium at 9 °C, and completely lost equilibrium and fell into a coma at 7 °C. As for cold-tolerant fugu, abnormal behavior barely appeared until the water temperature dropped to 7 °C for 12 h and eventually collapsed at 4 °C (Figure 1). Apparently, the lethal temperature of 50% (LT50) for wild-type fugu was 13.2 °C, at which only a few cold-tolerant individuals lost equilibrium. Compared to wild-type, LT50 in cold-tolerant fugu was as low as 6.2 °C. Surprisingly, the lowest survival temperature was nearly 3.9 °C (Figure S1).
After being treated at 12 °C for 12 h, the pectoral fins and lips of wild-type fugu that were in direct contact with water exhibited obvious congestion, while the physiological behavior of cold-tolerant fugu remained unchanged, such as normal swimming, feeding, and response to external stimuli (Figure 1). Compared with wild-type, cold-tolerant fugu had a stronger ability to cope with cold stress.
3.2. Cold Stress Induces Different Expression Patterns in Liver and Brain Tissues between Wild-Type and Cold-Tolerant Fugu
Identifying differences in a specific model of cold stress transcription in liver and brain tissues between wild-type and cold-tolerant fugu was classified using Mfuzz into significant discrete clusters, respectively, to illustrate relative expression changes of the transcriptome data. Then, cluster analysis was performed, and 9 clusters were obtained. Results indicated that the expression of genes in brain clusters 5, 9, and liver clusters 1, 12 showed a consistently increasing trend with the temperature decrease progression (Figure S2). The up-regulated genes were significantly enriched in 15 KEGG pathways. Regarding the top 50% gene list as standard, 751 and 525 genes were obtained in brain and liver tissues, respectively, which exhibited higher expression with the decrease in temperature.
During the process of low-temperature treatment, signaling pathways involved in energy metabolism and apoptosis, such as p53, MAPK, Ferroptosis, Ubiquitin mediated proteolysis, Adipocytokine, and PPAR signaling pathway were up-regulated in the brain tissues of wild-type. However, in cold-tolerant fugu, the repair and maintenance cell pathways, such as insulin, homologous recombination, mitophagy, FoxO, and mTOR signaling pathway (use programmed cell death to clean damaged cells and repair DNA damage) appeared to be expressed at a higher level (Figure S3). In conclusion, when temperatures dropped from 24 °C to 4 °C, wild-type fugu showed a higher expression of pathways pointed to apoptosis, while cold-tolerant fugu exhibited a significant improvement in DNA damage repair and mitochondrial oxidative stress elimination. This indicated that brain tissue damage during the cooling process could be the main reason for the existence of rollover in wild-type fugu, and there were no significant changes in signaling pathways about apoptosis in the brain tissues of cold-tolerant fugu; thus, cold-tolerant fugu only displayed weakened swimming vigor instead of the phenomenon of rollover.
The liver is the central hub for the regulation of nutrient metabolism and detoxification, thus constituting an excellent target to characterize mechanisms of acclimation to chronically reduced temperature. With temperatures dropping from 24 °C to 4 °C, the cold-tolerant fugu liver signaling pathways involved in energy metabolism, such as the TCA cycle, fatty acid metabolism, and PPAR signaling pathway, were significantly elevated (Figure 2). Therefore, the organism could produce enough energy and calories to maintain body homeostasis and adapt to low temperatures. This might be a potential way for cold-tolerant fugu to cope with cold stress.
3.3. Differential Apoptosis Progression in the Liver and Brain of Wild-Type and Cold-Tolerant Fugu
To further screen which pathways mediated the cold tolerance of fugu, Gene Set Enrichment Analysis (GSEA), a powerful method for interpreting the biological meaning of a list of genes by computing the overlaps with various previously defined gene sets, was performed. The wild-type and cold-tolerant fugu tissues were then compared with standard GSEA, which allows detecting statistically significant expression differences among a given set of genes in various phenotypes or conditions.
Low-temperature stress can induce endogenous reactive oxygen species (ROS) such as hydroxyl radicals, superoxide, and peroxyl radicals. Oxidative stress can trigger DNA damage, protein oxidation, lipid peroxidation, and a decline in physiological function, thus, ultimately leading to apoptosis. The vertebrate brain is one of the most active organs of all and is exquisitely sensitive to perturbations in energy metabolism. Glucose oxidation and mitochondrial oxidative phosphorylation provide most of the ATP required to drive various energetically demanding neuronal activities in teleost. We obtained the most strongly differentiated signaling pathways between the wild-type and cold-tolerant fugu using GSEA.
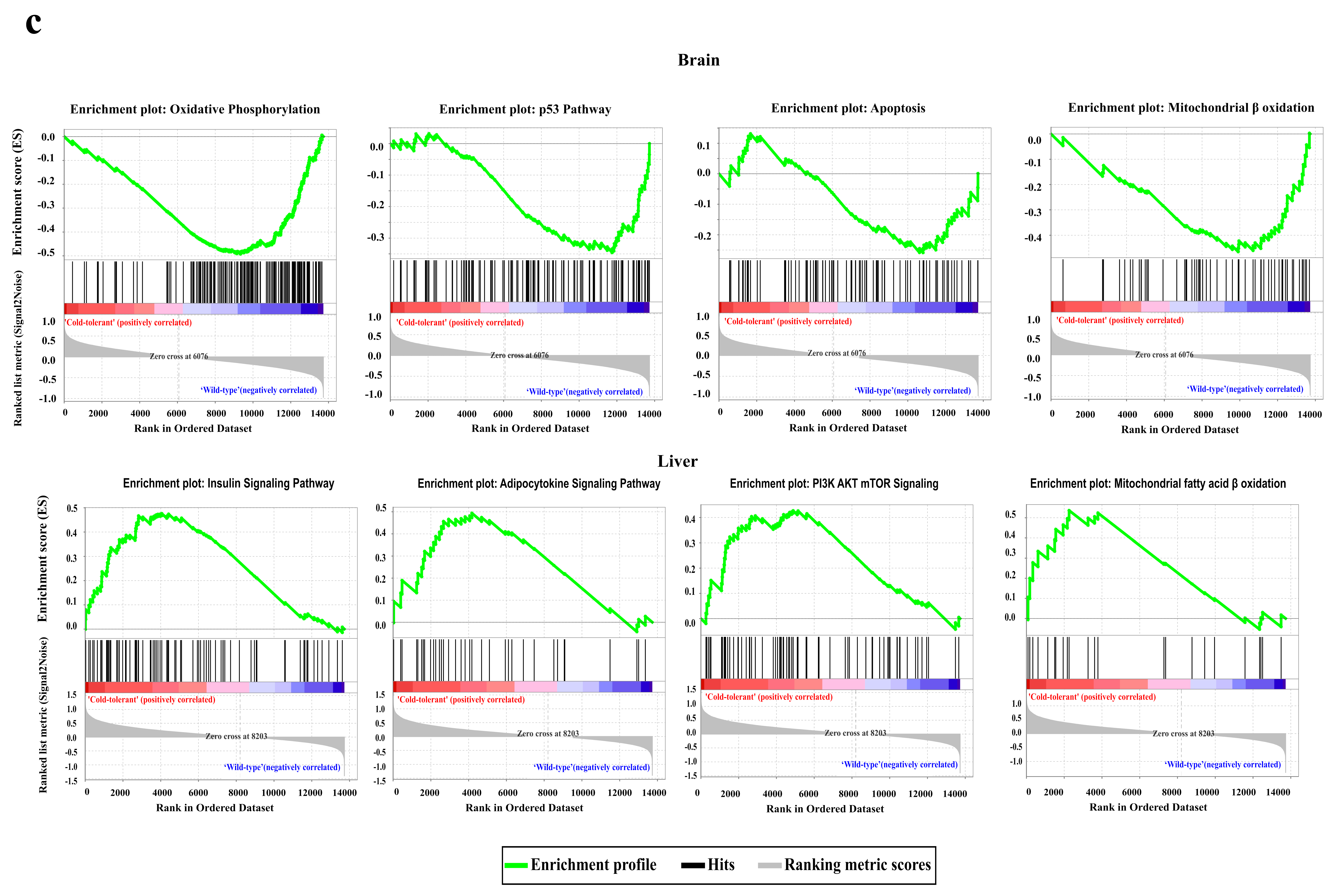
It was clearly demonstrated that oxidative phosphorylation, mitochondrial fatty acid beta-oxidation, and p53 pathway genes were significantly up-expressed in wild-type brain tissues (Figure 3). Compared with the cold-tolerant fugu, the expression of genes regulating oxidative phosphorylation signaling pathways in wild-type brain tissues, such as Cytochrome c oxidase subunit 4 (COX4), Dihydrolipoyllysine-residue succinyl transferase (DLST), Aconitate hydratase (ACO2), Isocitrate dehydrogenase (IDH2), NADH dehydrogenase 2(NDUFA2), and ATP synthase subunit gamma (ATP5F1C) increased by 2 or 3 times at 16 °C, and the maximum expression difference of wild-type fugu appeared at 7 °C (Figure 3).
Considering that the brain is one of the most active organs in vertebrate oxidation of glucose and oxidative phosphorylation, which provide most of the ATP required for brain function in teleost at low temperatures, the high metabolic rate of brain cells implies a high production of reactive oxygen species (ROS) [15,16]. The adaptive shift in metabolic and neuroprotective mechanisms is crucial for satisfying large energy demands in brains under cold stress. Obviously, these functional genes are involved in mitochondrial-related oxidative phosphorylation, and the critical step for intrinsic apoptosis is irreversible mitochondrial outer membrane permeabilization (MOMP), through which proteins of the intermembrane space, such as Voltage-dependent anion-selective channel protein 2 (VDAC2) form a channel through the mitochondrial outer membrane that allows diffusion of small hydrophilic molecules. Cytochrome c oxidase subunit 4 (COX4), a component of the cytochrome c oxidase, drives oxidative phosphorylation that creates an electrochemical gradient over the inner membrane that drives transmembrane transport or ATP synthase. ATP synthase subunit gamma (ATP5F1C), a mitochondrial membrane ATP synthase, produces ATP from ADP in the presence of a transmembrane proton which is generated by electron transport complexes of the respiratory chain. Generation of reactive oxygen species (ROS), as well as apoptosis initiation through the activation of the mitochondrial permeability transition pore and mitochondrial outer membrane permeabilization (MOMP), can both work in the apoptotic machinery of fish. Meanwhile, we enriched p53 and apoptosis pathway genes that are robustly up-regulated in brain tissues at low temperatures of wild-type fugu. As regards p53, it can induce ROS generation, thus stimulating apoptosis and DNA damage (Figure 3). MOMP is the point of no return in many forms of apoptotic cell death since it initiates both proteolytic cascades of pro-apoptotic enzymes and damages mitochondrial function. The expression patterns of these genes indicated that oxidative phosphorylation (OXPHOS) could be the crucial metabolic mitochondrial process that employs enzymes to oxidize nutrients, thereby releasing energy that is used for cell respiration and (ATP) synthesis during the cooling process. Excessive levels of mitochondrial metabolic process mediated the production of ATP accompanied by the robust ROS, which in turn increased the apoptosis in the brain and caused coma and lost swimming ability in wild-type fugu under cold stress.
In contrast, most assays were performed on samples from the fish liver because the liver plays a central role in the metabolism, storage, and redistribution of processed carbohydrates, proteins, and lipids in fish. As temperatures decreased from 24 °C to 16 °C, the energy metabolism in the liver of cold-tolerant fugu became stronger under cold stress. In cold-tolerant fugu, the expression of key genes involved in the insulin signaling pathway, adipocytokines, and mTOR signaling pathway in the liver was observed to have a significantly higher expression than in the wild-type, for the pathways mentioned above were pointed to the regulation of glycogen and lipid metabolism in the liver at 7 °C (Figure 3). The insulin signaling pathway regulates glucose metabolism through its actions in the liver. Some genes are classically known to be involved in the metabolic pathway regulated by the insulin signaling pathway. RAC-alpha serine/threonine-protein kinase (ATK1) is an important functional gene that plays a key role in regulating cellular energy metabolism. Sterol regulatory element-binding protein 1 (SREBP1) and Peroxisome proliferator-activated receptor gamma (PPARγ) are two genes that contribute to the modulation of cholesterol biosynthesis, lipid homeostasis, and insulin sensitivity, which are key markers of hepatic lipid metabolism. Another gene that participates in the rate-limiting step of glycolysis is called Hexokinase-1 (HK1), whose up-regulation can increase the release of mitochondrial membrane depolarization in cold-tolerant fugu by 3-fold at 7 °C (Figure 3). The liver provides energy-supplying substances for the whole body, which regulates the blood glucose concentration constantly through glycogen synthesis, glycogenolysis, and gluconeogenesis processes, and converts glucose into fat or secretes glucose into the blood in the form of very-low-density lipoprotein when the energy supply is sufficient. This process is an important source of fatty acids for the synthesis of triglycerides in adipose tissue. However, during a fasting or starving period, the liver turns many fatty acids mobilized from adipose tissue into ketone bodies for the organism to cope with environmental stress. In our study, results clearly showed that none of the apoptosis-related functional groups were over-expressed in cold-tolerant fugu.
When the temperature decreased, the fugu organism increased its energy metabolism rate to cope with cold stress; therefore, the peroxidation reaction and ROS accelerated the formation of apoptosis and apoptosis-related signaling pathways in brain and liver tissues. Comparing the transcriptome analysis of the wild-type and cold-tolerant fugu, it was found that the expression of genes such as Peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC1A), carnitine O-palmitoyl transferase 1 (CPT1B), pro-opiomelanocortin (POMC), mitogen-activated protein kinase kinase kinase 7 (MAP3K7), mothers against decapentaplegic homolog 2 (SMAD2), 3-ketoacyl-CoA thiolase (ACAA2), propionyl-CoA carboxylase alpha chain (PCCA), long-chain specific acyl-CoA dehydrogenase (ACADVL), and leptin receptor (LEPR) were significantly higher in cold-tolerant fugu, while apoptosis-related genes exhibited more expression in the wild-type at temperatures from 16 to 7 °C (Figure 4). In the cooling period, the brain was the most important organ, which played a key role in regulating fugu homeostasis and survival. Once the brain lost its function, the fugu would fall into a coma or die of shock directly. For the cold-tolerant fugu, the higher energy metabolism level of the liver, which produces energy in the form of glycogenolysis and fatty acids catabolism, ensures the energy demands of brain or muscle tissues are met and ultimately avoids dangerous situations for fish under hypothermia.
3.4. Model of Cold Tolerance Mechanism of Fugu
After 5 generations of low-temperature domestication for 10 consecutive years, the cold-tolerant variety “Zhongyang No.1” fugu has become more resistant to cold stress. In the experiment, with temperatures dropping from 24 °C to 16 °C, cold-tolerant fugu still had normal swimming ability and survived without coma at 7 °C. In the cooling experiment, we observed that half of the wild-type fugu died when the temperature dropped to 12 °C after 12 h, while cold-tolerant fugu survived. We further explored the mechanism of cold tolerance by examining gene expression levels of signaling pathways.
In wild-type fugu, we revealed higher expression of pathways such as mitochondrial fatty acid β oxidation, oxidative phosphorylation, and p53 pathway in brain tissues under lower temperature, which could induce apoptosis in the central nervous system (CNS) and brain damage, leading to coma or death of wild-type fugu (Figure 4). Surprisingly, when it comes to cold-tolerant fugu, we did not detect visible changes in the expression of pathways paralleled to that in wild-type brains under cold conditions. Meanwhile, the expression of pathways such as insulin signaling pathway, adipocytokines, mitochondrial fatty acid β oxidation, and AKT mTOR pathway significantly increased in the liver, which indicated the generation of liver glycogenolysis and fatty acids catabolism in cold-tolerant fugu when facing the cold.
We suppose that in cold-tolerant fugu, there exists a kind of protection strategy stimulated by lower temperature, where the liver mobilizes various energy substances to improve energy metabolism and, to some extent, protect the brain from damage with its active energy metabolism (Figure 5). As a result, cold-tolerant fugu can still live in colder temperatures compared to the wide type.
4. Discussion
The domestication of animals involves the transformation of morphological, physiological, developmental, and behavioral features, during which animals adapt to the environment that humans create for them [17]. Domestication is a key element for allowing sustainable expansion of aquacultural production, which is a priority of the sector in view of the ever-growing demand of consumers and constantly increasing human population [2,18,19]. Domestication will generationally lead to corresponding changes at the physiological and molecular level, thereby affecting a variety of biological functions in animals. There are abundant essays about the domestication effects on growth and stress response in teleosts [20,21]. It has been reported that domestication reduces fish stress response, manifesting less fear in salmonids and European sea bass or less sensitiveness to acute handling stressors in Eurasian perch and catfish [22,23]. The cold-tolerant fugu used in this experiment also obtained low-temperature domestication. After 5 generations of domestication, the semi-lethal temperature of cold-tolerant fugu was 6.2 °C, and the experimental fugu were equipped with an adaptation to the low temperatures at 7 °C. The underlying molecular mechanisms connecting physiological and transcriptional changes produced by domestication in response to low-temperature stress have yet to be revealed.
Temperature controls and limits all physiological and behavioral parameters of ectotherms; water temperature has been described as the ‘abiotic master factor’ for fishes [24]. At a range of non-lethal temperatures, fish have the ability to cope with gradual changes that are common in natural systems [25]. After artificial selection by gradually reducing the temperature with each generation, we eventually obtained the low-temperature domesticated fugu, which could effectively adapt to the cold environment. For most teleosts, the cold stress response is generally considered to be a kind of adaptive response to maintain homeostasis. Prolonged or severe temperature changes that occur outside a specific tolerance may ultimately result in mortality or otherwise affect the organism’s fitness [26,27]. Present knowledge of primary responses to cold stress has largely been gained by studying responses in the hypothalamic–pituitary–interrenal (HPI) axis and brain tissues. The vertebrate brain is metabolically one of the most active organs and is exquisitely sensitive to perturbations of energy metabolism. In fact, approximately 50–60% of the ATP consumed by the brain is devoted to its electrical activity [28]. Glucose oxidation and mitochondrial oxidative phosphorylation provide most of the ATP required to drive various energetically demanding neuronal activities in teleost. Since most physiological adaptation responses are initiated by the CNS, the brain is probably the organ most susceptible to thermal fluctuations [28,29,30].
At the temperature of Antarctic seawater, for instance, depression of nerve activity causes temperate ectotherm animals such as fish to get into trouble, though remaining quite active and well-coordinated. There must be some sort of cold compensation allowing higher rates of metabolism at low temperatures [31,32]. Thus, high rates of oxygen uptake have been observed in isolated brain slices of teleost fishes, and enzymes from the brains of Antarctic teleosts are more active at very low temperatures than those from animals adapted to warmer temperatures [31,33,34,35], suggesting that partial compensation to temperature may exist in ATP producing pathways in fish brains. A study on green sunfish (Lepomis cyanellus) demonstrates that activities of glycolytic enzymes such as glucose phosphate isomerase, glyceraldehyde phosphate dehydrogenase, and pyruvate kinase are elevated in the brain during cold exposure [15,36].
In our study of cold-tolerant fugu, we found that cold stress signaling pathways, such as insulin and PPAR signaling pathway, were significantly enriched in the brain, acting as a necessary regulator of lipid metabolism such as fatty acid biosynthesis and inositol phosphate metabolism. As expected, low temperature mobilized energy metabolic pathways in the brain. Energy generators of the cell through oxidative phosphorylation and electron transfer lead to the production of ATP [37,38]. In addition, reactive oxygen species (ROS), including the hydroxyl radical (OH) and the superoxide anion (O2−), are produced via mitochondrial bioenergetics [37,39].
Through transcriptome studies, we found that HK1 and VDAC2 were significantly up-regulated in the brain of wild-type fugu. The rate-limiting step of glycolysis is controlled by HK, which can convert glucose to glucose-6-phosphate in an ATP-dependent manner. HK’s tight association with VDAC provides ready access to mitochondrial-generated ATP [37,40], disruption of which can induce mitochondrial permeability transition (MPT). Oxidative stress mediating mitochondrial membrane permeabilization leads to permeability transition pore complex (PTPC) opening, which causes MPT [41]. Osmotic swelling and rupture of mitochondrial membranes, as well as the release of mitochondrial intermembrane proteins, such as CytC and apoptosis-inducing factor (AIF) after complete MPT, can eventually lead to cell death [41,42]. Meanwhile, we also observed enrichment of p53 and apoptotic pathways in brain tissues of wild-type fugu at temperatures from 16 to 7 °C. In the brain, apoptotic gene patterns initiated by the cold stress led to up-regulation of the initiator Caspase 9, followed by proteolytic maturation of the executioner Caspase 3 (Figure 3). As the temperature decreased, the number of apoptotic cells in the brain gradually increased, and fugu were able to show equilibrium at 12 °C while completely losing equilibrium and falling into a coma at 7 °C.
The liver is an essential metabolic organ whose metabolic activity is tightly controlled by insulin and other metabolic hormones. Glucose is metabolized into pyruvate through glycolysis in the cytoplasm, with pyruvate completely oxidized to generate ATP through the TCA cycle and oxidative phosphorylation in mitochondria [43]. In our study, we found that PIK3, AKT (AKT1, AKT2), SREBP1, PPARγ, and leptin receptor (LEPR) involved in that insulin signaling pathway were significantly enriched in the liver of cold-tolerant fugu, and their expression was also significantly higher than that of wild-type fugu. Hepatocytes have great flexibility in selecting metabolic fuels glucose and/or fatty acids; thus, insulin signaling is the primary hormone that stimulates hepatic lipogenesis in the body [44]. The PI3-kinase/Akt pathway is required for both insulin suppression of gluconeogenesis and stimulation of lipogenesis. However, lipogenesis and gluconeogenesis are mediated by two distinct pathways downstream of Akt [45]. Insulin stimulates the activation of mTORC1 through the PI3-kinase/Akt pathway, and mTORC1 is required for insulin to stimulate SREBP-1 expression and lipogenesis. Akt, particularly Akt 2, stimulates SREBP-1 activation and lipogenesis. Under cold stress, transcription factors and coactivators involved in energy regulation, including retinoid X receptor (RXR), SREBP1, and PPARγ, have the ability to control the expression of the enzymes that catalyze the rate-limiting steps of liver metabolic processes. The expression of transcription factors is significantly higher in the livers of cold-tolerant than that of wild-type fugu, thus strongly controlling liver energy metabolism.
Several lines of evidence suggest that regulation of mTORC1 signaling is crucial for the response in the liver under cold stress [46,47]. When blood glucose drops with the lowering temperature, the liver activates a compensatory response involving the induction of autophagy, gluconeogenesis, and the release of alternative energy sources in the form of ketone bodies [48]. Similarly, our study found that genes of the mTOR signaling pathway in the liver of cold-tolerant fugu were significantly up-regulated in expression under cold stress compared to that of wild-type fugu, and so was the Akt, Tuberous Sclerosis Complex1 (TSC1), a suppressor gene of mTORC1, succeed to generate ketone bodies by activating the expression of PPARα [49]. mTORC1 functions as a downstream effector for many pathways, including the PI3K/Akt pathway and the Ras/Raf/Mek/Erk (MAPK) pathways, each of which was most strongly expressed at 7 °C and 4 °C. Therefore, cold stress enhanced mTORC1-involved glucose metabolism and lipid metabolism in the liver of cold-tolerant fugu.
The production of heat and increased metabolic rate inevitably lead to increasing the production of adenosine triphosphate (ATP), which is mainly brought by mitochondrial oxidative phosphorylation [50]. Inevitably, the production of partially reduced oxygen and the formation of ROS occurs. The energetic cost of maintaining ‘primed’ mitochondria, together with the deleterious effects of reactive oxygen species (ROS) generated during electron transport, make the possession of mitochondria a double-edged sword [51]. For fish living at low temperatures, the increased polyinstantiation of mitochondrial membranes can raise rates of mitochondrial respiration, which, in turn, enhances the formation of reactive oxygen species (ROS), elevates proton leak, and favors peroxidation of these membranes. When the production of ROS is beyond the organism’s capacity to deal with these reactive species, oxidative stress can induce the production of apoptosis and result in the loss of function of organs.
Compared with cold-tolerant fugu, the level of expression of signaling pathways and genes involved in energy metabolism in their liver was significantly higher than that of wild-type fugu, which could provide better sufficient energy for the organism. The brain, possessing the most important central nervous system of the organism, is an organ that consumes the largest number of ATP [52]. The expression of apoptosis-related signaling pathways such as p53 and MAPK is higher in wild-type fugu at low temperatures, which could cause irreversible damage to the brain tissue and induce coma and death (Figure 5). The domestication of cold-tolerant fugu uses a “sacrifice minor things to save major ones” strategy to ensure the survival of organisms under low temperature, which provides a reference for our research on the dynamic response of fish under cold stress in aquaculture.
In our study, some signaling pathways involved in apoptosis and energy metabolism, such as MAPK, oxidative phosphorylation, and mitochondrial β-oxidation, which are enriched in brain and liver tissues, are usually modified by phosphorylation and methylation, therefore regulating the body’s homeostasis in response to cold. For the next step, we will identify these genes at the level of transcriptional regulation and epigenetic modifications to provide a deeper understanding of the mechanisms of cold tolerance in fish.
5. Conclusions
After artificial selection we obtained domesticated cold-tolerant fugu which could effectively adapt to the cold environment. Compared to the wild type, cold-tolerant fugu had no obvious subcutaneous congestion and frostbite phenotype at 12 °C. It even survived normally at 7 °C (with reduced swimming), and showed good adaptability to cold stress. The transcriptomic study revealed that in the brain of cold-tolerant fugu, the expression of signaling pathways involved in apoptosis, oxidative phosphorylation and mitochondrial β-oxidation was significantly lower than the wild-type, while signaling pathways involved in energy metabolism in response to cold stress, such as insulin, adipocytokines and PI3K AKT mTOR signaling pathways were significantly upregulated in the liver of cold-tolerant fugu, indicating that the liver provided energy in response to cold stress to compensate for the lack of energy supply to the brain, which may result in excessive generation of ROS. This phenomenon suggested that the domesticated cold-tolerant fugu respond to the low temperatures by mobilizing the dynamic regulatory mechanism of liver metabolism to reduce the effects of cold stress on brain apoptosis.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/fishes7040159/s1. Figure S1: LT50 of wild-type and cold-tolerant fugu; Figure S2: Mfuzz to clusters of brain and liver expression data in wild-type and cold-tolerant fugu. Figure S3: KEGG pathways for the cold-tolerant brain and wild-type liver genes.
Author Contributions
S.H. and L.C. conceived and designed the scientific objectives. M.N. collected and prepared the fish samples. S.W. and R.C. conducted the experiment and finished the bioinformatics analysis. M.N. and R.C. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the Natural Science Foundation of China (32130109) and the National Key Research and Development Program of China (2018YFD0900601). Fugu were treated appropriately to minimize suffering.
Institutional Review Board Statement
All experimental protocols in this study were approved by The Scientific Ethics Committee of Shanghai Ocean University, Shanghai, China (SHOU-DW-20171022).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data available upon request from authors.
Acknowledgments
The research was supported by Zhongyang Group, Jiangsu, China. We thank Yuping Lu and Bingbing Yan for their assistance with this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Anastasiadi, D.; Piferrer, F. Epimutations in Developmental Genes Underlie the Onset of Domestication in Farmed European Sea Bass. Mol. Biol. Evol. 2019, 36, 2252–2264. [Google Scholar] [CrossRef] [PubMed]
- Teletchea, F.; Fontaine, P. Levels of Domestication in Fish: Implications for the Sustainable Future of Aquaculture. Fish Fish. 2014, 15, 181–195. [Google Scholar] [CrossRef]
- Lorenzen, K.; Beveridge, M.C.M.; Mangel, M. Cultured Fish: Integrative Biology and Management of Domestication and Interactions with Wild Fish. Biol. Rev. 2012, 87, 639–660. [Google Scholar] [CrossRef] [PubMed]
- Milla, S.; Pasquet, A.; El Mohajer, L.; Fontaine, P. How Domestication Alters Fish Phenotypes. Rev. Aquac. 2021, 13, 388–405. [Google Scholar] [CrossRef]
- Price, E.O. Behavioral Development in Animals Undergoing Domestication. Appl. Anim. Behav. Sci. 1999, 65, 245–271. [Google Scholar] [CrossRef]
- Abram, Q.; Dixon, B.; Katzenback, B. Impacts of Low Temperature on the Teleost Immune System. Biology 2017, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Tan, X.; Xu, M.; Liu, Q.; Ye, H.; Zou, C.; Ye, C. Liver Transcriptome Analysis and de Novo Annotation of the Orange-Spotted Groupers (Epinephelus coioides) under Cold Stress. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 29, 264–273. [Google Scholar] [CrossRef]
- Sunday, J.M.; Bates, A.E.; Dulvy, N.K. Thermal Tolerance and the Global Redistribution of Animals. Nat. Clim. Change 2012, 2, 686–690. [Google Scholar] [CrossRef]
- Wen, X.; Hu, Y.; Zhang, X.; Wei, X.; Wang, T.; Yin, S. Integrated Application of Multi-Omics Provides Insights into Cold Stress Responses in Pufferfish Takifugu fasciatus. BMC Genom. 2019, 20, 563. [Google Scholar] [CrossRef] [Green Version]
- Long, Y.; Song, G.; Yan, J.; He, X.; Li, Q.; Cui, Z. Transcriptomic Characterization of Cold Acclimation in Larval Zebrafish. BMC Genom. 2013, 14, 612. [Google Scholar] [CrossRef] [Green Version]
- Vornanen, M.; Hassinen, M.; Koskinen, H.; Krasnov, A. Steady-State Effects of Temperature Acclimation on the Transcriptome of the Rainbow Trout Heart. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R1177–R1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, X.; Zhang, X.; Hu, Y.; Xu, J.; Wang, T.; Yin, S. ITRAQ-Based Quantitative Proteomic Analysis of Takifugu Fasciatus Liver in Response to Low-Temperature Stress. J. Proteom. 2019, 201, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, L.; Futschik, M.E. Mfuzz: A Software Package for Soft Clustering of Microarray Data. Bioinformation 2007, 2, 5–7. [Google Scholar] [CrossRef]
- Tseng, Y.-C.; Chen, R.-D.; Lucassen, M.; Schmidt, M.M.; Dringen, R.; Abele, D.; Hwang, P.-P. Exploring Uncoupling Proteins and Antioxidant Mechanisms under Acute Cold Exposure in Brains of Fish. PLoS ONE 2011, 6, e18180. [Google Scholar] [CrossRef] [Green Version]
- Blomgren, K.; Hagberg, H. Free Radicals, Mitochondria, and Hypoxia-Ischemia in the Developing Brain. Free Radic. Biol. Med. 2006, 40, 388–397. [Google Scholar] [CrossRef]
- Palińska-Żarska, K.; Woźny, M.; Kamaszewski, M.; Szudrowicz, H.; Brzuzan, P.; Żarski, D. Domestication Process Modifies Digestion Ability in Larvae of Eurasian Perch (Perca fluviatilis), a Freshwater Teleostei. Sci. Rep. 2020, 10, 2211. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.; Nguyen, T.; Turchini, G.M.; Amarasinghe, U.S.; Abery, N.W. Alien Species in Aquaculture and Biodiversity: A Paradox in Food Production. Ambio 2009, 38, 24–28. [Google Scholar] [CrossRef]
- Badiola, M.; Mendiola, D.; Bostock, J. Recirculating Aquaculture Systems (RAS) Analysis: Main Issues on Management and Future Challenges. Aquac. Eng. 2012, 51, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Tymchuk, W.E.; Biagi, C.; Withler, R.; Devlin, R.H. Growth and Behavioral Consequences of Introgression of a Domesticated Aquaculture Genotype into a Native Strain of Coho Salmon. Trans. Am. Fish. Soc. 2006, 135, 442–455. [Google Scholar] [CrossRef]
- Hassin, S.; De Monbrison, D.; Hanin, Y.; Elizur, A.; Zohar, Y.; Popper, D. Domestication of the White Grouper, Epinephelus Aeneus 1. Growth and Reproduction. Aquaculture 1997, 156, 305–316. [Google Scholar] [CrossRef]
- Douxfils, J.; Mathieu, C.; Mandiki, S.N.M.; Milla, S.; Henrotte, E.; Wang, N.; Vandecan, M.; Dieu, M.; Dauchot, N.; Pigneur, L.-M.; et al. Physiological and Proteomic Evidences That Domestication Process Differentially Modulates the Immune Status of Juvenile Eurasian Perch (Perca fluviatilis) under Chronic Confinement Stress. Fish Shellfish Immunol. 2011, 31, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Adloo, M.N.; Soltanian, S.; Hafeziyeh, M.; Ghadimi, N. Cortisol and Glucose Responses in Juvenile Striped Catfish Subjected to a Cold Shock. Vet. Sci. Dev. 2015, 5, 5892. [Google Scholar] [CrossRef]
- Brett, J.R. Energetic Responses of Salmon to Temperature. A Study of Some Thermal Relations in the Physiology and Freshwater Ecology of Sockeye Salmon (Oncorhynchus nerkd). Am. Zool. 1971, 11, 99–113. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, M.; Cooke, S.; Patterson, D.; Macdonald, J. Cold shock and fish. J. Fish Biol. 2008, 73, 1491–1530. [Google Scholar] [CrossRef]
- Schulte, P.M. What Is Environmental Stress? Insights from Fish Living in a Variable Environment. J. Exp. Biol. 2014, 217, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Soyano, K.; Mushirobira, Y. The Mechanism of Low-Temperature Tolerance in Fish. In Survival Strategies in Extreme Cold and Desiccation; Springer: Berlin/Heidelberg, Germany, 2018; pp. 149–164. [Google Scholar]
- Soengas, J.L.; Aldegunde, M. Energy Metabolism of Fish Brain. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2002, 131, 271–296. [Google Scholar] [CrossRef]
- Driedzic, W.R. Low Plasma Glucose Limits Glucose Metabolism by RBCs and Heart in Some Species of Teleosts. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2018, 224, 204–209. [Google Scholar] [CrossRef]
- Crawshaw, L.; Grahn, D.; Wollmuth, L.; Simpson, L. Central Nervous Regulation of Body Temperature in Vertebrates: Comparative Aspects. Pharmacol. Ther. 1985, 30, 19–30. [Google Scholar] [CrossRef]
- Kawall, H.; Torres, J.; Sidell, B.; Somero, G. Metabolic Cold Adaptation in Antarctic Fishes: Evidence from Enzymatic Activities of Brain. Mar. Biol. 2002, 140, 279–286. [Google Scholar]
- Chen, Z.; Cheng, C.-H.C.; Zhang, J.; Cao, L.; Chen, L.; Zhou, L.; Jin, Y.; Ye, H.; Deng, C.; Dai, Z. Transcriptomic and Genomic Evolution under Constant Cold in Antarctic Notothenioid Fish. Proc. Natl. Acad. Sci. USA 2008, 105, 12944–12949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somero, G.N.; Fields, P.A.; Hofmann, G.E.; Weinstein, R.B.; Kawall, H. Cold Adaptation and Stenothermy in Antarctic Notothenioid Fishes: What Has Been Gained and What Has Been Lost? In Fishes of Antarctica; Springer: Berlin/Heidelberg, Germany, 1998; pp. 97–109. [Google Scholar]
- Daane, J.M.; Detrich, H.W., III. Adaptations and Diversity of Antarctic Fishes: A Genomic Perspective. Annu. Rev. Anim. Biosci. 2022, 10, 39–62. [Google Scholar] [CrossRef]
- Hotaling, S.; Desvignes, T.; Sproul, J.S.; Lins, L.S.F.; Kelley, J.L. Pathways to Polar Adaptation in Fishes Revealed by Long-read Sequencing. Mol. Ecol. 2022, in press. [CrossRef] [PubMed]
- Shaklee, J.B.; Christiansen, J.A.; Sidell, B.D.; Prosser, C.L.; Whitt, G.S. Molecular Aspects of Temperature Acclimation in Fish: Contributions of Changes in Enzyme Activities and Isozyme Patterns to Metabolic Reorganization in the Green Sunfish. J. Exp. Zool. 1977, 201, 1–20. [Google Scholar] [CrossRef]
- Dawson, T.M.; Dawson, V.L. Mitochondrial Mechanisms of Neuronal Cell Death: Potential Therapeutics. Annu. Rev. Pharmacol. Toxicol. 2017, 57, 437–454. [Google Scholar] [CrossRef] [PubMed]
- Nath, S.; Villadsen, J. Oxidative Phosphorylation Revisited. Biotechnol. Bioeng. 2015, 112, 429–437. [Google Scholar] [CrossRef]
- Nohl, H.; Gille, L.; Staniek, K. Intracellular Generation of Reactive Oxygen Species by Mitochondria. Biochem. Pharmacol. 2005, 69, 719–723. [Google Scholar] [CrossRef]
- Chiara, F.; Castellaro, D.; Marin, O.; Petronilli, V.; Rasola, A. Hexokinase II Detachment from Mitochondria Triggers Apoptosis through the Permeability Transition Pore Independent of Voltage-Dependent Anion Channels. PLoS ONE 2008, 3, e1852. [Google Scholar] [CrossRef] [Green Version]
- Orrenius, S.; Gogvadze, V.; Zhivotovsky, B. Calcium and Mitochondria in the Regulation of Cell Death. Biochem. Biophys. Res. Commun. 2015, 460, 72–81. [Google Scholar] [CrossRef]
- Foskett, J.K.; Philipson, B. The Mitochondrial Ca2+ Uniporter Complex. J. Mol. Cell. Cardiol. 2015, 78, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Rui, L. Energy Metabolism in the Liver. Compr. Physiol. 2014, 4, 177. [Google Scholar]
- Li, Y.U.; Xu, S.; Mihaylova, M.M.; Zheng, B.; Hou, X.; Jiang, B.; Park, O.; Luo, Z.; Lefai, E.; Shyy, J.Y.-J.; et al. AMPK Phosphorylates and Inhibits SREBP Activity to Attenuate Hepatic Steatosis and Atherosclerosis in Diet-Induced Insulin-Resistant Mice. Cell Metab. 2011, 13, 376–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warne, J.; Alemi, F.; Reed, A.; Varonin, J.; Chan, H.; Piper, M.; Mullin, M.; Myers, M.; Corvera, C.; Xu, A. Impairment of Central Leptin-Mediated PI3K Signaling Manifested as Hepatic Steatosis Independent of Hyperphagia and Obesity. Cell Metab. 2015, 21, 648. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-W.; Storey, K.B. MTOR Signaling in Metabolic Stress Adaptation. Biomolecules 2021, 11, 681. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Yang, J.; Yang, L. Insights for Oxidative Stress and MTOR Signaling in Myocardial Ischemia/Reperfusion Injury under Diabetes. Oxidative Med. Cell. Longev. 2017, 2017, 6437467. [Google Scholar] [CrossRef] [Green Version]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [Green Version]
- Dibble, C.; Elis, W.; Menon, S.; Qin, W.; Klekota, J.; Asara, J.; Finan, P.; Kwiatkowski, D.; Murphy, L.; Manning, B. TBC1D7 Is a Third Subunit of the TSC1-TSC2 Complex Upstream of mTORC1. Mol. Cell 2012, 47, 535–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolfi-Donegan, D.; Braganza, A.; Shiva, S. Mitochondrial Electron Transport Chain: Oxidative Phosphorylation, Oxidant Production, and Methods of Measurement. Redox Biol. 2020, 37, 101674. [Google Scholar] [CrossRef]
- Guderley, H. Metabolic Responses to Low Temperature in Fish Muscle. Biol. Rev. 2010, 79, 409–427. [Google Scholar] [CrossRef] [PubMed]
- Ju, Z.; Dunham, R.; Liu, Z. Differential Gene Expression in the Brain of Channel Catfish (Ictalurus punctatus) in Response to Cold Acclimation. Mol. Genet. Genom. 2002, 268, 87–95. [Google Scholar] [CrossRef]
Figure 1.
Phenotypic distribution of cold-tolerant fugu and the model of the cooling experiment. (a)The temperature treatment model of the fugu cold stress experiment. (b)The phenotypic changes of fugu under low-temperature treatment can be observed. The subcutaneous congestion and frostbite are obvious in wild-type fugu.
Figure 1.
Phenotypic distribution of cold-tolerant fugu and the model of the cooling experiment. (a)The temperature treatment model of the fugu cold stress experiment. (b)The phenotypic changes of fugu under low-temperature treatment can be observed. The subcutaneous congestion and frostbite are obvious in wild-type fugu.

Figure 2.
Global overview of differentially expressed genes at cold temperature in wild-type and cold-tolerant fugu. (a,b) Cluster analysis of differentially expressed genes in cold-tolerant fugu. KEGG pathway enrichments for the genes differentially expressed under cold stress. (c) Bubble plots indicate the enriched KEGG pathways for the wild-type of brain up-regulated (right-top panel) and cold-tolerant of liver up-regulated (right-bottom panel) genes. The Mfuss enriched up-regulated genes at different temperatures under cold stress in wild-type brain and cold-tolerant liver tissues were used for analysis.
Figure 2.
Global overview of differentially expressed genes at cold temperature in wild-type and cold-tolerant fugu. (a,b) Cluster analysis of differentially expressed genes in cold-tolerant fugu. KEGG pathway enrichments for the genes differentially expressed under cold stress. (c) Bubble plots indicate the enriched KEGG pathways for the wild-type of brain up-regulated (right-top panel) and cold-tolerant of liver up-regulated (right-bottom panel) genes. The Mfuss enriched up-regulated genes at different temperatures under cold stress in wild-type brain and cold-tolerant liver tissues were used for analysis.

Figure 3.
Comparing brain and liver activation-related gene expressions between wild-type and cold-tolerant fugu under cold stress. (a,b) Analyses of gene expression of the GSEA for brain (top) and liver (bottom) under cold stress. (c) Genesets enriched the FDR < 0.05 pathways: oxidative phosphorylation (NES = −2.27), p53 (NES = −1.51), apoptosis (NES = −1.08), mitochondrial fatty acid β oxidation (NES = −1.81) in brain. Insulin signaling (NES = 1.52), adipocytokine (NES = 1.45), PI3K AKT mTOR (NES = 1.36), mitochondrial fatty acid β oxidation (NES = 1.45) in liver; NES, normalized enrichment score. FDR, false discovery rate. Positive and negative NES indicate higher and lower expression in the brain and liver, respectively.
Figure 3.
Comparing brain and liver activation-related gene expressions between wild-type and cold-tolerant fugu under cold stress. (a,b) Analyses of gene expression of the GSEA for brain (top) and liver (bottom) under cold stress. (c) Genesets enriched the FDR < 0.05 pathways: oxidative phosphorylation (NES = −2.27), p53 (NES = −1.51), apoptosis (NES = −1.08), mitochondrial fatty acid β oxidation (NES = −1.81) in brain. Insulin signaling (NES = 1.52), adipocytokine (NES = 1.45), PI3K AKT mTOR (NES = 1.36), mitochondrial fatty acid β oxidation (NES = 1.45) in liver; NES, normalized enrichment score. FDR, false discovery rate. Positive and negative NES indicate higher and lower expression in the brain and liver, respectively.


Figure 4.
qRT-PCR and Gene Set Enrichment Analysis ridge plot for significant KEGG terms, dynamic changes in the balance of gene regulation in liver and brain as a strategy for cold-tolerant fugu to cope with cold stress. (a) qRT-PCR detection of the expression of p53, caspase3, cpt1b, pparγ genes in the brain of wild-type fugu. n = 4 biologically independent experiments. * p < 0.05, ** p < 0.01 by unpaired Student’s t test. (b) qRT-PCR detection of the expression of p53, caspase3, cpt1b, pparγ genes in the liver of cold-tolerant fugu. n = 4 biologically independent experiments. * p < 0.05, ** p < 0.01, *** p < 0.001 by unpaired Student’s t test. (c) Ridge plots are density plots of the frequency of log2-fold change values per gene within each enriched KEGG group, which helps to interpret the up- or down-regulation of that KEGG category. X-axis is log2-fold change in expression for genes present in each KEGG category plotted, with positive values indicating up-regulated expression in the brain and negative values indicating down-regulated expression in the liver. Peaks are colored by corrected p-value as shown by the legend. Purple represents the regulation pattern for cold-tolerant, and red represents the regulation pattern of wild-type fugu.
Figure 4.
qRT-PCR and Gene Set Enrichment Analysis ridge plot for significant KEGG terms, dynamic changes in the balance of gene regulation in liver and brain as a strategy for cold-tolerant fugu to cope with cold stress. (a) qRT-PCR detection of the expression of p53, caspase3, cpt1b, pparγ genes in the brain of wild-type fugu. n = 4 biologically independent experiments. * p < 0.05, ** p < 0.01 by unpaired Student’s t test. (b) qRT-PCR detection of the expression of p53, caspase3, cpt1b, pparγ genes in the liver of cold-tolerant fugu. n = 4 biologically independent experiments. * p < 0.05, ** p < 0.01, *** p < 0.001 by unpaired Student’s t test. (c) Ridge plots are density plots of the frequency of log2-fold change values per gene within each enriched KEGG group, which helps to interpret the up- or down-regulation of that KEGG category. X-axis is log2-fold change in expression for genes present in each KEGG category plotted, with positive values indicating up-regulated expression in the brain and negative values indicating down-regulated expression in the liver. Peaks are colored by corrected p-value as shown by the legend. Purple represents the regulation pattern for cold-tolerant, and red represents the regulation pattern of wild-type fugu.

Figure 5.
A schematic model illustrating genetic pathways that promotes cold tolerance at low temperatures in fugu. When the temperature dropped to 12 °C, wild-type fugu would become comatose and eventually die. In the fugu’s brain, the most important regulator and effector of motor activity, significant enrichment in p53 and apoptosis pathways could be easily detected. Physiological changes at temperatures from 7 to 4 °C indicate that cold stress would increase brain damage. In contrast, domesticated cold-tolerant fugu still had the ability to swim normally at 7 °C. Transcriptomic studies have revealed an increase in hepatic genes involved in glycogenolysis and lipid metabolism. We also observed increased levels of oxidative phosphorylation, and a greater supply of energy from the liver would maintain the survival of cold-tolerant fugu in cold stress.
Figure 5.
A schematic model illustrating genetic pathways that promotes cold tolerance at low temperatures in fugu. When the temperature dropped to 12 °C, wild-type fugu would become comatose and eventually die. In the fugu’s brain, the most important regulator and effector of motor activity, significant enrichment in p53 and apoptosis pathways could be easily detected. Physiological changes at temperatures from 7 to 4 °C indicate that cold stress would increase brain damage. In contrast, domesticated cold-tolerant fugu still had the ability to swim normally at 7 °C. Transcriptomic studies have revealed an increase in hepatic genes involved in glycogenolysis and lipid metabolism. We also observed increased levels of oxidative phosphorylation, and a greater supply of energy from the liver would maintain the survival of cold-tolerant fugu in cold stress.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Specific primers for the selected and reference gene.
| Acc. Number | Gene | Primers Sequences (5′ to 3′) | Product Length |
|---|---|---|---|
| NC_042292 | p53 | CCTGGGTAATCGGTGGTAA ATCTGTGGGAGAATGTGGC | 205 |
| NC_042301 | caspase 3 | GACAACAGTCGGGTTCGTCT CCGAGGCTCAAGAACACTTT | 205 |
| NC_042293 | cpt1b | TCTACCTGCTGAGATACACC GAACATCTTCACGAGGGTCA | 116 |
| NC_042303 | pparγ | TCTGAAAGTCCCGTCATGC TTTAACCTGATGGTGCGTCT | 161 |
| NC_042301 | β-actin | AATCGTGCGTGACATCAA CTGGGCAACGGAACCTCT | 155 |
| NW_021821647 | gapdh | TGGCCATCAATGACCCCTTC CCTCTCGTGGAAAACGGTGA | 149 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Han, S.; Wei, S.; Chen, R.; Ni, M.; Chen, L. Tissue-Specific and Differential Cold Responses in the Domesticated Cold Tolerant Fugu. Fishes 2022, 7, 159. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes7040159
AMA Style
Han S, Wei S, Chen R, Ni M, Chen L. Tissue-Specific and Differential Cold Responses in the Domesticated Cold Tolerant Fugu. Fishes. 2022; 7(4):159. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes7040159
Chicago/Turabian StyleHan, Shuang, Shang Wei, Ruoyu Chen, Man Ni, and Liangbiao Chen. 2022. "Tissue-Specific and Differential Cold Responses in the Domesticated Cold Tolerant Fugu" Fishes 7, no. 4: 159. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes7040159