
Inflammation and Preterm Birth: A Systematic Review


1
Obstetrics Department, Faculty of Medicine, Centro Hospitalar e Universitário de Coimbra, 3000-076 Coimbra, Portugal
2
Faculty of Medicine, Coimbra Institute for Clinical and Biomedical Research (iCBR), University of Coimbra, 3004-504 Coimbra, Portugal
3
Centre of Investigation in Environment, Genetics and Oncobiology (CIMAGO), Faculty of Medicine, University of Coimbra, 3004-504 Coimbra, Portugal
*
Author to whom correspondence should be addressed.
Reprod. Med. 2022, 3(2), 101-111; https://0-doi-org.brum.beds.ac.uk/10.3390/reprodmed3020009
Submission received: 1 March 2022
/
Revised: 21 April 2022
/
Accepted: 26 April 2022
/
Published: 1 May 2022
(This article belongs to the Special Issue Inflammation in Pregnancy)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Inflammatory mechanisms have a critical role in parturition, which results from a gathering of different stimuli that collectively initiate labour. In fact, a sophisticated interaction occurs between contractile and immuno-inflammatory pathways, whereby proinflammatory amplification is intensified by collaborative connections between cells, ligands, and tissues. Preterm birth (PTB) is one of the major challenges of modern obstetrics and still lacks an efficient treatment. Therefore, the scientific research of modern therapies is warranted. This systematic review aims to provide an overview of recent research into inflammation and PTB. The main inclusion criterion was articles concerning birth and inflammation, and searches were performed in the electronic databases MEDLINE, Embase, Scopus, Web of Science and Cochrane Library, from 2017 to 2021. A literature search from all databases yielded 1989 results which, applying the specified eligibility criteria, resulted in the 16 articles included in this review. Delivery is the consequence of an inequity between maternal inflammation and hormonal-driven uterine quiescence. Studies show that the distinction between term and preterm labour could consist of a pre-existing disproportion of decidual inflammatory signalling, or an unusual stimulus eliciting inflammatory pathways, comparable to both. Thus, controlling inflammation could be hopeful for detaining PTB.
1. Introduction
Inflammatory mechanisms are accepted to have important roles in ovulation, implantation, decidualization and parturition [1]. Moreover, these inflammatory mechanisms have an impact not only on maternal and placental health, but also on foetal development and conventional growth [1]. In fact, successful implantation relies on an injury induced inflammatory reaction [2], and localized membrane inflammation contributes to tissue remodelling during pregnancy [3]. Since 1995, it has generally been accepted that labour results from a constellation of diverse stimuli that together initiate it (as opposed to a single-stimulus theory) [4].
Parturition is physiologically initiated when foetal maturation is completed [3]. Biochemical signalling from matured foetal organs indicates the completion of pregnancy and initiates the contractile pathway of the uterus [3]. Inflammatory mediators from damaged membranes are propagated via extracellular vesicles to maternal tissues, which activate labour [3]. More recently, the terms inflammatory load and inflammatory burden were introduced, emphasizing their importance in reproductive medicine [1].
The expression uterine transition refers to the complex process of modifying the uterus from a quiescent to an active phase. In fact, there is an intricate interaction between contractile and immune pathways, whereby proinflammatory amplification is heightened by collaborative connections among cells, ligands, and tissues [5]. These relations generate positive feedback interactions regarding paracrine and autocrine signalling in the uterus and other reproductive tissues, leading to a rise in the inflammatory burden that prompts labour [5].
Therefore, the transition to labour is an inflammatory event not limited to a single gestational tissue, involving more than the myometrium; nevertheless, studies show that this uterine transition is a localized intrauterine inflammatory response [5]. Labour is comparable to a volcano in which many processes take place before we see it burst; it is like a high-speed train that is impossible to stop beyond a certain threshold [5]. Similarly, labour results from breaching an inflammatory line that leads to progesterone withdrawal and the activation of uterine tissues [5]. Therefore, delivery is the result of an imbalance between maternal inflammation and hormone-driven uterine quiescence [1]. Indeed, for the parturition process to occur, there must be a joint action of inflammatory cells, proinflammatory mediators, and signalling cascades [1].
Preterm birth (PTB) affects 11% of all births, corresponding to 15 million babies/annually, of which two-thirds are spontaneous [6]. The rates of PTB range from 5% in Europe, 10.2% in USA, to 18% in certain African countries [7,8]. The majority of women who deliver prematurely (85%) do not have any risk factors [9]. Moreover, women who unexpectedly present with threatened PTB, notwithstanding the usage of foetal fibronectin, tocolytic therapy and steroids, will give birth before 34 weeks in 15 to 20% of cases [9]. So, new strategies for PTB treatment are one of the most important goals in present day maternal–foetal medicine worldwide.
This systematic review aims to discover if recent research about inflammation and PTB can answer the question of whether there are anti-inflammatory strategies that can stop preterm labour.
2. Materials and Methods
The present protocol was registered within PROSPERO database (registration number CRD42022302372) and was reported in accordance with the reporting guidance provided in the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) statement (see checklist in Supplementary File S1).
2.1. Eligibility Criteria
The strict inclusion criterion were that articles should concern birth and inflammation, both term and preterm births.
Exclusion criteria were opinion articles; no data on inflammation; PTB preventive strategies; articles comparing different PTB treatment strategies; immunology of the placenta; and foetal information only.
2.2. Information Sources and Search Strategy
Searches were performed in the electronic databases MEDLINE, Embase, Scopus, Web of Science and Cochrane Library, from January 2017 to December 2021. The literature searches were designed and conducted by the review team ALA and AMP.
Example of a full electronic search (PubMed): (“Premature Birth” [Mesh]) AND “Inflammation” [Mesh]) OR (“Premature Birth/prevention and control” [Mesh]) OR (“Premature Birth” [Mesh]) AND “Genetic Predisposition to disease” [Mesh] OR “Inflammation” [Mesh]) OR (“Premature Birth” [Mesh]) AND “Immunity, Innate” [Mesh] OR “Adaptive Immunity” [Mesh] OR “Cytokines” [Mesh] OR (“Premature Birth/prevention and control” [Mesh]) AND “Progesterone” [Mesh]) OR (“Prostaglandins” [Mesh]) OR (“Premature Birth/prevention and control” [Mesh]) AND “Tocolytic Agents” [Mesh]) OR (“Premature Birth” [Mesh] AND “Tocolytic Agents/therapeutic use” [Mesh]).
2.3. Study Selection
The search was limited to human studies, with English or French as languages of report writing, conducted in the last five years. The following medical subject headings (MeSH) were used: premature birth; inflammation; cytokines; prostaglandins; tocolytic agents; immunity, innate; adaptive immunity; progesterone. Secondary sources of literature consisted of the hand-searching of reference lists of included studies, relevant reviews, or clinical practice guidelines.
2.4. Data Extraction
The following processes were carried out: the analysis of titles and abstracts (two researchers); when necessary, discussion of differences between choices and definition of a consensual final choice (two researchers); reading of the full text of the chosen articles (two researchers) and analysis of the results (two researchers). Disagreements were solved by debate between both authors.
Original authors were contacted for missing data, and an Excel file was used to record data.
Variables for which data were taken were genetic predisposition; cytokines; prostaglandins; tocolytic agents; innate immunity components; adaptive immunity components; estrogens, progesterone, and new therapies.
3. Results
The PRISMA flow diagram showing the inclusion and exclusion of published articles is shown in Figure 1.
The literature searches from all databases yielded 1989 results which, after duplicate exclusion, resulted in 200 references. Studies were then screened, using the specified eligibility criteria, and studies that did not meet the criteria were excluded, resulting in 49 full-text studies for deep analysis and data extraction. A review of the reference lists of these articles also added relevant citations. All levels of screening were conducted by two independent reviewers and any disputes between reviewers were solved by active discussion and unanimous agreement. None of the reviewers were blinded to titles, authors, journals, or institutions.
- Risk of bias within studies and across studies
Due to the study design, a clear evaluation of the risk of bias could not be conducted. Nevertheless, the potential reporting, selection and publication bias should be considered. Despite all efforts to reduce the risk of bias and increase data quality, some selection bias might be present in the current analysis, as other databases could have been included. Moreover, selective reporting bias cannot be excluded, as well as publication bias caused by the inclusion of published studies only.
4. Discussion
4.1. Immuno-Inflammation and Normal Delivery
As explained, the uterine transition event encompasses several components: (1) myometrium, where there are uterine activation proteins (such as COX2) that change prior to the beginning of labour and are regulated by proinflammatory mediators (interleukin (IL) 6 and 1β) and steroid hormones; (2) cervix, where the cervical ripening process is influenced by steroid hormones, prostaglandins, growth factors and invading leukocytes; (3) decidua, which upregulates uterine activation products and undergoes activation; (4) foetal tissues, where an increase in the amount of oxidative stress and cell senescence during the gestational period occurs, prompting the release of sterile inflammatory mediators of parturition (DAMPs) and the upregulation of senescence-associated secretory phenotypes, both of which lead to an inflammatory upregulation that determines birth timing and membrane rupture; and (5) maternal leukocytes, which are attracted by the release of chemoattractants by foetal membranes [5].
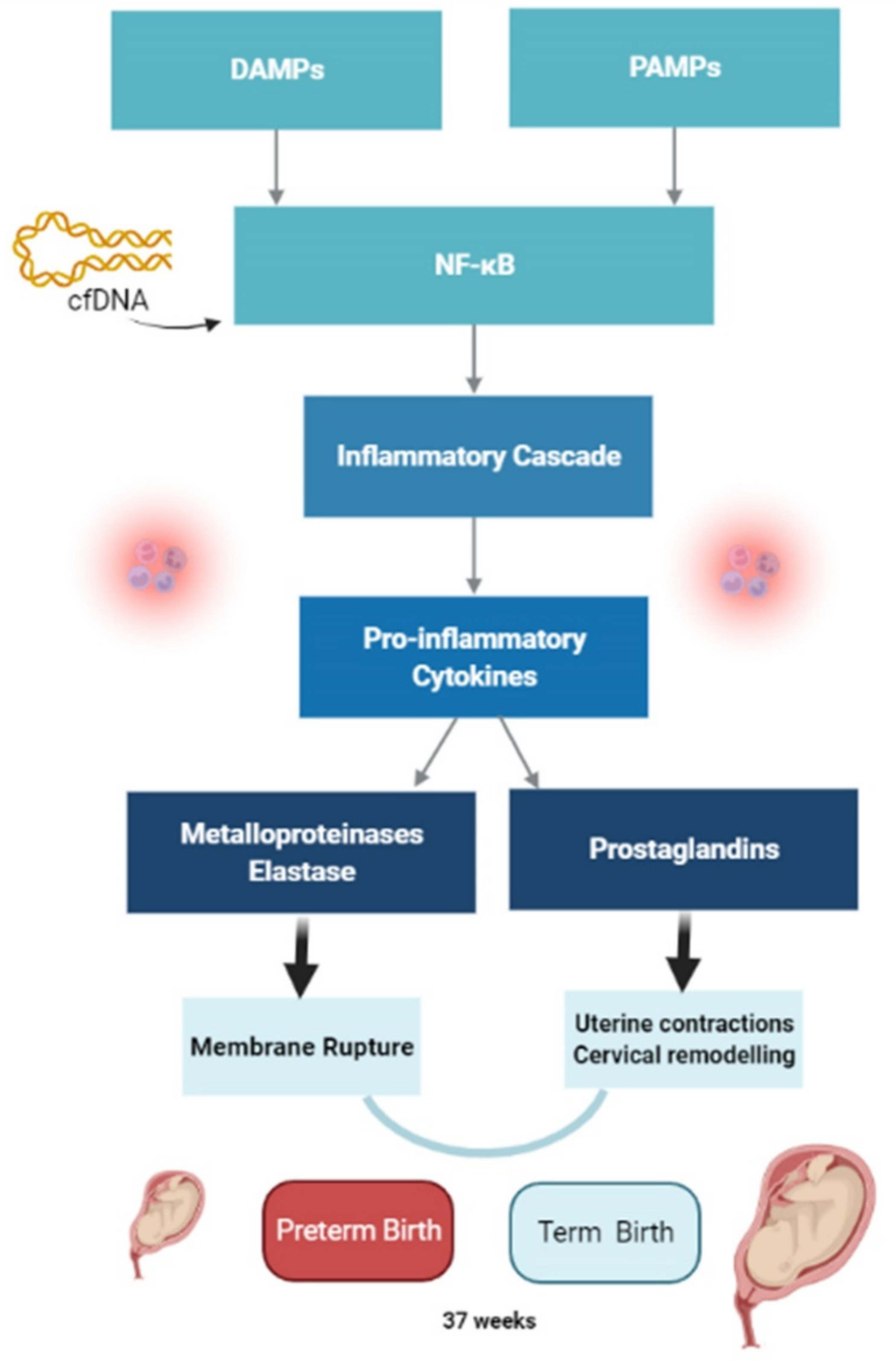
Regardless of gestational age, inflammatory activation occurs in intrauterine tissues during labour, and it is self-propagating [1]. The switch of the myometrium from a quiescent to a contractile state is accompanied by a shift in signalling between anti-inflammatory and proinflammatory pathways, including chemokines (IL-8), cytokines (IL-1, IL-6) and contraction-associated proteins (expression of conexin 43, oxytocin receptors, prostaglandin receptors) [12]. The process starts with pathogen-associated molecular patterns (PAMPs) that act through pattern-recognition receptors (PRRs), activating the nuclear factor kappa light chain enhancer of activated B cells (NF-κB), which in turn elicits the inflammatory cascade [6]. As inflammatory cytokines are released, there is an accumulation of neutrophils, with the subsequent production of metalloproteinases, elastase, and elevated prostaglandin (PGF2α). Metalloproteinases and elastase are responsible for membrane rupture, and elevated PGF2α for both uterine contractions and cervical remodelling. All of these processes elicit birth [6] (Figure 2). Damage-associated molecular patterns (DAMPs) or alarmins are released not only from far away infections from the intrauterine tissues, but also from cell death due to trauma, stress, toxins, or tissue ischemia [6]. Oxidative stress and cellular senescence of foetal amnion and chorion may trigger human parturition through the release of these DAMPs [14]. In fact, oxidative stress in response to risk factors leads to accelerated senescence and promotes a terminal state of cellular transition from the epithelial to mesenchymal state. These events promote inflammation, which degrades the matrix and destabilizes membrane function [3]. Recently it has been argued that cell-free DNA (cfDNA) has a role as a signal for the onset of labour, due to its proinflammatory properties [12]. Cell-free DNA can bind to Toll-like receptor (TLR) 9, which leads to NF-κB activation and enhances inflammatory cytokine gene expression [12]. Studies disclosed that women with spontaneous-term labour show higher levels of cfDNA than non-labouring women [12,15].
Inflammasomes are cytosolic protein complexes that orchestrate an inflammatory response to pathogens and endogenous danger signals. The high-mobility group box 1 protein induces an inflammatory response in chorioamniotic membranes, which is partially mediated by inflammasomes [12]. The participation of the inflammasome in labour mechanisms related to inflammation is proven by the presence of elevated CASP1, IL-1β and IL-18 in the amniotic fluid of women with PTB [17].
Subpopulations of maternal peripheral leukocytes (granulocytes, monocytes and lymphocytes) specifically activated with labour onset were identified [12]. Natural killer (NK) cells from the decidua are different from the other NK cells in the body: they are critical in producing cytokines and shifting the cytokine profile at the maternal foetal interface [2]. Maternal macrophages and foetal antigen-specific cells may activate, expand and participate in the labour process, either locally or systemically [10].
4.2. Immuno-Inflammation and Preterm Birth
Accordingly, the difference between term and preterm labour might be a precocious imbalance of decidual inflammatory signalling, or a potent abnormal stimulus (internal or external) that triggers inflammatory pathways, similar in term and preterm labour [12].
PTB is characterized by an increase in proinflammatory mediators, such as IL-1, IL-6, IL-8, TNF-α, INF-γ, as well as a decrease in anti-inflammatory mediators, such as IL-10 and IL-4. In fact, IL-10 is a key cytokine in pregnancy maintenance, with a preponderant role in preterm labour and delivery [2].
PTB syndrome includes placental dysfunction, premature uterine contractions, rupture of membranes and cervix dilatation [10]. Additionally, PTB may be considered as a disease of the myometrium, cervix, endometrium/decidua and/or placenta [3]. Around 30% of patients with PTB have placental lesions consistent with maternal vascular underperfusion [12]. Therefore, PTB may be regarded as a disorder that results from defective deep placentation, such as preeclampsia [14]. Moreover, early immune alterations caused by maternal stress perception, infection and diet may result in distorted immune tolerance and excess inflammation [14].
The placenta is not sterile with its bacterial flora being closer to that of the oral cavity than that of the vagina [11]. Inflammation and infection are implicated in 20–25% of preterm deliveries. However, intrauterine inflammation may present without evidence of pathological intrauterine infection, providing the recent notion of triple “I”, which stands for intrauterine inflammation, infection, or both [8,13]. Furthermore, sterile intra-amniotic inflammation is more frequent than microbial-associated intraamniotic inflammation and is reported to be present in 10% of women with a short cervix [1,2]. Nevertheless, both induce the same cytokine mediators [2].
During the uterine transition phase, most important uterine mediators are IL-6 and IL-1β [5]. IL-6 is involved in implantation, pregnancy, and parturition, with high levels at labour onset. It also has the ability to stimulate the amnion and decidual cells leading to prostaglandin increase. Moreover, IL-6 has been intensively investigated as a diagnostic biomarker of preterm delivery, measured in the vaginal fluid [5,13]. The degree of inflammation determined by IL-6 concentration correlated with the rate of perinatal morbidity and mortality [2,19]. However, others argue that the diagnosis of intraamniotic inflammation based on amniotic fluid IL-6 under-represents the true extent of intrauterine inflammation [1].
On the other hand, IL-1β influences the coordination of proinflammatory and pro-labour gene regulation in intrauterine tissues [5]. IL-1β is also a potent stimulator of prostaglandin synthesis via COX2 induction and contributes to progesterone (P4) withdrawal by increasing nuclear progesterone receptor A [5]. Additionally, IL-1-related pathways are upregulated during the third trimester in women who deliver preterm [18].
In terms of cells, PTB is associated with increased choriodecidual inflammation and increased M1 macrophages and NK cells [10].
A damaging condition might stimulate the innate system through PAMPs (infectious stimuli) or DAMPs (sterile stimulus) [5]. DAMPs can be released by the stressed uterus, maturing foetus, or ageing placenta [5]. Recent evidence demonstrated that foetal membrane senescence is an important sign at the beginning of labour [5] and some studies revealed that cfDNA was a trigger of sterile inflammation [1]. Accordingly, innate stimulation activates TLRs that gather the inflammasome, stimulate the release of proinflammatory cytokines and chemokines, and activate leukocytes, unequivocally initiating labour [5]. If the dysregulation of decidual inflammatory signalling happens early, spontaneous PTB will supervene [12]. Nevertheless, PTB may also result from the active induction of decidual inflammation in the mid trimester of pregnancy due to, for instance, intrauterine infection or placental abruption [12]. Moreover, risk factors, such as cigarette smoking, anemia, cervical shortening, lower genital tract infection, ethnicity, extreme birth weight (low or high), may be mere indicators of dysfunctional immunological protection within uterine tissues [12].
- Genetic factors and PTB
Genetic factors play a role in PTB as it shows a familial aggregation; has measures of heritability; can be detected by susceptibility genes; and reveals a racial disparity [17]. Additionally, it recurs in mothers and women who were preterm babies and are at a higher risk of delivering preterm babies themselves [18]. Twin studies show that genetics affects at least one-third of the variation seen in PTB [18]. A study based on maternal data revealed that, from the 210 genes differentially expressed in spontaneous PTB, genes and cell types associated with innate immunity were upregulated, while those associated with adaptive immunity were downregulated [18]. From these genes, 18 of them displayed different expressions according to trimesters (namely in the second trimester) [18]. Both IL-1R1 and tissue factor pathway inhibitor encode immune-related proteins that were found to be differently expressed and longitudinally secreted [18]. Therefore, PTB has a polygenic basis involving rare mutations or damaging variants in multiple genes involved in innate immunity and host defence mechanisms against microbes and their hazardous products [17]. Hence, genes involved in the inflammatory process are upregulated and amplify these processes [1], and there are identity genetic variants that influence the inflammatory response in PTB [17].
Fascinating recent evidence suggests that some genes are reversed in maternal foetal environments: TLR5 upregulated in the mother and downregulated in the foetus [18]. Additionally, evidence points out that adult-onset diseases seen in premature babies are partly related to epigenetic changes/variations due to wrong foetal programming [3].
4.3. Reproductive Hormones and Labour
Gestational hormones are important in inflammatory reactions during pregnancy [8]. Estrogens inhibit proinflammatory cytokines (IL-1; TNF-α; IFN-γ) and stimulate anti-inflammatory cytokines such as IL-10, IL-4 and TGF-β [2]. Nevertheless, IL-10 has been accepted as a part of a pro-labour inflammatory response, having both an active and a tolerant part in labour that changes with the tissue involved [1].
Progesterone (P4) has anti-inflammatory properties, helps prevent uterine contractions and protects foetal membranes [8]. However, human pregnancy is unique in that peripheral progesterone levels remain unchanged, and only fall after the delivery of both the foetus and placenta [2,5]. Serum P4 levels rise, and its effect on pregnancy maintenance decreases just before delivery [5]. So, the differential expression of its nuclear progesterone receptors (nPR), A or B, might be the feature responsible for the changes during labour [2]. During pregnancy, nPRB, the main receptor, has anti-inflammatory effects, acting to prevent labour and delivery [1]. On the contrary, just before labour, there is an increase in nPRA versus nPRB, with its proinflammatory actions and the inhibition of nPRB activity [5]. Myometrial cells with a high nPRA/nPRB ratio show a proinflammatory status [5]. Therefore, we can assume that parturition is inflammation-induced, whereby proinflammatory/prolabour stimuli induce nPRA-mediated adjustments [16], and that the key mechanism by which P4 is believed to block labour is through decreasing uterine cell responsiveness to proinflammatory/prolabour stimuli [16]. P4 also weakens the response to infection in PTB, as evidenced by high levels of TLR4 and low levels of TLR2 [2]. As a consequence, external P4 therapy given to women with a high risk of PTB seems to accomplish its actions by reducing maternal inflammation in both the decidua and cervix [1], and by diminishing the expression of contraction-associated proteins, inflammatory cytokines and hindering cervical ripening [6].
Consequently, PTB requires an inflammatory stimulus sufficient to overcome progesterone effects [1]. At earlier gestation, intrauterine infection is a robust pathological drive for PTB. At later gestation, as the maternal inflammatory burden increases, P4 effects are eliminated and tissue remodelling initiates [1].
4.4. New Approaches in PTB Treatment
Until now, the PTB treating strategy has been to antagonize contractile mediators’ actions that stimulate myometrial contractions, none being very effective. For this reason, the approach ought to be revised, focusing on the newly discovered pathways [5]. Accordingly, the new experimental drugs in PTB inhibition/arrest encompass (Figure 3):
- Drugs that interfere with nPRA/nPRB balance:
- (a)
- Selective progesterone receptor modulators (SPRMs) boost P4 anti-inflammatory activity that blocks labour, while at the same time preventing PR phosphorylation (of serine residue 344 and 345 in nPRA) [8], leading to a loss of P4 anti-inflammatory activity [14]. SPRMs are compounds that compete with P4 for binding to the nuclear PR ligand-binding pocket and induce mixed agonist/antagonist activity [16]. Nuclear PRs are presumably saturated by progesterone during pregnancy [14]. Since membrane PR have a lower binding affinity, they may be pharmacologically affected by additional P4 [14]. Moreover, it is possible that nuclear PR signalling in myometrial, endometrial and decidual cells might be affected by an interaction with membrane PR activity, altered by exogenous P4 [14].
- (b)
- Histone deacetylase inhibitors (Trichostatin A): seems to promote nPRB expression in human myometrial samples [8].
- (c)
- Chloroquine: human myometrial cells express bitter taste receptors, and the activation of these receptors leads to muscle relaxation. Additionally, chloroquine, as a bitter tastant, is also able to prevent inflammation induced PTB [8].
- PGF2 α antagonists: recent research showed that, when used in combination with Atosiban, delivery was reduced 55% more efficiently than with Atosiban alone [5].
- TLR4 antagonists (naloxone, naltrexone, and rosiglitazone) diminish proinflammatory cytokines [8]. Naloxone (+)- is an isomer of the antagonist of opioid receptor naloxone (−)-. Naloxone (+)- has anti-inflammatory properties with the capacity to specifically antagonize TLR4; nevertheless, it has no opioid activity [6]. In animal models, it reduces PTB, foetal demise and cytokine expression in intrauterine tissues [6]. Additionally, it has the benefit of providing a greater blockage than other more distally located targets [6]. However, drugs targeting TLR4 may only be useful in PTB triggered by Gram-negative bacteria (which can elicit inflammation by another TLR, rendering some freedom to the birth pathway) [6].
- Cytokine-suppressive anti-inflammatory drugs (CSAIDs) inhibit intracellular pathways that result in NF-κB activation [6]. These drugs are more selective and effective than NSAIDs with no adverse neonatal effects (specifically abortion and renal impairment). The classes being studied are TAK1 inhibitors; P38 MAPK inhibitors and IKK complex inhibitors (of which TPCA 1 is the most promising) [6].
- NF-κB inhibitors: N-acetylcysteine is involved in reducing oxidative stress, with promising results in PTB reduction and an additional benefit in foetal neuroprotection. Sulfasalazine, a drug frequently used in inflammatory bowel disease, inhibits IKK kinases and seems to reduce inflammatory cytokine release in vitro [6].
- Drugs targeting specific proinflammatory cytokines:
- TNF-α inhibitors (infliximab and etanercept) are only studied in mice but raise concerns regarding the increased risk of neonatal infection [6].
- Polyunsaturated fatty acids (PUFA) and lipid metabolites are known to resolve oxidative stress and inflammation through arachidonic acid pathways. The balance between metabolites of omega-3 and omega-6 PUFA seems to be important in pregnancy maintenance and have a significant role in cervical effacement, dilation and initiation of labour. θ3 PUFA have anti-inflammatory actions but θ6 have proinflammatory properties [6]. The recent ORIP trial found that they did not reduce PTB [6]. However, the new molecules (specialized pro-resolving lipid mediators, such as lipoxin A4 and resolving E2) have not been studied in humans [6]. Moreover, plasma N-acylethanolamines were recently proposed as a single simple test that could be used to predict PTB in high-risk women [20].
- Polyphenols suppress the production of proinflammatory cytokines such as TNF-α and IL-1β. Resveratrol has shown good results, but the increase in foetal pancreatic mass makes its use undesirable [6].
According to these results, the best candidates for future PTB treatments are CSAIDs (Naloxone could be chosen for low-income countries), and the IL-1 receptor inhibitor (Rytvela might be the ideal for high-income countries).
As no drug has proven unequivocal efficiency in PTB arrest, new delivery systems are also being developed and tested [8]. These new delivery systems are: (1) targeted liposomes (systemically deliver tocolytics to the uterus); (2) mucoinert vaginal progesterone nanosuspension formulations, which carry drugs vaginally; (3) exosomes; (4) hydroxyl polyamidoamine dendrimers, that allow the coupling of therapeutic and imaging mediators; and (5) an acetate vaginal ring with progesterone [8].
5. Final Comment
Current PTB treatment focuses on the inhibition of myometrial contractions which are a consequence of the inflammatory cascade responsible for the beginning of PTB [6]. Nonetheless, none of these medications are especially efficient. So, pharmacological agents that aim to reduce or eradicate the inflammatory process seem to be the way forward [6]. Of these, the two with the best results are CSAIDs (Naloxone) and an IL-1 receptor inhibitor (Rytvela) [6]. Additionally, the idea of designing SPRMs that simultaneously boost P4/nPR anti-inflammatory activity and prevent the phosphorylation of Ser 344/345 nPRA-mediated progesterone withdrawal, is promising [16].
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/reprodmed3020009/s1, PRISMA Checklist [21].
Author Contributions
Conceptualization, A.L.A. and A.M.-P.; methodology, A.L.A.; software, A.L.A.; validation, A.L.A. and A.M.-P.; formal analysis, A.L.A.; investigation, A.L.A.; writing—original draft preparation, A.L.A.; writing—review and editing, A.L.A. and A.M.-P.; supervision, A.M.-P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Keelan, J.A. Intrauterine inflammatory activation, functional progesterone withdrawal, and the timing of term and preterm birth. J. Reprod. Immunol. 2018, 125, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Kalagiri, R.R.; Carder, T.; Choudhury, S.; Vora, N.; Ballard, A.R.; Govande, V.; Drever, N.; Beeram, M.R.; Uddin, M.N. Inflammation in Complicated Pregnancy and Its Outcome. Am. J. Perinatol. 2016, 33, 1337–1356. [Google Scholar] [CrossRef] [PubMed]
- Menon, R.; Behnia, F.; Polettini, J.; Richardson, L.S. Novel pathways of inflammation in human fetal membranes associated with preterm birth and preterm pre-labor rupture of the membranes. Semin. Immunopathol. 2020, 42, 431–450. [Google Scholar] [CrossRef]
- Olson, D.M.; Mijovic, J.E.; Sadowsky, D.W. Control of human parturition. Semin. Perinatol. 1995, 19, 52–63. [Google Scholar] [CrossRef]
- Leimert, K.B.; Xu, W.; Princ, M.M.; Chemtob, S.; Olson, D.M. Inflammatory Amplification: A Central Tenet of Uterine Transition for Labor. Front. Cell Infect. Microbiol. 2021, 11, 660983. [Google Scholar] [CrossRef] [PubMed]
- Triggs, T.; Kumar, S.; Mitchell, M. Experimental drugs for the inhibition of preterm labor. Expert. Opin. Investig. Drugs 2020, 29, 507–523. [Google Scholar] [CrossRef] [PubMed]
- Boyle, A.K.; Rinaldi, S.F.; Norman, J.E.; Stock, S.J. Preterm birth: Inflammation, fetal injury and treatment strategies. J. Reprod. Immunol. 2017, 119, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Zierden, H.C.; Shapiro, R.L.; DeLong, K.; Carter, D.M.; Ensign, L.M. Next generation strategies for preventing preterm birth. Adv. Drug Deliv. Rev. 2021, 174, 190–209. [Google Scholar] [CrossRef]
- Singh, N.; Bonney, E.; McElrath, T.; Lamont, R.F.; Preterm Birth International collaborative (PREBIC). Prevention of preterm birth: Proactive and reactive clinical practice-are we on the right track? Placenta 2020, 98, 6–12. [Google Scholar] [CrossRef]
- Bonney, E.A.; Johnson, M.R. The role of maternal T cell and macrophage activation in preterm birth: Cause or consequence? Placenta 2019, 79, 53–61. [Google Scholar] [CrossRef]
- Di Renzo, G.C.; Cabero Roura, L.; Facchinetti, F.; Helmer, H.; Hubinont, C.; Jacobsson, B.; Jorgensen, J.S.; Lamont, R.F.; Mikhailov, A.; Papantoniou, N.; et al. Preterm Labor and Birth Management: Recommendations from the European Association of Perinatal Medicine. J. Matern. Fetal Neonatal. Med. 2017, 30, 2011–2030. [Google Scholar] [CrossRef]
- Di Renzo, G.C.; Tosto, V.; Giardina, I. The biological basis and prevention of preterm birth. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 52, 13–22. [Google Scholar] [CrossRef]
- Gilman-Sachs, A.; Dambaeva, S.; Salazar Garcia, M.D.; Hussein, Y.; Kwak-Kim, J.; Beaman, K. Inflammation induced preterm labor and birth. J. Reprod. Immunol. 2018, 129, 53–58. [Google Scholar] [CrossRef]
- Green, E.S.; Arck, P.C. Pathogenesis of preterm birth: Bidirectional inflammation in mother and fetus. Semin. Immunopathol. 2020, 42, 413–429. [Google Scholar] [CrossRef]
- Herrera, C.A.; Stoerker, J.; Carlquist, J.; Stoddard, G.J.; Jackson, M.; Esplin, S.; Rose, N.C. Cell-free DNA, inflammation, and the initiation of spontaneous term labor. Am. J. Obstet. Gynecol. 2017, 217, 583.e1–583.e8. [Google Scholar] [CrossRef]
- Mesiano, S.A.; Peters, G.A.; Amini, P.; Wilson, R.A.; Tochtrop, G.P.; van Den Akker, F. Progestin therapy to prevent preterm birth: History and effectiveness of current strategies and development of novel approaches. Placenta 2019, 79, 46–52. [Google Scholar] [CrossRef]
- Strauss, J.F., 3rd; Romero, R.; Gomez-Lopez, N.; Haymond-Thornburg, H.; Modi, B.P.; Teves, M.E.; Pearson, L.N.; York, T.P.; Schenkein, H.A. Spontaneous preterm birth: Advances toward the discovery of genetic predisposition. Am. J. Obstet. Gynecol. 2018, 218, 294–314.e2. [Google Scholar] [CrossRef]
- Vora, B.; Wang, A.; Kosti, I.; Huang, H.; Paranjpe, I.; Woodruff, T.J.; MacKenzie, T.; Sirota, M. Meta-Analysis of Maternal and Fetal Transcriptomic Data Elucidates the Role of Adaptive and Innate Immunity in Preterm Birth. Front. Immunol. 2018, 9, 993. [Google Scholar] [CrossRef]
- Combs, C.A.; Gravett, M.; Garite, T.J.; Hickok, D.E.; Lapidus, J.; Porreco, R.; Rael, J.; Grove, T.; Morgan, T.K.; Clewell, W.; et al. Amniotic fluid infection, inflammation, and colonization in preterm labor with intact membranes. Am. J. Obstet. Gynecol. 2014, 210, 125.e1–125.e15. [Google Scholar] [CrossRef]
- Bachkangi, P.; Taylor, A.H.; Bari, M.; Maccarrone, M.; Konje, J.C. Prediction of preterm labour from a single blood test: The role of the endocannabinoid system in predicting preterm birth in high-risk women. Eur. J. Obstet. Gynecol. Reprod. Biol. 2019, 243, 1–6. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; The PRISMA Group. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
PRISMA flow diagram.

Figure 2.
Pathways to labour. DAMPs: damage-associated molecular patterns; PAMPs: pathogen-associated molecular patterns; NF-κB: nuclear factor kappa light chain enhancer of activated B cells; cfDNA: cell-free DNA. Created with BioRender.com (accessed on 3 February 2022).
Figure 2.
Pathways to labour. DAMPs: damage-associated molecular patterns; PAMPs: pathogen-associated molecular patterns; NF-κB: nuclear factor kappa light chain enhancer of activated B cells; cfDNA: cell-free DNA. Created with BioRender.com (accessed on 3 February 2022).

Figure 3.
Investigational treatments in preterm birth (PTB). Created with BioRender.com (accessed on 3 February 2022).
Figure 3.
Investigational treatments in preterm birth (PTB). Created with BioRender.com (accessed on 3 February 2022).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Areia, A.L.; Mota-Pinto, A. Inflammation and Preterm Birth: A Systematic Review. Reprod. Med. 2022, 3, 101-111. https://0-doi-org.brum.beds.ac.uk/10.3390/reprodmed3020009
AMA Style
Areia AL, Mota-Pinto A. Inflammation and Preterm Birth: A Systematic Review. Reproductive Medicine. 2022; 3(2):101-111. https://0-doi-org.brum.beds.ac.uk/10.3390/reprodmed3020009
Chicago/Turabian StyleAreia, Ana Luísa, and Anabela Mota-Pinto. 2022. "Inflammation and Preterm Birth: A Systematic Review" Reproductive Medicine 3, no. 2: 101-111. https://0-doi-org.brum.beds.ac.uk/10.3390/reprodmed3020009