
Wheat Breeding, Fertilizers, and Pesticides: Do They Contribute to the Increasing Immunogenic Properties of Modern Wheat?



Abstract
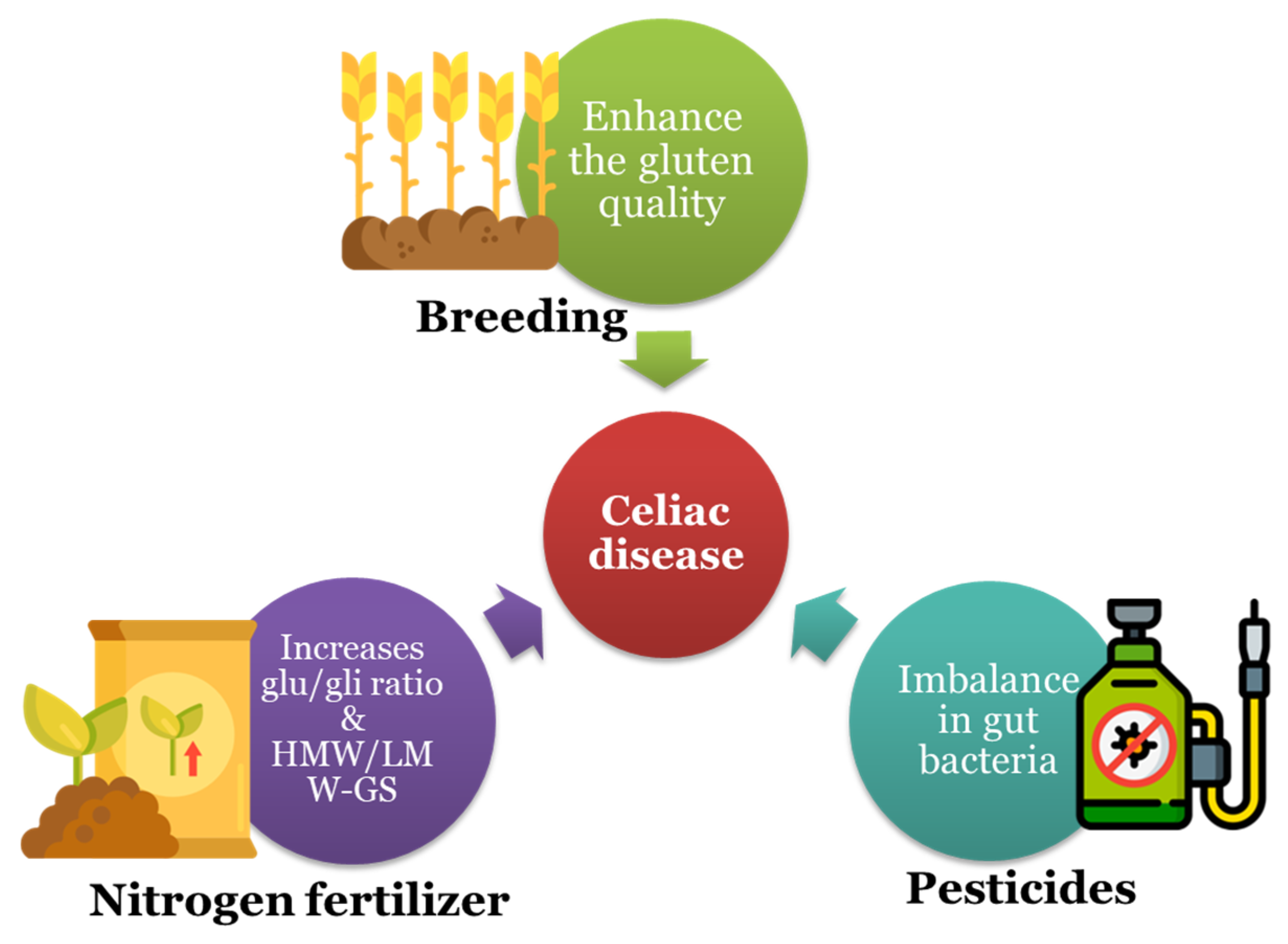
:1. Introduction
2. Wheat Breeding Strategies: Current Challenges
Modern Wheat Cultivars Have a Greater Immunostimulatory Capacity, Linked with Better Seed Quality
3. Nitrogen Fertilizers: Substantially Affects Grain Yield and Glutenin Protein
4. Glyphosate-Based Herbicides (GBHs): Role in Modern Agriculture
Glyphosate: As a Potent Herbicide Associated with Increasing Incidence of CD
5. Methods to Reduce the Immunogenic Properties of Modern Wheat
5.1. RNA Interference (RNAi)-Based Gene Silencing in Wheat
5.2. Developing Overexpressing Transgenic Wheat
5.3. Wheat Deletion Lines
5.4. Genome Editing through CRISPR
6. Developing DNA-Free CRISPR-Edited Wheat to Minimize Regulatory Concerns (Ribonucleoprotein, RNP)
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Verma, A.K.; Singh, A.; Gatti, S.; Lionetti, E.; Galeazzi, T.; Monachesi, C.; Franceschini, E.; Ahuja, V.; Catassi, C.; Makharia, G.K. Validation of a novel single-drop rapid human leukocyte antigen-DQ2/-DQ8 typing method to identify subjects susceptible to celiac disease. JGH Open 2018, 2, 311–316. [Google Scholar] [CrossRef] [PubMed]
- King, J.A.; Jeong, J.; Underwood, F.E.; Quan, J.; Panaccione, N.; Windsor, J.W.; Coward, S.; Debruyn, J.; Ronksley, P.E.; Shaheen, A.-A.; et al. Incidence of Celiac Disease Is Increasing Over Time: A Systematic Review and Meta-analysis. Am. J. Gastroenterol. 2020, 115, 507–525. [Google Scholar] [CrossRef] [PubMed]
- Mustalahti, K.; Catassi, C.; Reunanen, A.; Fabiani, E.; Heier, M.; McMillan, S.; Murray, L.; Metzger, M.H.; Gasparin, M.; Bravi, E.; et al. The prevalence of celiac disease in Europe: Results of a centralized, international mass screening project. Ann. Med. 2010, 42, 587–595. [Google Scholar] [CrossRef]
- Ivarsson, A.; Myléus, A.; Norström, F.; Van Der Pals, M.; Rosén, A.; Högberg, L.; Danielsson, L.; Halvarsson, B.; Hammarroth, S.; Hernell, O.; et al. Prevalence of Childhood Celiac Disease and Changes in Infant Feeding. Pediatrics 2013, 131, e687–e694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatti, S.; Lionetti, E.; Balanzoni, L.; Verma, A.K.; Galeazzi, T.; Gesuita, R.; Scattolo, N.; Cinquetti, M.; Fasano, A.; Catassi, C.; et al. Increased Prevalence of Celiac Disease in School-age Children in Italy. Clin. Gastroenterol. Hepatol. 2019, 18, 596–603. [Google Scholar] [CrossRef]
- Catassi, C.; Kryszak, D.; Bhatti, B.; Sturgeon, C.; Helzlsouer, K.; Clipp, S.L.; Gelfond, D.; Puppa, E.; Sferruzza, A.; Fasano, A. Natural history of celiac disease autoimmunity in a USA cohort followed since 1974. Ann. Med. 2010, 42, 530–538. [Google Scholar] [CrossRef]
- Giersiepen, K.; Lelgemann, M.; Stuhldreher, N.; Ronfani, L.; Husby, S.; Koletzko, S.; Korponay-Szabó, I.R. Accuracy of Diagnostic Antibody Tests for Coeliac Disease in Children: Summary of an evidence report. J. Pediatr. Gastroenterol. Nutr. 2012, 54, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Verma, A.K.; Gatti, S.; Lionetti, E.; Galeazzi, T.; Monachesi, C.; Franceschini, E.; Balanzoni, L.; Scattolo, N.; Cinquetti, M.; Catassi, C. Comparison of Diagnostic Performance of the IgA Anti-tTG Test vs IgA Anti–Native Gliadin Antibodies Test in Detection of Celiac Disease in the General Population. Clin. Gastroenterol. Hepatol. 2018, 16, 1997–1998. [Google Scholar] [CrossRef] [Green Version]
- Bradauskiene, V.; Vaiciulyte-Funk, L.; Martinaitiene, D.; Andruskiene, J.; Verma, A.K.; Lima, J.P.M.; Serin, Y.; Catassi, C. Wheat consumption and prevalence of celiac disease: Correlation from a multilevel analysis. Crit. Rev. Food Sci. Nutr. 2021, 1–15. [Google Scholar] [CrossRef]
- Cummins, A.G.; Roberts-Thomson, I.C. Prevalence of celiac disease in the Asia-Pacific region. J. Gastroenterol. Hepatol. 2009, 24, 1347–1351. [Google Scholar] [CrossRef]
- Lionetti, E.; Catassi, C. Co-localization of gluten consumption and HLA-DQ2 and -DQ8 genotypes, a clue to the history of celiac disease. Dig. Liver Dis. 2014, 46, 1057–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiferaw, B.; Smale, M.; Braun, H.-J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef] [Green Version]
- Hatfield, J.L.; Beres, B.L. Yield Gaps in Wheat: Path to Enhancing Productivity. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Boukid, F.; Vittadini, E.; Prandi, B.; Mattarozzi, M.; Marchini, M.; Sforza, S.; Sayar, R.; Seo, Y.W.; Yacoubi, I.; Mejri, M. Insights into a century of breeding of durum wheat in Tunisia: The properties of flours and starches isolated from landraces, old and modern genotypes. LWT 2018, 97, 743–751. [Google Scholar] [CrossRef]
- De Santis, M.A.; Kosik, O.; Passmore, D.; Flagella, Z.; Shewry, P.R.; Lovegrove, A. Comparison of the dietary fibre composition of old and modern durum wheat (Triticum turgidum spp. durum) genotypes. Food Chem. 2017, 244, 304–310. [Google Scholar] [CrossRef]
- De Santis, M.A.; Giuliani, M.M.; Giuzio, L.; De Vita, P.; Lovegrove, A.; Shewry, P.R.; Flagella, Z. Differences in gluten protein composition between old and modern durum wheat genotypes in relation to 20th century breeding in Italy. Eur. J. Agron. 2017, 87, 19–29. [Google Scholar] [CrossRef]
- Maccaferri, M.; Sanguineti, M.C.; Corneti, S.; Ortega, J.L.A.; Ben Salem, M.; Bort, J.; DeAmbrogio, E.; Del Moral, L.F.G.; Demontis, A.; El-Ahmed, A.; et al. Quantitative Trait Loci for Grain Yield and Adaptation of Durum Wheat (Triticum durum Desf.) Across a Wide Range of Water Availability. Genetics 2008, 178, 489–511. [Google Scholar] [CrossRef] [Green Version]
- Collard, B.; Mackill, D.J. Marker-assisted selection: An approach for precision plant breeding in the twenty-first century. Philos. Trans. R. Soc. B Biol. Sci. 2007, 363, 557–572. [Google Scholar] [CrossRef] [Green Version]
- De Vita, P.; Matteu, L.; Mastrangelo, A.M.; Di Fonzo, N.; Cattivelli, L. Effects of breeding activity on durum wheat traits breed in Italy during the 20th century. Ital. J. Agron. 2007, 2, 451–462. [Google Scholar] [CrossRef] [Green Version]
- Raciti, C.; Doust, M.; Lombardo, G.; Boggini, G.; Pecetti, L. Characterization of durum wheat mediterranean germplasm for high and low molecular weight glutenin subunits in relation with quality. Eur. J. Agron. 2002, 19, 373–382. [Google Scholar] [CrossRef]
- Rossini, F.; Provenzano, M.E.; Sestili, F.; Ruggeri, R. Synergistic Effect of Sulfur and Nitrogen in the Organic and Mineral Fertilization of Durum Wheat: Grain Yield and Quality Traits in the Mediterranean Environment. Agronomy 2018, 8, 189. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Niu, Y.; Ruan, Y.; Depauw, R.M.; Singh, A.K.; Gan, Y. Agronomic Advancement in Tillage, Crop Rotation, Soil Health, and Genetic Gain in Durum Wheat Cultivation: A 17-Year Canadian Story. Agronomy 2018, 8, 193. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.E.; Sands, D.C. The breeder’s dilemma—yield or nutrition? Nat. Biotechnol. 2006, 24, 1078–1080. [Google Scholar] [CrossRef]
- Kucek, L.K.; Veenstra, L.D.; Amnuaycheewa, P.; Sorrells, M.E. A Grounded Guide to Gluten: How Modern Genotypes and Processing Impact Wheat Sensitivity. Compr. Rev. Food Sci. Food Saf. 2015, 14, 285–302. [Google Scholar] [CrossRef]
- Scherf, K.A. Immunoreactive cereal proteins in wheat allergy, non-celiac gluten/wheat sensitivity (NCGS) and celiac disease. Curr. Opin. Food Sci. 2019, 25, 35–41. [Google Scholar] [CrossRef]
- Yue, H.; Jiang, D.; Dai, T.; Qin, X.; Jing, Q.; Cao, W. Effect of nitrogen application rate on content of glutenin macropolymer and high molecular weight glutenin subunits in grains of two winter wheat cultivars. J. Cereal Sci. 2007, 45, 248–256. [Google Scholar] [CrossRef]
- García-Molina, M.D.; Barro, F. Characterization of Changes in Gluten Proteins in Low-Gliadin Transgenic Wheat Lines in Response to Application of Different Nitrogen Regimes. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, C.; Triboi, E. Effects of Temperature and Nitrogen Nutrition on the Grain Composition of Winter Wheat: Effects on Gliadin Content and Composition. J. Cereal Sci. 2000, 32, 45–56. [Google Scholar] [CrossRef]
- Ducsay, L.; Ložek, O. Effect of topdressing with nitrogen on the yield and quality of winter wheat grain. Plant Soil Environ. 2011, 50, 309–314. [Google Scholar] [CrossRef] [Green Version]
- Tea, I.; Genter, T.; Naulet, N.; Boyer, V.; Lummerzheim, M.; Kleiber, D. Effect of Foliar Sulfur and Nitrogen Fertilization on Wheat Storage Protein Composition and Dough Mixing Properties. Cereal Chem. J. 2004, 81, 759–766. [Google Scholar] [CrossRef]
- Varga, B.; Svečnjak, Z.; Jurković, Z.; Pospíšil, M. Quality responses of winter wheat cultivars to nitrogen and fungicide applications in Croatia. Acta Agron. Hung. 2007, 55, 37–48. [Google Scholar] [CrossRef]
- Knapowski, T.; Ralcewicz, M.; Barczak, B.; Kozera, W. Effect of Nitrogen and Zinc Fertilizing on Bread-Making Quality of Spring Triticale Cultivated in Noteć Valley. Pol. J. Environ. Sustain. 2009, 18, 223–232. [Google Scholar]
- Ralcewicz, M.; Knapowski, T.; Kozera, W.; Barczak, B. Technological value of spring wheat of zebra cultivar as related to the way of nitrogen and magnesium application. J. Cent. Eur. Agric. 2009, 10, 223–232. [Google Scholar]
- Zörb, C.; Grover, C.; Steinfurth, D.; Mühling, K.H. Quantitative proteome analysis of wheat gluten as influenced by N and S nutrition. Plant Soil 2009, 327, 225–234. [Google Scholar] [CrossRef]
- Zecevic, V.; Knezevic, D.; Boskovic, J.; Milenkovic, S. Effect of nitrogen and ecological factors on quality of winter triticale cultivars. Genetika 2010, 42, 465–474. [Google Scholar] [CrossRef]
- Fuertes-Mendizábal, T.; Aizpurua, A.; Moro, G.; Estavillo, J. Improving wheat breadmaking quality by splitting the N fertilizer rate. Eur. J. Agron. 2010, 33, 52–61. [Google Scholar] [CrossRef]
- Wan, Y.; Shewry, P.R.; Hawkesford, M. A novel family of γ-gliadin genes are highly regulated by nitrogen supply in developing wheat grain. J. Exp. Bot. 2012, 64, 161–168. [Google Scholar] [CrossRef]
- Fuertes-Mendizábal, T.; Torralba, J.G.; Arregui, L.M.; Gonzalez-Murua, C.; Moro, G.; Estavillo, J.M. Ammonium as sole N source improves grain quality in wheat. J. Sci. Food Agric. 2012, 93, 2162–2171. [Google Scholar] [CrossRef]
- Wojtkowiak, K.; Stpie, A.; Taska, M.; Konopka, I.; Konopka, S. Impact of nitrogen fertilization on the yield and content of protein fractions in spring triticale grain. Afr. J. Agric. Res. 2013, 8, 3778–3783. [Google Scholar] [CrossRef]
- Wan, Y.; Gritsch, C.S.; Hawkesford, M.; Shewry, P.R. Effects of nitrogen nutrition on the synthesis and deposition of the ω-gliadins of wheat. Ann. Bot. 2013, 113, 607–615. [Google Scholar] [CrossRef] [Green Version]
- Rodrighero, M.B.; Caires, E.; Lopes, R.B.; Zielinski, A.; Granato, D.; Demiate, I.M. Wheat technological quality as affected by nitrogen fertilization under a no-till system. Acta Sci. Technol. 2015, 37, 175. [Google Scholar] [CrossRef]
- Klikocka, H.; Cybulska, M.; Barczak, B.; Narolski, B.; Szostak, B.; Kobiałka, A.; Nowak, A.; Wójcik, E. The effect of sulphur and nitrogen fertilization on grain yield and technological quality of spring wheat. Plant Soil Environ. 2016, 62, 230–236. [Google Scholar] [CrossRef] [Green Version]
- Galieni, A.; Stagnari, F.; Visioli, G.; Marmiroli, N.; Speca, S.; Angelozzi, G.; D’Egidio, S.; Pisante, M. Nitrogen fertilisation of durum wheat: A case of study in Mediterranean area during transition to conservation agriculture. Ital. J. Agron. 2015, 11, 12. [Google Scholar] [CrossRef] [Green Version]
- Zhen, S.; Zhou, J.; Deng, X.; Zhu, G.; Cao, H.; Wang, Z.; Yan, Y. Metabolite profiling of the response to high-nitrogen fertilizer during grain development of bread wheat (Triticum aestivum L.). J. Cereal Sci. 2016, 69, 85–94. [Google Scholar] [CrossRef]
- Xue, C.; Erley, G.S.A.; Rücker, S.; Koehler, P.; Obenauf, U.; Mühling, K.H. Late nitrogen application increased protein concentration but not baking quality of wheat. J. Plant Nutr. Soil Sci. 2016, 179, 591–601. [Google Scholar] [CrossRef]
- Zheng, T.; Qi, P.-F.; Cao, Y.-L.; Han, Y.-N.; Ma, H.-L.; Guo, Z.-R.; Wang, Y.; Qiao, Y.-Y.; Hua, S.-Y.; Yu, H.-Y.; et al. Mechanisms of wheat (Triticum aestivum) grain storage proteins in response to nitrogen application and its impacts on processing quality. Sci. Rep. 2018, 8, 11928. [Google Scholar] [CrossRef]
- Litke, L.; Gaile, Z. Effect of nitrogen fertilization on winter wheat yield and yield quality. Agron. Res. 2018, 16, 500. [Google Scholar] [CrossRef]
- Cho, S.-W.; Kang, C.-S.; Kang, T.-G.; Cho, K.-M.; Park, C.S. Influence of different nitrogen application on flour properties, gluten properties by HPLC and end-use quality of Korean wheat. J. Integr. Agric. 2018, 17, 982–993. [Google Scholar] [CrossRef]
- Dubois, B.; Bertin, P.; Hautier, L.; Muhovski, Y.; Escarnot, E.; Mingeot, D. Genetic and environmental factors affecting the expression of α-gliadin canonical epitopes involved in celiac disease in a wide collection of spelt (Triticum aestivum ssp. spelta) cultivars and landraces. BMC Plant Biol. 2018, 18, 262. [Google Scholar] [CrossRef] [Green Version]
- Guardia, G.; Sanz-Cobena, A.; Sanchez-Martin, L.; Fuertes-Mendizábal, T.; Gonzalez-Murua, C.; Alvarez, J.M.A.; Chadwick, D.; Vallejo, A. Urea-based fertilization strategies to reduce yield-scaled N oxides and enhance bread-making quality in a rainfed Mediterranean wheat crop. Agric. Ecosyst. Environ. 2018, 265, 421–431. [Google Scholar] [CrossRef]
- Klikocka, H.; Marks, M.; Marx, M. Sulphur and Nitrogen Fertilization as a Potential Means of Agronomic Biofortification to Improve the Content and Uptake of Microelements in Spring Wheat Grain DM. J. Chem. 2018, 2018, 9326820. [Google Scholar] [CrossRef]
- Xue, C.; Matros, A.; Mock, H.-P.; Mühling, K.H. Protein Composition and Baking Quality of Wheat Flour as Affected by Split Nitrogen Application. Front. Plant Sci. 2019, 10, 642. [Google Scholar] [CrossRef]
- Eser, A.; Kassai, K.; Kato, H.; Kunos, V.; Tarnava, A.; Jolánkai, M. Impact of nitrogen topdressing on the quality parameters of winter wheat (Triticum aestivum L.) yield. Acta Aliment. 2020, 49, 244–253. [Google Scholar] [CrossRef]
- Zhen, S.; Deng, X.; Xu, X.; Liu, N.; Zhu, D.; Wang, Z.; Yan, Y. Effect of high-nitrogen fertilizer on gliadin and glutenin subproteomes during kernel development in wheat (Triticum aestivum L.). Crop. J. 2020, 8, 38–52. [Google Scholar] [CrossRef]
- Song, L.; Li, L.; Zhao, L.; Liu, Z.; Li, X. Effects of Nitrogen Application in the Wheat Booting Stage on Glutenin Polymerization and Structural–Thermal Properties of Gluten with Variations in HMW-GS at the Glu-D1 Locus. Foods 2020, 9, 353. [Google Scholar] [CrossRef]
- Pepo, P.; Sipos, P.; Győri, Z. Effects of fertilizer application on the baking quality of winter wheat varieties in a long term experiment under continental climatic conditions in Hungary. Cereal Res. Commun. 2005, 33, 825–832. [Google Scholar] [CrossRef]
- Turhan, D.; Güngördü, A.; Ozmen, M. Developmental and lethal effects of glyphosate and a glyphosate-based product on Xenopus laevis embryos and tadpoles. Bull. Environ. Contam. Toxicol. 2020, 104, 173–179. [Google Scholar] [CrossRef]
- Mertens, M.; Höss, S.; Neumann, G.; Afzal, J.; Reichenbecher, W. Glyphosate, a chelating agent—relevant for ecological risk assessment? Environ. Sci. Pollut. Res. 2018, 25, 5298–5317. [Google Scholar] [CrossRef] [Green Version]
- Van Bruggen, A.H.C.; He, M.M.; Shin, K.; Mai, V.; Jeong, K.C.; Finckh, M.R.; Morris, J.G., Jr. Environmental and health effects of the herbicide glyphosate. Sci. Total Environ. 2018, 616, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Blot, N.; Veillat, L.; Rouzé, R.; Delatte, H. Glyphosate, but not its metabolite AMPA, alters the honeybee gut microbiota. PLoS ONE 2019, 14, e0215466. [Google Scholar] [CrossRef] [PubMed]
- Maddalon, A.; Galbiati, V.; Colosio, C.; Mandić-Rajčević, S.; Corsini, E. Glyphosate-based herbicides: Evidence of immune-endocrine alteration. Toxicology 2021, 459, 152851. [Google Scholar] [CrossRef] [PubMed]
- Gorbach, S. Microbiology of the Gastrointestinal Tract. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch: Galveston, TX, USA, 1996. [Google Scholar]
- Sanz, Y.; De Palma, G.; Laparra, M. Unraveling the Ties between Celiac Disease and Intestinal Microbiota. Int. Rev. Immunol. 2011, 30, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; De Angelis, M.; De Pasquale, I.; Ndagijimana, M.; Vernocchi, P.; Ricciuti, P.; Gagliardi, F.; Laghi, L.; Crecchio, C.; Guerzoni, M.E.; et al. Duodenal and faecal microbiota of celiac children: Molecular, phenotype and metabolome characterization. BMC Microbiol. 2011, 11, 219. [Google Scholar] [CrossRef] [Green Version]
- Collado, M.C.; Calabuig, M.S.Y. Differences between the Faecal Microbiota of Coeliac Infants and Healthy Controls. Curr. Issues Intest Microbiol. 2007, 8, 9–14. [Google Scholar]
- Francavilla, R.; De Angelis, M.; Rizzello, C.G.; Cavallo, N.; Bello, F.D.; Gobbetti, M. Selected Probiotic Lactobacilli Have the Capacity To Hydrolyze Gluten Peptides during Simulated Gastrointestinal Digestion. Appl. Environ. Microbiol. 2017, 83, e00376-17. [Google Scholar] [CrossRef] [Green Version]
- Greco, L.; Gobbetti, M.; Auricchio, R.; Di Mase, R.; Landolfo, F.; Paparo, F.; Di Cagno, R.; De Angelis, M.; Rizzello, C.G.; Cassone, A.; et al. Safety for Patients With Celiac Disease of Baked Goods Made of Wheat Flour Hydrolyzed During Food Processing. Clin. Gastroenterol. Hepatol. 2011, 9, 24–29. [Google Scholar] [CrossRef]
- Tang, Q.; Tang, J.; Ren, X.; Li, C. Glyphosate exposure induces inflammatory responses in the small intestine and alters gut microbial composition in rats. Environ. Pollut. 2020, 261, 114129. [Google Scholar] [CrossRef]
- Mao, Q.; Manservisi, F.; Panzacchi, S.; Mandrioli, D.; Menghetti, I.; Vornoli, A.; Bua, L.; Falcioni, L.; Lesseur, C.; Chen, J.; et al. The Ramazzini Institute 13-week pilot study on glyphosate and Roundup administered at human-equivalent dose to Sprague Dawley rats: Effects on the microbiome. Environ. Health 2018, 17, 50. [Google Scholar] [CrossRef] [Green Version]
- Lerner, A.; Arleevskaya, M.; Schmiedl, A.; Matthias, T. Microbes and Viruses Are Bugging the Gut in Celiac Disease. Are They Friends or Foes? Front. Microbiol. 2017, 8, 1392. [Google Scholar] [CrossRef]
- Ou, G.; Hedberg, M.; Hörstedt, P.; Baranov, V.; Forsberg, G.; Drobni, M.; Sandström, O.; Wai, S.N.; Johansson, I.; Hammarström, M.-L.; et al. Proximal Small Intestinal Microbiota and Identification of Rod-Shaped Bacteria Associated With Childhood Celiac Disease. Am. J. Gastroenterol. 2009, 104, 3058–3067. [Google Scholar] [CrossRef]
- Zamakhchari, M.; Wei, G.; Dewhirst, F.; Lee, J.; Schuppan, D.; Oppenheim, F.G.; Helmerhorst, E.J. Identification of Rothia Bacteria as Gluten-Degrading Natural Colonizers of the Upper Gastro-Intestinal Tract. PLoS ONE 2011, 6, e24455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.H.; Han, D.S.; Oh, Y.-H.; Lee, A.-R.; Lee, Y.-R.; Eun, C.S. Role of Fusobacteria in the serrated pathway of colorectal carcinogenesis. Sci. Rep. 2016, 6, 25271. [Google Scholar] [CrossRef]
- Kittana, H.; Gomes-Neto, J.C.; Heck, K.; Geis, A.L.; Muñoz, R.R.S.; Cody, L.A.; Schmaltz, R.J.; Bindels, L.B.; Sinha, R.; Hostetter, J.; et al. Commensal Escherichia coli Strains Can Promote Intestinal Inflammation via Differential Interleukin-6 Production. Front. Immunol. 2018, 9, 2318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koliada, A.; Syzenko, G.; Moseiko, V.; Budovska, L.; Puchkov, K.; Perederiy, V.; Gavalko, Y.; Dorofeyev, A.; Romanenko, M.; Tkach, S.; et al. Association between body mass index and Firmicutes/Bacteroidetes ratio in an adult Ukrainian population. BMC Microbiol. 2017, 17, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rustgi, S.; Shewry, P.; Brouns, F.; Deleu, L.J.; Delcour, J.A. Wheat Seed Proteins: Factors Influencing Their Content, Composition, and Technological Properties, and Strategies to Reduce Adverse Reactions. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1751–1769. [Google Scholar] [CrossRef] [Green Version]
- Altenbach, S.B.; Tanaka, C.K.; Allen, P.V. Quantitative proteomic analysis of wheat grain proteins reveals differential effects of silencing of omega-5 gliadin genes in transgenic lines. J. Cereal Sci. 2014, 59, 118–125. [Google Scholar] [CrossRef]
- Altenbach, S.B.; Tanaka, C.K.; Seabourn, B.W. Silencing of omega-5 gliadins in transgenic wheat eliminates a major source of environmental variability and improves dough mixing properties of flour. BMC Plant Biol. 2014, 14, 393. [Google Scholar] [CrossRef] [Green Version]
- Blechl, A.; Beecher, B.; Vensel, W.; Tanaka, C.; Altenbach, S. RNA interference targeting rye secalins alters flour protein composition in a wheat variety carrying a 1BL.1RS translocation. J. Cereal Sci. 2016, 68, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Barro, F.; Iehisa, J.C.M.; Gimenez, M.J.; García-Molina, M.D.; Ozuna, C.V.; Comino, I.; Sousa, C.; Gil Humanes, J. Targeting of prolamins by RNAi in bread wheat: Effectiveness of seven silencing-fragment combinations for obtaining lines devoid of coeliac disease epitopes from highly immunogenic gliadins. Plant Biotechnol. J. 2015, 14, 986–996. [Google Scholar] [CrossRef] [Green Version]
- Altenbach, S.B.; Allen, P.V. Transformation of the US bread wheat ‘Butte 86’ and silencing of omega-5 gliadin genes. GM Crop. 2011, 2, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Altenbach, S.B.; Tanaka, C.K.; Pineau, F.; Lupi, R.; Drouet, M.; Beaudouin, E.; Morisset, M.; Denery-Papini, S. Assessment of the Allergenic Potential of Transgenic Wheat (Triticum aestivum) with Reduced Levels of ω5-Gliadins, the Major Sensitizing Allergen in Wheat-Dependent Exercise-Induced Anaphylaxis. J. Agric. Food Chem. 2015, 63, 9323–9332. [Google Scholar] [CrossRef] [PubMed]
- Becker, D.; Wieser, H.; Koehler, P.; Folck, A.; Mühling, K.H.; Zörb, C. Protein composition and techno-functional properties of transgenic wheat with reduced α-gliadin content obtained by RNA interference. J. Appl. Bot. Food Quality 2012, 85, 23–33. [Google Scholar]
- Gil-Humanes, J.; Piston, F.; Tollefsen, S.; Sollid, L.M.; Barro, F. Effective shutdown in the expression of celiac disease-related wheat gliadin T-cell epitopes by RNA interference. Proc. Natl. Acad. Sci. USA 2010, 107, 17023–17028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil-Humanes, J.; Piston, F.; Gimenez, M.J.; Martín, A.; Barro, F. The Introgression of RNAi Silencing of γ-Gliadins into Commercial Lines of Bread Wheat Changes the Mixing and Technological Properties of the Dough. PLoS ONE 2012, 7, e45937. [Google Scholar] [CrossRef] [Green Version]
- Gil-Humanes, J.; Pistón, F.; Barro, F.; Rosell, C.M. The Shutdown of Celiac Disease-Related Gliadin Epitopes in Bread Wheat by RNAi Provides Flours with Increased Stability and Better Tolerance to Over-Mixing. PLoS ONE 2014, 9, e91931. [Google Scholar] [CrossRef] [Green Version]
- Gil-Humanes, J.; Pistón, F.; Altamirano-Fortoul, R.; Real, A.; Comino, I.; Sousa, C.; Rosell, C.M.; Barro, F. Reduced-Gliadin Wheat Bread: An Alternative to the Gluten-Free Diet for Consumers Suffering Gluten-Related Pathologies. PLoS ONE 2014, 9, e90898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil Humanes, J.; Piston, F.; Hernando, A.; Alvarez, J.B.; Shewry, P.R.; Barro, F. Silencing of γ-gliadins by RNA interference (RNAi) in bread wheat. J. Cereal Sci. 2008, 48, 565–568. [Google Scholar] [CrossRef]
- Pistón, F.; Gil Humanes, J.; Rodríguez-Quijano, M.; Barro, F. Down-Regulating γ-Gliadins in Bread Wheat Leads to Non-Specific Increases in Other Gluten Proteins and Has No Major Effect on Dough Gluten Strength. PLoS ONE 2011, 6, e24754. [Google Scholar] [CrossRef] [Green Version]
- Osorio, C.E.; Wen, N.; Mejías, J.; Liu, B.; Reinbothe, S.; Von Wettstein, D.; Rustgi, S. Development of wheat genotypes expressing a glutamine-specific endoprotease from barley and a prolyl endopeptidase from Flavobacterium meningosepticum or Pyrococcus furiosus as a potential remedy to celiac disease. Funct. Integr. Genom. 2018, 19, 123–136. [Google Scholar] [CrossRef]
- Li, D.; Jin, H.; Zhang, K.; Wang, Z.; Wang, F.; Zhao, Y.; Huo, N.; Liu, X.; Gu, Y.Q.; Wang, D.; et al. Analysis of the Gli-D2 locus identifies a genetic target for simultaneously improving the breadmaking and health-related traits of common wheat. Plant J. 2018, 95, 414–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.-W.; Li, D.; Wang, J.; Zhao, Y.; Wang, Z.; Yue, G.; Liu, X.; Qin, H.; Zhang, K.; Dong, L.; et al. Genome-wide analysis of complex wheat gliadins, the dominant carriers of celiac disease epitopes. Sci. Rep. 2017, 7, 44609. [Google Scholar] [CrossRef] [Green Version]
- Camerlengo, F.; Sestili, F.; Silvestri, M.; Colaprico, G.; Margiotta, B.; Ruggeri, R.; Lupi, R.; Masci, S.; Lafiandra, D. Production and molecular characterization of bread wheat lines with reduced amount of α-type gliadins. BMC Plant Biol. 2017, 17, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waga, J.; Skoczowski, A. Development and characteristics of ω-gliadin-free wheat genotypes. Euphytica 2013, 195, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Waga, J.; Zientarski, J.; Szaleniec, M.; Obtułowicz, K.; Dyga, W.; Skoczowski, A. Null Alleles in Gliadin Coding Loci and Wheat Allergenic Properties. Am. J. Plant Sci. 2013, 4, 160–168. [Google Scholar] [CrossRef] [Green Version]
- León, S.S.; Gil Humanes, J.; Ozuna, C.V.; Gimenez, M.J.; Sousa, C.; Voytas, D.; Barro, F. Low-gluten, nontransgenic wheat engineered with CRISPR/Cas9. Plant Biotechnol. J. 2017, 16, 902–910. [Google Scholar] [CrossRef]
- Jouanin, A.; Schaart, J.G.; Boyd, L.A.; Cockram, J.; Leigh, F.; Bates, R.; Wallington, E.J.; Visser, R.G.F.; Smulders, M.J.M. Outlook for coeliac disease patients: Towards bread wheat with hypoimmunogenic gluten by gene editing of α- and γ-gliadin gene families. BMC Plant Biol. 2019, 19, 333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zörb, C.; Becker, D.; Hasler, M.; Muehling, K.H.; Gödde, V.; Niehaus, K.; Geilfus, C.-M. Silencing of the sulfur rich α-gliadin storage protein family in wheat grains (Triticum aestivum L.) causes no unintended side-effects on other metabolites. Front. Plant Sci. 2013, 4, 369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altenbach, S.B.; Chang, H.-C.; Simon-Buss, A.; Mohr, T.; Huo, N.; Gu, Y.Q. Exploiting the reference genome sequence of hexaploid wheat: A proteomic study of flour proteins from the cultivar Chinese Spring. Funct. Integr. Genom. 2020, 20, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Altenbach, S.B.; Chang, H.-C.; Yu, X.B.; Seabourn, B.W.; Green, P.H.; Alaedini, A. Elimination of Omega-1,2 Gliadins From Bread Wheat (Triticum aestivum) Flour: Effects on Immunogenic Potential and End-Use Quality. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Chai, J.-F.; Wang, H.-B.; Ma, X.-Y.; Zhang, C.-M.; Dong, F.-S. Effect of ω-Secalin Gene Silencing on Processing Quality of Wheat 1B/1R Translocation Line. Acta Agron. Sin. 2016, 42. [Google Scholar] [CrossRef]
- Chai, J.-F.; Zhang, C.-M.; Ma, X.-Y.; Wang, H.-B. Molecular identification of ω-secalin gene expression activity in a wheat 1B/1R translocation cultivar. J. Integr. Agric. 2016, 15, 2712–2718. [Google Scholar] [CrossRef] [Green Version]
- García-Molina, M.D.; Giménez, M.J.; León, S.S.; Barro, F. Gluten Free Wheat: Are We There? Nutrients 2019, 11, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. CRISPR/Cas Genome Editing and Precision Plant Breeding in Agriculture. Annu. Rev. Plant Biol. 2019, 70, 667–697. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Joshi, R.K.; Zhao, K. Base editing in crops: Current advances, limitations and future implications. Plant Biotechnol. J. 2019, 18, 20–31. [Google Scholar] [CrossRef]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.K.; Mandal, S.; Tiwari, A.; Monachesi, C.; Catassi, G.N.; Srivastava, A.; Gatti, S.; Lionetti, E.; Catassi, C. Current Status and Perspectives on the Application of CRISPR/Cas9 Gene-Editing System to Develop a Low-Gluten, Non-Transgenic Wheat Variety. Foods 2021, 10, 2351. [Google Scholar] [CrossRef]
- Ishii, T.; Araki, M. A future scenario of the global regulatory landscape regarding genome-edited crops. GM Crop. Food 2017, 8, 44–56. [Google Scholar] [CrossRef] [Green Version]
- Wolt, J.D.; Wang, K.; Yang, B. The Regulatory Status of Genome-edited Crops. Plant Biotechnol. J. 2015, 14, 510–518. [Google Scholar] [CrossRef] [Green Version]
- Callaway, E. CRISPR plants now subject to tough GM laws in European Union. Nature 2018, 560, 16. [Google Scholar] [CrossRef]
- Sprink, T.; Eriksson, D.; Schiemann, J.; Hartung, F. Regulatory hurdles for genome editing: Process- vs. product-based approaches in different regulatory contexts. Plant Cell Rep. 2016, 35, 1493–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Pribil, M.; Palmgren, M.; Gao, C. A CRISPR way for accelerating improvement of food crops. Nat. Food 2020, 1, 200–205. [Google Scholar] [CrossRef]
- He, Y.; Zhao, Y. Technological breakthroughs in generating transgene-free and genetically stable CRISPR-edited plants. aBIOTECH 2019, 1, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Abe, F.; Haque, E.; Hisano, H.; Tanaka, T.; Kamiya, Y.; Mikami, M.; Kawaura, K.; Endo, M.; Onishi, K.; Hayashi, T.; et al. Genome-Edited Triple-Recessive Mutation Alters Seed Dormancy in Wheat. Cell Rep. 2019, 28, 1362–1369.e4. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Liu, H.; Du, L.; Ye, X. Generation of marker-free transgenic hexaploid wheat via an Agrobacterium-mediated co-transformation strategy in commercial Chinese wheat varieties. Plant Biotechnol. J. 2016, 15, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liang, Z.; Zong, Y.; Wang, Y.; Liu, J.; Chen, K.; Qiu, J.-L.; Gao, C. Efficient and transgene-free genome editing in wheat through transient expression of CRISPR/Cas9 DNA or RNA. Nat. Commun. 2016, 7, 12617. [Google Scholar] [CrossRef] [Green Version]
- Zong, Y.; Wang, Y.; Li, C.; Zhang, R.; Chen, K.; Ran, Y.; Qiu, J.-L.; Wang, D.; Gao, C. Precise base editing in rice, wheat and maize with a Cas9-cytidine deaminase fusion. Nat. Biotechnol. 2017, 35, 438–440. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zong, Y.; Wang, Y.; Jin, S.; Zhang, D.; Song, Q.; Zhang, R.; Gao, C. Expanded base editing in rice and wheat using a Cas9-adenosine deaminase fusion. Genome Biol. 2018, 19, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Liu, J.; Chai, Z.; Chen, S.; Bai, Y.; Zong, Y.; Chen, K.; Li, J.; Jiang, L.; Gao, C. Generation of herbicide tolerance traits and a new selectable marker in wheat using base editing. Nat. Plants 2019, 5, 480–485. [Google Scholar] [CrossRef]
- Liang, Z.; Chen, K.; Li, T.; Zhang, Y.; Wang, Y.; Zhao, Q.; Liu, J.; Zhang, H.; Liu, C.; Ran, Y.; et al. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nat. Commun. 2017, 8, 14261. [Google Scholar] [CrossRef]


{kind=link}
| S. No | Genotype | Effect of Nitrogen Application | Year | Reference |
|---|---|---|---|---|
| 1. | Triticum aestivum L. cv. Neepawa | Proteins and gliadins increased in the flour | 2000 | [28] |
| 2. | T. aestivum L. cv. Astella (Winter wheat) | Increase in wet gluten concentrations and total protein | 2004 | [29] |
| 3. | T. aestivum L. cv. Soissons (Winter wheat) | Gliadins, LMW-GS, and HMW-GS levels increase significantly | 2004 | [30] |
| 4. | T. aestivum L. cv. Marija, Žitarka, Srpanjka, Soissons, Renan and Kuna | Gluten content increased in the wheat grain | 2007 | [31] |
| 5. | T. aestivum L. Kargo (Spring triticale) | Highest significant amount of wet gluten | 2009 | [32] |
| 6. | T. aestivum L. cv. Zebra (Spring triticale) | Increase in grain yield and gluten content | 2009 | [33] |
| 7. | T. aestivum L. cv. Privileg (winter wheat) | ω-gliadins and HMW glutenin were increased by high levels of N | 2009 | [34] |
| 8. | T. aestivum L. cv. Trijumf and Favorit (winter triticale) | Wet gluten content increased | 2010 | [35] |
| 9. | T. aestivum L. cv. Soissons (Winter wheat) | Increase in the HMW glutenin subunits (HMW-GS) | 2010 | [36] |
| 10. | T. aestivum L. cv. Cordiale, Hereward, Istabraq, Malacca, Marksman and Xi 19 | γ-gliadin gene expression increased | 2012 | [37] |
| 11. | T. aestivum L. cv. Cezanne | Improves grain yield, quality as well gliadin and gliadin/protein ratio | 2013 | [38] |
| 12. | T. aestivum L. cv. Andrus (Spring triticale) | Increase in albumins with globulins and ω and α/β prolamins in grain | 2013 | [39] |
| 13. | T. aestivum L. cv. Cordiale, Hereward, Istabraq, Malacca, Marksman and Xi 19 | increased accumulation of ω-5 and ω-2 gliadins | 2014 | [40] |
| 14. | T. aestivum L. cv. Quartzo | Increasing kernel protein content and gluten content of the flour | 2015 | [41] |
| 15. | T. aestivum L. cv. Tybalt (spring wheat) | Increase in gluten content | 2016 | [42] |
| 16. | T. turgidum L. subsp. durum (Desf.) Husn., cv. Achille | Increase in GPC, total gluten, and gluten fractions | 2016 | [43] |
| 17. | T. aestivum L. cv. Zhongmai 175 | High-nitrogen fertilizer enhanced grain protein and gluten contents | 2016 | [44] |
| 18. | T. aestivum L. cv. Tobak and JB Asano | Enhances GPC and the relative abundance of HMW-GS | 2016 | [45] |
| 19. | T. aestivum L. cv. Shumai 969 (SM969), Shumai 482 (SM482), Chuannong 16 (CN16) and Mianmai 51 (MM51) | HMW-GSs and ω-gliadin, greatly responded to increased nitrogen | 2018 | [46] |
| 20. | T. aestivum L. cv. Skagen | Higher average gluten content in grain | 2018 | [47] |
| 21. | T. aestivum L. (Korean wheat cultivars) | Increases in gluten concentrations in flour, α + β-gliadin and decreases in ω and γ-gliadin concentrations | 2018 | [48] |
| 22. | T. aestivum ssp. Spelta | Increase in α-gliadin epitopes related to CD | 2018 | [49] |
| 23. | T. aestivum L. cv. Ingenio and Marcopolo | Increase in total gluten proteins (gliadins and glutenins) | 2018 | [50] |
| 24. | T. aestivum L. cv. Tybalt (spring wheat) | Increase in gluten content | 2018 | [51] |
| 25. | T. aestivum L. cv. Tobak and JB Asano | Increase in globulins, LMW-GS, α-, and γ-gliadins | 2019 | [52] |
| 26. | T. aestivum L. (Winter wheat) | Gluten content was significantly increasing | 2020 | [53] |
| 27. | T. aestivum L. Zhongmai 175 (Chinese winter wheat) | Increases in gluten, total gliadin, and glutenin content | 2020 | [54] |
| 28. | T. aestivum L. cv. Xinong 1718, Fa 710 and Fa 790 (NILs) | Increased the accumulation of total protein, glutenin, glu/gli ratio, HMW/LMW-GS, and GMP | 2020 | [55] |
| S. No. | Cultivar/Transgenic Lines | Target Gene | Output | References |
|---|---|---|---|---|
| RNA interference | ||||
| 1. | Triticum aestivum cv. Chinese Spring; T. aestivum cv. Butte 86 | ω-1,2 gliadins | ω-1,2 gliadins were effectively silenced and one transgenic line showed improved dough functionality. This reduced the immunogenicity of flour protein to antibodies from CD patients. | [78,79] |
| 2. | T. aestivum cv. Bobwhite (BW) | Secalins in 1BL/1RS wheat line | Multiple secalins and closely related ω-gliadins showed substantial reductions; HMW-GSs and α-gliadins were elevated; and dough functioning for two transgenic lines was improved. | [80] |
| 3. | T. aestivum cv. Bobwhite 208 (BW208) | α-, γ- and ω-gliadins, LMW-GSs | Gliadins and LMW-GS have been effectively silenced; there are many lines that lack CD epitopes and therefore are much less immunogenic due to the absence of α- and ω-gliadins and overall protein and starch contents of the grains are unaltered in the transgenic lines | [81] |
| 4. | T. aestivum cv. Butte 86, Transgenic lines SA-8-35b-5 and SA-8-45a-2 | ω-5 gliadins | In one line, the content of ω-5 gliadin was reduced by 80%, while in another line, it was completely removed. WDEIA patients’ responsiveness to the IgE antibody was significantly decreased; enhancing the characteristics of dough functioning. | [78,82,83] |
| 5. | T. aestivum cv. Florida | α-Gliadins | In two transgenic lines, the content of α-gliadin was significantly decreased, but this was accounted for by higher amounts of γ- and ω-gliadins, HMW-GSs, and other grain proteins; no substantial impact on flour functioning was found. | [84] |
| 6. | T. aestivum cv. Bobwhite (BW2003 and BW208); Transgenic line A1152, G613, G622, G626, G845, G658 and G664 | α-, ω- and/or γ-gliadins | Strongly reduced gliadin expression by 85.6%; transgenic wheat lines with extremely low toxicity for CD patients produced; and several of the transgenic lines demonstrated enhanced end-use quality and nutritional value. | [85,86,87,88] |
| 7. | T. aestivum cv. Bobwhite 208 (BW208), Lines A1152, A1158, A1406, C655, C657, D445, D623, C217 and D598 | γ-Gliadins | Increased levels of α- and ω-gliadins together with decreased expression of γ-gliadins in nine transgenic lines, six transgenic lines exhibited increased SDS-sedimentation values. | [89,90] |
| Overexpressing transgenics lines | ||||
| 8. | T. aestivum cv. Brundage 96 | α-Gliadins | Transgenic wheat expressing engineered glutenase, which were capable of degrading the CD epitopes carried by α-gliadins under simulated gastrointestinal condition | [91] |
| Deletion wheat lines | ||||
| 9. | Xiaoyan 81, Xy81, DLGliD2, Zhengmai 366 and ZDLGliD2. | α-Gliadins | Six deletion lines deficient in the gliadin chromosomal loci were developed; the line lacking Gli-D2 demonstrated better dough functioning and breadmaking quality, with a substantial reduction in the number of CD epitopes. | [92,93] |
| 10. | T. aestivum cv. Pegaso (bread wheat) | α-Gliadins | Three deletion lines lacking Gli-A2, Gli-D2, or Gli-A2/Gli-D2 were created; significant reductions in α-gliadin expression were found in all three lines; the 33-mer peptide containing gliadins was not identified in the lines lacking Gli-D2 or Gli-A2/Gli-D2. | [94] |
| 11. | T. aestivum 39N, winter wheat (contained inactive genes in ω-gliadin loci that code no proteins) | ω-1,2 and ω-5 gliadins | Gliadin immunoreactivity was reduced by around 30% in wheat genotypes missing both ω-1,2 and ω-5 gliadins, while gluten functioning was enhanced. | [95] |
| 12. | Hybrid wheat (spelt wheat, cultivar Oberkummler Rotkorn and Polish high quality wheat line LAD 480) | α-, γ- and ω-gliadins | Three deletion lines were generated, each with a null allele at Gli-D1, Gli-B1, or Gli-B2; immunoreactivity of flour proteins was decreased by 6–18% in the deletion lines. | [96] |
| Genome editing through CRISPR/Cas9 | ||||
| 13. | T. aestivum cv. Fielder; T. aestivum cv. BW208 and THA53 (bread wheat), and T. aestivum, cv Don Pedro (DP, durum wheat) | α-Gliadins | Using CRISPR/Cas9 genome editing, simultaneous mutation of several α-gliadin genes was used to successfully lower the level of immunoreactivity for gluten proteins by up to 85%. | [97,98] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mandal, S.; Verma, A.K. Wheat Breeding, Fertilizers, and Pesticides: Do They Contribute to the Increasing Immunogenic Properties of Modern Wheat? Gastrointest. Disord. 2021, 3, 247-264. https://0-doi-org.brum.beds.ac.uk/10.3390/gidisord3040023
Mandal S, Verma AK. Wheat Breeding, Fertilizers, and Pesticides: Do They Contribute to the Increasing Immunogenic Properties of Modern Wheat? Gastrointestinal Disorders. 2021; 3(4):247-264. https://0-doi-org.brum.beds.ac.uk/10.3390/gidisord3040023
Chicago/Turabian StyleMandal, Sayanti, and Anil K. Verma. 2021. "Wheat Breeding, Fertilizers, and Pesticides: Do They Contribute to the Increasing Immunogenic Properties of Modern Wheat?" Gastrointestinal Disorders 3, no. 4: 247-264. https://0-doi-org.brum.beds.ac.uk/10.3390/gidisord3040023