Global Properties of Latent Virus Dynamics Models with Immune Impairment and Two Routes of Infection


1
Department of Mathematics, Faculty of Science, King Abdulaziz University, P.O. Box 80203, Jeddah 21589, Saudi Arabia
2
Department of Mathematics, Faculty of Science, King Khalid University, P.O. Box 25145, Abha 61466, Saudi Arabia
*
Author to whom correspondence should be addressed.
High-Throughput 2019, 8(2), 16; https://0-doi-org.brum.beds.ac.uk/10.3390/ht8020016
Submission received: 19 March 2019
/
Revised: 19 April 2019
/
Accepted: 15 May 2019
/
Published: 3 June 2019
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:This paper studies the global stability of viral infection models with CTL immune impairment. We incorporate both productively and latently infected cells. The models integrate two routes of transmission, cell-to-cell and virus-to-cell. In the second model, saturated virus–cell and cell–cell incidence rates are considered. The basic reproduction number is derived and two steady states are calculated. We first establish the nonnegativity and boundedness of the solutions of the system, then we investigate the global stability of the steady states. We utilize the Lyapunov method to prove the global stability of the two steady states. We support our theorems by numerical simulations.
1. Introduction
In the literature, several mathematical models of within-host virus dynamics have been constructed and analyzed [1,2,3,4,5,6,7,8,9]. The cytotoxic T Lymphocyte (CTL) is one of the central components of the immune system against viral infections. CTLs lyse the viral-infected cells which participate in reducing or clearing the viruses from the body. Several mathematical models have been presented which integrate the effect of the CTL immune response on viral dynamics (see e.g., [10,11,12]). Nowak and Bangham [10] have presented a mathematical model to characterize the dynamics of the virus (J) with uninfected cells (G), infected cells (I) and CTLs (K) as:
The uninfected cells are replenished at rate , die at rate and become infected at rate , where is the virus–cell incidence rate constant. is the killer rate of infected cells by CTL and is the death rate of the infected cells, where and are constants. The CTLs are proliferated and die at rates and , respectively, where and are constants.
Models (1)–(4) assume that the presence of antigen can activate the CTL immune response, however, the CTL immune impairment is negelcted. To model the immune impairment, Regoes et al. [13] have modified models (1)–(4) as:
where the terms and represents the proliferation rate and the immune impairment, respectively, and h is a constant. Mathematical models of virus dynamics with impairment of CTL functions have been constructed in seveal papers (see e.g., [13,14,15]). The works presented in [13,14,15] assume that the virus infects the uninfected cells by virus-to-cell transmission.
The uninfected target cells can be infected via two ways of transmissions, namely, the diffusion-limited virus-to-cell transmission and the direct cell-to-cell transfer using virological synapses [16]. The cell-to-cell transmission has been recognized in several works (see e.g., [17,18,19,20]). Recent studies have revealed that over 50% of viral infection is due to cell-to-cell transmission [21] and even with an antiretroviral therapy, the cell-to-cell spread of the virus can still permit ongoing replication [22]. Thus, for some viruses, cell-to-cell transmission seems to be a more powerful means of virus propagation than the virus-to-cell transmission [23,24]. Several mathematical models of virus dynamics with two ways of infection have been developed by many researchers (see [25,26,27,28,29,30]). However, in these papers, the impairment of CTL functions is not included. In a very recent work, Elaiw et al. [31] have studied the dynamic behavior of virus infection with impairment of CTL functions and two routes of infection, but with one class of infected cells, productively infected cells.
In case of human immunodeficiency virus (HIV) infection, current treatment consisting of several antiretroviral drugs can suppress viral replication to a low level but cannot completely eradicate the HIV [29]. An important reason is that HIV provirus can reside in latently infected cells [32,33]. Latently infected cells live long, are not affected by antiretroviral drugs or immune responses, but can be activated to produce HIV by relevant antigens.
The aim of the present paper is to propose and analyze viral infection models which include (i) both productively infected cells and latently infected cells, (ii) both virus-to-cell and cell-to-cell transmissions, and (iii) impairment of CTL functions. We first show that the solutions of the models are nonnegative and bounded, then we derive the basic reproduction number which determines the existence and global stability of the steady states. We utilize the Lyapunov method to prove the global stability of the two steady states. We support our theorems by numerical simulations.
2. The Model
We study the following model:
where, L is the concentration of the latently infected cells. The uninfected cells become infected at rates and due to virus-to-cell and cell-to-cell infections, respectively, where and are the incidence rate constants. The fractions and with are the probabilities that upon infection, an uninfected cell will becomes either latently infected or productively infected, respectively. Parameter b denotes the average number of latently infected cells cells that become productively infected cells, and d denotes the death rate constant of the latently infected cells.
2.1. Nonnegativity and Boundedness
Let us define
Proof.
We observe that
This confirms that with . Let Then
where, . Hence, for all if where . Consequently, and for all if , where and . This establishes the bondedness of and . □
Lemma2.
- (i)
- if then there exists a disease-free steady state ,
- (ii)
- if , then there exist two steady states and endemic steady state .
Proof.
The steady states of the system satisfy
By solving Equations (16)–(20) we get two steady states, disease-free steady state where . In addition, we have
where
Define a function by
Then, when and . Hence, there exists such that . Hence, when , then
It follows that, an endemic steady state exists if . □
2.2. Global Stability
We define . We note that for any and . To investigate the global stability of the steady states, we construct Lyapunov functions using the method presented [4] and followed by [5,6,7].
Proof.
Constructing a function as:
Clearly, for all , while reaches its global minimum at . Calculating along the trajectories of (9)–(13) we get
Since then for all we have . The solutions of the system tend to the largest invariant subset of [34]. It can be easily show that at . Applying LaSalle’s invariance principle (LIP), we get that is globally asymptotically stable.
We calculate the characteristic equation at the steady state as:
where
Define
We have . Hence, when . We have also which shows that has a positive real root and then, is unstable for . □
Proof.
Let a function be defined as:
We have if then . The geometrical and arithmetical means relationship implies that
Hence for all we have and when , , and . Utilizing LIP we obtain that if , then is globally asymptotically stable. □
3. Model with Saturated Incidence Rate
The rate of infection in model (9)–(13) is bilinear in the virus and the uninfected cell. Actual incidence rates are probably not strictly linear. A less than linear response in viruses and infected cells could occur due to saturation at high virus or infected cell concentrations [35]. Therefore, it is reasonable for us to assume that the infection rate of modeling viral infection is given by saturated mass action. In this section, we study a vial infection model with saturation:
where are saturation constants. All parameters and variables have the same meaning as (9)–(13).
3.1. Basic Properties
The next lemma shows the nonnegativity and boundedness of the solutions of system (27)–(31)
Lemma 3.
The compact set Ω is positively invariant for system (27)–(31).
The proof is similar to that of Lemma 1.
Lemma 4.
- (i)
- A disease-free steady state exists when
- (ii)
- An endemic steady state exists when .
3.2. Global Properties
Theorem 3.
Let , then of system (27)–(31) is globally asymptotically stable and it is unstable if .
Proof.
Define as the following
It is seen that for all while reaches its global minimum at . We calculate as:
Clearly if then for all we have , and when and . Applying LIP implies we get that if then is globally asymptotically stable. Similar to the previous section we can easily show that if , then is unstable. □
Theorem 4.
Let then Δ ofsystem (27)–(31) is globally asymptotically stable.
Proof.
Define a function as:
It is seen that for all while reaches its global minimum at . Calculating as:
The steady state conditions of implies that:
we get
The geometrical and arithmetical means relationship implies that
Thus for all and at . Using LIP one can easily show that is globally asymptotically stable. □
4. Numerical Simulations
In this section, we solve system (27)–(31) numerically with values of the parameters given as: , , , , , , , , and . The parameters , , and h will be varied. We take and choose different initial conditions as:
IC1: ,
IC2: ,
IC3:
IC4:
IC5:
Case(1) Stability of steady states:
We take and is varied as:
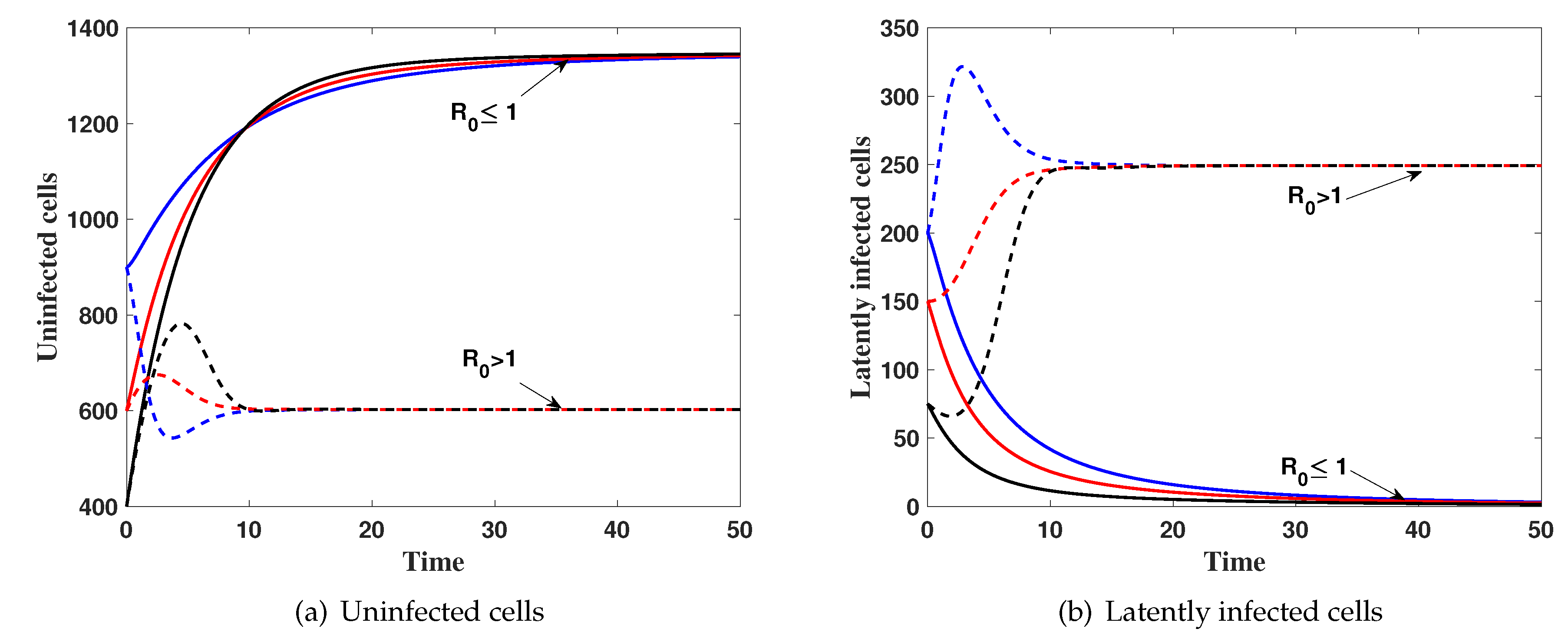
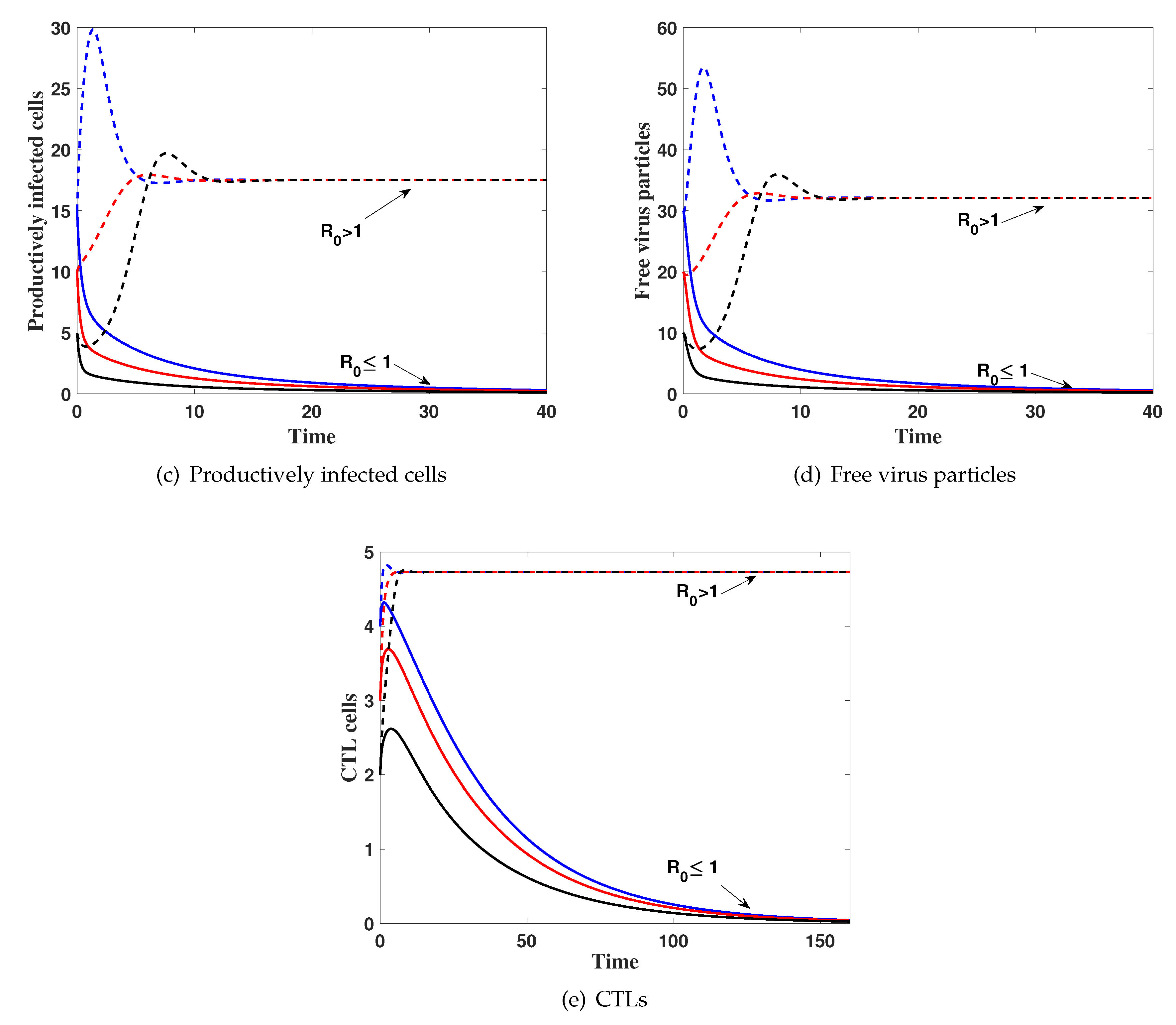
(i) then . Figure 1 shows that, the solution of the system with different initial conditions IC1–IC3 tends to . This result implies that is globally asymptotically stable which confirms Theorem 3.
(ii) then, . The numerical results show that the solutions of the system tends to the steady state for all IC1–IC3. This supports the global stability result of Theorem 4.
Case(2) Virus dynamics with variation of:
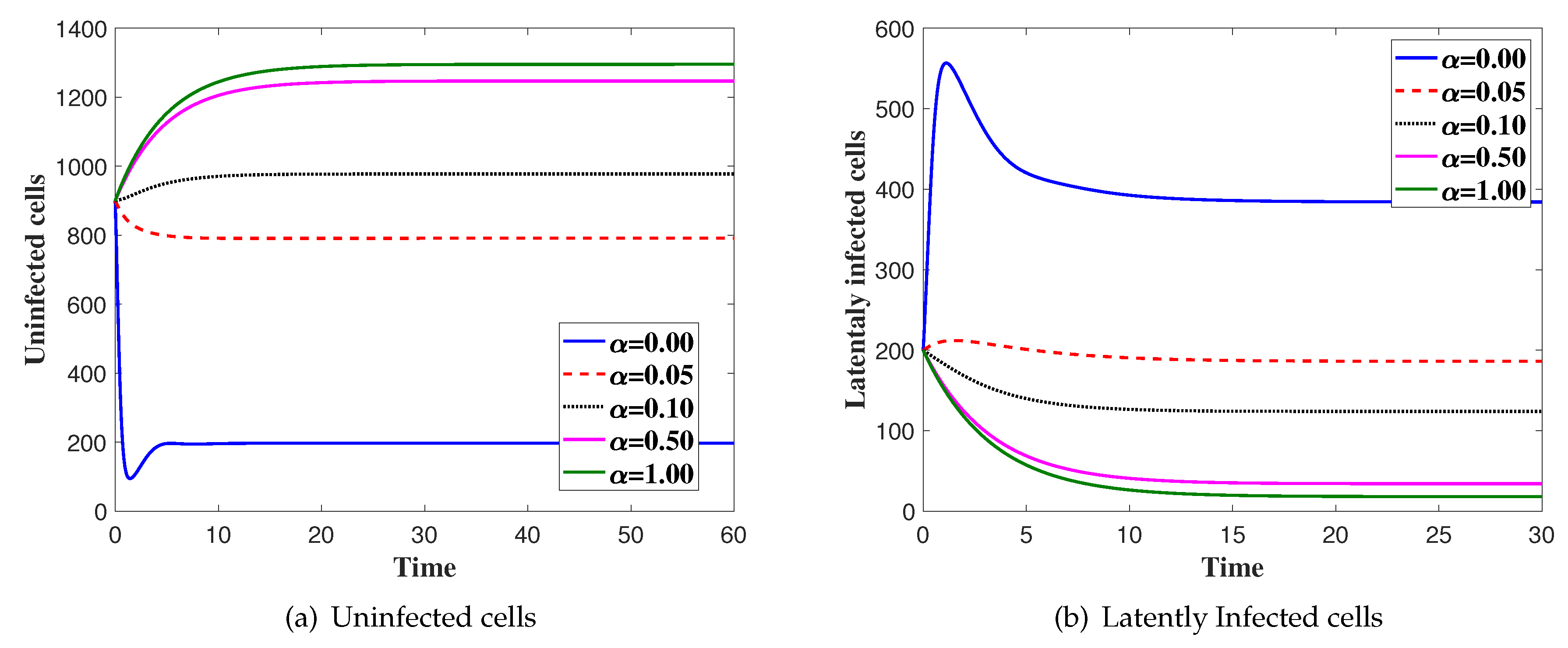
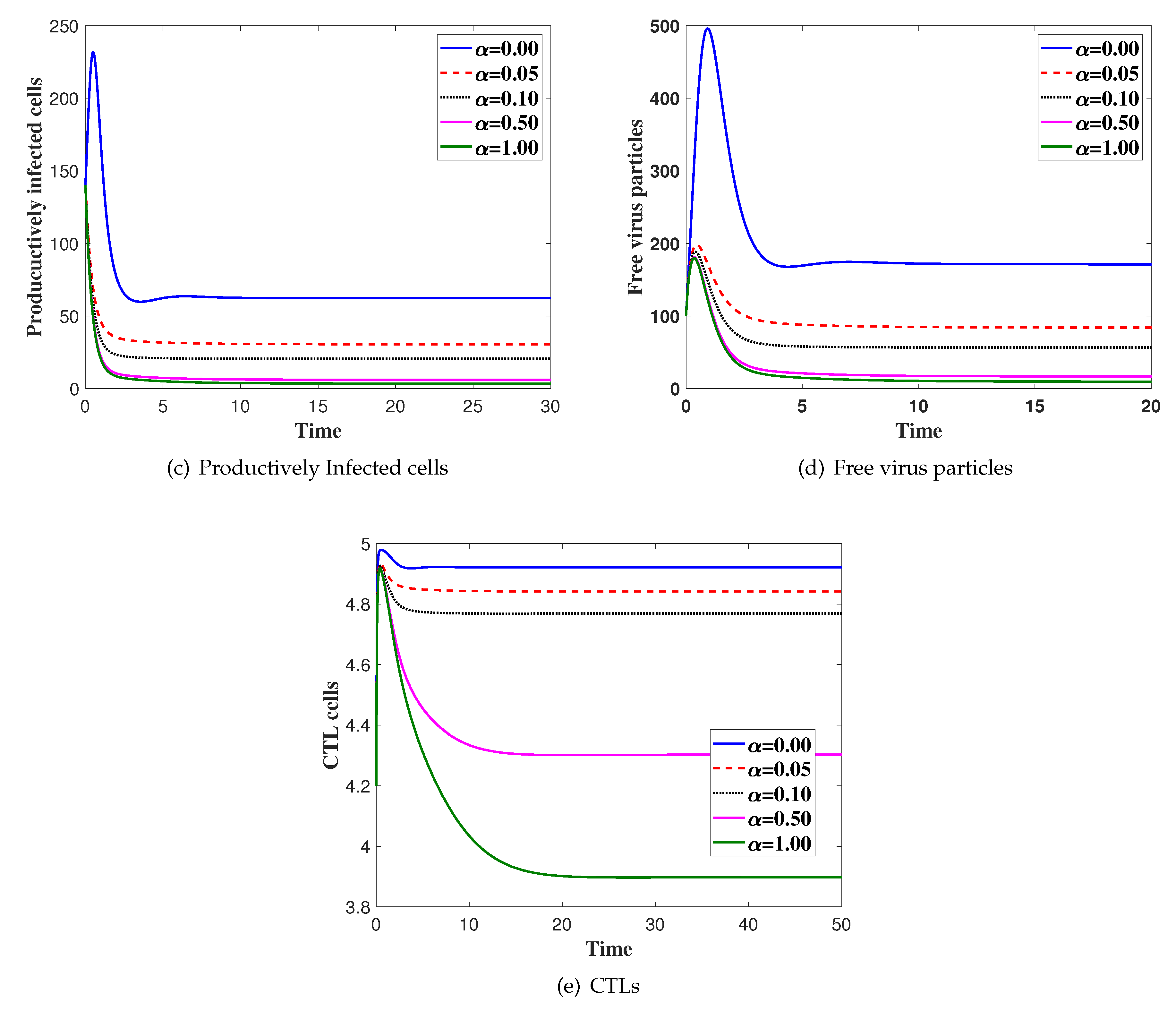
In this case, we fix and is changed. We solve the system numerically with the initial condition IC4. In Figure 2, we show the effect of saturated incidence parameter . We can see that the concentration of the uninfected cells is increased as is increased. Moreover, the concentration of latently infected cells, productively infected, viruses and CTLs are decreased as is increased.
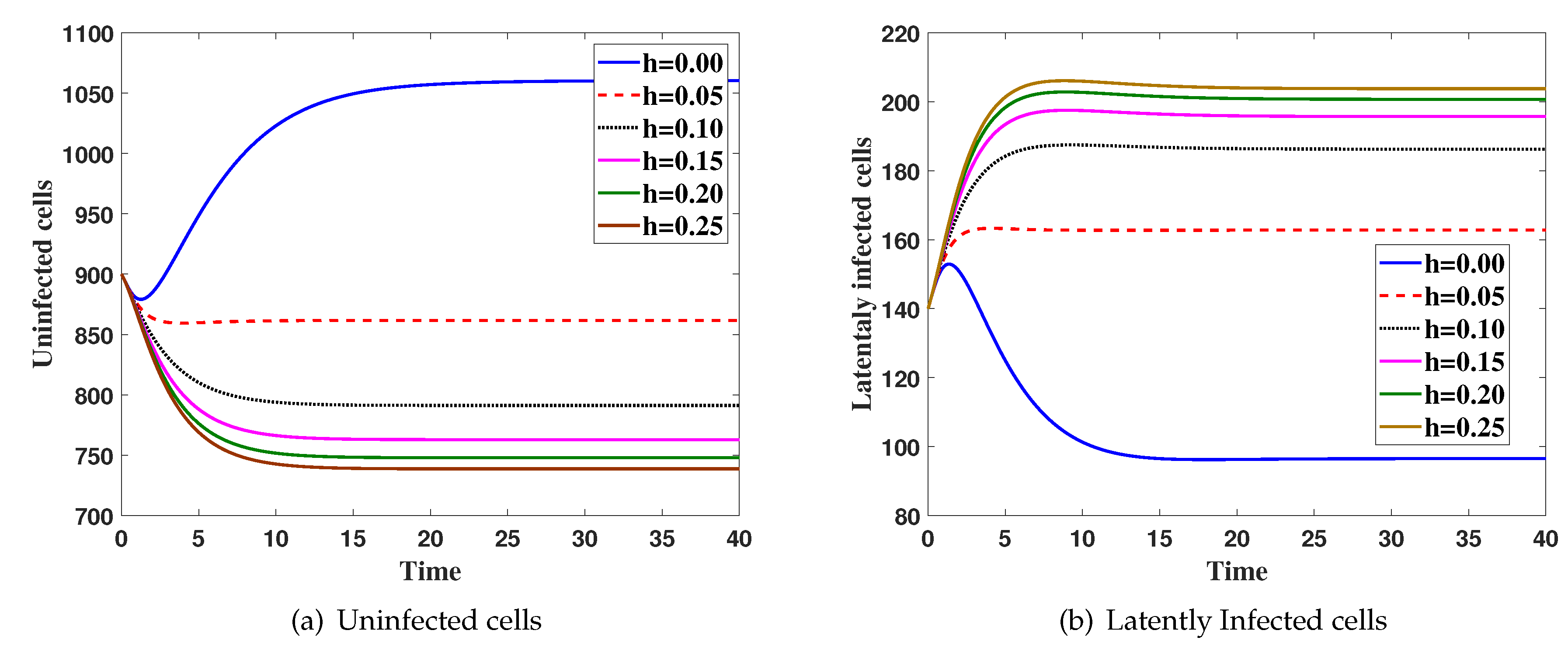
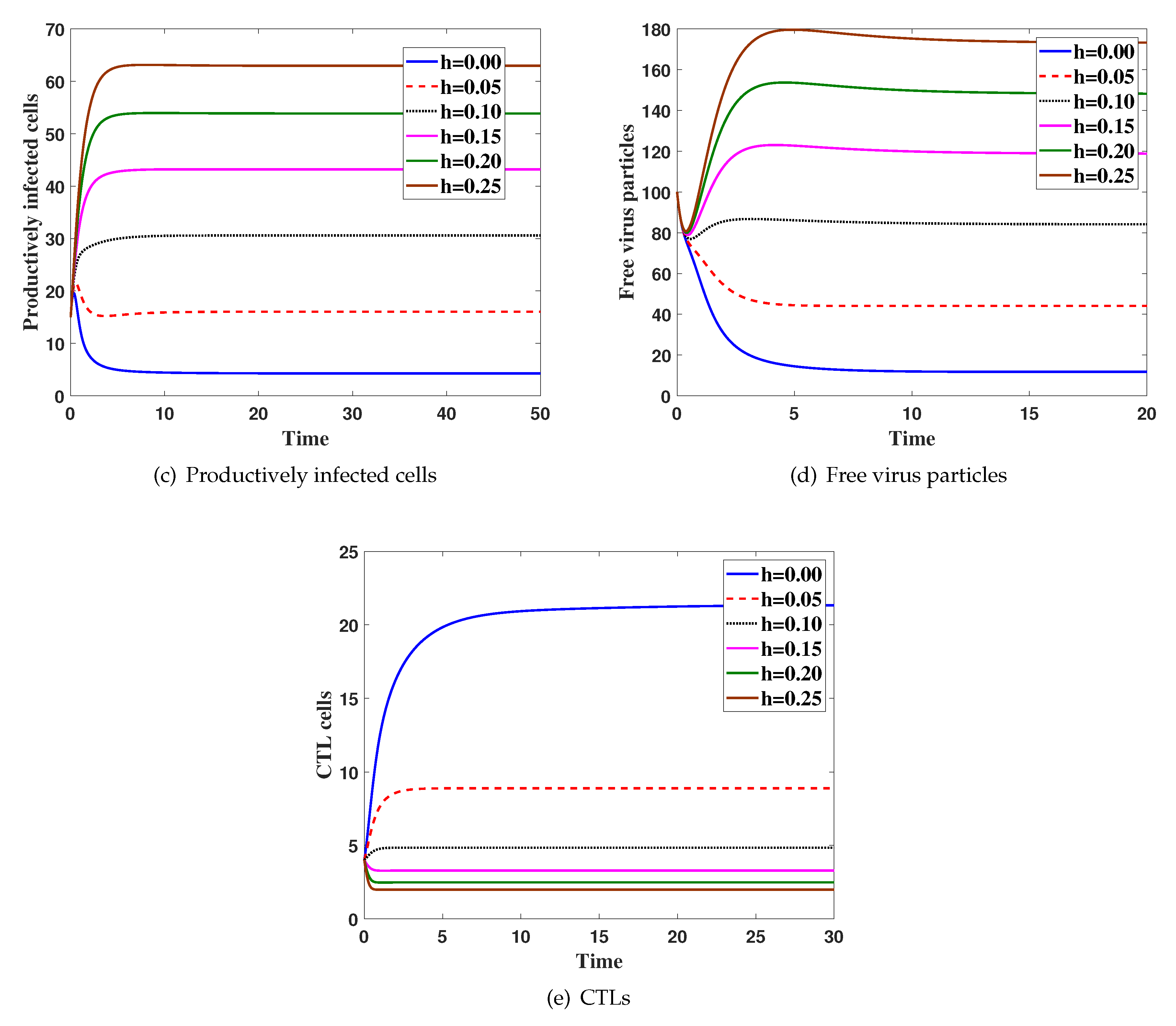
Case(3) Effect ofhon the virus dynamics:
Here, we fix and h is changed. The system is solved with initial condition IC5, Figure 3 shows that the increasing of h will increase both and and decrease all of , and .
5. Discussion and Conclusions
In this paper, we have proposed two virus dynamics models with impairment of CTL functions. We consider that the healthy cells are infected by two ways, viral and cellular infections. We have considered both latently and productively infected cells. The incidence rate is represented by bilinear and saturation in the first and second models, respectively. We have established the well-posedness of the model. We have derived the basic reproduction numbers which determine the existence and stability of the disease-free steady state and endemic steady state of the model. We have investigated the global stability of the steady states of the model by using the Lyapunov method and LaSalle’s invariance principle. We have proven that (i) if , then is globally asymptotically stable and the viruses is cleared (ii) if then exists then it is globally asymptotically stable. This case corresponds to the persistence of the viruses. The effects of saturation and CTL impairment have been studied. We have supported the theoretical results by numerical simulations.
Models (1)–(4) have three steady states; disease-free steady state endemic steady state without a CTL immune response and endemic steady state with a CTL immune response . Moreover, the existence and stability of the steady states are determined by two threshold parameters, the basic reproduction number (which determines whether or not the disease will progress) and the CTL immune response activation number (which determines whether or not a persistent CTL immune response can be established), where
In contrast, models (5)–(8) as well as our proposed models (9)–(13) and (27)–(31) have two steady states ( and ) and their existence and stability are determined by only the basic reproduction number .
It has been reported in several works (see e.g., [10,13,36]) that viruses mutate fast and there is a generation of quasi species that may vary in infectivity. In fact, mutations are one of the ways of immune evasion whereby viruses can evade CTL activity. The high mutation rate of viruses naturally leads to the study of the interplay between immune response and virus diversity for a number of different strains [36]. A viral infection model with CTL immune response and mutations has been proposed in [10] as:
where, is the concentration of actively infected cells with virus strain i, denotes the concentration of different strains of virus particles and denotes the concentration of strain specific immune responses. It has been assumed that there are n diffierent strains of virus. Models (42)–(45) can be extended to take into account (i) cell-to-cell transmision, (ii) latently infected cells, (iii) immune impairment, and (iv) time delay as:
where is the concentration of latently infected cells with virus strain i. Here, is the time between a virus strain i entering an uninfected cell to become latently infected cell with virus strain i, and is the time between a virus strain i entering an uninfected cell and the production of immature viruses of type i. The immature viruses of type i need time to be mature. The factors , and represent the probability of surviving to the age of , and , respectively, where , and, are positive constants. It is worth stressing that the role of the delay term does not only take into account the delay in the dynamical response of the interacting entities, but also their heterogeneity. This can be accounted for by modeling interactions as shown in [37].
Effects of Latent Infection on the Virus Dynamics
In this subsection, we show the effect of the presence of latently infected cells on virus dynamics. Let us incorporate an antiviral drug with efficacy where . The virus dynamics model (9)–(13) under the effect of treatment is given by:
The basic reproduction number for system (51)–(55) is given by
When the population of the latently infected cells are not modeled then models (51)–(55) will become:
and stabilize the disease-free steady state for systems (51)–(55) and (56)–(59). Now, we calculate and as:
Since , then
Clearly, the presence of latently infected cells deceases the basic reproduction number of the system. Now, we aim to determine the minimum drug efficacy that can clear the viruses from the body. We determine and that make
Author Contributions
Conceptualization, A.A.R., A.M.E. and B.S.A.; methodology, A.A.R., A.M.E. and B.S.A.; formal analysis, A.A.R. and B.S.A.; investigation, A.M.E.; writing original draft preparation, A.A.R. and B.S.A.; writing review and editing, A.M.E.
Funding
This research was funded by King Abdulaziz City for Science and Technology (KACST), Saudi Arabia, under grant No. (1-17-01-009-0103).
Acknowledgments
This work was funded by King Abdulaziz City for Science and Technology (KACST), Saudi Arabia, under grant No. (1-17-01-009-0103). The authors, therefore, acknowledge with thanks KACST for technical and financial support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nowak, M.A.; May, R.M. Virus Dynamics: Mathematical Principles of Immunology and Virology; University of Oxford: Oxford, UK, 2000. [Google Scholar]
- Perelson, A.S.; Nelson, P.W. Mathematical analysis of HIV-1 dynamics in vivo. SIAM Rev. 1999, 41, 3–44. [Google Scholar] [CrossRef]
- Huang, G.; Takeuchi, Y.; Ma, W. Lyapunov functionals for delay differential equations model of viral infections. SIAM J. Appl. Math. 2010, 70, 2693–2708. [Google Scholar] [CrossRef]
- Korobeinikov, A. Global properties of basic virus dynamics models. Bull. Math. Biol. 2004, 66, 879–883. [Google Scholar] [CrossRef] [PubMed]
- Elaiw, A.M. Global properties of a class of HIV models. Nonlinear Anal. 2010, 11, 2253–2263. [Google Scholar] [CrossRef]
- Elaiw, A.M. Global properties of a class of virus infection models with multitarget cells. Nonlinear Dyn. 2012, 69, 423–435. [Google Scholar] [CrossRef]
- Elaiw, A.M.; AlShamrani, N.H. Global stability of humoral immunity virus dynamics models with nonlinear infection rate and removal. Nonlinear Anal. 2015, 26, 161–190. [Google Scholar] [CrossRef]
- Zhang, S.; Xu, X. Dynamic analysis and optimal control for a model of hepatitis C with treatment. Commun. Nonlinear Sci. Numer. Simulat. 2017, 46, 14–25. [Google Scholar] [CrossRef]
- Acevedo, H.G.; Li, M.Y. Backward bifurcation in a model for HTLV-I infection of CD4+ T cells. Bull. Math. Biol. 2005, 67, 101–114. [Google Scholar]
- Nowak, M.A.; Bangham, C.R.M. Population dynamics of immune responses to persistent viruses. Science 1996, 272, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Shu, H. Global dynamics of a mathematical model for HTLV-I infection of CD4+ T cells with delayed CTL response. Nonlinear Anal. 2012, 13, 1080–1092. [Google Scholar] [CrossRef]
- Shu, H.; Wang, L.; Watmough, J. Global stability of a nonlinear viral infection model with infinitely distributed intracellular delays and CTL imune responses. SIAM J. Appl. Math. 2013, 73, 1280–1302. [Google Scholar] [CrossRef]
- Regoes, R.; Wodarz, D.; Nowak, M.A. Virus dynamics: The effect to target cell limitation and immune responses on virus evolution. J. Theor. Biol. 1998, 191, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Krishnapriya, P.; Pitchaimani, M. Modeling and bifurcation analysis of a viral infection with time delay and immune impairment. Jpn. J. Ind. Appl. Math. 2017, 34, 99–139. [Google Scholar] [CrossRef]
- Jia, J.; Shi, X. Analysis of a viral infection model with immune impairment and cure rate. J. Nonlinear Sci. Appl. 2016, 9, 3287–3298. [Google Scholar] [CrossRef]
- Shu, H.; Chen, Y.; Wang, L. Impacts of the cell-free and cell-to-cell infection modes on viral dynamics. J. Dyn. Diff. Equat. 2018, 30, 1817–1836. [Google Scholar] [CrossRef]
- Jolly, C.; Sattentau, Q. Retroviral spread by induction of virological synapses. Traffic 2004, 5, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, M.; Nikolic, D.S.; Piguet, V. How HIV-1 takes advantage of the cytoskeleton during replication and cell-to-cell transmission. Viruses 2011, 3, 1757–1776. [Google Scholar] [CrossRef]
- Phillips, D.M. The role of cell-to-cell transmission in HIV infection. AIDS 1994, 8, 719–731. [Google Scholar] [CrossRef]
- Sato, H.; Orenstein, J.; Dimitrov, D.; Martin, M. Cell-to-cell spread of HIV-1 occurs within minutes and may not involve the participation of virus particles. Virology 1992, 186, 712–724. [Google Scholar] [CrossRef]
- Iwami, S.; Takeuchi, J.S.; Nakaoka, S.; Mammano, F.; Clavel, F.; Inaba, H.; Kobayashi, T.; Misawa, N.; Aihara, K.; Koyanagi, Y.; et al. Cell-to-cell infection by HIV contributes over half of virus infection. Elife 2015, 4, e08150. [Google Scholar] [CrossRef]
- Sigal, A.; Kim, J.T.; Balazs, A.B.; Dekel, E.; Mayo, A.; Milo, R.; Baltimore, D. Cell-to-cell spread of HIV permits ongoing replication despite antiretroviral therapy. Nature 2011, 47, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Komarova, N.L.; Anghelina, D.; Voznesensky, I.; Trinite, B.; Levy, D.N.; Wodarz, D. Relative contribution of free-virus and synaptic transmission to the spread of HIV-1 through target cell populations. Biol. Lett. 2012, 9, 1049–1055. [Google Scholar] [CrossRef]
- Komarova, N.L.; Wodarz, D. Virus dynamics in the presence of synaptic transmission. Math. Biosci. 2013, 242, 161–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culshaw, R.V.; Ruan, S.; Webb, G. A mathematical model of cell-to-cell spread of HIV-1 that includes a time delay. J. Math. Biol. 2003, 46, 425–444. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lang, J.; Zou, X. Analysis of an age structured HIV infection model with virus-to-cell infection and cell-to-cell transmission. Nonlinear Anal. 2017, 34, 75–96. [Google Scholar] [CrossRef]
- Chen, S.-S.; Cheng, C.-Y.; Takeuchi, Y. Stability analysis in delayed within-host viral dynamics with both viral and cellular infections. J. Math. Anal. Appl. 2016, 442, 642–672. [Google Scholar] [CrossRef]
- Lai, X.; Zou, X. Modelling HIV-1 virus dynamics with both virus-to-cell infection and cell-to-cell transmission. SIAM J. Appl. Math. 2014, 74, 898–917. [Google Scholar] [CrossRef]
- Wang, X.; Tang, S.; Song, X.; Rong, L. Mathematical analysis of an HIV latent infection model including both virus-to-cell infection and cell-to-cell transmission. J. Biol. Dyn. 2017, 11, 455–483. [Google Scholar] [CrossRef]
- Yang, Y.; Zou, L.; Ruan, S. Global dynamics of a delayed within-host viral infection model with both virus-to-cell and cell-to-cell transmissions. Math. Biosci. 2015, 270, 183–191. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Raezah, A.A.; Alofi, B.S. Dynamics of delayed pathogen infection models with pathogenic and cellular infections and immune impairment. AIP Adv. 2018, 8, 025323. [Google Scholar] [CrossRef]
- Chun, T.-W.; Stuyver, L.; Mizell, S.B.; Ehler, L.A.; Mican, J.A.M.; Baseler, M.; Lloyd, A.L.; Nowak, M.A.; Fauci, A.S. Presence of an inducible HIV-1 latent reservoir during highly active antiretroviral therapy. Proc. Natl. Acad. Sci. USA 1997, 94, 13193–13197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.K.; Hezareh, M.; Gunthard, H.F.; Havlir, D.V.; Ignacio, C.C.; Spina, C.A.; Richman, D.D. Recovery of replication-competent HIV despite prolonged suppression of plasma viremia. Science 1997, 278, 1291–1295. [Google Scholar] [CrossRef]
- Hale, J.K.; Lunel, S.M.V. Introduction to Functional Differential Equations; Springer: New York, NY, USA, 1993. [Google Scholar]
- Song, X.; Neumann, A. Global stability and periodic solution of the viral dynamics. J. Math. Anal. Appl. 2007, 329, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Souza, M.O.; Zubelli, J.P. Global stability for a class of virus models with Cytotoxic T Lymphocyte immune response and antigenic variation. Bull. Math. Biol. 2011, 73, 609–625. [Google Scholar] [CrossRef] [PubMed]
- Gibelli, L.; Elaiw, A.; Alghamdi, M.A.; Althiabi, A.M. Heterogeneous population dynamics of active particles: Progression, mutations, and selection dynamics. Math. Models Methods Appl. Sci. 2017, 27, 617–640. [Google Scholar] [CrossRef]
Figure 1.
The simulation of trajectories of system (27)–(31) with IC1–IC3.


Figure 2.
The simulation of trajectories of system (27)–(31) with different values of α.


Figure 3.
The simulation of trajectories of system (27)–(31) with different value h.


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Raezah, A.A.; Elaiw, A.M.; Alofi, B.S. Global Properties of Latent Virus Dynamics Models with Immune Impairment and Two Routes of Infection. High-Throughput 2019, 8, 16. https://0-doi-org.brum.beds.ac.uk/10.3390/ht8020016
AMA Style
Raezah AA, Elaiw AM, Alofi BS. Global Properties of Latent Virus Dynamics Models with Immune Impairment and Two Routes of Infection. High-Throughput. 2019; 8(2):16. https://0-doi-org.brum.beds.ac.uk/10.3390/ht8020016
Chicago/Turabian StyleRaezah, Aeshah A., Ahmed M. Elaiw, and Badria S. Alofi. 2019. "Global Properties of Latent Virus Dynamics Models with Immune Impairment and Two Routes of Infection" High-Throughput 8, no. 2: 16. https://0-doi-org.brum.beds.ac.uk/10.3390/ht8020016
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.