
The Effect of Manure from Cattle Fed Barley- vs. Corn-Based Diets on Greenhouse Gas Emissions Depends on Soil Type



Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
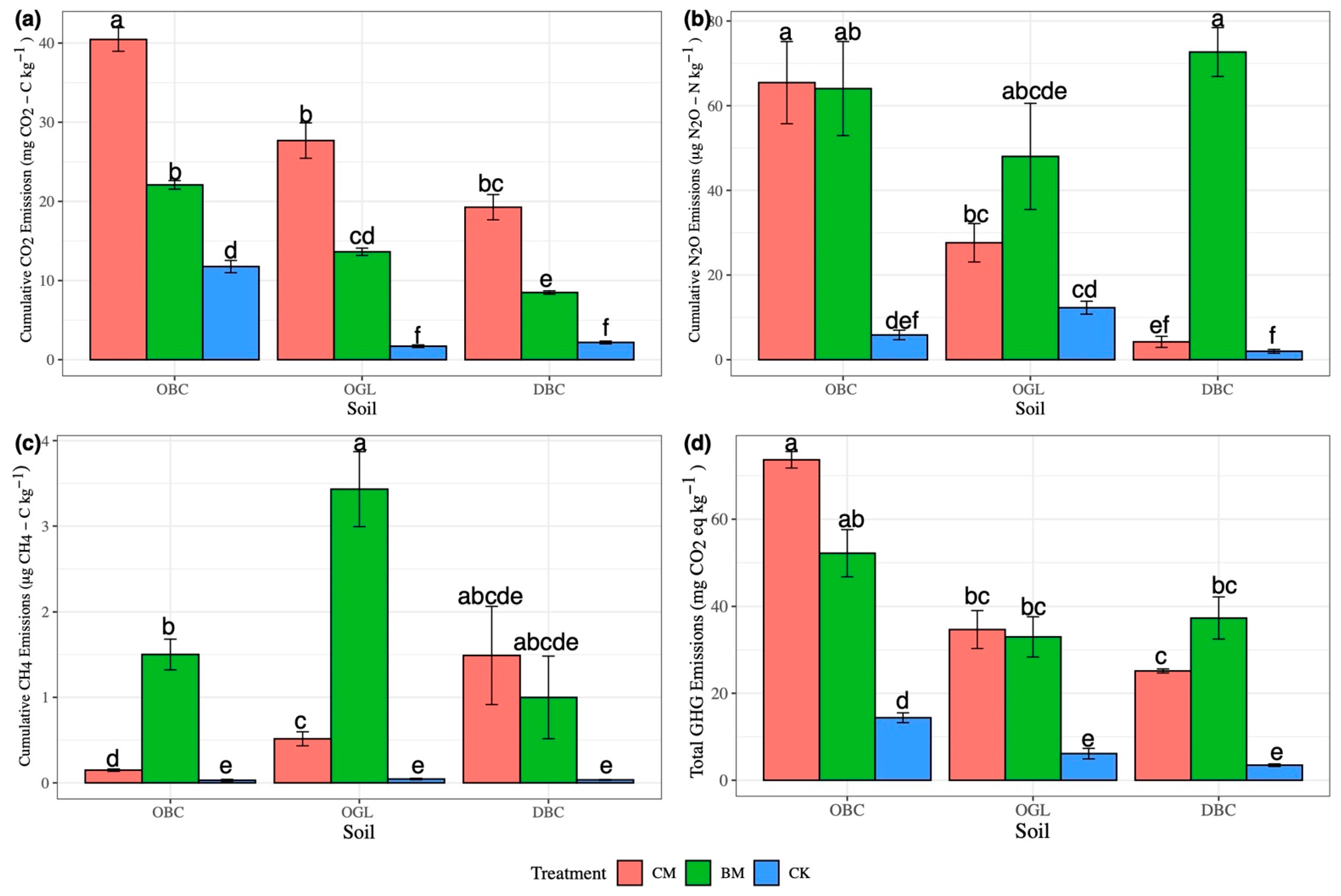
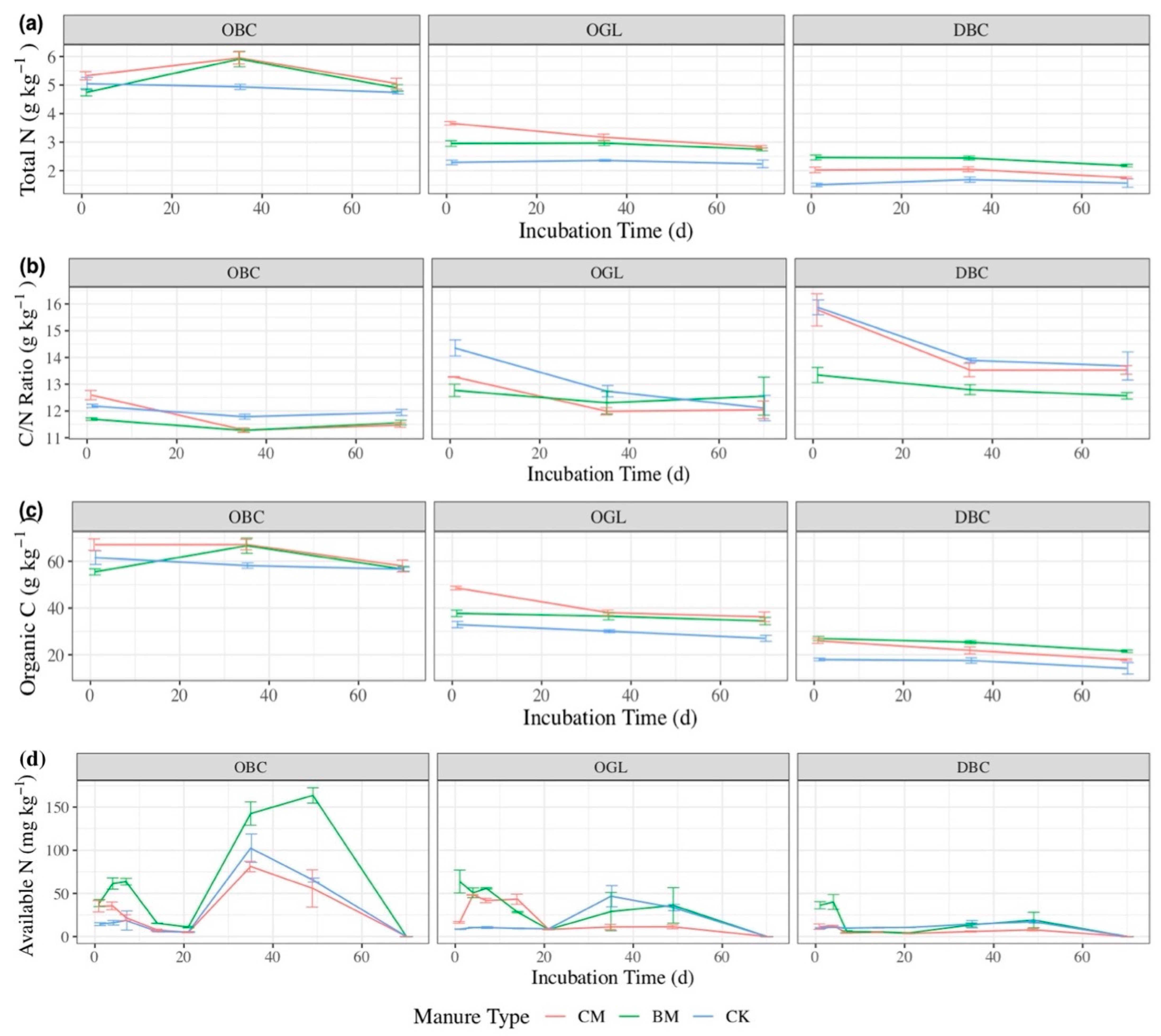
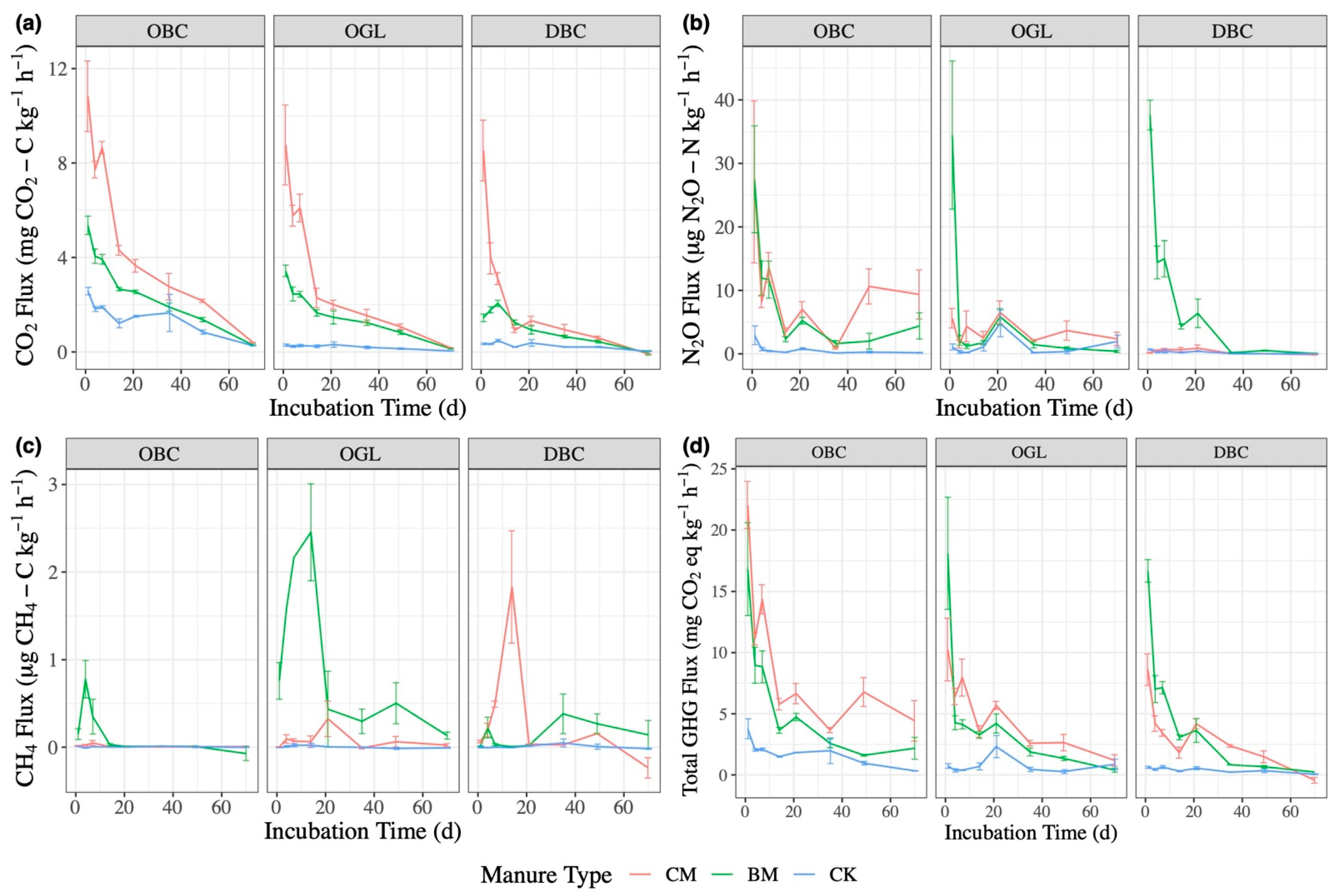
3.1. CO2 Emissions
3.2. N2O Emissions
3.3. CH4 Emissions
3.4. Total GHG Emissions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Vries, M.; de Boer, I.J.M. Comparing Environmental Impacts for Livestock Products: A Review of Life Cycle Assessments. Livest. Sci. 2010, 128, 1–11. [Google Scholar] [CrossRef]
- Environment and Climate Change Canada National Inventory Report 1990–2018: Greenhouse Gas Sources and Sinks in Canada: Executive Summary; Environment and Climate Change Canada: Gatineau, QC, Canada, 2020.
- Van Vliet, P.C.J.; Reijs, J.W.; Bloem, J.; Dijkstra, J.; de Goede, R.G.M. Effects of Cow Diet on the Microbial Community and Organic Matter and Nitrogen Content of Feces. J. Dairy Sci. 2007, 90, 5146–5158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozbayram, E.; Ince, O.; Ince, B.; Harms, H.; Kleinsteuber, S. Comparison of Rumen and Manure Microbiomes and Implications for the Inoculation of Anaerobic Digesters. Microorganisms 2018, 6, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spicer, L.A.; Theurer, C.B.; Sowe, J.; Noon, T.H. Ruminal and Post-Ruminal Utilization of Nitrogen and Starch from Sorghum Grain-, Corn- and Barley-Based Diets by Beef Steers. J. Anim. Sci. 1986, 62, 521–530. [Google Scholar] [CrossRef]
- Guyader, J.; Baron, V.; Beauchemin, K. Corn Forage Yield and Quality for Silage in Short Growing Season Areas of the Canadian Prairies. Agronomy 2018, 8, 164. [Google Scholar] [CrossRef] [Green Version]
- Beauchemin, K.A.; McGinn, S.M. Methane Emissions from Feedlot Cattle Fed Barley or Corn Diets. J. Anim. Sci. 2005, 83, 653–661. [Google Scholar] [CrossRef] [Green Version]
- Eckard, R.J.; Grainger, C.; de Klein, C.A.M. Options for the Abatement of Methane and Nitrous Oxide from Ruminant Production: A Review. Livest. Sci. 2010, 130, 47–56. [Google Scholar] [CrossRef]
- Hassanat, F.; Gervais, R.; Massé, D.I.; Petit, H.V.; Benchaar, C. Methane Production, Nutrient Digestion, Ruminal Fermentation, N Balance, and Milk Production of Cows Fed Timothy Silage- or Alfalfa Silage-Based Diets. J. Dairy Sci. 2014, 97, 6463–6474. [Google Scholar] [CrossRef]
- Chibisa, G.E.; Beauchemin, K.A. Effects of Feeding Corn Silage from Short-Season Hybrids and Extending the Backgrounding Period on Production Performance and Carcass Traits of Beef Cattle. J. Anim. Sci. 2018, 96, 2490–2503. [Google Scholar] [CrossRef]
- Hunt, C.W. Factors Affecting the Feeding Quality of Barley for Ruminants. Anim. Feed Sci. Technol. 1996, 62, 37–48. [Google Scholar] [CrossRef]
- Gibb, D.J.; McAllister, T.A. Corn Compared to Barley in Feedlot Diets. In Proceedings of the 3rd Canadian Barley Symposium, Red Deer, AB, Canada, 19–20 June 2003. [Google Scholar]
- Weber, T.L.; Hao, X.; Gross, C.D.; Beauchemin, K.A.; Chang, S.X. Effect of Manure from Cattle Fed 3-Nitrooxypropanol on Anthropogenic Greenhouse Gas Emissions Depends on Soil Type. Agronomy 2021, 11, 371. [Google Scholar] [CrossRef]
- Romero, C.M.; Redman, A.-A.P.H.; Terry, S.A.; Hazendonk, P.; Hao, X.; McAllister, T.A.; Okine, E. Molecular Speciation and Aromaticity of Biochar-Manure: Insights from Elemental, Stable Isotope and Solid-State DPMAS 13C NMR Analyses. J. Environ. Manag. 2021, 280, 111705. [Google Scholar] [CrossRef] [PubMed]
- Soil Classification Working Group. The Canadian System of Soil Classification; Agriculture and Agri-Food Canada Publication: Ottowa, ON, Canada, 1998; ISBN 0-660-17404-9.
- Calderón, F.J.; McCarty, G.W.; Van Kessel, J.A.S.; Reeves, J.B. Carbon and Nitrogen Dynamics during Incubation of Manured Soil. Soil Sci. Soc. Am. J. 2004, 68, 1592–1599. [Google Scholar] [CrossRef]
- Cai, Y.; Chang, S.X.; Ma, B.; Bork, E.W. Watering Increased DOC Concentration but Decreased N2O Emission from a Mixed Grassland Soil under Different Defoliation Regimes. Biol. Fertil. Soils 2016, 52, 987–996. [Google Scholar] [CrossRef]
- Jones, C.A. Effect of Soil Texture on Critical Bulk Densities for Root Growth. Soil Sci. Soc. Am. J. 1983, 47, 1208–1211. [Google Scholar] [CrossRef]
- Franzluebbers, A.J. Microbial Activity in Response to Water-Filled Pore Space of Variably Eroded Southern Piedmont Soils. Appl. Soil Ecol. 1999, 11, 91–101. [Google Scholar] [CrossRef]
- Li, P.; Lang, M.; Li, C.; Hao, X. Nitrous Oxide and Carbon Dioxide Emissions from Soils Amended with Compost and Manure from Cattle Fed Diets Containing Wheat Dried Distillers’ Grains with Solubles. Can. J. Soil Sci. 2016, 97, 522–531. [Google Scholar] [CrossRef] [Green Version]
- Murphy, D.V.; Recous, S.; Stockdale, E.A.; Fillery, I.R.P.; Jensen, L.S.; Hatch, D.J.; Goulding, K.W.T. Gross Nitrogen Fluxes in Soil: Theory, Measurement and Application of 15N Pool Dilution Techniques. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2003; Volume 79, pp. 69–118. [Google Scholar]
- Sadeghpour, A.; Ketterings, Q.M.; Vermeylen, F.; Godwin, G.S.; Czymmek, K.J. Soil Properties under Nitrogen- vs. Phosphorus-Based Manure and Compost Management of Corn. Soil Sci. Soc. Am. J. 2016, 80, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An Extraction Method for Measuring Soil Microbial Biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Pattey, E.; Trzcinski, M.K.; Desjardins, R.L. Quantifying the Reduction of Greenhouse Gas Emissions as a Result of Composting Dairy and Beef Cattle Manure. Nutr. Cycl. Agroecosystems 2005, 72, 173–187. [Google Scholar] [CrossRef]
- Norman, A.G.; Bremner, J.M. Inorganic Forms of Nitrogen. In Methods of Soil Analysis. Part 2: Chemical and Microbiological Properties; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1965; pp. 672–676. ISBN 978-0-89118-204-7. [Google Scholar]
- Ramnarine, R.; Voroney, R.P.; Wagner-Riddle, C.; Dunfield, K.E. Carbonate Removal by Acid Fumigation for Measuring the δ13C of Soil Organic Carbon. Can. J. Soil Sci. 2011, 91, 247–250. [Google Scholar] [CrossRef] [Green Version]
- Premrov, A.; Cummins, T.; Byrne, K.A. Bulk-Density Modelling Using Optimal Power-Transformation of Measured Physical and Chemical Soil Parameters. Geoderma 2018, 314, 205–220. [Google Scholar] [CrossRef]
- Bremner, J.M. Total Nitrogen. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties Soil Analysis; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1965; pp. 1149–1178. [Google Scholar]
- Van Zwieten, L.; Kimber, S.A.; Morris, S.A.; Downie, A.B.; Berger, E.A.; Rust, J.A.; Scheer, C.D. Influence of Biochars on Flux of N2O and CO2 from Ferrosol. Aust. J. Soil Res. 2010, 48, 555–568. [Google Scholar] [CrossRef]
- Moraghan, J.T.; Buresh, R. Correction for Dissolved Nitrous Oxide in Nitrogen Studies. Soil Sci. Soc. Am. J. 1977, 41, 1201–1202. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014; ISBN 92-9169-113-5. [Google Scholar]
- Hwang, H.Y.; Kim, S.H.; Kim, M.S.; Park, S.J.; Lee, C.H. Co-Composting of Chicken Manure with Organic Wastes: Characterization of Gases Emissions and Compost Quality. Appl. Biol. Chem. 2020, 63, 3. [Google Scholar] [CrossRef]
- Shakoor, A.; Shakoor, S.; Rehman, A.; Ashraf, F.; Abdullah, M.; Shahzad, S.M.; Farooq, T.H.; Ashraf, M.; Manzoor, M.A.; Altaf, M.M.; et al. Effect of Animal Manure, Crop Type, Climate Zone, and Soil Attributes on Greenhouse Gas Emissions from Agricultural Soils—A Global Meta-Analysis. J. Clean. Prod. 2021, 278, 124019. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Friedel, J.K.; Stahr, K. Review of Mechanisms and Quantification of Priming Effects. Soil Biol. Biochem. 2000, 32, 1485–1498. [Google Scholar] [CrossRef]
- Gregorich, E.G.; Rochette, P.; McGuire, S.; Liang, B.C.; Lessard, R. Soluble Organic Carbon and Carbon Dioxide Fluxes in Maize Fields Receiving Spring-Applied Manure. J. Environ. Qual. 1998, 27, 209–214. [Google Scholar] [CrossRef]
- Coelho, J.J.; Hennessy, A.; Casey, I.; Bragança, C.R.S.; Woodcock, T.; Kennedy, N. Biofertilisation with Anaerobic Digestates: A Field Study of Effects on Soil Microbial Abundance and Diversity. Appl. Soil Ecol. 2020, 147, 103403. [Google Scholar] [CrossRef]
- Roman-Perez, C.C.; Hernandez-Ramirez, G. Nitrous Oxide Production and Nitrogen Transformations in a Soil Amended with Biosolids. Can. J. Soil Sci. 2021, 1–14. [Google Scholar] [CrossRef]
- Domsch, K.H.; Anderson, T.-H. The Metabolic Quotient for CO2 (QCO2) as a Specific Activity Parameter to Assess the Effects of Environmental Conditions, Such as pH, on the Microbial Biomass of Forest Soils. Soil Biol. Biochem. 1993, 25, 393–395. [Google Scholar] [CrossRef]
- Ørskov, E.R. Starch Digestion and Utilization in Ruminants. J. Anim. Sci. 1986, 63, 1624–1633. [Google Scholar] [CrossRef] [PubMed]
- Staley, B.F.; de los Reyes, F.L.; Barlaz, M.A. Effect of Spatial Differences in Microbial Activity, pH, and Substrate Levels on Methanogenesis Initiation in Refuse. Appl. Environ. Microbiol. 2011, 77, 2381–2391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampanpanish, P. Effect of Organic Fertilizer on CO2, CH4 and N2O Emissions in a Paddy Field. Mod. Appl. Sci. 2012, 6, 239. [Google Scholar] [CrossRef]
- Thangarajan, R.; Bolan, N.S.; Tian, G.; Naidu, R.; Kunhikrishnan, A. Role of Organic Amendment Application on Greenhouse Gas Emission from Soil. Sci. Total Environ. 2013, 465, 72–96. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Parameter | pH ** | TN | OC | C/N | AN |
|---|---|---|---|---|---|
| (g kg−1) | (g kg−1) | (mg kg−1) | |||
| Manure Type * | |||||
| CM | 8.31 ± 0.02 | 17.0 ± 2.39 | 238 ± 33.4 | 14.1 ± 0.61 | 51.6 ± 0.53 |
| BM | 8.42 ± 0.01 | 18.2 ± 0.20 | 247 ± 14.9 | 13.6 ± 0.70 | 54.9 ± 0.71 |
| Soil Type | |||||
| OBC | 5.75 ± 0.05 | 5.05 ± 0.22 | 61.6 ± 2.94 | 12.2 ± 0.06 | 14.93 ± 0.59 |
| OGL | 4.92 ± 0.02 | 2.29 ± 0.08 | 32.9 ± 1.35 | 14.4 ± 0.30 | 8.71 ± 0.34 |
| DBC | 7.26 ± 0.03 | 1.51 ± 0.06 | 18.0 ± 0.58 | 15.9 ± 0.28 | 9.85 ± 0.54 |
| Source of Variation | S | M | S × M |
|---|---|---|---|
| CO2 emissions | <0.001 | <0.001 | <0.001 |
| N2O emissions | <0.001 | <0.001 | <0.001 |
| CH4 emissions | 0.155 | <0.001 | <0.001 |
| Total GHG emissions (sum of CO2, N2O, and CH4) | <0.001 | <0.001 | <0.001 |
| Variable | Total Nitrogen | Carbon/Nitrogen | Organic Carbon | Available Nitrogen |
|---|---|---|---|---|
| CO2 | 0.59 ** | −0.52 * | 0.57 ** | 0.42 * |
| N2O | 0.61 ** | −0.20 | 0.49 * | 0.76 *** |
| CH4 | 0.10 | −0.25 | 0.07 | 0.62 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weber, T.L.; Hao, X.; Gross, C.D.; Beauchemin, K.A.; Chang, S.X. The Effect of Manure from Cattle Fed Barley- vs. Corn-Based Diets on Greenhouse Gas Emissions Depends on Soil Type. Soil Syst. 2022, 6, 47. https://0-doi-org.brum.beds.ac.uk/10.3390/soilsystems6020047
Weber TL, Hao X, Gross CD, Beauchemin KA, Chang SX. The Effect of Manure from Cattle Fed Barley- vs. Corn-Based Diets on Greenhouse Gas Emissions Depends on Soil Type. Soil Systems. 2022; 6(2):47. https://0-doi-org.brum.beds.ac.uk/10.3390/soilsystems6020047
Chicago/Turabian StyleWeber, Tien L., Xiying Hao, Cole D. Gross, Karen A. Beauchemin, and Scott X. Chang. 2022. "The Effect of Manure from Cattle Fed Barley- vs. Corn-Based Diets on Greenhouse Gas Emissions Depends on Soil Type" Soil Systems 6, no. 2: 47. https://0-doi-org.brum.beds.ac.uk/10.3390/soilsystems6020047