
Aerial Branch Sampling to Detect Forest Pathogens
by
 , , , and
, , , and
Ryan L. Perroy
1,2,* ,
,
Philip Meier
3,
Eszter Collier
2,
Marc A. Hughes
4,5,
Eva Brill
6,
Timo Sullivan
7,
Thomas Baur
3,
Nina Buchmann
3 and
Lisa M. Keith
6 1
Department of Geography & Environmental Science, University of Hawai‘i at Hilo, Hilo, HI 96720, USA
2
Spatial Data Analysis & Visualization Laboratory, University of Hawai‘i at Hilo, Hilo, HI 96720, USA
3
Department of Environmental Systems Science, Institute of Agricultural Sciences, ETH Zurich, 8092 Zurich, Switzerland
4
Pacific Cooperative Studies Unit, University of Hawai‘i at Manoa, Hilo, HI 96720, USA
5
Institute of Pacific Islands Forestry, United States Forest Service, United States Department of Agriculture, Hilo, HI 96720, USA
6
Daniel K. Inouye U.S. Pacific Basin Agricultural Research Center, Agricultural Research Service, United States Department of Agriculture, Hilo, HI 96720, USA
7
Island Conservation, Santa Cruz, CA 95060, USA
*
Author to whom correspondence should be addressed.
Drones 2022, 6(10), 275; https://0-doi-org.brum.beds.ac.uk/10.3390/drones6100275
Submission received: 8 August 2022
/
Revised: 13 September 2022
/
Accepted: 21 September 2022
/
Published: 24 September 2022
(This article belongs to the Special Issue Drones for Biodiversity Conservation)
Abstract
:Diagnostic testing to detect forest pathogens requires the collection of physical samples from affected trees, which can be challenging in remote or rugged environments. As an alternative to traditional ground-based sampling at breast height by field crews, we examined the feasibility of aerially sampling and testing material collected from upper canopy branches using a small unoccupied aerial system (sUAS). The pathogen of interest in this study is Ceratocystis lukuohia, the fungal pathogen responsible for Ceratocystis wilt of ‘ōhi‘a, a vascular wilt disease which has caused widespread mortality to ‘ōhi‘a in native forests across the state of Hawai‘i. To characterize the minimum branch diameter needed to successfully detect the pathogen of interest in infected trees, we tested 63 branch samples (0.8–9.6 cm in diameter) collected from felled trees inoculated with C. lukuohia on Hawai‘i Island. Subsequently, we aerially sampled branches from ten symptomatic ‘ōhi‘a (Metrosideros polymorpha) trees using two different branch sampling systems, the Flying Tree Top Sampler from ETH Zurich and the new Kūkūau branch sampler system introduced in this work, producing 29 branch samples with a maximum diameter of 4.2 cm and length of >2 m. We successfully detected the target fungal pathogen from the collected branches and found that branch diameter, leaf presence and condition, as well as wood moisture content are important factors in pathogen detection in sampled branches. None of the smallest branch samples (those <1 cm in diameter) tested positive for C. lukuohia, while 77% of the largest diameter branch samples (5–10 cm) produced positive results. The Kūkūau branch sampler system is capable of retrieving branches up to 7 cm diameter, providing important capacity for pathogenic research requiring larger diameter samples for successful diagnostic testing. Inconclusive and/or non-detection laboratory results were obtained from sample materials that were either too desiccated or from a branch with asymptomatic leaves, suggesting there is an optimal temporal window for sampling.
1. Introduction
Small unoccupied aerial vehicles (sUAS) play an increasing role in conservation and forestry, primarily through the collection of high-resolution imagery for use in management decisions and situational awareness in densely forested and treacherous environments [1,2,3]. As sUAS platforms become more capable, opportunities exist to move beyond typical imagery collection into more active applications. Here, we present results from sUAS aerial branch sampling operations to detect a forest tree fungal pathogen on Hawai‘i Island using two different systems, the Flying Tree Top Sampler device [4] and the Kūkūau branch sampling system developed for this project. Our objective was to demonstrate that aerial branch sampling via sUAS can produce viable samples to detect forest fungal pathogens.
1.1. Physical Branch Sample Collection and Retrieval
The collection and retrieval of physical samples in forested environments has applications in plant pathology [5,6], the propagation of threatened and endangered species [7,8,9], and canopy nutrient analyses [10,11,12]. Forestry vegetation samples have been collected using a range of methods, from simple handheld pruners to crossbows to sophisticated canopy crane installations [13,14,15,16,17]. These ground-based methods, while proven, are limited in scope and coverage area and can be highly inefficient in densely vegetated rugged areas. Manned helicopters have been successfully used to collect physical samples in remote or otherwise difficult to access locations [18,19], but the inherent dangers and costs associated with these operations limit their utility for routine use. sUAS provide another means of aerial branch sampling, with the potential for increased efficiency over ground-based methods and without the risks associated with complicated low altitude manned aviation operations.
A number of sUAS systems exist for sampling leaves and small branches or twigs, from platforms modified with simple fixed blades to complicated attachments deploying circular saws and gripping mechanisms with autonomous features [4,20,21,22,23]. For a thorough review of recent sUAS systems used in vegetation sampling and engineering design considerations, please see [22,23]. The more capable of these systems use a circular saw for the cutting device, with a maximum reported cut stem diameter of <2.5 cm. While this branch diameter is more than sufficient for applications relating to canopy leaf measurements [23,24], in the case of diagnostic sampling of woody material for vascular tree pathogens, thicker branch diameters may be needed to provide adequate material for successful laboratory analyses. Determining the minimum tree branch diameter needed to confidently detect infection by Ceratocystis lukuohia [25] was one of the goals of this study.
1.2. Sampling Application: Ceratocystis wilt of ‘ōhi‘a in Hawai’i
In Hawai‘i, the introduced fungal pathogen Ceratocystis lukuohia I. Barnes, T.C. Harr. and L.M. Keith is causing widespread mortality to the native keystone tree species Metrosideros polymorpha Gaud. (‘ōhi‘a) [26,27]. C. lukuohia is introduced to ‘ōhi‘a trees through wounds and causes a systemic vascular wilt disease that spreads throughout the infected tree [28,29]. While remote sensing imagery can be used to identify ‘ōhi‘a trees suspected of infection [30,31,32] (Figure 1), physical samples are required for laboratory testing to confirm presence of the pathogen’s DNA as the cause of death [33]. This requirement becomes problematic when the suspect trees are located in remote or otherwise inaccessible areas or intended samples are in impossible to reach locations on a tree. In such cases, many months may pass between the identification of the suspect tree and the actual collection of a physical sample, and in this period the wood may dry out to the point of no longer producing viable materials for laboratory testing. In the most extreme cases, the suspect tree is never visited for sampling because its location is too dangerous or impossible to access [34].
Physical samples for laboratory analysis are typically collected by field crews using a hatchet and/or electric drill at breast height to remove woody material from a depth of 2–5 cm beneath the outer bark (Figure 2). A systemic pathogen, C. lukuohia spreads passively via conidia (spores) though xylem sap flow, often into the upper canopy. However, due to the differences in branch and xylem architecture of individual trees, there is uncertainty and an inconsistent degree of colonization into the uppermost and smallest branches and twigs [28]. This uncertainty is heightened by the spatial variability of the initial fungal infection point(s) on the tree, which can impact fungal distribution patterns. Infections and xylem occlusion of the lower bole could potentially lead to foliar symptoms in upper canopy branches without fungal colonization. Distal stem samples from emergent canopy trees are also subject to increased solar radiation that causes desiccation in outer branches [35]. Once the branch material becomes too dry, it is no longer suitable for laboratory analyses. Hence, an aerial sampling device that can collect larger diameter branches is needed, increasing the chance of providing viable wood for testing.
2. Materials and Methods
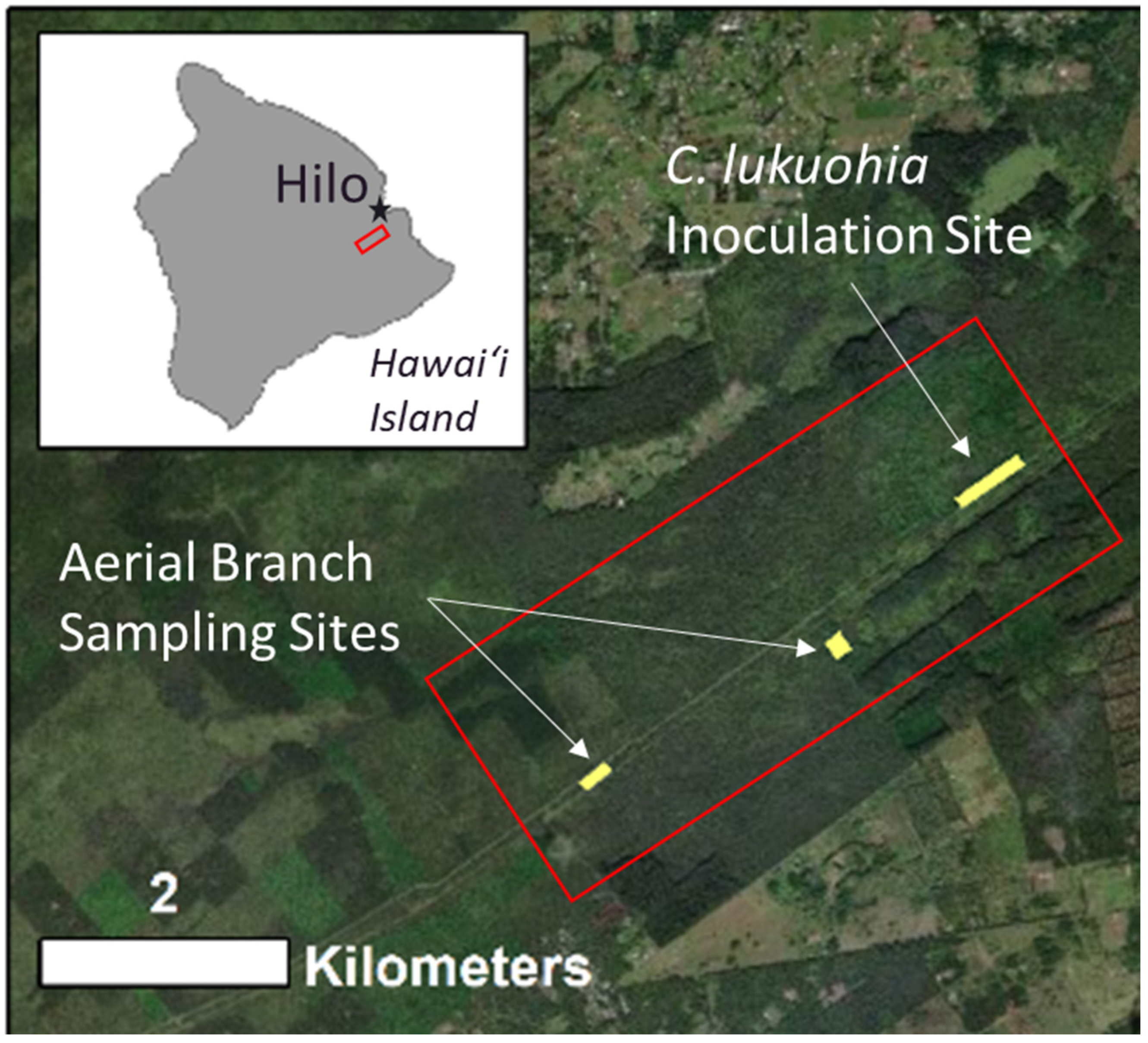
The field sites for this work are located on east Hawai‘i Island outside of the town of Hilo within the Waiākea Forest Reserve (Figure 3), an area heavily impacted by both C. lukuohia and invasive species including strawberry guava (Psidium cattleianum). The understory vegetation is dominated by a fern species, Uluhe (Dicranopteris linearis). Site elevations range from 300–500 m, increasing in elevation to the southwest. The area receives annual rainfall between 4500–5250 mm [36,37].
2.1. Determining the Minimum Branch Diameter for Detecting C. lukuohia
In order to determine the minimum branch diameter required to consistently generate a positive laboratory test for trees infected with C. lukuohia, an opportunistic branch sampling experiment was conducted using inoculated trees felled in Waiākea Forest Reserve on Hawai‘i Island (Figure 3) as part of a kiln-heating inoculation study (M. Hughes, in press). Branch samples from eleven C. lukuohia inoculated trees, ranging in diameter from 0.8–9.6 cm, were collected immediately after felling via hand saw and stored in Ziploc bags prior to delivery to the USDA Agricultural Research Service, Pacific Basic Agricultural Research Center (USDA ARS PBARC) in Hilo for testing via qPCR assay [33]. Sixty-three samples were collected from these branches and tested for the presence of C. lukuohia (Figure 4). Additional branch materials <1 cm in diameter were collected but these were determined to be too small or desiccated to be successfully tested via qPCR assay. All laboratory measurements were conducted at the USDA ARS facility in Hilo, HI, USA.
2.2. Aerial Branch Sampling with the FTTS
In an initial attempt to aerially sample ‘ōhi‘a trees for C. lukuohia, we used the FTTS (“Flying Tree Top Sampler”) device, developed by ETH Zurich [4]. We conducted six test flights with the FTTS sampler, attached to a DJI Matrice 600 sUAS platform via a 2 m extension pole (Figure 5) and were able to successfully collect small diameter (0.8–1.6 cm) branch material from each sampling flight. Sampling flights took place during daylight hours on 5 September 2019, and 18 October 2019. Sampled trees were chosen on the basis of accessibility and presence of the characteristic reddish canopy coloration suggesting likely infection with C. lukuohia. Some of the collected twig samples were too small and/or desiccated to extract enough material to run laboratory tests, but five FTTS-collected samples were eventually tested. The sampled trees were located within 50 m of the flight crew within direct line of sight, and first-person view camera footage (Figure 5B) was used to guide operation of the FTTS gripper and saw blade. We experienced challenges maneuvering the FTTS into place for sampling, as the dense canopy of the ‘ōhi‘a trees and shape of the FTTS sometimes made it difficult to position the gripper and the saw blade accurately for sample collection.
2.3. Development of the Kūkūau Branch Sampler
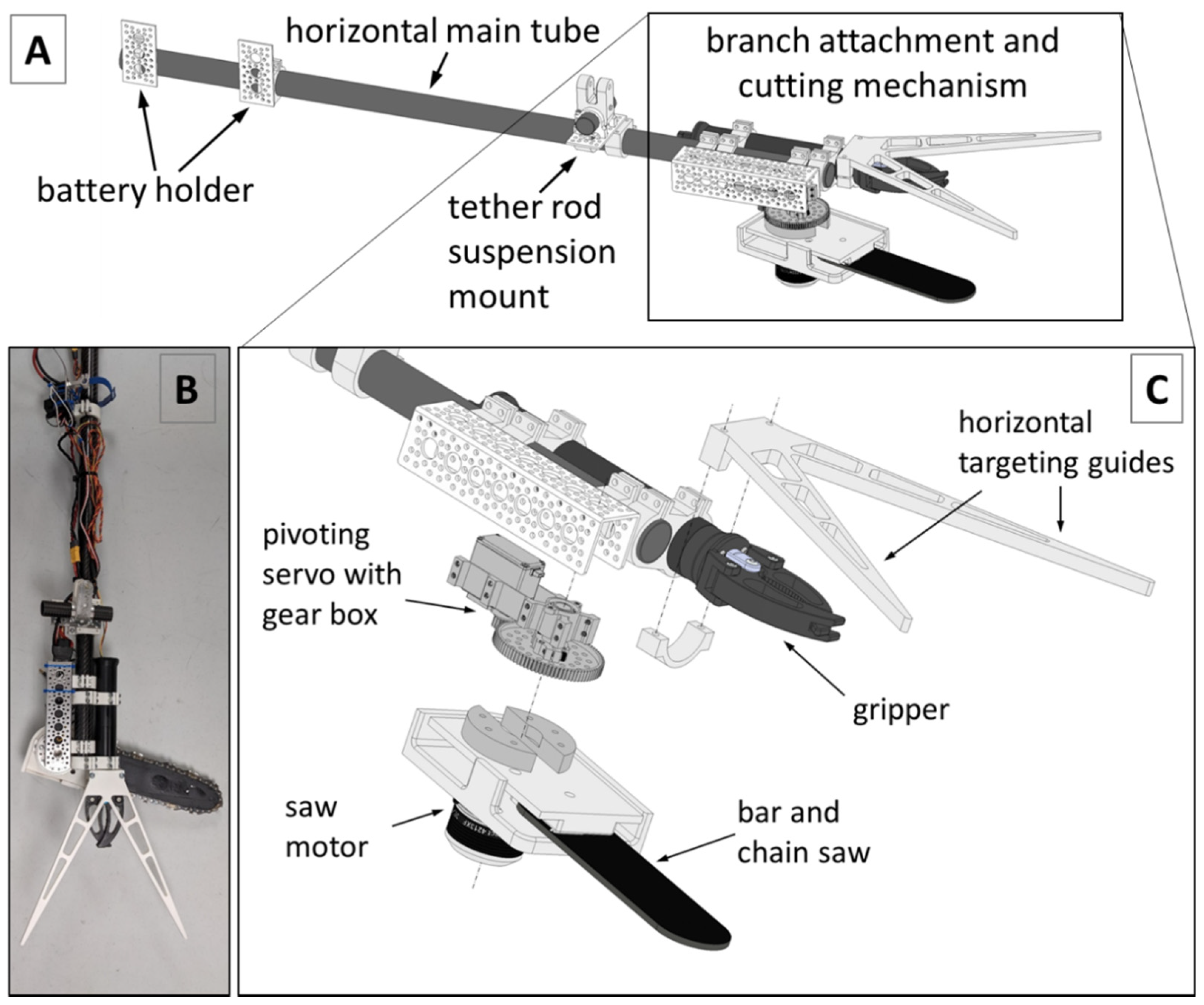
Based on our experiences with the FTTS in the field and the difficulties laboratory staff had in testing the collected small diameter twig samples, we determined that a new design was needed to better manuever within the canopy of the ‘ōhi‘a trees and collect larger diameter woody branch samples for diagnostic pathogen testing. Towards this end we developed and tested a series of prototypes, evolving from a small circular saw attached to a linear servo to the Kūkūau V2 branch sampler presented here, a robust bar and chain cutting system and robotic gripper claw mounted to a horizontal carbon fiber pole via a rotating servo gearbox (Figure 6).
The bar and chain of the Kūkūau V2 cutting mechanism originate from a commercially available oil-free pruning chainsaw (P5452, Ryobi Ltd., Hiroshima, Japan). As the chain does not require continuous lubrication, no reservoir of lubricating oil is needed. The chain is directly driven by a brushless motor (KDE4213XF-360, KDEdirect LLC, Bend, OR, USA) and electronic speed controller (ESC) (KDEXF-UAS55, KDEdirect LLC, Bend, OR, USA) and mounted to a servo driven gear box (Actobotics SG20-70, Robotzone LLC, Winfield, KS, USA) for horizontal pivoting. The mount was custom designed and 3D printed (RAISE3D, Pro2 Plus, Irvine, CA, USA) using poly-lactic acid (PLA) filament. The system attaches to target tree branches via a robotic gripper claw (Newton Gripper, Blue Robotics Torrance, CA, USA). The Kūkūau V2 branch sampler is powered by a 14.8 V lithium polymer battery (Turnigy Nano-Tech 4000 mAh 4S). A battery eliminating circuit (BEC) (Turnigy 6A (6 V~25 V) Switch Mode UBEC) steps down the voltage to 6.4 volts for the receiver and the pivoting servo. Saw motor speed, pivoting servo position and claw operation (open or close) are controlled via standard RC pulse width modulated (PWM) signals from the receiver. All these parts are mounted to a horizontally balanced carbon fiber tube (OD 28 mm) and attached to the sUAS platform via a vertical carbon fiber extension pole (3 m, OD 22 mm). A downward-looking oblique first person view (FPV) camera system (FPV Air Unit Module, DJI Inc., Shenzhen, China) is embedded into the 2 m vertical extension pole, which can be remotely detached from the sUAS platform by a servo in case of an emergency (not shown in Figure 6).
The bar and chain system would allow us to cut branches up to 15 cm in diameter, though this capacity is beyond the 7 cm maximum opening of the gripper claw currently used to secure the system to the tree during cutting and to hold the collected sample during retrieval. These components are manually controlled via a RC controller and connected to the sUAS platform by a 2 m extension pole and electronic quick-release mechanism. Our Kūkūau branch sampler was named to reflect its origins in Hawai‘i. Kūkūau is the name of an ahupua’a (land subdivision) in the Hilo area, and is also a term for a type of crab, Metopograpsus thukuhar, or ‘alamihi in Hawaiian [37]. The Kūkūau V2 system was developed by the University of Hawai‘i at Hilo Spatial Data Analysis & Visualization (SDAV) Laboratory in collaboration with ETH Zurich and R&R Machining/Welding in Hilo, Hawai‘i, USA. Kūkūau sampling flights took place during daylight hours on 6 May 2020, 13 May 2020, and 28 October 2020.
3. Results
3.1. Branch Diameter and C. lukuohia Detections
Summary results from the kiln inoculation felled-tree branch study, to determine the minimum branch diameter needed to confidently interpret results from qPCR testing, are shown in Table 1 (see Appendix A for the full dataset of tested branches from the felled tree inoculation trials). Small diameter branches from inoculated trees (1.0–2.5 cm) returned positive test results 43% of the time. As branch diameter increased, the positivity rate also increased, reaching 65% for branches between 2.5 and 5.0 cm, and 77% for branches 5.0 to 10.0 cm in diameter. For the smallest branches sampled, those ≤1 cm in diameter, the positivity rate was 0%. Each inoculated source tree that provided branch samples for the study was also sampled via the standard electric drill method to remove woody material from the main stem at breast height, and those samples all tested positive for C. lukuohia.
3.2. Aerial Branch Sampling Results
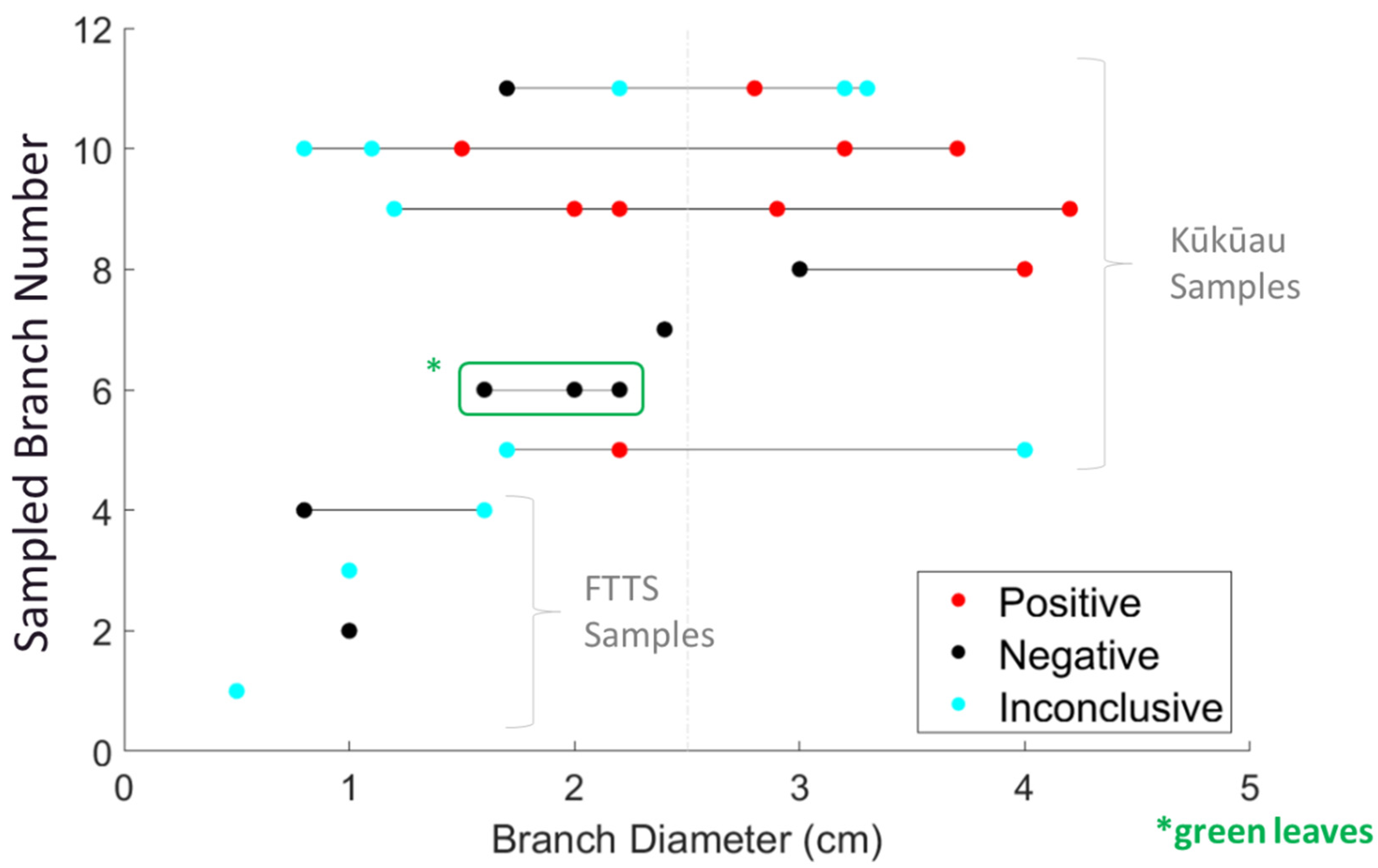
In addition to the FTTS flights, we conducted five sampling flights with the Kūkūau attachment (Figure 7), retrieving 11 different branches in total from ten trees exhibiting characteristic C. lukuohia symptoms. Branches were partitioned into 29 individual samples for laboratory testing. The maximum FTTS stem sample diameter was 1.6 cm and the maximum Kūkūau sample diameter was 4.2 cm. None of the five FTTS samples came back positive for C. lukuohia, while five of seven branches (10 of 24 samples) collected by the Kūkūau attachment were positive (Figure 8). Eleven samples, out of the overall 29 collected, returned inconclusive results, indicating a failed qPCR reaction [33]. There was a marked increase in the positivity rate for the higher diameter branches, with 0% testing positive for branches ≤1 cm, 25% testing positive for diameters of 1.0–2.5 cm, and 60% testing positive for branch diameters >2.5 cm (Table 1). One of the branches (number 6) contained healthy green leaves, though it was collected from a tree exhibiting symptoms on another branch that did test positive for C. lukuohia (Figure 7C).
4. Discussion
Our results show that sUAS aerial branch sampling is a viable technique for detecting vascular tree pathogens, with branch diameter and condition being important factors in obtaining informative qPCR results for the pathogen C. lukuohia. Branch and twig sampling for the detection of tree pathogens (though non-aerial) has been used in previous studies, including olive branch dieback [38], Dutch elm disease [39], and thousand cankers disease [40]. We found that larger diameter branches had a better chance of returning positive C. lukuohia results for aerially sampled, naturally infected trees, rising from a positivity rate of 0% for branches < 1 cm up to 60% for branches 2.5–5 cm in diameter. A similar pattern was found for the inoculated felled tree branch dataset, rising from 0% for branches <1 cm up to 77% for branches >5 cm in diameter. We acknowledge that these results are based on a relatively small dataset (n = 92, 29 of these from aerial operations) and support additional sampling and testing to further examine this relationship.
The current configuration of the Kūkūau branch sampler allows retrieval of a branch up to 7 cm in diameter, though the maximum diameter aerially sampled branch included in this study was 4.2 cm. As branch diameter increases, so does the weight of the resulting sample, eventually surpassing the carrying capacity of the sUAS and potentially inducing failure. One means of circumventing this issue would be to perform multiple cuts on a branch, discarding the smaller diameter outer branches and only retrieving a short section of thicker diameter material for testing. As sawdust and wood chips from cutting into infected trees may contain infectious material [41], a vacuum system for the collection of produced sawdust and wood chips to minimize spread of inoculum may be added in the future, though this would increase the weight and complexity of the system.
Branch diameter is not the only factor for successful aerial sampling for the detection of vascular tree pathogens. Within our dataset, two branches (number five and eleven) returned inconclusive results at the largest diameters but positive results at smaller diameter samples (Figure 8). Inconclusive results indicate a failed qPCR reaction and not a lack of pathogen detection [33]. Desiccated and degraded samples often result in inconclusive results, likely due to an accumulation of PCR inhibitors. Xylem vessel density and vessel diameter, both also related to xylem cavitation vulnerability, are influenced by branch architecture and species identity, with tension wood and available sunlight two factors that can affect branch xylem characteristics and therefore collected samples [39,42,43,44]. To combat these negative factors in high value samples and increase the chances of a positive result, it would be advised to remove the bark from stem samples and target symptomatic regions (showing xylem stain). Additionally, testing multiple samples from different sections of any given branch sample and screening the extracted DNA at various dilutions could improve capturing fungus-infested tissue while avoiding PCR inhibition. Of the two Kūkūau-collected branches that tested negative for C. lukuohia, one (branch six) contained only healthy green leaves from a tree exhibiting partial canopy symptoms (Figure 7C; highlighted green box in Figure 8). The other one (branch seven) was an older decomposed and desiccated branch that was difficult to process for laboratory testing.
To increase the likelihood of successful detection of a vascular fungal pathogen such as C. lukuohia from aerial branch sampling, we suggest the following in terms of candidate branch characteristics. The minimum branch diameter should be >2.5 cm (the thicker the better), located near the main stem, and support a full set of symptomatic leaves (reddish coloration). Branches that have lost their leaves may be too desiccated to produce viable sample material, while branches with asymptomatic leaves are unlikely to return positive test results, based on the single green leaf branch included in this study. The negative results for the green leaf branch also suggest that aerial branch sampling may not be a reliable means of confirming suspicions of early detection of infection by C. lukuohia via imaging spectroscopy [45] or other techniques, though more data are needed.
Even for the largest diameter branches sampled in this study, those between 5–10 cm from the felled tree inoculation trial, positivity rates were only 77%. To maximize the likelihood of aerially obtaining a positive test result from an infected tree, the best approach may be to mimic existing ground-based sampling protocols by sampling the main stem. This would require a system that can reliably attach to (and detach from) the main trunk of a tree and drill in and extract wood material from various points around the circumference. Aerially accessing the trunks of isolated trees is more feasible than it is for trees in closed canopy forests, but recent developments in autonomous navigation through forested environments and target selection are making this more practical [46,47,48].
5. Conclusions
In this study, we explored the utility of aerial branch sampling for the detection of C. lukuohia, the fungal pathogen responsible for widespread ‘ōhi‘a mortality across the state of Hawai‘i. We found that branch diameter and health play important roles in successful pathogen detection, with larger branches supporting symptomatic leaves most likely to test positive for C. lukuohia. Inconclusive and/or non-detection laboratory results were most frequently obtained for samples collected from branches that were small diameter and/or desiccated, or from branches supporting asymptomatic leaves. We have shown that aerial sUAS branch sampling is a viable method for fungal pathogen detection for high value inaccessible target trees, though additional improvements to reduce the fraction of inconclusive results and to minimize the release of inoculum during cutting are needed. As fungal pathogens affect forests worldwide [49,50], the results presented here have applications beyond the state of Hawai’i. In addition to aerial sampling for pathogen detection, the Kūkūau V2 branch sampler has utility in other conservation applications, including the collection and propagation of threatened and endangered species [7,8,9], canopy nutrient analyses [10,11,12], the mechanical control of incipient populations in inaccessible areas [51,52], and aerial pruning [53].
Author Contributions
Conceptualization, R.L.P., P.M., M.A.H., T.S. and T.B.; Data curation, E.B.; Formal analysis, E.B.; Funding acquisition, R.L.P.; Investigation, R.L.P., E.C. and E.B.; Methodology, R.L.P., P.M., E.C., T.S. and T.B.; Resources, N.B. and L.M.K.; Visualization, R.L.P.; Writing—original draft, R.L.P., P.M., M.A.H. and E.B.; Writing—review and editing, R.L.P., P.M., E.C., M.A.H., E.B., T.S., T.B., N.B. and L.M.K. All authors have read and agreed to the published version of the manuscript.
Funding
Funding for this project (via the The ‘Ōhi’a Challenge Prize) provided by the U.S. Department of the Interior’s Office of Native Hawaiian Relations, the National Park Service, and Conservation X Labs. P.M. was supported by ETH Zurich during a staff sabbatical.
Data Availability Statement
Not applicable.
Acknowledgments
We acknowledge the inspiration and support of Werner Eugster, ETH Zurich, who passed away before seeing this work. We also acknowledge and thank Cheyenne Perry for naming the Kūkūau branch sampling system.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
Tested Branch Sample Results from Felled Tree Inoculation Trial.
| Tree Sample | Diameter (cm) | CODE | Results |
|---|---|---|---|
| 211 | 1.70 | 1 | positive for C. lukuohia |
| 211 | 1.90 | 1 | positive for C. lukuohia |
| 211 | 2.50 | 0 | inconclusive |
| 211 | 2.80 | 1 | positive for C. lukuohia |
| 211 | 5.10 | 1 | positive for C. lukuohia |
| 211 | 5.10 | 1 | positive for C. lukuohia |
| 212 | 2.00 | 1 | positive for C. lukuohia |
| 212 | 2.30 | 0 | inconclusive |
| 212 | 3.40 | 0 | inconclusive |
| 212 | 4.40 | 1 | positive for C. lukuohia |
| 212 | 7.40 | 1 | positive for C. lukuohia |
| 214 | 1.00 | 0 | inconclusive |
| 214 | 1.60 | 1 | positive for C. lukuohia |
| 214 | 2.20 | 0 | inconclusive |
| 214 | 2.70 | 1 | positive for C. lukuohia |
| 214 | 4.40 | 0 | inconclusive |
| 214 | 5.50 | 1 | positive for C. lukuohia |
| 214 | 6.70 | 1 | positive for C. lukuohia |
| 215 | 0.8 | −1 | Ceratocystis not detected |
| 215 | 1.4 | 1 | Positive for C. lukuohia |
| 215 | 1.6 | 1 | Positive for C. lukuohia |
| 215 | 2.5 | 1 | Positive for C. lukuohia |
| 215 | 3.5 | 1 | Positive for C. lukuohia |
| 215 | 4.5 | 1 | Positive for C. lukuohia |
| 215 | 2.2 | 0 | inconclusive |
| 215 | 2.5 | 0 | inconclusive |
| 215 | 2.8 | 0 | inconclusive |
| 215 | 3.6 | 0 | inconclusive |
| 215 | 5.1 | 0 | inconclusive |
| 216 | 5.60 | −1 | no Ceratocystis detected |
| 216 | 6.00 | −1 | no Ceratocystis detected |
| 217 | 7.20 | −1 | no Ceratocystis detected |
| 217 | 9.60 | 1 | positive for C. lukuohia |
| 218 | 4.30 | −1 | no Ceratocystis detected |
| 218 | 5.60 | −1 | no Ceratocystis detected |
| 219 | 4.20 | −1 | no Ceratocystis detected |
| 219 | 7.40 | −1 | no Ceratocystis detected |
| 220 | 1.40 | 1 | Positive for C. lukuohia |
| 220 | 1.70 | −1 | no Ceratocystis detected |
| 220 | 2.60 | 1 | Positive for C. lukuohia |
| 220 | 3.40 | 0 | inconclusive |
| 220 | 5.20 | 1 | positive for C. lukuohia |
| 220 | 8.40 | 1 | positive for C. lukuohia |
| Tree 224 | 1.2 | 0 | inconclusive |
| Tree 224 | 1.5 | 0 | inconclusive |
| Tree 224 | 1.9 | 0 | inconclusive |
| Tree 224 | 2.2 | −1 | no Ceratocystis detected |
| Tree 224 | 3 | −1 | no Ceratocystis detected |
| Tree 224 | 3.4 | 1 | C. lukuohia detected |
| Tree 224 | 0.8 | −1 | no Ceratocystis detected |
| Tree 224 | 1.3 | −1 | no Ceratocystis detected |
| Tree 224 | 2.1 | −1 | no Ceratocystis detected |
| Tree 224 | 2.4 | −1 | no Ceratocystis detected |
| Tree 224 | 2.5 | −1 | no Ceratocystis detected |
| Tree 224 | 2.6 | −1 | no Ceratocystis detected |
| Tree 224 | 3 | −1 | no Ceratocystis detected |
| Tree 224 | 4.8 | 1 | C. lukuohia detected |
| Tree 225 | 1.40 | 0 | inconclusive |
| Tree 225 | 1.80 | −1 | No Ceratocystis detected |
| Tree 225 | 2.20 | 0 | inconclusive |
| Tree 225 | 2.30 | −1 | No Ceratocystis detected |
| Tree 225 | 3.00 | −1 | No Ceratocystis detected |
| Tree 225 | 3.90 | 1 | Positive for C. lukuohia |
| Tree 225 | 4.20 | −1 | No Ceratocystis detected |
| Tree 226 | 1.30 | 1 | Positive for C. lukuohia (weak positive) |
| Tree 226 | 1.60 | 0 | inconclusive |
| Tree 226 | 2.00 | 1 | Positive for C. lukuohia (ohia internal marker was not detected in sample) |
| Tree 226 | 2.50 | 1 | Positive for C. lukuohia |
| Tree 226 | 2.70 | 1 | Positive for C. lukuohia |
| Tree 226 | 3.60 | 1 | Positive for C. lukuohia |
| Tree 226 | 5.00 | 1 | Positive for C. lukuohia |
| Tree 226_Rot_Core | 5.10 | 1 | Positive for C. lukuohia |
| Tree 226 | 7.40 | 0 | inconclusive |
| 228 | 1.50 | −1 | No Ceratocystis detected |
| 228 | 2.20 | 1 | positive for C. lukuohia |
| 228 | 2.40 | −1 | No Ceratocystis detected |
| 228 | 4.30 | 1 | positive for C. lukuohia |
| 228 | 4.30 | 1 | positive for C. lukuohia |
| 228 | 6.30 | 1 | positive for C. lukuohia |
| 228 | 6.30 | 1 | positive for C. lukuohia |
References
- Baena, S.; Boyd, D.S.; Moat, J. UAVs in Pursuit of Plant Conservation—Real World Experiences. Ecol. Inform. 2018, 47, 2–9. [Google Scholar] [CrossRef]
- Guimarães, N.; Pádua, L.; Marques, P.; Silva, N.; Peres, E.; Sousa, J.J. Forestry Remote Sensing from Unmanned Aerial Vehicles: A Review Focusing on the Data, Processing and Potentialities. Remote Sens. 2020, 12, 1046. [Google Scholar] [CrossRef]
- Sharma, J.B. Applications of Small Unmanned Aircraft Systems: Best Practices and Case Studies; CRC Press: Boca Raton, FL, USA, 2019; ISBN 978-0-429-52085-3. [Google Scholar]
- Käslin, F.; Baur, T.; Meier, P.; Koller, P.; Buchmann, N.; D’Odorico, P.; Eugster, W. Novel Twig Sampling Method by Unmanned Aerial Vehicle (UAV). Front. For. Glob. Change 2018, 1, 2. [Google Scholar] [CrossRef]
- Lucas, J.A. Plant Pathology and Plant Pathogens; John Wiley & Sons: New York, NY, USA, 2020; ISBN 978-1-118-89386-9. [Google Scholar]
- West, J.S.; Canning, G.G.; Perryman, S.A.; King, K. Novel Technologies for the Detection of Fusarium Head Blight Disease and Airborne Inoculum. Trop. Plant Pathol. 2017, 42, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Szczecińska, M.; Sramko, G.; Wołosz, K.; Sawicki, J. Genetic Diversity and Population Structure of the Rare and Endangered Plant Species Pulsatilla Patens (L.) Mill in East Central Europe. PLoS ONE 2016, 11, e0151730. [Google Scholar] [CrossRef]
- Luna, T. Native Plant Restoration on Hawai‘i. Native Plants J. 2018, 19, 58–69. [Google Scholar] [CrossRef]
- Weisenberger, L.A.; Weller, S.G.; Sakai, A.K. Remnants of Populations Provide Effective Source Material for Reintroduction of an Endangered Hawaiian Plant, Schiedea Kaalae (Caryophyllaceae). Am. J. Bot. 2014, 101, 1954–1962. [Google Scholar] [CrossRef] [PubMed]
- Reuter, D.; Robinson, J.B. Plant Analysis: An Interpretation Manual; Csiro Publishing: Clayton, Australia, 1997; ISBN 978-0-643-10126-5. [Google Scholar]
- Asner, G.P.; Martin, R.E.; Tupayachi, R.; Anderson, C.B.; Sinca, F.; Carranza-Jiménez, L.; Martinez, P. Amazonian Functional Diversity from Forest Canopy Chemical Assembly. Proc. Natl. Acad. Sci. USA 2014, 111, 5604–5609. [Google Scholar] [CrossRef] [PubMed]
- Lira-Martins, D.; Humphreys-Williams, E.; Strekopytov, S.; Ishida, F.Y.; Quesada, C.A.; Lloyd, J. Tropical Tree Branch-Leaf Nutrient Scaling Relationships Vary With Sampling Location. Front. Plant Sci. 2019, 10, 877. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Serbin, S.P.; McNeil, B.E.; Kingdon, C.C.; Townsend, P.A. Imaging Spectroscopy Algorithms for Mapping Canopy Foliar Chemical and Morphological Traits and Their Uncertainties. Ecol. Appl. 2015, 25, 2180–2197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Wang, T.; Darvishzadeh, R.; Skidmore, A.K.; Jones, S.; Suarez, L.; Woodgate, W.; Heiden, U.; Heurich, M.; Hearne, J. Vegetation Indices for Mapping Canopy Foliar Nitrogen in a Mixed Temperate Forest. Remote Sens. 2016, 8, 491. [Google Scholar] [CrossRef]
- Nakamura, A.; Kitching, R.L.; Cao, M.; Creedy, T.J.; Fayle, T.M.; Freiberg, M.; Hewitt, C.N.; Itioka, T.; Koh, L.P.; Ma, K.; et al. Forests and Their Canopies: Achievements and Horizons in Canopy Science. Trends Ecol. Evol. 2017, 32, 438–451. [Google Scholar] [CrossRef]
- Malenovský, Z.; Homolová, L.; Lukeš, P.; Buddenbaum, H.; Verrelst, J.; Alonso, L.; Schaepman, M.E.; Lauret, N.; Gastellu-Etchegorry, J.-P. Variability and Uncertainty Challenges in Scaling Imaging Spectroscopy Retrievals and Validations from Leaves Up to Vegetation Canopies. Surv. Geophys. 2019, 40, 631–656. [Google Scholar] [CrossRef]
- Cannon, C.H.; Borchetta, C.; Anderson, D.L.; Arellano, G.; Barker, M.; Charron, G.; LaMontagne, J.M.; Richards, J.H.; Abercrombie, E.; Banin, L.F.; et al. Extending Our Scientific Reach in Arboreal Ecosystems for Research and Management. Front. For. Glob. Change 2021, 4, 160. [Google Scholar] [CrossRef]
- Jackaman, W.; Sacco, D.A. Reconnaissance Biogeochemical Survey Using Spruce Tops in the West Road (Blackwater) River Area, Fraser Plateau, Central British Columbia (Parts of NTS 093C/14, /15, 093F/02, /03). 2015, 4. Available online: https://cdn.geosciencebc.com/pdf/SummaryofActivities2015/SoA2015_Jackaman.pdf (accessed on 7 August 2022).
- Hildes. Turanich-Noyen Aerochem: An Introduction and Comparison with Traditional Stream Sediment Sampling. Available online: https://www.semanticscholar.org/paper/Aerochem-%3A-An-Introduction-and-Comparison-with-Hildes-Turanich-Noyen/1b36eccf3d2cbc3bb4dac4c5544740e10809e467 (accessed on 11 December 2020).
- UC Berkeley Forest Pathology and Mycology Lab Sampler Drones for Forestry Reseach. Available online: https://nature.berkeley.edu/garbelottowp/?p=1801 (accessed on 14 December 2020).
- Kutia, J. Aerial Manipulation for Canopy Sampling. Ph.D. Thesis, University of Auckland, Auckland, New Zealand, 2019. [Google Scholar]
- Charron, G.; Robichaud-Courteau, T.; Vigne, H.L.; Weintraub, S.; Hill, A.; Justice, D.; Bélanger, N.; Desbiens, A.L. The DeLeaves: A UAV Device for Efficient Tree Canopy Sampling. J. Unmanned Veh. Syst. 2020, 8, 245–264. [Google Scholar] [CrossRef]
- Krisanski, S.; Taskhiri, M.S.; Montgomery, J.; Turner, P. Design and Testing of a Novel Unoccupied Aircraft System for the Collection of Forest Canopy Samples. Forests 2022, 13, 153. [Google Scholar] [CrossRef]
- Schweiger, A.K.; Desbiens, A.L.; Charron, G.; Vigne, H.L.; Laliberté, E. Foliar Sampling with an Unmanned Aerial System (UAS) Reveals Spectral and Functional Trait Differences within Tree Crowns. Can. J. For. Res. 2020, 50, 966–974. [Google Scholar] [CrossRef]
- Barnes, I.; Fourie, A.; Wingfield, M.J.; Harrington, T.C.; McNew, D.L.; Sugiyama, L.S.; Luiz, B.C.; Heller, W.P.; Keith, L.M. New Ceratocystis Species Associated with Rapid Death of Metrosideros Polymorpha in Hawai’i. Persoonia Mol. Phylogeny Evol. Fungi 2018, 40, 154–181. [Google Scholar] [CrossRef]
- Camp, R.J.; LaPointe, D.A.; Hart, P.J.; Sedgwick, D.E.; Canale, L.K. Large-Scale Tree Mortality from Rapid Ohia Death Negatively Influences Avifauna in Lower Puna, Hawaii Island, USA. Condor 2019, 121, duz007. [Google Scholar] [CrossRef]
- Fortini, L.B.; Kaiser, L.R.; Keith, L.M.; Price, J.; Hughes, R.F.; Jacobi, J.D.; Friday, J.B. The Evolving Threat of Rapid ‘Ōhi‘a Death (ROD) to Hawai‘i’s Native Ecosystems and Rare Plant Species. For. Ecol. Manag. 2019, 448, 376–385. [Google Scholar] [CrossRef]
- Hughes, M.A.; Juzwik, J.; Harrington, T.C.; Keith, L.M. Pathogenicity, Symptom Development, and Colonization of Metrosideros Polymorpha by Ceratocystis Lukuohia. Plant Dis. 2020, 104, 2233–2241. [Google Scholar] [CrossRef] [PubMed]
- Keith, L.M.; Hughes, R.F.; Sugiyama, L.S.; Heller, W.P.; Bushe, B.C.; Friday, J.B. First Report of Ceratocystis Wilt on ˋŌhiˋa (Metrosideros Polymorpha). Plant Dis. 2015, 99, 1276. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E.; Keith, L.M.; Heller, W.P.; Hughes, M.A.; Vaughn, N.R.; Hughes, R.F.; Balzotti, C. A Spectral Mapping Signature for the Rapid Ohia Death (ROD) Pathogen in Hawaiian Forests. Remote Sens. 2018, 10, 404. [Google Scholar] [CrossRef]
- Vaughn, N.R.; Asner, G.P.; Brodrick, P.G.; Martin, R.E.; Heckler, J.W.; Knapp, D.E.; Hughes, R.F. An Approach for High-Resolution Mapping of Hawaiian Metrosideros Forest Mortality Using Laser-Guided Imaging Spectroscopy. Remote Sens. 2018, 10, 502. [Google Scholar] [CrossRef]
- Perroy, R.L.; Hughes, M.; Keith, L.M.; Collier, E.; Sullivan, T.; Low, G. Examining the Utility of Visible Near-Infrared and Optical Remote Sensing for the Early Detection of Rapid ‘Ōhi‘a Death. Remote Sens. 2020, 12, 1846. [Google Scholar] [CrossRef]
- Heller, W.P.; Keith, L.M. Real-Time PCR Assays to Detect and Distinguish the Rapid ʻŌhiʻa Death Pathogens Ceratocystis Lukuohia and C. Huliohia. Phytopathology 2018, 108, 1395–1401. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, M. (Hawaii Department of Land and Natural Resources, Honolulu, Hawaii, USA). Personal communication. 2020. [Google Scholar]
- Kunert, N.; Aparecido, L.M.T.; Wolff, S.; Higuchi, N.; dos Santos, J.; de Araujo, A.C.; Trumbore, S. A Revised Hydrological Model for the Central Amazon: The Importance of Emergent Canopy Trees in the Forest Water Budget. Agric. For. Meteorol. 2017, 239, 47–57. [Google Scholar] [CrossRef]
- Giambelluca, T.W.; Chen, Q.; Frazier, A.G.; Price, J.P.; Chen, Y.-L.; Chu, P.-S.; Eischeid, J.K.; Delparte, D.M. Online Rainfall Atlas of Hawai‘i. Bull. Am. Meteorol. Soc. 2013, 94, 313–316. [Google Scholar] [CrossRef]
- Frazier, A.G.; Giambelluca, T.W. Spatial Trend Analysis of Hawaiian Rainfall from 1920 to 2012-Frazier-2017-International Journal of Climatology—Wiley Online Library. Available online: https://0-rmets-onlinelibrary-wiley-com.brum.beds.ac.uk/doi/abs/10.1002/joc.4862 (accessed on 9 September 2022).
- Moral, J.; Agustí-Brisach, C.; Pérez-Rodríguez, M.; Xaviér, C.; Raya, M.C.; Rhouma, A.; Trapero, A. Identification of Fungal Species Associated with Branch Dieback of Olive and Resistance of Table Cultivars to Neofusicoccum Mediterraneum and Botryosphaeria Dothidea. Plant Dis. 2017, 101, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Beier, G.L.; Blanchette, R.A. Xylem Characteristics in Ulmus Americana Cultivars and Their Potential Use as a Preliminary Screening Method for Dutch Elm Disease Resistance. For. Pathol. 2020, 50, e12638. [Google Scholar] [CrossRef]
- Oren, E.; Klingeman, W.; Gazis, R.; Moulton, J.; Lambdin, P.; Coggeshall, M.; Hulcr, J.; Seybold, S.J.; Hadziabdic, D. A Novel Molecular Toolkit for Rapid Detection of the Pathogen and Primary Vector of Thousand Cankers Disease. PLoS ONE 2018, 13, e0185087. [Google Scholar] [CrossRef] [PubMed]
- Harrington, T.C. Ceratocystis Diseases. In Infectious Forest Diseases; Gonthier, P., Nicolotti, G., Eds.; CABI: Wallingford, UK, 2013; pp. 230–255. ISBN 978-1-78064-040-2. [Google Scholar]
- Lemoine, D.; Jacquemin, S.; Granier, A. Beech ( Fagus Sylvatica L.) Branches Show Acclimation of Xylem Anatomy Andhydraulic Properties to Increased Light after Thinning. Ann. For. Sci. 2002, 59, 761–766. [Google Scholar] [CrossRef]
- Jourez, B.; Riboux, A.; Leclercq, A. Anatomical Characteristics Of Tension Wood And Opposite Wood In Young Inclined Stems Of Poplar (Populus Euramericana Cv ’Ghoy’). IAWA J. 2001, 22, 133–157. [Google Scholar] [CrossRef]
- Yamamoto, F.; Kozlowski, T.T. Effects of Flooding, Tilting of Stems, and Ethrel Application on Growth, Stem Anatomy and Ethylene Production of Pinus Densiflora Seedlings. J. Exp. Bot. 1987, 38, 293–310. [Google Scholar] [CrossRef]
- Weingarten, E.; Martin, R.E.; Hughes, R.F.; Vaughn, N.R.; Shafron, E.; Asner, G.P. Early Detection of a Tree Pathogen Using Airborne Remote Sensing. Ecol. Appl. 2022, 32, e2519. [Google Scholar] [CrossRef]
- Lee, T.; Mckeever, S.; Courtney, J. Flying Free: A Research Overview of Deep Learning in Drone Navigation Autonomy. Drones 2021, 5, 52. [Google Scholar] [CrossRef]
- da Silva, D.Q.; dos Santos, F.N.; Sousa, A.J.; Filipe, V.; Boaventura-Cunha, J. Unimodal and Multimodal Perception for Forest Management: Review and Dataset. Computation 2021, 9, 127. [Google Scholar] [CrossRef]
- Salles, R.N.; de Campos Velho, H.F.; Shiguemori, E.H. Automatic Position Estimation Based on Lidar × Lidar Data for Autonomous Aerial Navigation in the Amazon Forest Region. Remote Sens. 2022, 14, 361. [Google Scholar] [CrossRef]
- Wingfield, M.J.; Slippers, B.; Wingfield, B.D.; Barnes, I. The Unified Framework for Biological Invasions: A Forest Fungal Pathogen Perspective. Biol. Invasions 2017, 19, 3201–3214. [Google Scholar] [CrossRef]
- Ghelardini, L.; Pepori, A.L.; Luchi, N.; Capretti, P.; Santini, A. Drivers of Emerging Fungal Diseases of Forest Trees. For. Ecol. Manag. 2016, 381, 235–246. [Google Scholar] [CrossRef]
- Howard, P.L. Human Adaptation to Invasive Species: A Conceptual Framework Based on a Case Study Metasynthesis. Ambio 2019, 48, 1401–1430. [Google Scholar] [CrossRef] [PubMed]
- Van Wilgen, B.; Richardson, D.; Higgins, S.I. Integrated Control of Invasive Alien Plants in Terrestrial Ecosystems. Land Use Water Resour. Res. 2001, 1, 1732-2016-140256. [Google Scholar] [CrossRef]
- Molina, J.; Hirai, S. Aerial Pruning Mechanism, Initial Real Environment Test. Robot. Biomim. 2017, 4, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
High-resolution aerial image from east Hawai‘i Island showing an ‘ōhi‘a tree with the characteristic reddish canopy coloration suggesting likely infection with C. lukuohia, the fungal pathogen responsible for Ceratocystis wilt of ‘ōhi‘a.
Figure 1.
High-resolution aerial image from east Hawai‘i Island showing an ‘ōhi‘a tree with the characteristic reddish canopy coloration suggesting likely infection with C. lukuohia, the fungal pathogen responsible for Ceratocystis wilt of ‘ōhi‘a.

Figure 2.
Ground-based sampling with a handheld drill for the collection of material for qPCR testing for C. lukuohia, the fungal pathogen responsible for Ceratocystis wilt of ‘ōhi‘a.
Figure 2.
Ground-based sampling with a handheld drill for the collection of material for qPCR testing for C. lukuohia, the fungal pathogen responsible for Ceratocystis wilt of ‘ōhi‘a.

Figure 3.
Location of field study sites on Hawai‘i Island for the aerial branch sampling described in this work.
Figure 3.
Location of field study sites on Hawai‘i Island for the aerial branch sampling described in this work.

Figure 4.
(Left) Samples from kiln-study branches to be tested; (Right) Aerial branch sample during laboratory processing.
Figure 4.
(Left) Samples from kiln-study branches to be tested; (Right) Aerial branch sample during laboratory processing.

Figure 5.
FTTS in process of collecting a twig sample. (A) View of the FTTS from the ground prior to sampling; (B) Screenshot from video recording from first-person view camera located on the extension pole looking downwards at the FTTS; (C) close-up view of the FTTS.
Figure 5.
FTTS in process of collecting a twig sample. (A) View of the FTTS from the ground prior to sampling; (B) Screenshot from video recording from first-person view camera located on the extension pole looking downwards at the FTTS; (C) close-up view of the FTTS.

Figure 6.
(A) Kūkūau V2 branch sampler components. (B) Photo of Kūkūau V2 branch sampler. (C) Expanded view of branch attachment and cutting mechanism.
Figure 6.
(A) Kūkūau V2 branch sampler components. (B) Photo of Kūkūau V2 branch sampler. (C) Expanded view of branch attachment and cutting mechanism.

Figure 7.
Aerial branch sampling. (A) Kūkūau branch sampler in the air following a successful cut; (B) Samples from a single branch, U.S. Quarter dollar (25 cents coin) for scale; (C) Two different collected branches, branch on the right contains healthy green leaves from a tree exhibiting partial C. lukuohia canopy symptoms; (D) >2 m tall branch collected by the Kūkūau branch sampler.
Figure 7.
Aerial branch sampling. (A) Kūkūau branch sampler in the air following a successful cut; (B) Samples from a single branch, U.S. Quarter dollar (25 cents coin) for scale; (C) Two different collected branches, branch on the right contains healthy green leaves from a tree exhibiting partial C. lukuohia canopy symptoms; (D) >2 m tall branch collected by the Kūkūau branch sampler.

Figure 8.
Laboratory test results for infection with C. lukuohia of aerially sampled branches.

Table 1.
Laboratory qPCR test results from collected branch samples.
| Felled Inoculated Tree Branches | Aerially Sampled Branches | |||||
|---|---|---|---|---|---|---|
| Branch Diameter (cm) | # of Samples | # Positive for C. lukuohia | % Positive for C. lukuohia | # of Samples | # Positive for C. lukuohia | % Positive for C. lukuohia |
| <1.0 | 2 | 0 | 0% | 3 | 0 | 0% |
| 1.0–2.5 | 28 | 12 | 43% | 16 | 4 | 25% |
| 2.51–5.0 | 20 | 13 | 65% | 10 | 6 | 60% |
| 5.1–10.0 | 13 | 10 | 77% | N/A | N/A | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Perroy, R.L.; Meier, P.; Collier, E.; Hughes, M.A.; Brill, E.; Sullivan, T.; Baur, T.; Buchmann, N.; Keith, L.M. Aerial Branch Sampling to Detect Forest Pathogens. Drones 2022, 6, 275. https://0-doi-org.brum.beds.ac.uk/10.3390/drones6100275
AMA Style
Perroy RL, Meier P, Collier E, Hughes MA, Brill E, Sullivan T, Baur T, Buchmann N, Keith LM. Aerial Branch Sampling to Detect Forest Pathogens. Drones. 2022; 6(10):275. https://0-doi-org.brum.beds.ac.uk/10.3390/drones6100275
Chicago/Turabian StylePerroy, Ryan L., Philip Meier, Eszter Collier, Marc A. Hughes, Eva Brill, Timo Sullivan, Thomas Baur, Nina Buchmann, and Lisa M. Keith. 2022. "Aerial Branch Sampling to Detect Forest Pathogens" Drones 6, no. 10: 275. https://0-doi-org.brum.beds.ac.uk/10.3390/drones6100275