The Effects of Iron Supplementation and Fortification on the Gut Microbiota: A Review

 , ,
, , {kind=link}
Abstract
:1. Introduction
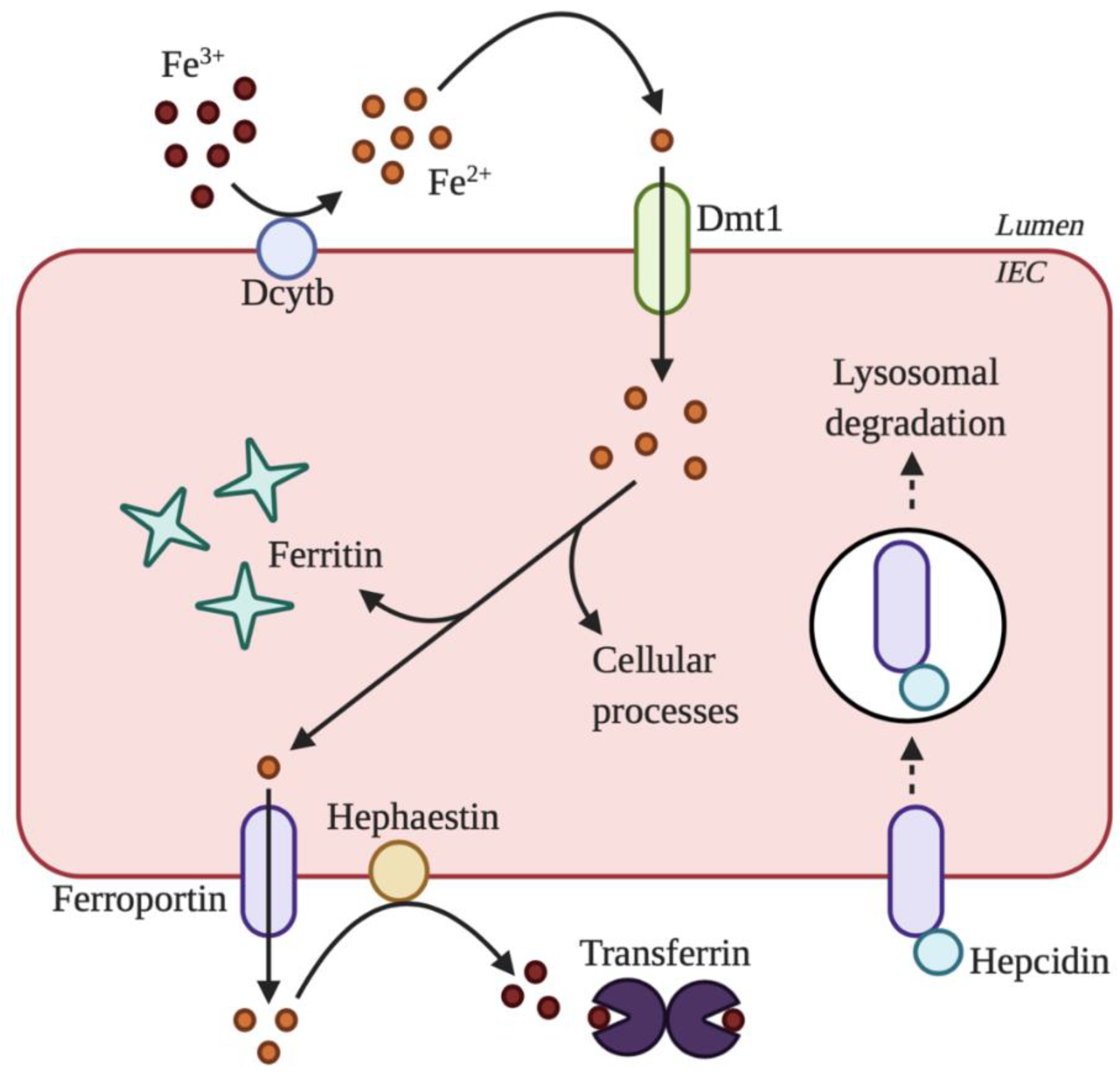
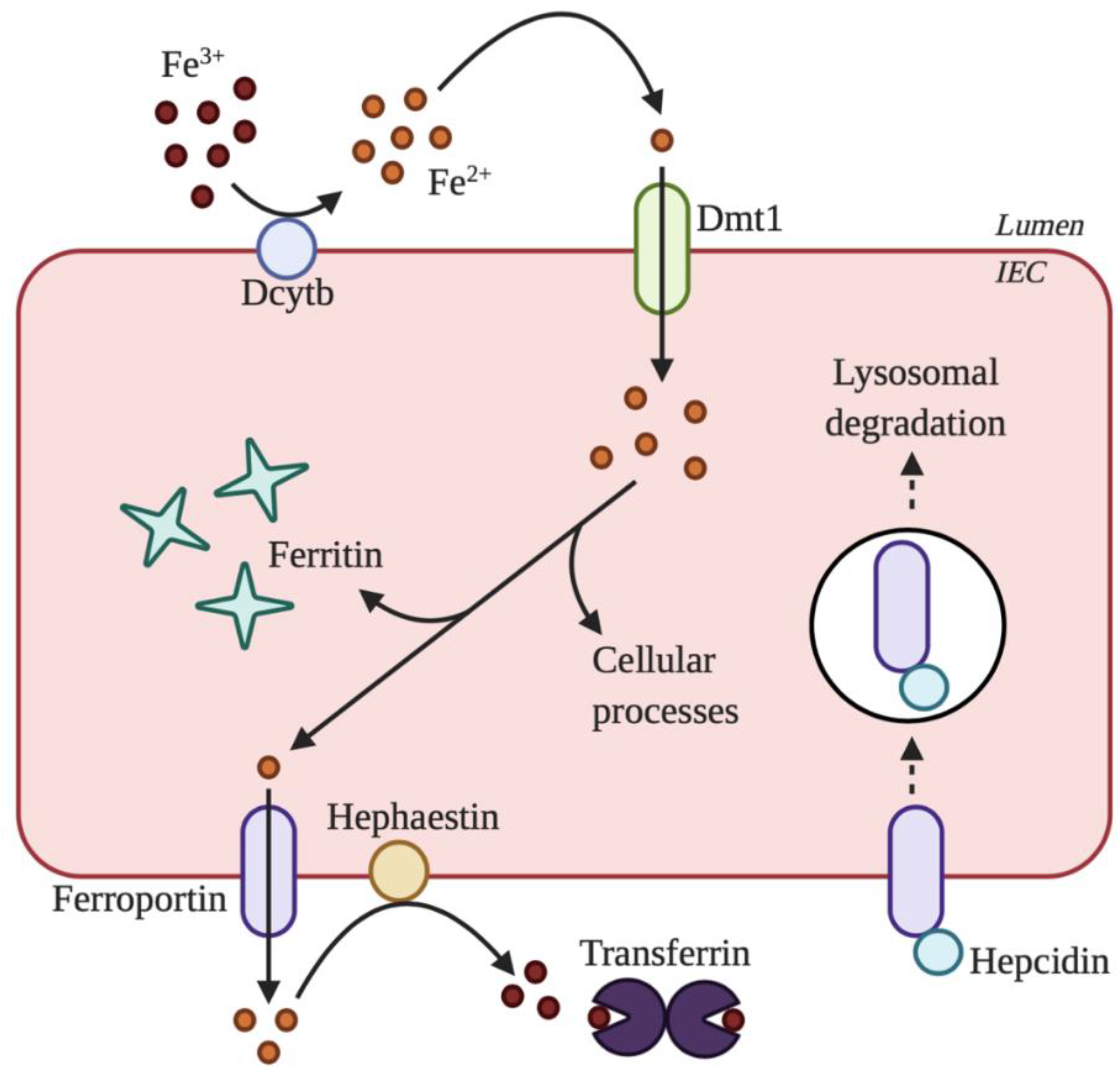
2. Overview of Iron Absorption
3. Maintenance of Systemic Iron Homeostasis
4. Role of the Gut Microbiota in Maintaining Iron Homeostasis
5. Effect of Iron Supplementation and Fortification on the Gut Microbiota
5.1. Animal Studies
5.2. Human Studies
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cassat, J.E.; Skaar, E.P. Iron in Infection and Immunity. Cell Host Microbe 2013, 13, 509–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yilmaz, B.; Li, H. Gut Microbiota and Iron: The Crucial Actors in Health and Disease. Pharmaceuticals 2018, 11, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, S.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef] [Green Version]
- Abbaspour, N.; Hurrell, R.; Kelishadi, R. Review on iron and its importance for human health. J. Res. Med. Sci. 2014, 19, 164–174. [Google Scholar] [PubMed]
- Winter, W.E.; Bazydlo, L.A.L.; Harris, N.S. The Molecular Biology of Human Iron Metabolism. Lab. Med. 2014, 45, 92–102. [Google Scholar] [CrossRef]
- Gulec, S.; Anderson, G.J.; Collins, J.F. Mechanistic and regulatory aspects of intestinal iron absorption. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G397–G409. [Google Scholar] [CrossRef] [Green Version]
- Ganz, T. Hepcidin, a key regulator of iron metabolism and mediator of anemia of inflammation. Blood 2003, 102, 783–788. [Google Scholar] [CrossRef] [Green Version]
- McKie, A.T. An Iron-Regulated Ferric Reductase Associated with the Absorption of Dietary Iron. Science 2001, 291, 1755–1759. [Google Scholar] [CrossRef]
- Garrick, M.D.; Dolan, K.G.; Horbinski, C.; Ghio, A.J.; Higgins, D.; Porubcin, M.; Moore, E.G.; Hainsworth, L.N.; Umbreit, J.N.; Conrad, M.E.; et al. DMT1: A mammalian transporter for multiple metals. BioMetals 2003, 16, 41–54. [Google Scholar] [CrossRef]
- Drakesmith, H.; Nemeth, E.; Ganz, T. Ironing out Ferroportin. Cell Metab. 2015, 22, 777–787. [Google Scholar] [CrossRef] [Green Version]
- Anderson, G.J.; Frazer, D.M. Current understanding of iron homeostasis. Am. J. Clin. Nutr. 2017, 106, 1559S–1566S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, A.R.; Oates, P.S. Mechanisms of heme iron absorption: Current questions and controversies. World J. Gastroenterol. 2008, 14, 4101. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.F. The Regulation of Iron Absorption and Homeostasis. Clin. Biochem. Rev. 2016, 37, 51–62. [Google Scholar]
- Conway, D.; Henderson, M.A. Iron metabolism. Anaesth. Intensive Care Med. 2019, 20, 175–177. [Google Scholar] [CrossRef]
- Aschemeyer, S.; Qiao, B.; Stefanova, D.; Valore, E.V.; Sek, A.C.; Ruwe, T.A.; Vieth, K.R.; Jung, G.; Casu, C.; Rivella, S.; et al. Structure-function analysis of ferroportin defines the binding site and an alternative mechanism of action of hepcidin. Blood 2018, 131, 899–910. [Google Scholar] [CrossRef]
- Jones, E.; Pasricha, S.-R.; Allen, A.; Evans, P.; Fisher, C.A.; Wray, K.; Premawardhena, A.; Bandara, D.; Perera, A.; Webster, C.; et al. Hepcidin is suppressed by erythropoiesis in hemoglobin E β-thalassemia and β-thalassemia trait. Blood 2015, 125, 873–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paganini, D.; Zimmermann, M.B. The effects of iron fortification and supplementation on the gut microbiome and diarrhea in infants and children: A review. Am. J. Clin. Nutr. 2017, 106, 1688S–1693S. [Google Scholar] [CrossRef] [Green Version]
- Taher, A.T.; Saliba, A.N. Iron overload in thalassemia: Different organs at different rates. Hematology 2017, 2017, 265–271. [Google Scholar] [CrossRef] [Green Version]
- Papanikolaou, G.; Tzilianos, M.; Christakis, J.I.; Bogdanos, D.; Tsimirika, K.; MacFarlane, J.; Goldberg, Y.P.; Sakellaropoulos, N.; Ganz, T.; Nemeth, E. Hepcidin in iron overload disorders. Blood 2005, 105, 4103–4105. [Google Scholar] [CrossRef]
- Porter, J.L.; Rawla, P. Hemochromatosis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Xue, X.; Shah, Y. Intestinal Iron Homeostasis and Colon Tumorigenesis. Nutrients 2013, 5, 2333–2351. [Google Scholar] [CrossRef]
- Nielsen, O.H.; Ainsworth, M.; Coskun, M.; Weiss, G. Management of Iron-Deficiency Anemia in Inflammatory Bowel Disease: A Systematic Review. Medicine 2015, 94, e963. [Google Scholar] [CrossRef] [PubMed]
- Moran-Lev, H.; Galai, T.; Yerushalmy-Feler, A.; Weisman, Y.; Anafy, A.; Deutsch, V.; Cipok, M.; Lubetzky, R.; Cohen, S. Vitamin D Decreases Hepcidin and Inflammatory Markers in Newly Diagnosed Inflammatory Bowel Disease Paediatric Patients: A Prospective Study. J. Crohns Colitis 2019, 13, 1287–1291. [Google Scholar] [CrossRef] [PubMed]
- Krawiec, P.; Mroczkowska-Juchkiewicz, A.; Pac-Kożuchowska, E. Serum Hepcidin in Children with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2017, 23, 2165–2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paköz, Z.B.; Çekiç, C.; Arabul, M.; Sarıtaş Yüksel, E.; İpek, S.; Vatansever, S.; Ünsal, B. An Evaluation of the Correlation between Hepcidin Serum Levels and Disease Activity in Inflammatory Bowel Disease. Gastroenterol. Res. Pract. 2015, 2015, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanmugam, N.K.N.; Trebicka, E.; Fu, L.; Shi, H.N.; Cherayil, B.J. Intestinal Inflammation Modulates Expression of the Iron-Regulating Hormone Hepcidin Depending on Erythropoietic Activity and the Commensal Microbiota. J. Immunol. 2014, 193, 1398–1407. [Google Scholar] [CrossRef]
- Shu, W.; Pang, Z.; Xu, C.; Lin, J.; Li, G.; Wu, W.; Sun, S.; Li, J.; Li, X.; Liu, Z. Anti-TNF-α Monoclonal Antibody Therapy Improves Anemia through Downregulating Hepatocyte Hepcidin Expression in Inflammatory Bowel Disease. Mediat. Inflamm. 2019, 2019, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Caza, M.; Kronstad, J.W. Shared and distinct mechanisms of iron acquisition by bacterial and fungal pathogens of humans. Front. Cell. Infect. Microbiol. 2013, 3. [Google Scholar] [CrossRef]
- Lönnerdal, B. Excess iron intake as a factor in growth, infections, and development of infants and young children. Am. J. Clin. Nutr. 2017, 106, 1681S–1687S. [Google Scholar] [CrossRef]
- Page, M.G.P. The Role of Iron and Siderophores in Infection, and the Development of Siderophore Antibiotics. Clin. Infect. Dis. 2019, 69, S529–S537. [Google Scholar] [CrossRef]
- Wilson, B.R.; Bogdan, A.R.; Miyazawa, M.; Hashimoto, K.; Tsuji, Y. Siderophores in Iron Metabolism: From Mechanism to Therapy Potential. Trends Mol. Med. 2016, 22, 1077–1090. [Google Scholar] [CrossRef] [Green Version]
- Golonka, R.; Yeoh, B.S.; Vijay-Kumar, M. The Iron Tug-of-War between Bacterial Siderophores and Innate Immunity. J. Innate Immun. 2019, 11, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Qi, B.; Han, M. Microbial Siderophore Enterobactin Promotes Mitochondrial Iron Uptake and Development of the Host via Interaction with ATP Synthase. Cell 2018, 175, 571–582.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosbahi, K.; Wojnowska, M.; Albalat, A.; Walker, D. Bacterial iron acquisition mediated by outer membrane translocation and cleavage of a host protein. Proc. Natl. Acad. Sci. USA 2018, 115, 6840–6845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archibald, F. Lactobacillus plantarum, an organism not requiring iron. FEMS Microbiol. Lett. 1983, 19, 29–32. [Google Scholar] [CrossRef]
- Weinberg, E.D. The Lactobacillus Anomaly: Total Iron Abstinence. Perspect. Biol. Med. 1997, 40, 578–583. [Google Scholar] [CrossRef]
- Das, N.K.; Schwartz, A.J.; Barthel, G.; Inohara, N.; Liu, Q.; Sankar, A.; Hill, D.R.; Ma, X.; Lamberg, O.; Schnizlein, M.K.; et al. Microbial Metabolite Signaling Is Required for Systemic Iron Homeostasis. Cell Metab. 2020, 31, 115–130.e6. [Google Scholar] [CrossRef]
- Buhnik-Rosenblau, K.; Moshe-Belizowski, S.; Danin-Poleg, Y.; Meyron-Holtz, E.G. Genetic modification of iron metabolism in mice affects the gut microbiota. BioMetals 2012, 25, 883–892. [Google Scholar] [CrossRef]
- Balamurugan, R.; Mary, R.R.; Chittaranjan, S.; Jancy, H.; Shobana Devi, R.; Ramakrishna, B.S. Low levels of faecal lactobacilli in women with iron-deficiency anaemia in south India. Br. J. Nutr. 2010, 104, 931–934. [Google Scholar] [CrossRef] [Green Version]
- Kalipatnapu, S.; Kuppuswamy, S.; Venugopal, G.; Kaliaperumal, V.; Ramadass, B. Fecal total iron concentration is inversely associated with fecal Lactobacillus in preschool children: Fecal iron is inversely associated with Fecal Lactobacillus. J. Gastroenterol. Hepatol. 2017, 32, 1475–1479. [Google Scholar] [CrossRef]
- Vonderheid, S.C.; Tussing-Humphreys, L.; Park, C.; Pauls, H.; OjiNjideka Hemphill, N.; LaBomascus, B.; McLeod, A.; Koenig, M.D. A Systematic Review and Meta-Analysis on the Effects of Probiotic Species on Iron Absorption and Iron Status. Nutrients 2019, 11, 2938. [Google Scholar] [CrossRef] [Green Version]
- Sandberg, A.-S.; Önning, G.; Engström, N.; Scheers, N. Iron Supplements Containing Lactobacillus plantarum 299v Increase Ferric Iron and Up-regulate the Ferric Reductase DCYTB in Human Caco-2/HT29 MTX Co-Cultures. Nutrients 2018, 10, 1949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, G.M.; Morrissette, S.; Larson, A.; Stading, P.; Griffin, K.H.; Barnes, T.L. Use of a Probiotic to Enhance Iron Absorption in a Randomized Trial of Pediatric Patients Presenting with Iron Deficiency. J. Pediatr. 2019, 207, 192–197.e1. [Google Scholar] [CrossRef] [PubMed]
- Rusu, I.G.; Suharoschi, R.; Vodnar, D.C.; Pop, C.R.; Socaci, S.A.; Vulturar, R.; Istrati, M.; Moroșan, I.; Fărcaș, A.C.; Kerezsi, A.D.; et al. Iron Supplementation Influence on the Gut Microbiota and Probiotic Intake Effect in Iron Deficiency—A Literature-Based Review. Nutrients 2020, 12, 1993. [Google Scholar] [CrossRef]
- Deschemin, J.; Noordine, M.; Remot, A.; Willemetz, A.; Afif, C.; Canonne-Hergaux, F.; Langella, P.; Karim, Z.; Vaulont, S.; Thomas, M.; et al. The microbiota shifts the iron sensing of intestinal cells. FASEB J. 2016, 30, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Reddy, B.S.; Pleasants, J.R.; Wostmann, B.S. Effect of Intestinal Microflora on Iron and Zinc Metabolism, and on Activities of Metalloenzymes in Rats. J. Nutr. 1972, 102, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Forrester, R.H.; Conrad, M.E.; Crosby, W.H. Measurement of total body iron in animals using whole-body liquid scintillation detectors. Exp. Biol. Med. 1962, 111, 115–119. [Google Scholar] [CrossRef]
- Stern, P.; Košak, R.; Misirlija, A. The problem of iron resorption. Experientia 1954, 10, 227. [Google Scholar] [CrossRef]
- Sanchez, K.K.; Chen, G.Y.; Schieber, A.M.P.; Redford, S.E.; Shokhirev, M.N.; Leblanc, M.; Lee, Y.M.; Ayres, J.S. Cooperative Metabolic Adaptations in the Host Can Favor Asymptomatic Infection and Select for Attenuated Virulence in an Enteric Pathogen. Cell 2018, 175, 146–158.e15. [Google Scholar] [CrossRef] [Green Version]
- Kortman, G.A.M.; Boleij, A.; Swinkels, D.W.; Tjalsma, H. Iron Availability Increases the Pathogenic Potential of Salmonella Typhimurium and Other Enteric Pathogens at the Intestinal Epithelial Interface. PLoS ONE 2012, 7, e29968. [Google Scholar] [CrossRef]
- Abbott, M.; Galloway, A.; Cunningham, J.L. Haemochromatosis presenting with a double yersinia infection. J. Infect. 1986, 13, 143–145. [Google Scholar] [CrossRef]
- Van Asbeck, B.S.; Verbrugh, H.A.; van Oost, B.A.; Marx, J.J.; Imhof, H.W.; Verhoef, J. Listeria monocytogenes meningitis and decreased phagocytosis associated with iron overload. BMJ 1982, 284, 542–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moura; Verheul; Marx A functional defect in hereditary haemochromatosis monocytes and monocyte-derived macrophages. Eur. J. Clin. Invest. 1998, 28, 164–173. [CrossRef] [PubMed]
- Núñez, G.; Sakamoto, K.; Soares, M.P. Innate Nutritional Immunity. J. Immunol. 2018, 201, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Barton, J.C.; Acton, R.T. Hepcidin, iron, and bacterial infection. In Vitamins and Hormones; Elsevier: Amsterdam, The Netherlands, 2019; Volume 110, pp. 223–242. ISBN 978-0-12-817842-3. [Google Scholar]
- Chen, G.Y.; Ayres, J.S. Beyond tug-of-war: Iron metabolism in cooperative host–microbe interactions. PLoS Pathog. 2020, 16, e1008698. [Google Scholar] [CrossRef] [PubMed]
- Parmanand, B.A.; Kellingray, L.; Le Gall, G.; Basit, A.W.; Fairweather-Tait, S.; Narbad, A. A decrease in iron availability to human gut microbiome reduces the growth of potentially pathogenic gut bacteria; an in vitro colonic fermentation study. J. Nutr. Biochem. 2019, 67, 20–27. [Google Scholar] [CrossRef]
- Kortman, G.A.M.; Mulder, M.L.M.; Richters, T.J.W.; Shanmugam, N.K.N.; Trebicka, E.; Boekhorst, J.; Timmerman, H.M.; Roelofs, R.; Wiegerinck, E.T.; Laarakkers, C.M.; et al. Low dietary iron intake restrains the intestinal inflammatory response and pathology of enteric infection by food-borne bacterial pathogens: Immunity to infection. Eur. J. Immunol. 2015, 45, 2553–2567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phipps, O.; Al-Hassi, H.O.; Quraishi, M.N.; Kumar, A.; Brookes, M.J. Influence of Iron on the Gut Microbiota in Colorectal Cancer. Nutrients 2020, 12, 2512. [Google Scholar] [CrossRef] [PubMed]
- Ng, O. Iron, microbiota and colorectal cancer. Wien. Med. Wochenschr. 2016, 166, 431–436. [Google Scholar] [CrossRef]
- Baltussen, R.; Knai, C.; Sharan, M. Iron Fortification and Iron Supplementation are Cost-Effective Interventions to Reduce Iron Deficiency in Four Subregions of the World. J. Nutr. 2004, 134, 2678–2684. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Nutritional Anaemias: Tools for Effective Prevention and Control; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Patil, S.S.; Khanwelkar, C.C.; Patil, S.K.; Thorat, V.M.; Jadhav, S.A.; Sontakke, A.V. Comparison of efficacy, tolerability, and cost of newer with conventional oral iron preparation. Al Ameen. J. Med. Sci. 2013, 6, 29–33. [Google Scholar]
- Cancelo-Hidalgo, M.J.; Castelo-Branco, C.; Palacios, S.; Haya-Palazuelos, J.; Ciria-Recasens, M.; Manasanch, J.; Pérez-Edo, L. Tolerability of different oral iron supplements: A systematic review. Curr. Med. Res. Opin. 2013, 29, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Tondeur, M.C.; Schauer, C.S.; Christofides, A.L.; Asante, K.P.; Newton, S.; Serfass, R.E.; Zlotkin, S.H. Determination of iron absorption from intrinsically labeled microencapsulated ferrous fumarate (sprinkles) in infants with different iron and hematologic status by using a dual-stable-isotope method. Am. J. Clin. Nutr. 2004, 80, 1436–1444. [Google Scholar] [CrossRef] [PubMed]
- Bagna, R.; Spada, E.; Mazzone, R.; Saracco, P.; Boetti, T.; Cester, E.A.; Bertino, E.; Coscia, A. Efficacy of Supplementation with Iron Sulfate Compared to Iron Bisglycinate Chelate in Preterm Infants. Curr. Pediatr. Rev. 2018, 14, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Milman, N.; Jønsson, L.; Dyre, P.; Pedersen, P.L.; Larsen, L.G. Ferrous bisglycinate 25 mg iron is as effective as ferrous sulfate 50 mg iron in the prophylaxis of iron deficiency and anemia during pregnancy in a randomized trial. J. Perinat. Med. 2014, 42. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, J.; Choudhury, P. Iron formulations in pediatric practice. Indian Pediatr. 2004, 41, 807–815. [Google Scholar] [PubMed]
- Ramsay, L.C.; Charles, C.V. Review of Iron Supplementation and Fortification. In Topics in Public Health; Claborn, D., Ed.; InTech: London, UK, 2015; ISBN 978-953-51-2132-9. [Google Scholar]
- De-Regil, L.M.; Suchdev, P.S.; Vist, G.E.; Walleser, S.; Peña-Rosas, J.P. Home fortification of foods with multiple micronutrient powders for health and nutrition in children under two years of age. Cochrane Database Syst. Rev. 2011. [Google Scholar] [CrossRef]
- Pachón, H.; Spohrer, R.; Mei, Z.; Serdula, M.K. Evidence of the effectiveness of flour fortification programs on iron status and anemia: A systematic review. Nutr. Rev. 2015, 73, 780–795. [Google Scholar] [CrossRef]
- Pereira, D.I.A.; Aslam, M.F.; Frazer, D.M.; Schmidt, A.; Walton, G.E.; McCartney, A.L.; Gibson, G.R.; Anderson, G.J.; Powell, J.J. Dietary iron depletion at weaning imprints low microbiome diversity and this is not recovered with oral nano Fe(III). Microbiol. Open 2015, 4, 12–27. [Google Scholar] [CrossRef]
- Dostal, A.; Chassard, C.; Hilty, F.M.; Zimmermann, M.B.; Jaeggi, T.; Rossi, S.; Lacroix, C. Iron Depletion and Repletion with Ferrous Sulfate or Electrolytic Iron Modifies the Composition and Metabolic Activity of the Gut Microbiota in Rats. J. Nutr. 2012, 142, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Dostal, A.; Fehlbaum, S.; Chassard, C.; Zimmermann, M.B.; Lacroix, C. Low iron availability in continuous in vitro colonic fermentations induces strong dysbiosis of the child gut microbial consortium and a decrease in main metabolites. FEMS Microbiol. Ecol. 2013, 83, 161–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dostal, A.; Lacroix, C.; Bircher, L.; Pham, V.T.; Follador, R.; Zimmermann, M.B.; Chassard, C. Iron Modulates Butyrate Production by a Child Gut Microbiota In Vitro. mBio 2015, 6, e01453-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tompkins, G.R.; O’Dell, N.L.; Bryson, I.T.; Pennington, C.B. The Effects of Dietary Ferric Iron and Iron Deprivation on the Bacterial Composition of the Mouse Intestine. Curr. Microbiol. 2001, 43, 38–42. [Google Scholar] [CrossRef]
- La Carpia, F.; Wojczyk, B.S.; Annavajhala, M.K.; Rebbaa, A.; Culp-Hill, R.; D’Alessandro, A.; Freedberg, D.E.; Uhlemann, A.-C.; Hod, E.A. Transfusional iron overload and intravenous iron infusions modify the mouse gut microbiota similarly to dietary iron. NPJ Biofilms Microbiomes 2019, 5, 26. [Google Scholar] [CrossRef] [PubMed]
- Coffey, R.; Ganz, T. Iron homeostasis: An anthropocentric perspective. J. Biol. Chem. 2017, 292, 12727–12734. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Pan, X.; Liu, Z.; Han, M.; Xu, G.; Dai, X.; Wang, W.; Zhang, H.; Xie, L. Fecal microbiota as a noninvasive biomarker to predict the tissue iron accumulation in intestine epithelial cells and liver. FASEB J. 2020, 34, 3006–3020. [Google Scholar] [CrossRef] [Green Version]
- Alexeev, E.E.; He, X.; Slupsky, C.M.; Lönnerdal, B. Effects of iron supplementation on growth, gut microbiota, metabolomics and cognitive development of rat pups. PLoS ONE 2017, 12, e0179713. [Google Scholar] [CrossRef] [Green Version]
- Mahalhal, A.; Williams, J.M.; Johnson, S.; Ellaby, N.; Duckworth, C.A.; Burkitt, M.D.; Liu, X.; Hold, G.L.; Campbell, B.J.; Pritchard, D.M.; et al. Oral iron exacerbates colitis and influences the intestinal microbiome. PLoS ONE 2018, 13, e0202460. [Google Scholar] [CrossRef] [Green Version]
- Lucas López, R.; Grande Burgos, M.J.; Gálvez, A.; Pérez Pulido, R. The human gastrointestinal tract and oral microbiota in inflammatory bowel disease: A state of the science review. APMIS 2017, 125, 3–10. [Google Scholar] [CrossRef]
- Constante, M.; Fragoso, G.; Lupien-Meilleur, J.; Calvé, A.; Santos, M.M. Iron Supplements Modulate Colon Microbiota Composition and Potentiate the Protective Effects of Probiotics in Dextran Sodium Sulfate-induced Colitis. Inflamm. Bowel Dis. 2017, 23, 753–766. [Google Scholar] [CrossRef]
- Ettreiki, C. Juvenile ferric iron prevents microbiota dysbiosis and colitis in adult rodents. World J. Gastroenterol. 2012, 18, 2619. [Google Scholar] [CrossRef] [PubMed]
- Werner, T.; Wagner, S.J.; Martinez, I.; Walter, J.; Chang, J.-S.; Clavel, T.; Kisling, S.; Schuemann, K.; Haller, D. Depletion of luminal iron alters the gut microbiota and prevents Crohn’s disease-like ileitis. Gut 2011, 60, 325–333. [Google Scholar] [CrossRef]
- Dostal, A.; Baumgartner, J.; Riesen, N.; Chassard, C.; Smuts, C.M.; Zimmermann, M.B.; Lacroix, C. Effects of iron supplementation on dominant bacterial groups in the gut, faecal SCFA and gut inflammation: A randomised, placebo-controlled intervention trial in South African children. Br. J. Nutr. 2014, 112, 547–556. [Google Scholar] [CrossRef]
- Tang, M.; Frank, D.N.; Sherlock, L.; Ir, D.; Robertson, C.E.; Krebs, N.F. Effect of Vitamin E with Therapeutic Iron Supplementation on Iron Repletion and Gut Microbiome in US Iron Deficient Infants and Toddlers. J. Pediatr. Gastroenterol. Nutr. 2016, 63, 379–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitert, M.D.; Gomez-Arango, L.F.; Barrett, H.L.; McIntyre, H.D.; Anderson, G.J.; Frazer, D.M.; Callaway, L.K. Iron supplementation has minor effects on gut microbiota composition in overweight and obese women in early pregnancy. Br. J. Nutr. 2018, 120, 283–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaeggi, T.; Kortman, G.A.M.; Moretti, D.; Chassard, C.; Holding, P.; Dostal, A.; Boekhorst, J.; Timmerman, H.M.; Swinkels, D.W.; Tjalsma, H.; et al. Iron fortification adversely affects the gut microbiome, increases pathogen abundance and induces intestinal inflammation in Kenyan infants. Gut 2015, 64, 731–742. [Google Scholar] [CrossRef]
- Tang, M.; Frank, D.; Hendricks, A.; Ir, D.; Esamai, F.; Liechty, E.; Hambidge, K.; Krebs, N. Iron in Micronutrient Powder Promotes an Unfavorable Gut Microbiota in Kenyan Infants. Nutrients 2017, 9, 776. [Google Scholar] [CrossRef] [Green Version]
- Sjödin, K.S.; Domellöf, M.; Lagerqvist, C.; Hernell, O.; Lönnerdal, B.; Szymlek-Gay, E.A.; Sjödin, A.; West, C.E.; Lind, T. Administration of ferrous sulfate drops has significant effects on the gut microbiota of iron-sufficient infants: A randomised controlled study. Gut 2019, 68, 2095–2097. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V.K.; Paul, S.; Dutta, C. Geography, Ethnicity or Subsistence-Specific Variations in Human Microbiome Composition and Diversity. Front. Microbiol. 2017, 8, 1162. [Google Scholar] [CrossRef] [Green Version]
- Behrouzi, A.; Nafari, A.H.; Siadat, S.D. The significance of microbiome in personalized medicine. Clin. Transl. Med. 2019, 8, 16. [Google Scholar] [CrossRef]
- Sazawal, S.; Black, R.E.; Ramsan, M.; Chwaya, H.M.; Stoltzfus, R.J.; Dutta, A.; Dhingra, U.; Kabole, I.; Deb, S.; Othman, M.K.; et al. Effects of routine prophylactic supplementation with iron and folic acid on admission to hospital and mortality in preschool children in a high malaria transmission setting: Community-based, randomised, placebo-controlled trial. Lancet 2006, 367, 133–143. [Google Scholar] [CrossRef]
- Zhang, D.-L.; Wu, J.; Shah, B.N.; Greutélaers, K.C.; Ghosh, M.C.; Ollivierre, H.; Su, X.; Thuma, P.E.; Bedu-Addo, G.; Mockenhaupt, F.P.; et al. Erythrocytic ferroportin reduces intracellular iron accumulation, hemolysis, and malaria risk. Science 2018, 359, 1520–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Sun, Q.; Liu, J.; Hu, Y.; Liu, S.; Zhang, J.; Sheng, X.; Hambidge, K.M. The Effect of Iron Fortification on Iron (Fe) Status and Inflammation: A Randomized Controlled Trial. PLoS ONE 2016, 11, e0167458. [Google Scholar] [CrossRef] [PubMed]
- Qasem, W.; Azad, M.B.; Hossain, Z.; Azad, E.; Jorgensen, S.; Castillo San Juan, S.; Cai, C.; Khafipour, E.; Beta, T.; Roberts, L.J.; et al. Assessment of complementary feeding of Canadian infants: Effects on microbiome & oxidative stress, a randomized controlled trial. BMC Pediatr. 2017, 17, 54. [Google Scholar] [CrossRef] [Green Version]
- Ghanchi, A.; James, P.T.; Cerami, C. Guts, Germs, and Iron: A Systematic Review on Iron Supplementation, Iron Fortification, and Diarrhea in Children Aged 4–59 Months. Curr. Dev. Nutr. 2019, 3. [Google Scholar] [CrossRef]
- Chen, K.; Zhang, X.; Li, T.; Chen, L.; Wei, X.; Qu, P.; Liu, Y. Effect of vitamin A, vitamin A plus iron and multiple micronutrient-fortified seasoning powder on infectious morbidity of preschool children. Nutrition 2011, 27, 428–434. [Google Scholar] [CrossRef]
- Javaid, N.; Haschke, F.; Pietschnig, B.; Schuster, E.; Huemer, C.; Shebaz, A.; Ganesh, P.; Steffan, I.; Hurrel, R.; Secretin, M.C. Interactions between infections, malnutrition and iron nutritional status in Pakistani infants. A longitudinal study. Acta Paediatr. Scand. Suppl. 1991, 374, 141–150. [Google Scholar] [CrossRef]
- Barth-Jaeggi, T.; Moretti, D.; Kvalsvig, J.; Holding, P.A.; Njenga, J.; Mwangi, A.; Chhagan, M.K.; Lacroix, C.; Zimmermann, M.B. In-home fortification with 2.5 mg iron as NaFeEDTA does not reduce anaemia but increases weight gain: A randomised controlled trial in Kenyan infants: Low-dose iron fortification in Kenyan infants. Matern. Child. Nutr. 2015, 11, 151–162. [Google Scholar] [CrossRef] [Green Version]
- Christofides, A.; Schauer, C.; Sharieff, W.; Zlotkin, S.H. Acceptability of micronutrient sprinkles: A new food-based approach for delivering iron to First Nations and Inuit children in Northern Canada. Chronic Dis. Can. 2005, 26, 114–120. [Google Scholar]
- Lemaire, M.; Islam, Q.S.; Shen, H.; Khan, M.A.; Parveen, M.; Abedin, F.; Haseen, F.; Hyder, Z.; Cook, R.J.; Zlotkin, S.H. Iron-containing micronutrient powder provided to children with moderate-to-severe malnutrition increases hemoglobin concentrations but not the risk of infectious morbidity: A randomized, double-blind, placebo-controlled, noninferiority safety trial. Am. J. Clin. Nutr. 2011, 94, 585–593. [Google Scholar] [CrossRef] [Green Version]
- Paganini, D.; Uyoga, M.A.; Kortman, G.A.M.; Cercamondi, C.I.; Moretti, D.; Barth-Jaeggi, T.; Schwab, C.; Boekhorst, J.; Timmerman, H.M.; Lacroix, C.; et al. Prebiotic galacto-oligosaccharides mitigate the adverse effects of iron fortification on the gut microbiome: A randomised controlled study in Kenyan infants. Gut 2017, 66, 1956–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovannini, M.; Sala, D.; Usuelli, M.; Livio, L.; Francescato, G.; Braga, M.; Radaelli, G.; Riva, E. Double-Blind, Placebo-Controlled Trial Comparing Effects of Supplementation with Two Different Combinations of Micronutrients Delivered as Sprinkles on Growth, Anemia, and Iron Deficiency in Cambodian Infants. J. Pediatr. Gastroenterol. Nutr. 2006, 42, 306–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witzig, R.S.; Black, R.E.; Caulfield, L.E.; Zavaleta, N.; Shankar, A.H.; Richard, S.A. Zinc and Iron Supplementation and Malaria, Diarrhea, and Respiratory Infections in Children in the Peruvian Amazon. Am. J. Trop. Med. Hyg. 2006, 75, 126–132. [Google Scholar] [CrossRef] [Green Version]
- Luabeya, K.-K.A.; Mpontshane, N.; Mackay, M.; Ward, H.; Elson, I.; Chhagan, M.; Tomkins, A.; den Broeck, J.V.; Bennish, M.L. Zinc or Multiple Micronutrient Supplementation to Reduce Diarrhea and Respiratory Disease in South African Children: A Randomized Controlled Trial. PLoS ONE 2007, 2, e541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Chen, X.; Zhang, L.; Luo, H.; Gao, N.; Wang, J.; Fu, G.-Y.; Mao, M. Effect of simultaneous supplementation of vitamin A and iron on diarrheal and respiratory tract infection in preschool children in Chengdu City, China. Nutrition 2013, 29, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Rosado, J.L.; López, P.; Muñoz, E.; Martinez, H.; Allen, L.H. Zinc supplementation reduced morbidity, but neither zinc nor iron supplementation affected growth or body composition of Mexican preschoolers. Am. J. Clin. Nutr. 1997, 65, 13–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelrazik, N.; Al-Haggar, M.; Al-Marsafawy, H.; Abdel-Hadi, H.; Al-Baz, R.; Mostafa, A.-H. Impact of long-term oral iron supplementation in breast-fed infants. Indian J. Pediatr. 2007, 74, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Dewey, K.G.; Domellöf, M.; Cohen, R.J.; Landa Rivera, L.; Hernell, O.; Lönnerdal, B. Iron supplementation affects growth and morbidity of breast-fed infants: Results of a randomized trial in Sweden and Honduras. J. Nutr. 2002, 132, 3249–3255. [Google Scholar] [CrossRef] [Green Version]
- Baqui, A.H.; Zaman, K.; Persson, L.A.; El Arifeen, S.; Yunus, M.; Begum, N.; Black, R.E. Simultaneous weekly supplementation of iron and zinc is associated with lower morbidity due to diarrhea and acute lower respiratory infection in Bangladeshi infants. J. Nutr. 2003, 133, 4150–4157. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.; El Arifeen, S.; Bari, S.; Wahed, M.A.; Rahman, K.M.; Rahman, M.T.; Mahmud, A.B.A.; Begum, N.; Zaman, K.; Baqui, A.H.; et al. Supplementing iron and zinc: Double blind, randomized evaluation of separate or combined delivery. Eur. J. Clin. Nutr. 2010, 64, 153–160. [Google Scholar] [CrossRef]
- Menon, P.; Ruel, M.T.; Loechl, C.U.; Arimond, M.; Habicht, J.-P.; Pelto, G.; Michaud, L. Micronutrient Sprinkles reduce anemia among 9- to 24-mo-old children when delivered through an integrated health and nutrition program in rural Haiti. J. Nutr. 2007, 137, 1023–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soofi, S.; Cousens, S.; Iqbal, S.P.; Akhund, T.; Khan, J.; Ahmed, I.; Zaidi, A.K.M.; Bhutta, Z.A. Effect of provision of daily zinc and iron with several micronutrients on growth and morbidity among young children in Pakistan: A cluster-randomised trial. Lancet. 2013, 382, 29–40. [Google Scholar] [CrossRef]
- Mitra, A.K.; Akramuzzaman, S.M.; Fuchs, G.J.; Rahman, M.M.; Mahalanabis, D. Long-term oral supplementation with iron is not harmful for young children in a poor community of Bangladesh. J. Nutr. 1997, 127, 1451–1455. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative Stress: An Essential Factor in the Pathogenesis of Gastrointestinal Mucosal Diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natoli, M.; Felsani, A.; Ferruzza, S.; Sambuy, Y.; Canali, R.; Scarino, M.L. Mechanisms of defence from Fe(II) toxicity in human intestinal Caco-2 cells. Toxicol. In Vitro 2009, 23, 1510–1515. [Google Scholar] [CrossRef]
- Ferruzza, S.; Scarino, M.L.; Gambling, L.; Natella, F.; Sambuy, Y. Biphasic effect of iron on human intestinal Caco-2 cells: Early effect on tight junction permeability with delayed onset of oxidative cytotoxic damage. Cell Mol. Biol. Noisy Gd. Fr. 2003, 49, 89–99. [Google Scholar]
- Zimmermann, M.B.; Chassard, C.; Rohner, F.; N’Goran, E.K.; Nindjin, C.; Dostal, A.; Utzinger, J.; Ghattas, H.; Lacroix, C.; Hurrell, R.F. The effects of iron fortification on the gut microbiota in African children: A randomized controlled trial in Côte d’Ivoire. Am. J. Clin. Nutr. 2010, 92, 1406–1415. [Google Scholar] [CrossRef]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Finlayson-Trick, E.C.; Fischer, J.A.; Goldfarb, D.M.; Karakochuk, C.D. The Effects of Iron Supplementation and Fortification on the Gut Microbiota: A Review. Gastrointest. Disord. 2020, 2, 327-340. https://0-doi-org.brum.beds.ac.uk/10.3390/gidisord2040030
Finlayson-Trick EC, Fischer JA, Goldfarb DM, Karakochuk CD. The Effects of Iron Supplementation and Fortification on the Gut Microbiota: A Review. Gastrointestinal Disorders. 2020; 2(4):327-340. https://0-doi-org.brum.beds.ac.uk/10.3390/gidisord2040030
Chicago/Turabian StyleFinlayson-Trick, Emma CL, Jordie AJ Fischer, David M Goldfarb, and Crystal D Karakochuk. 2020. "The Effects of Iron Supplementation and Fortification on the Gut Microbiota: A Review" Gastrointestinal Disorders 2, no. 4: 327-340. https://0-doi-org.brum.beds.ac.uk/10.3390/gidisord2040030