
Two Types of PPARγ Ligands Identified in the Extract of Artemisia campestris

 , and
, and 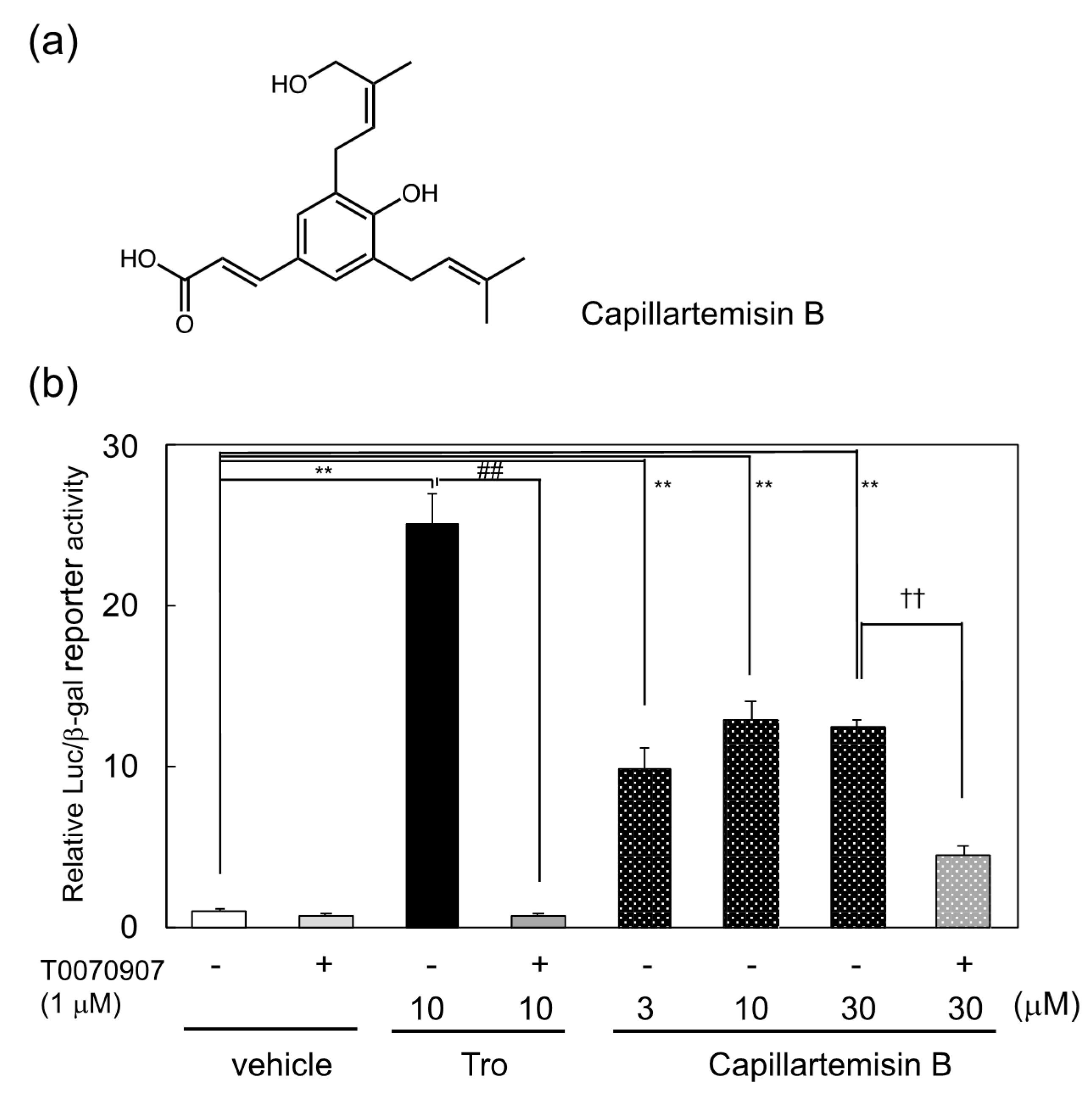{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical Reagents and Instruments
2.2. Preparation of Plant Extracts and Purification of PPARγ Ligands
2.3. Cell Culture
2.4. PPARγ Reporter Assay
2.5. Analysis of Lipid Accumulation in the Differentiated 3T3-L1 Cells
2.6. Docking Study of Eupatilin to PPARγ LBD
2.7. Statistics
3. Results
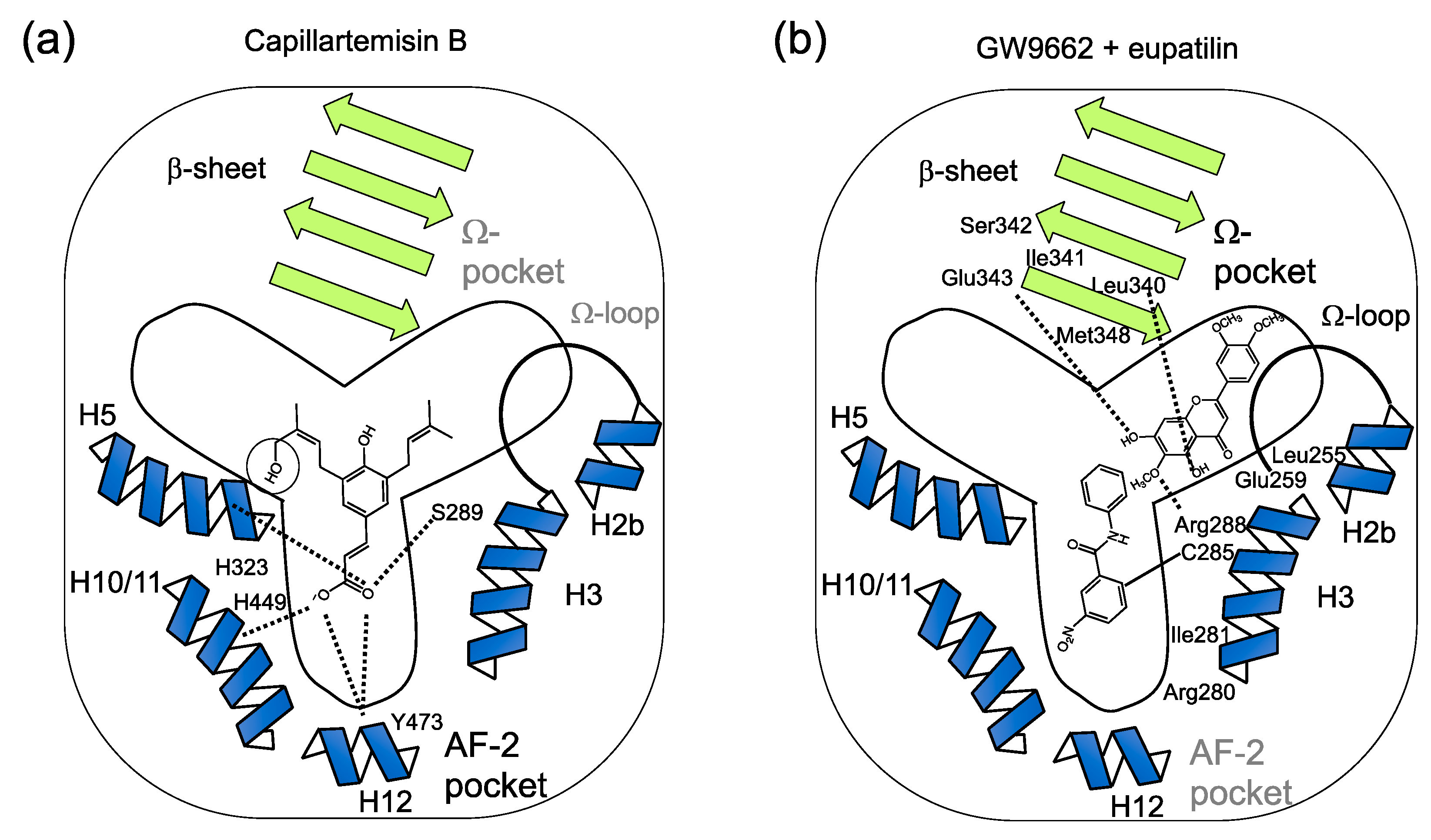
3.1. Identification of Capillartemisin B as a PPARγ Agonist in the Methanol Extract of Artemisia Campestris and Its Biological Activity
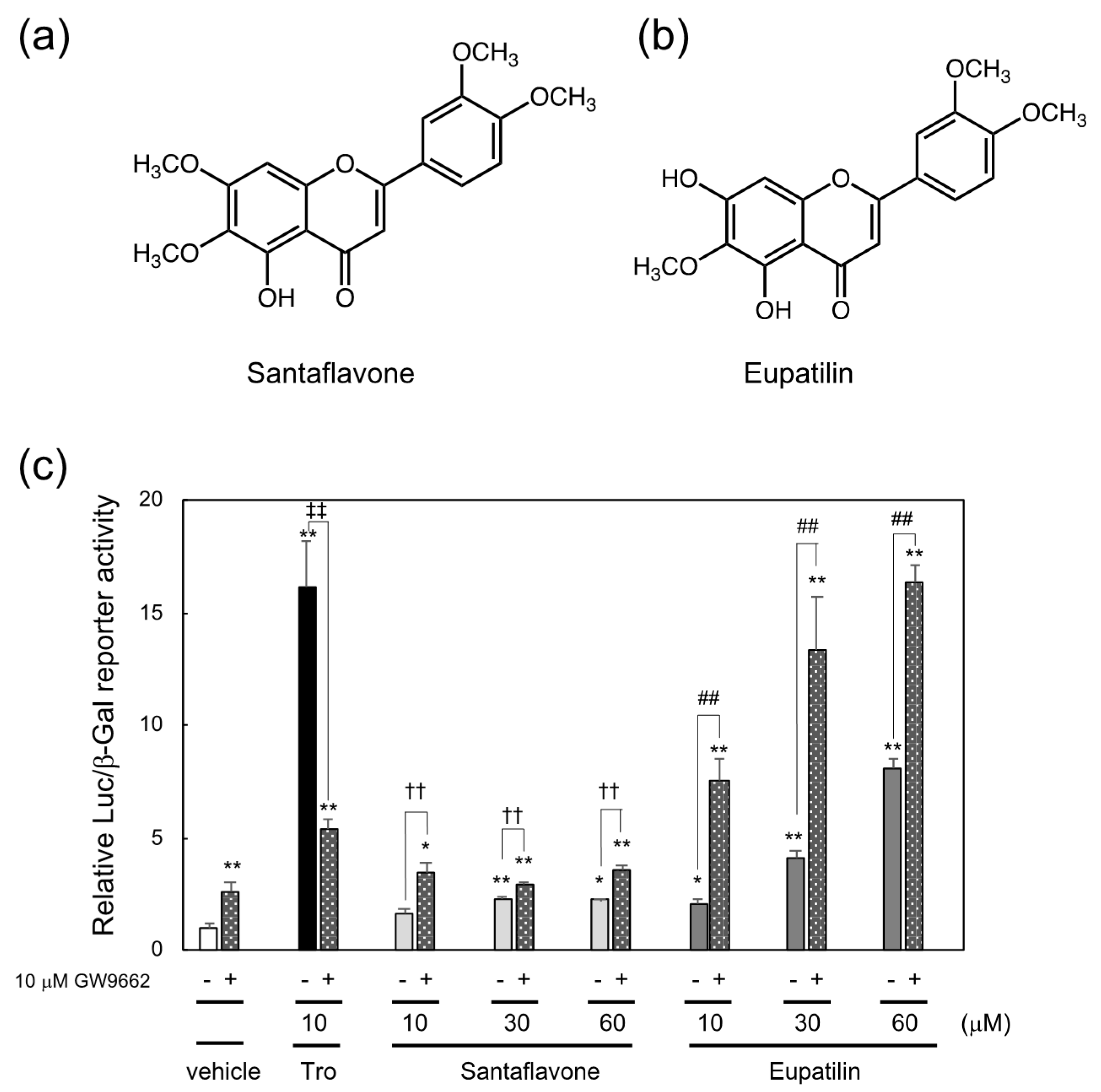
3.2. Identification of Santaflavone and Eupatilin as PPARγ Ligands in the Methanol Extracts of Artemisia Campestris That Function as the Cooperating PPARγ Ligands with GW9662
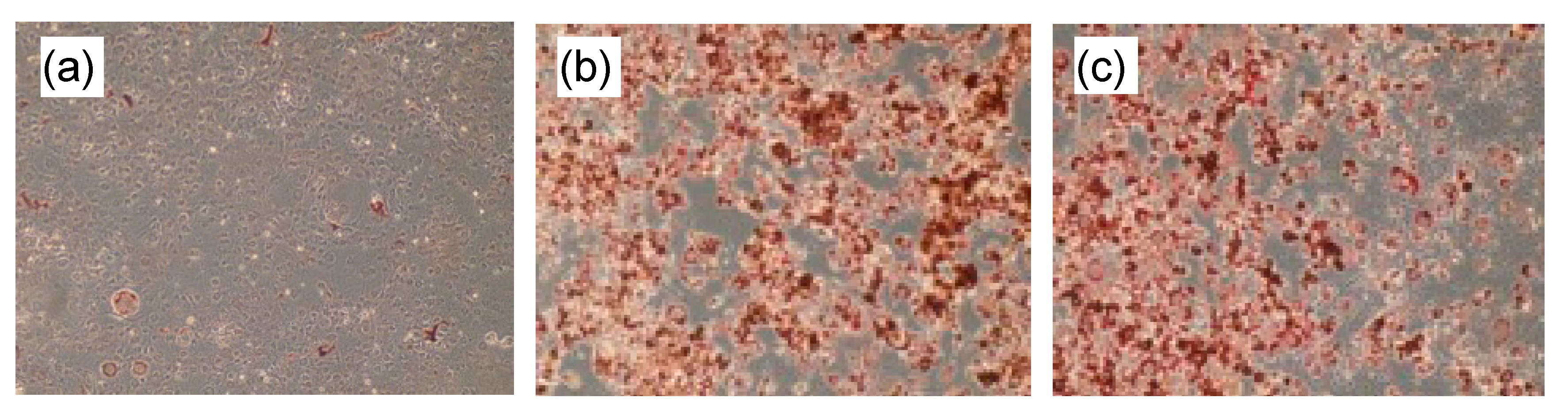
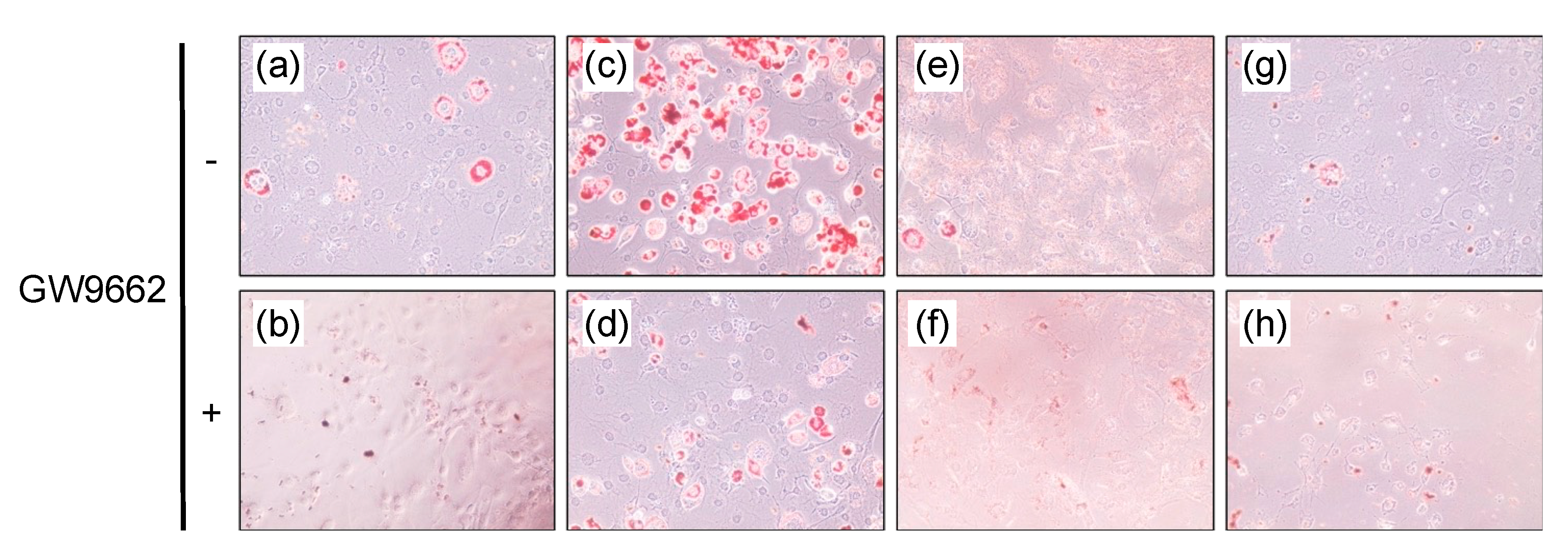
3.3. Lipid Accumulation in the 3T3-L1 cells
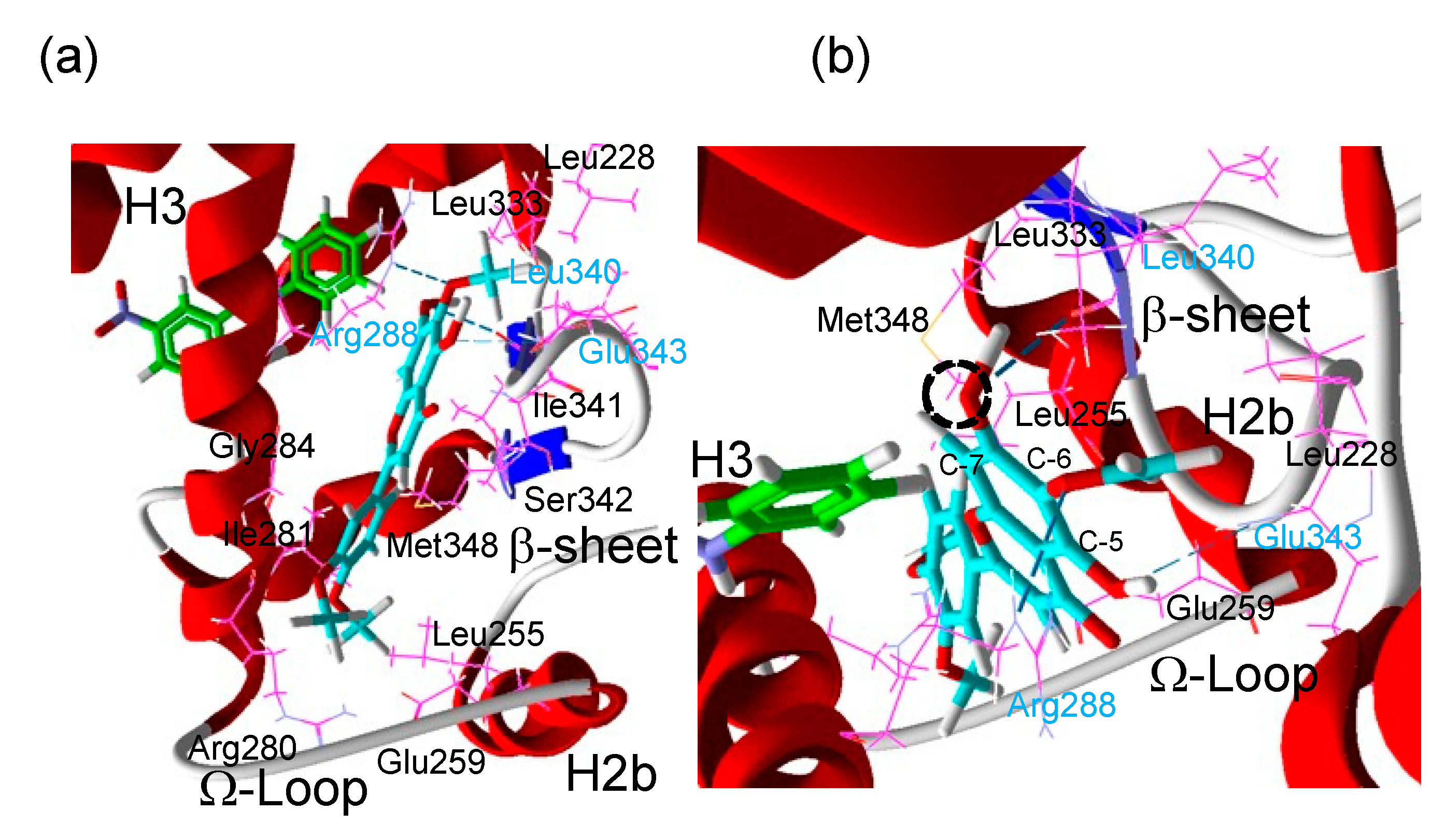
3.4. Analysis of the Putative Binding Mode of Eupatilin in the GW9662-Bound PPARγ LBD
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Dib, I.; El Alaoui-Faris, F.E. Artemisia campestris L.: Review on taxonomical aspects, cytogeography, biological activities and bioactive compounds. Biomed. Pharm. 2019, 109, 1884–1906. [Google Scholar] [CrossRef]
- Megdiche-Ksouri, W.; Trabelsi, N.; Mkadmini, K.; Bourgou, S.; Noumi, A.; Snoussi, M.; Barbria, R.; Tebourbi, O.; Ksouri, R. Artemisia campestris phenolic compounds have antioxidant and antimicrobial activity. Ind. Crop. Prod. 2015, 63, 104–113. [Google Scholar] [CrossRef]
- Sefi, M.; Fetoui, H.; Soudani, N.; Chtourou, Y.; Makni, M.; Zeghal, N. Artemisia campestris leaf extract alleviates early diabetic nephropathy in rats by inhibiting protein oxidation and nitric oxide end products. Pathol. Res. Pr. 2012, 208, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Tontonoz, P.; Spiegelman, B.M. Fat and beyond: The diverse biology of PPARγamma. Annu Rev. Biochem. 2008, 77, 289–312. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.V.; Brüne, B.; von Knethen, A. The nuclear hormone receptor PPARγ as a therapeutic target in major diseases. Sci. World J. 2010, 10, 2181–2197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubenstrunk, A.; Hanf, R.; Hum, D.W.; Fruchart, J.C.; Staels, B. Safety issues and prospects for future generations of PPAR modulators. Biochim. Biophys Acta 2007, 1771, 1065–1081. [Google Scholar] [CrossRef]
- Higgins, L.S.; Depaoli, A.M. Selective peroxisome proliferator-activated receptor gamma (PPARγamma) modulation as a strategy for safer therapeutic PPARγamma activation. Am. J. Clin. Nutr. 2010, 91, 267s–272s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waku, T.; Shiraki, T.; Oyama, T.; Maebara, K.; Nakamori, R.; Morikawa, K. The nuclear receptor PPARγ individually responds to serotonin- and fatty acid-metabolites. Embo. J. 2010, 29, 3395–3407. [Google Scholar] [CrossRef] [Green Version]
- Nolte, R.T.; Wisely, G.B.; Westin, S.; Cobb, J.E.; Lambert, M.H.; Kurokawa, R.; Rosenfeld, M.G.; Willson, T.M.; Glass, C.K.; Milburn, M.V. Ligand binding and co-activator assembly of the peroxisome proliferator-activated receptor-gamma. Nature 1998, 395, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Nagy, L.; Schwabe, J.W. Mechanism of the nuclear receptor molecular switch. Trends Biochem. Sci. 2004, 29, 317–324. [Google Scholar] [CrossRef]
- Leesnitzer, L.M.; Parks, D.J.; Bledsoe, R.K.; Cobb, J.E.; Collins, J.L.; Consler, T.G.; Davis, R.G.; Hull-Ryde, E.A.; Lenhard, J.M.; Patel, L.; et al. Functional consequences of cysteine modification in the ligand binding sites of peroxisome proliferator activated receptors by GW9662. Biochemistry 2002, 41, 6640–6650. [Google Scholar] [CrossRef]
- Ohtera, A.; Miyamae, Y.; Yoshida, K.; Maejima, K.; Akita, T.; Kakizuka, A.; Irie, K.; Masuda, S.; Kambe, T.; Nagao, M. Identification of a New Type of Covalent PPARγ Agonist using a Ligand-Linking Strategy. Acs Chem. Biol. 2015, 10, 2794–2804. [Google Scholar] [CrossRef]
- Ohtera, A.; Miyamae, Y.; Nakai, N.; Kawachi, A.; Kawada, K.; Han, J.; Isoda, H.; Neffati, M.; Akita, T.; Maejima, K.; et al. Identification of 6-octadecynoic acid from a methanol extract of Marrubium vulgare L. as a peroxisome proliferator-activated receptor γ agonist. Biochem. Biophys Res. Commun. 2013, 440, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Sakaue, H.; Ogawa, W.; Matsumoto, M.; Kuroda, S.; Takata, M.; Sugimoto, T.; Spiegelman, B.M.; Kasuga, M. Posttranscriptional control of adipocyte differentiation through activation of phosphoinositide 3-kinase. J. Biol. Chem. 1998, 273, 28945–28952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitagawa, I.; Fukuda, Y.; Yoshihara, M.; Yamahara, J.; Yoshikawa, M. Capillartemisin A and B, two new choleretic principles from Artemisiae Capillaris Herba. Chem Pharm Bull. 1983, 31, 352–355. [Google Scholar] [CrossRef] [Green Version]
- Akkal, S.; Benayache, F.; Bentamene, A.; Medjroubi, K.; Seguin, E.; Tillequin, F. Flavonoid Aglycones from Centaurea napifolia. Chem. Nat. Compd. 2003, 39, 219–220. [Google Scholar] [CrossRef]
- Deng, Y.-r.; Song, A.; Wang, H. Chemical Components of Seriphidium santolium Poljak. J. Chin. Chem. Soc. 2004, 51, 629–636. [Google Scholar] [CrossRef]
- Puhl, A.C.; Bernardes, A.; Silveira, R.L.; Yuan, J.; Campos, J.L.; Saidemberg, D.M.; Palma, M.S.; Cvoro, A.; Ayers, S.D.; Webb, P.; et al. Mode of peroxisome proliferator-activated receptor γ activation by luteolin. Mol. Pharm. 2012, 81, 788–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, R.; Yanagisawa, M.; Takahashi, N.; Kawada, T.; Kumazawa, S.; Yamaotsu, N.; Nakagome, I.; Hirono, S.; Tsuda, T. Brazilian propolis-derived components inhibit TNF-α-mediated downregulation of adiponectin expression via different mechanisms in 3T3-L1 adipocytes. Biochim. Biophys. Acta 2011, 1810, 695–703. [Google Scholar] [CrossRef]
- Shang, J.; Mosure, S.A.; Zheng, J.; Brust, R.; Bass, J.; Nichols, A.; Solt, L.A.; Griffin, P.R.; Kojetin, D.J. A molecular switch regulating transcriptional repression and activation of PPARγ. Nat. Commun. 2020, 11, 956. [Google Scholar] [CrossRef]
- Hughes, T.S.; Giri, P.K.; de Vera, I.M.; Marciano, D.P.; Kuruvilla, D.S.; Shin, Y.; Blayo, A.L.; Kamenecka, T.M.; Burris, T.P.; Griffin, P.R.; et al. An alternate binding site for PPARγ ligands. Nat. Commun. 2014, 5, 3571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.S.; Lee, S.G.; Min, K.; Kwon, T.K.; Kim, H.J.; Nam, J.O. Eupatilin inhibits adipogenesis through suppression of PPARγ activity in 3T3-L1 cells. Biomed. Pharm. 2018, 103, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.; Kim, J.C.; Park, N.J.; Bong, S.K.; Lee, S.; Jegal, H.; Jin, L.T.; Kim, S.M.; Kim, Y.K.; Kim, S.N. Eupatilin, an activator of PPARα, inhibits the development of oxazolone-induced atopic dermatitis symptoms in Balb/c mice. Biochem. Biophy.s Res. Commun 2018, 496, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.; Kim, J.C.; Choi, Y.; Lee, S.; Kang, K.S.; Kim, Y.K.; Kim, S.N. Eupatilin with PPARα agonistic effects inhibits TNFα-induced MMP signaling in HaCaT cells. Biochem. Biophys. Res. Commun. 2017, 493, 220–226. [Google Scholar] [CrossRef]
- Lai, C.S.; Tsai, M.L.; Cheng, A.C.; Li, S.; Lo, C.Y.; Wang, Y.; Xiao, H.; Ho, C.T.; Wang, Y.J.; Pan, M.H. Chemoprevention of colonic tumorigenesis by dietary hydroxylated polymethoxyflavones in azoxymethane-treated mice. Mol. Nutr. Food Res. 2011, 55, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.S.; Ho, M.H.; Tsai, M.L.; Li, S.; Badmaev, V.; Ho, C.T.; Pan, M.H. Suppression of adipogenesis and obesity in high-fat induced mouse model by hydroxylated polymethoxyflavones. J. Agric. Food Chem. 2013, 61, 10320–10328. [Google Scholar] [CrossRef]
- Bruning, J.B.; Chalmers, M.J.; Prasad, S.; Busby, S.A.; Kamenecka, T.M.; He, Y.; Nettles, K.W.; Griffin, P.R. Partial agonists activate PPARγamma using a helix 12 independent mechanism. Structure 2007, 15, 1258–1271. [Google Scholar] [CrossRef]
- Scott, D.E.; Coyne, A.G.; Hudson, S.A.; Abell, C. Fragment-based approaches in drug discovery and chemical biology. Biochemistry 2012, 51, 4990–5003. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasegawa, T.; Osaka, M.; Miyamae, Y.; Nishino, K.; Isoda, H.; Kawada, K.; Neffati, M.; Irie, K.; Nagao, M. Two Types of PPARγ Ligands Identified in the Extract of Artemisia campestris. Chemistry 2021, 3, 647-657. https://0-doi-org.brum.beds.ac.uk/10.3390/chemistry3020045
Hasegawa T, Osaka M, Miyamae Y, Nishino K, Isoda H, Kawada K, Neffati M, Irie K, Nagao M. Two Types of PPARγ Ligands Identified in the Extract of Artemisia campestris. Chemistry. 2021; 3(2):647-657. https://0-doi-org.brum.beds.ac.uk/10.3390/chemistry3020045
Chicago/Turabian StyleHasegawa, Tokio, Mayo Osaka, Yusaku Miyamae, Katsutoshi Nishino, Hiroko Isoda, Kiyokazu Kawada, Mohamed Neffati, Kazuhiro Irie, and Masaya Nagao. 2021. "Two Types of PPARγ Ligands Identified in the Extract of Artemisia campestris" Chemistry 3, no. 2: 647-657. https://0-doi-org.brum.beds.ac.uk/10.3390/chemistry3020045