
Power Spectrum and Connectivity Analysis in EEG Recording during Attention and Creativity Performance in Children

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
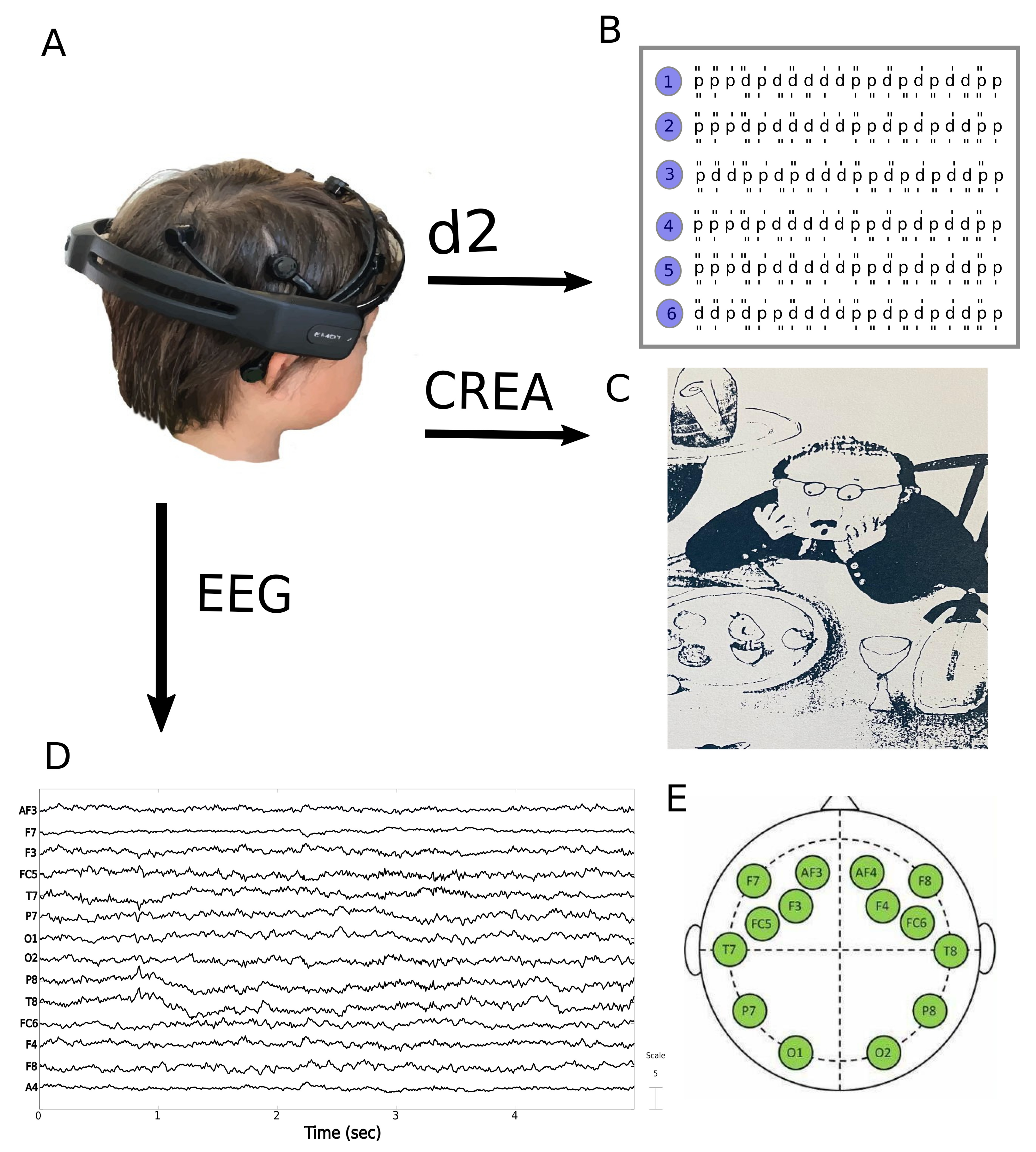
2.2. Data Acquisition
2.3. Raven Test of Progressive Matrices
2.4. Swanson, Nolan, and Pelham Rating Scale, Fourth Version (SNAP-IV)
2.5. d2 Attention Test
2.6. CREA (Creative Intelligence) Test
2.7. Data Analysis
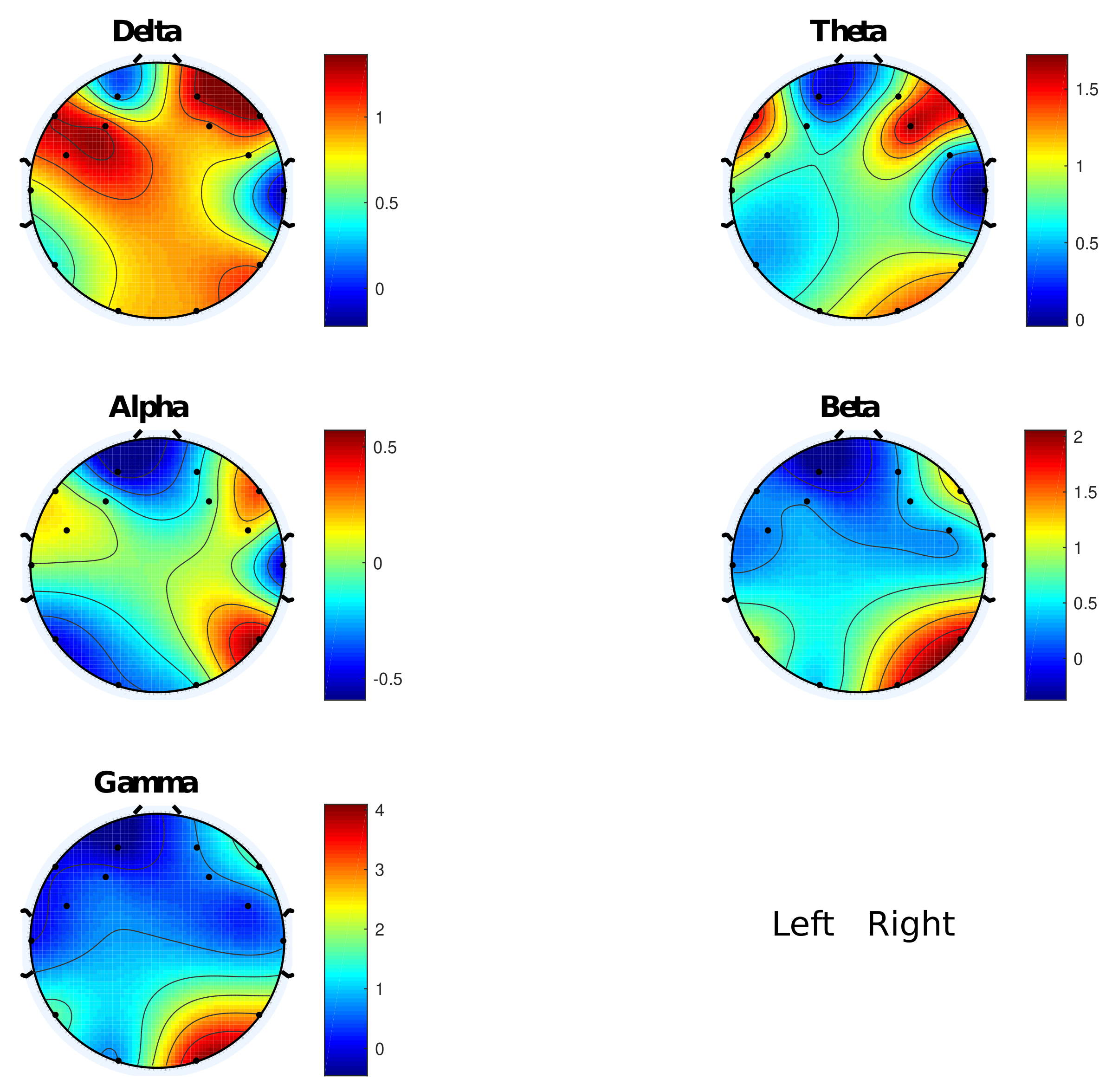
3. Results
3.1. d2 Test
3.2. CREA Test
4. Discussion
4.1. d2 Study
4.2. CREA Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brauch Lehman, E.; Naglieri, J.A.; Aquilino, S. A national study on the development of visual attention using the cognitive assessment system. J. Atten. Disord. 2010, 14, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Manly, T.; Anderson, V.; Nimmo-Smith, I.; Turner, A.; Watson, P.; Robertson, I. The differential assessment of children’s attention: The Test of Everyday Attention for Children (TEA-Ch), normative sample and ADHD performance. J. Child Psychol. Psychiatry Allied Discip. 2001, 42, 1065–1081. [Google Scholar] [CrossRef] [PubMed]
- Benedek, M.; Franz, F.; Heene, M.; Neubauer, A. Differential effects of cognitive inhibition and intelligence on creativity. Personal. Individ. Differ. 2012, 53, 480–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benedek, M.; Konen, T.; Neubauer, A. Associative abilities underlying creativity. Psychol. Aesthet. Creat. Arts 2012, 6, 273. [Google Scholar] [CrossRef]
- Amabile, T. The social psychology of creativity: A componential conceptualization. J. Personal. Soc. Psychol. 1983, 45, 357. [Google Scholar] [CrossRef]
- Csikszentmihalyi, M. The Flow Experience and Its Significance for Human Psychology; Optimal Experience: Psychological Studies of Flow in Consciousness; Csikszentmihalyi, I.S., Ed.; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Guilford, J. The structure of intellect. Psychol. Bull. 1956, 53, 267. [Google Scholar] [CrossRef]
- Mednick, S. The associative basis of the creative process. Psychol. Rev. 1962, 69, 220–232. [Google Scholar] [CrossRef] [Green Version]
- Simonton, D. Scientific Genius: A Psychology of Science; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Sternberg, R.; Lubart, T. Investing in creativity. Am. Psychol. 1996, 51, 677. [Google Scholar] [CrossRef]
- Boden, M. Creativity and artificial intelligence. Artif. Intell. 1998, 103, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Pfenninger, K.; Shubik, V. Insights into the foundation of creativity: A synthesis. Orig. Creat. 2001, 103, 213–236. [Google Scholar]
- Arán Filippetti, V.; Gutierrez, M.; Krumm, G.; Mateos, D. Convergent validity, academic correlates and age-and SES-based normative data for the d2 Test of attention in children. Appl. Neuropsychol. Child 2021, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Garaigordobil, M.; Torres, E. Evaluación de la creatividad en sus correlatos con inteligencia y rendimiento académico. Univ. Tarracon. 1996, 18, 87–101. [Google Scholar]
- Brickenkamp, R.; Zillmer, E. The d2 Test of Attention; Hogrefe & Huber: Cambridge, MA, USA, 1998. [Google Scholar]
- Lord, W.; Clarke, P. Use of the d2 Test of Attention as a Predictor of Driving Proficiency. In Driver Behaviour and Training; Routledge: Oxfordshire, UK, 2017; pp. 437–441. [Google Scholar]
- Arán Filippetti, V.; Krumm, G. A hierarchical model of cognitive flexibility in children: Extending the relationship between flexibility, creativity and academic achievement. Child Neuropsychol. 2020, 26, 770–800. [Google Scholar] [CrossRef] [PubMed]
- Krumm, G.; Filippetti, A. The contribution of executive functions to creativity in children: What is the role of crystallized and fluid intelligence? Think. Skills Creat. 2018, 29, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Gras, R.; Corbalán, B. Creatividad y estilos de personalidad: Aproximación a un perfil creativo en estudiantes universitarios. Psicol. Ann. Psychol. 2010, 26, 273–278. [Google Scholar]
- Elisondo, R.; Donolo, D. ¿Creatividad o inteligencia? That is not the question. An. Psicol. Ann. Psychol. 2010, 26, 220–225. [Google Scholar]
- Berná Corbalán, J.; Gras, R. El genio en una botella. El test CREA, las preguntas y la creatividad. Introducción al monográfico “El test CREA, inteligencia creativa”. An. Psicol. Ann. Psychol. 2010, 26, 197–205. [Google Scholar]
- Perry, N.; Swingler, M.; Calkins, S.; Bell, M. Neurophysiological correlates of attention behavior in early infancy: Implications for emotion regulation during early childhood. J. Exp. Child Psychol. 2016, 142, 245–261. [Google Scholar] [CrossRef] [Green Version]
- Cheung, C.; McLoughlin, G.; Brandeis, D.; Banaschewski, T.; Asherson, P.; Kuntsi, J. Neurophysiological correlates of attentional fluctuation in attention-deficit/hyperactivity disorder. Brain Topogr. 2017, 30, 320–332. [Google Scholar] [CrossRef] [Green Version]
- Taylor, M.; Baldeweg, T. Application of EEG, ERP and intracranial recordings to the investigation of cognitive functions in children. Dev. Sci. 2002, 5, 318–334. [Google Scholar] [CrossRef]
- Mueller, V.; Brehmer, Y.; Von Oertzen, T.; Li, S.C.; Lindenberger, U. Electrophysiological correlates of selective attention: A lifespan comparison. BMC Neurosci. 2008, 9, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardi, G.; Betta, M.; Ricciardi, E.; Pietrini, P.; Tononi, G.; Siclari, F. Regional delta waves in human rapid eye movement sleep. J. Neurosci. 2019, 39, 2686–2697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Güntekin, B.; Başar, E. Review of evoked and event-related delta responses in the human brain. Int. J. Psychophysiol. 2016, 103, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orekhova, E.; Stroganova, T.; Posikera, I.; Elam, M. EEG theta rhythm in infants and preschool children. Clin. Neurophysiol. 2006, 117, 1047–1062. [Google Scholar] [CrossRef]
- Lagopoulos, J.; Xu, J.; Rasmussen, I.; Vik, A.; Malhi, G.S.; Eliassen, C.; Arntsen, I.; Sæther, J.; Hollup, S.; Holen, A.; et al. Increased theta and alpha EEG activity during nondirective meditation. J. Altern. Complement. Med. 2009, 15, 1187–1192. [Google Scholar] [CrossRef]
- Wienbruch, C.; Paul, I.; Bauer, S.; Kivelitz, H. The influence of methylphenidate on the power spectrum of ADHD children—An MEG study. BMC Psychiatry 2005, 5, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Womelsdorf, T.; Fries, P. The role of neuronal synchronization in selective attention. Curr. Opin. Neurobiol. 2007, 17, 154–160. [Google Scholar] [CrossRef]
- Benasich, A.; Gou, Z.; Choudhury, N.; Harris, K. Early cognitive and language skills are linked to resting frontal gamma power across the first 3 years. Behav. Brain Res. 2008, 195, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Engel, A.; Fries, P.; Singer, W. Dynamic predictions: Oscillations and synchrony in top-down processing. Nat. Rev. Neurosci. 2001, 2, 704–716. [Google Scholar] [CrossRef]
- Fell, J.; Fernandez, G.; Klaver, P.; Elger, C.E.; Fries, P. Is synchronized neuronal gamma activity relevant for selective attention? Brain Res. Rev. 2003, 42, 265–272. [Google Scholar] [CrossRef]
- Paul, R.; Richard Clark, C.; Lawrence, J.; Goldberg, E.; Williams, L.; Cooper, N.; Cohen, R.A.; Brickman, A.; Gordon, E. Age-dependent change in executive function and gamma 40 Hz phase synchrony. J. Integr. Neurosci. 2005, 4, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Barry, R.; Clarke, A.; Hajos, M.; McCarthy, R.; Selikowitz, M.; Dupuy, F. Resting-state EEG gamma activity in children with attention-deficit/hyperactivity disorder. Clin. Neurophysiol. 2010, 121, 1871–1877. [Google Scholar] [CrossRef] [PubMed]
- Yordanova, J.; Banaschewski, T.; Kolev, V.; Woerner, W.; Rothenberger, A. Abnormal early stages of task stimulus processing in children with attention-deficit hyperactivity disorder-evidence from event-related gamma oscillations. Clin. Neurophysiol. 2001, 112, 1096–1108. [Google Scholar] [CrossRef]
- Hebert, R.; Lehmann, D.; Tan, G.; Travis, F.; Arenander, A. Enhanced EEG alpha time-domain phase synchrony during Transcendental Meditation: Implications for cortical integration theory. Signal Process. 2005, 85, 213–2232. [Google Scholar] [CrossRef]
- Xie, W.; Mallin, B.M.; Richards, J. Development of infant sustained attention and its relation to EEG oscillations: An EEG and cortical source analysis study. Dev. Sci. 2018, 21, e12562. [Google Scholar] [CrossRef]
- Thut, G.; Nietzel, A.; Brandt, S.A.; Pascual-Leone, A. α-Band electroencephalographic activity over occipital cortex indexes visuospatial attention bias and predicts visual target detection. J. Neurosci. 2006, 26, 9494–9502. [Google Scholar] [CrossRef] [Green Version]
- Tucker, D.; Dawson, S.; Roth, D.L.; Penland, J. Regional changes in EEG power and coherence during cognition: Intensive study of two individuals. Behav. Neurosci. 1985, 99, 564. [Google Scholar] [CrossRef]
- Klimesch, W.; Doppelmayr, M.; Schimke, H.P.T. Alpha frequency, reaction time, and the speed of processing information. J. Clin. Neurophysiol. 1996, 13, 511–518. [Google Scholar] [CrossRef]
- Arden, R.; Chavez, R.S.; Grazioplene, R.; Jung, R. Neuroimaging creativity: A psychometric view. Behav. Brain Res. 2010, 214, 143–156. [Google Scholar] [CrossRef]
- Dietrich, A.; Kanso, R. A review of EEG, ERP, and neuroimaging studies of creativity and insight. Psychol. Bull. 2010, 136, 822. [Google Scholar] [CrossRef]
- Bazanova, O.; Aftanas, L. Individual measures of electroencephalogram alpha activity and non-verbal creativity. Neurosci. Behav. Physiol. 2008, 38, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Danko, S.; Shemyakina, N.; Nagornova, Z.; Starchenko, M. Comparison of the effects of the subjective complexity and verbal creativity on EEG spectral power parameters. Hum. Physiol. 2009, 35, 381–383. [Google Scholar] [CrossRef]
- Fink, A.; Grabner, R.; Benedek, M.; Reishofer, G.; Hauswirth, V.; Fally, M.; Neuper, C.; Ebner, F.; Neubauer, A. The creative brain: Investigation of brain activity during creative problem solving by means of EEG and fMRI. Hum. Brain Mapp. 2009, 30, 734–748. [Google Scholar] [CrossRef]
- Fink, A.; Neubauer, A. EEG alpha oscillations during the performance of verbal creativity tasks: Differential effects of sex and verbal intelligence. Int. J. Psychophysiol. 2006, 62, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Fink, A.; Neubauer, A. Eysenck meets Martindale: The relationship between extraversion and originality from the neuroscientific perspective. Personal. Individ. Differ. 2008, 44, 299–310. [Google Scholar] [CrossRef]
- Fink, A.; Benedek, M. EEG alpha power and creative ideation. Neurosci. Biobehav. Rev. 2014, 44, 111–123. [Google Scholar] [CrossRef] [Green Version]
- Grabner, R.; Fink, A.; Neubauer, A.C. Brain correlates of self-rated originality of ideas: Evidence from event-related power and phase-locking changes in the EEG. Behav. Neurosci. 2007, 121, 224. [Google Scholar] [CrossRef] [Green Version]
- Jaušovec, N. Differences in cognitive processes between gifted, intelligent, creative, and average individuals while solving complex problems: An EEG study. Intelligence 2000, 28, 213–237. [Google Scholar] [CrossRef]
- Pidgeon, L.M.; Grealy, M.; Duffy, A.H.; Hay, L.; McTeague, C.; Vuletic, T.; Coyle, D.; Gilbert, S. Functional neuroimaging of visual creativity: A systematic review and meta-analysis. Brain Behav. 2016, 6, e00540. [Google Scholar] [CrossRef] [Green Version]
- Rominger, C.; Papousek, I.; Perchtold, C.; Benedek, M.; Weiss, E.; Schwerdtfeger, A.; Fink, A. Creativity is associated with a characteristic U-shaped function of alpha power changes accompanied by an early increase in functional coupling. Cogn. Affect. Behav. Neurosci. 2019, 19, 1012–1021. [Google Scholar] [CrossRef] [Green Version]
- Stevens, C., Jr.; Zabelina, D. Creativity comes in waves: An EEG-focused exploration of the creative brain. Curr. Opin. Behav. Sci. 2019, 27, 154–162. [Google Scholar] [CrossRef] [Green Version]
- Razumnikova, O. Gender differences in hemispheric organization during divergent thinking: An EEG investigation in human subjects. Neurosci. Lett. 2004, 362, 193–195. [Google Scholar] [CrossRef] [PubMed]
- Razumnikova, O.; Volf, N.; Tarasova, I. Strategy and results: Sex differences in electrographic correlates of verbal and figural creativity. Hum. Physiol. 2009, 35, 285–294. [Google Scholar] [CrossRef]
- Volf, N.; Tarasova, I. The relationships between EEG θ and β oscillations and the level of creativity. Hum. Physiol. 2010, 36, 132–138. [Google Scholar] [CrossRef]
- Jauk, E.; Benedek, M.; Neubauer, A. Tackling creativity at its roots: Evidence for different patterns of EEG alpha activity related to convergent and divergent modes of task processing. Int. J. Psychophysiol. 2012, 84, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Benedek, M. The Neuroscience of Creative Idea Generation. In Exploring Transdisciplinarity in Art and Sciences; Springer: Berlin, Germany, 2018; pp. 31–48. [Google Scholar]
- Benedek, M.; Bergner, S.; Könen, T.; Fink, A.; Neubauer, A. EEG alpha synchronization is related to top-down processing in convergent and divergent thinking. Neuropsychologia 2011, 49, 3505–3511. [Google Scholar] [CrossRef] [Green Version]
- Stevens, C., Jr.; Zabelina, D. Classifying creativity: Applying machine learning techniques to divergent thinking EEG data. NeuroImage 2020, 219, 116990. [Google Scholar] [CrossRef]
- Nobukawa, S.; Yamanishi, T.; Ueno, K.; Mizukami, K.; Nishimura, H.; Takahashi, T. High phase synchronization in alpha band activity in older subjects with high creativity. Front. Hum. Neurosci. 2020, 14, 420. [Google Scholar] [CrossRef]
- Neubauer, A.; Fink, A.; Grabner, R. Sensitivity of alpha band ERD to individual differences in cognition. Prog. Brain Res. 2006, 159, 167–178. [Google Scholar]
- Neubauer, A.C.; Fink, A. Intelligence and neural efficiency. Neurosci. Biobehav. Rev. 2009, 33, 1004–1023. [Google Scholar] [CrossRef]
- Shemyakina, N.; Danko, S.; Nagornova, Z.; Starchenko, M.; Bechtereva, N. Changes in the power and coherence spectra of the EEG rhythmic components during solution of a verbal creative task of overcoming a stereotype. Hum. Physiol. 2007, 33, 524–530. [Google Scholar] [CrossRef]
- Boot, N.; Baas, M.; Mühlfeld, E.; de Dreu, C.; van Gaal, S. Widespread neural oscillations in the delta band dissociate rule convergence from rule divergence during creative idea generation. Neuropsychologia 2017, 104, 8–17. [Google Scholar] [CrossRef]
- Wokke, M.; Padding, L.; Ridderinkhof, K. Creative Brains Show Reduced Mid Frontal Theta. bioRxiv 2019, 370494. [Google Scholar]
- Cavanagh, J.F.; Frank, M. Frontal theta as a mechanism for cognitive control. Trends Cogn. Sci. 2014, 18, 414–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fink, A.; Benedek, M. The Creative brain: Brain correlates underlying the generation of original ideas. Neurosci. Creat. 2013, 1, 207–232. [Google Scholar]
- Jin, S.H.; Kwon, Y.J.; Jeong, J.S.; Kwon, S.W.; Shin, D.H. Differences in brain information transmission between gifted and normal children during scientific hypothesis generation. Brain Cogn. 2006, 62, 191–197. [Google Scholar] [CrossRef]
- Privodnova, E.; Volf, N. Features of temporal dynamics of oscillatory brain activity during creative problem solving in young and elderly adults. Hum. Physiol. 2016, 42, 469–475. [Google Scholar] [CrossRef]
- Privodnova, E.; Volf, N.; Knyazev, G. Specific features of the oscillatory brain activity during the final stage of creative problem solving in young and elderly subjects. Hum. Physiol. 2017, 43, 241–247. [Google Scholar] [CrossRef]
- Torrance, E.; Ball, O.; Safter, H. Torrance Tests of Creative Thinking: Streamlined Scoring Guide. Figural A and B; Scholastic Testing Service: Bensenville, IL, USA, 1992. [Google Scholar]
- Corbalán Berná, J.; Martínez Zaragoza, F.; Donolo, D.; Alonso Monreal, C.; Tejerina Arreal, M.; Limiñana Gras, R. CREA. In Inteligencia Creativa. Una Medida Cognitiva de la Creatividad; TEA Ediciones: Madrid, Spain, 2003. [Google Scholar]
- Emotiv Epoc+ Emotiv. Available online: https://www.emotiv.com/epoc/ (accessed on 8 September 2021).
- Bobrov, P.; Frolov, A.; Cantor, C.; Fedulova, I.; Bakhnyan, M.; Zhavoronkov, A. Brain-computer interface based on generation of visual images. PLoS ONE 2011, 6, e20674. [Google Scholar] [CrossRef]
- Amjad, I.; Toor, H.; Niazi, I.K.; Afzal, H.; Jochumsen, M.; Shafique, M.; Allen, K.; Haavik, H.; Ahmed, T. Therapeutic effects of aerobic exercise on EEG parameters and higher cognitive functions in mild cognitive impairment patients. Int. J. Neurosci. 2019, 129, 551–562. [Google Scholar] [CrossRef]
- Raven, J.; Court, J.; Raven, J. Test de Matrices Progresivas. Escalas Coloreada, General y Avanzada. Manual; Paidós: Buenos Aires, Argentina, 2008. [Google Scholar]
- Grañana, N.; Richaudeau, A.; Gorriti, C.; O’Flaherty, M.; Scotti, M.E.; Sixto, L.; Allegri, R.; Fejerman, N. Evaluación de déficit de atención con hiperactividad: La escala SNAP IV adaptada a la Argentina. Rev. Panam. Salud PÚBlica 2011, 29, 344–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brickenkamp, R. d2, Test de Atención (Revisada y Ampliada), 2nd ed.; TEA Ediciones: Madrid, Spain, 2004. [Google Scholar]
- Elisondo, R.; Donolo, D. Los estímulos en un test de creatividad. Incidencias según género, edad y escolaridad. Bol. Psicol. 2011, 101, 51–65. [Google Scholar]
- Torrance, E. Torrance Tests of Creative Thinking: Manual for Scoring and Interpreting Results; Scholastic Testing Service: Bensenville, IL, USA, 1990. [Google Scholar]
- Stam, C.; Nolte, G.; Daffertshofer, A. Phase lag index: Assessment of functional connectivity from multi channel EEG and MEG with diminished bias from common sources. Hum. Brain Mapp. 2007, 28, 1178–1193. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M. Analyzing Neural Time Series Data: Theory and Practice; MIT Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Carter, G.C. Coherence and time delay estimation. Proc. IEEE 1987, 75, 236–255. [Google Scholar] [CrossRef]
- Lachaux, J.P.; Rodriguez, E.; Martinerie, J.; Varela, F. Measuring phase synchrony in brain signals. Hum. Brain Mapp. 1999, 8, 194–208. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, J.; Petsche, H. Phase synchrony analysis of EEG during music perception reveals changes in functional connectivity due to musical expertise. Signal Process. 2005, 85, 2161–2177. [Google Scholar] [CrossRef]
- Guevara Erra, R.; Mateos, D.; Wennberg, R.; Perez Velazquez, J. Statistical mechanics of consciousness: Maximization of information content of network is associated with conscious awareness. Phys. Rev. E 2016, 94, 052402. [Google Scholar] [CrossRef] [Green Version]
- Mateos, D.; Wennberg, R.; Guevara, R.; Perez Velazquez, J. Consciousness as a global property of brain dynamic activity. Phys. Rev. E 2017, 96, 062410. [Google Scholar] [CrossRef] [Green Version]
- Dressler, O.; Schneider, G.; Stockmanns, G.; Kochs, E. Awareness and the EEG power spectrum: Analysis of frequencies. Br. J. Anaesth. 2004, 93, 806–809. [Google Scholar] [CrossRef] [Green Version]
- Doesburg, S.; Roggeveen, A.; Kitajo, K.; Ward, L. Large-scale gamma-band phase synchronization and selective attention. Cereb. Cortex 2008, 18, 386–396. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, E.; George, N.; Lachaux, J.P.; Martinerie, J.; Renault, B.; Varela, F.J. Perception’s shadow: Long-distance synchronization of human brain activity. Nature 1999, 397, 430–433. [Google Scholar] [CrossRef] [PubMed]
- Aoki, F.; Fetz, E.; Shupe, L.; Lettich, E.; Ojemann, G. Increased gamma-range activity in human sensorimotor cortex during performance of visuomotor tasks. Clin. Neurophysiol. 1999, 110, 524–537. [Google Scholar] [CrossRef]
- Ferraracci, J.; Anzalone, C.; Bridges, R.; Moore, R.; Decker, S. QEEG correlates of cognitive processing speed in children and adolescents with traumatic brain injuries. Appl. Neuropsychol. Child 2019, 10, 1–11. [Google Scholar] [CrossRef]
- González-Garrido, A.; Gómez-Velázquez, F.; Salido-Ruiz, R.; Espinoza-Valdez, A.; Vélez-Pérez, H.; Romo-Vazquez, R.; Gallardo-Moreno, G.; Ruiz-Stovel, V.; Martínez-Ramos, A.; Berumen, G. The analysis of EEG coherence reflects middle childhood differences in mathematical achievement. Brain Cogn. 2018, 124, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Marrufo, M.; Vaquero, E.; Cardoso, M.; Gomez, C.M. Temporal evolution of α and β bands during visual spatial attention. Cogn. Brain Res. 2001, 12, 315–320. [Google Scholar] [CrossRef]
- Fleck, J.; Kuti, J.; Brown, J.; Mahon, J.; Gayda-Chelder, C. Frontal-posterior coherence and cognitive function in older adults. Int. J. Psychophysiol. 2016, 110, 217–230. [Google Scholar] [CrossRef]
- Shaw, J. Intention as a component of the alpha-rhythm response to mental activity. Int. J. Psychophysiol. 1996, 24, 7–23. [Google Scholar] [CrossRef]
- Clarke, A.; Barry, R.; Bond, D.; McCarthy, R.; Selikowitz, M. Effects of stimulant medications on the EEG of children with attention-deficit/hyperactivity disorder. Psychopharmacology 2002, 164, 277–284. [Google Scholar] [CrossRef]
- Satterfield, J.; Cantwell, D. CNS function and response to methylphenidate in hyperactive children. Psychopharmacol. Bull. 1974, 10, 36–37. [Google Scholar]
- Loo, S.; Teale, P.; Reite, M. EEG correlates of methylphenidate response among children with ADHD: A preliminary report. Biol. Psychiatry 1999, 45, 1657–1660. [Google Scholar] [CrossRef]
- Ahani, A.; Wahbeh, H.; Nezamfar, H.; Miller, M.; Erdogmus, D.; Oken, B. Quantitative change of EEG and respiration signals during mindfulness meditation. J. Neuroeng. Rehabil. 2014, 11, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aftanas, L.; Golocheikine, S. Human anterior and frontal midline theta and lower alpha reflect emotionally positive state and internalized attention: High-resolution EEG investigation of meditation. Neurosci. Lett. 2001, 310, 57–60. [Google Scholar] [CrossRef]
- Bagherzadeh, Y.; Baldauf, D.; Pantazis, D.; Desimone, R. Alpha synchrony and the neurofeedback control of spatial attention. Neuron 2020, 105, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Roux, F.; Uhlhaas, P. Working memory and neural oscillations: Alpha–gamma versus theta–gamma codes for distinct WM information? Trends Cogn. Sci. 2014, 18, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Brito, N.; Fifer, W.; Myers, M.; Elliott, A.; Noble, K. Associations among family socioeconomic status, EEG power at birth, and cognitive skills during infancy. Dev. Cogn. Neurosci. 2016, 19, 144–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gou, Z.; Choudhury, N.; Benasich, A. Resting frontal gamma power at 16, 24 and 36 months predicts individual differences in language and cognition at 4 and 5 years. Behav. Brain Res. 2011, 220, 263–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarullo, A.; Obradović, J.; Keehn, B.; Rasheed, M.; Siyal, S.; Nelson, C.; Yousafzai, A. Gamma power in rural Pakistani children: Links to executive function and verbal ability. Dev. Cogn. Neurosci. 2017, 26, 1–8. [Google Scholar] [CrossRef]
- Njiokiktjien, C.; De Rijke, W.; Jonkman, E. Children with nonverbal learning disabilities (NLD): Coherence values in the resting state may reflect hypofunctional long distance connections in the right hemisphere. Hum. Physiol. 2001, 27, 523–528. [Google Scholar] [CrossRef]
- Drechsler, R.; Straub, M.; Doehnert, M.; Heinrich, H.; Steinhausen, H.C.; Brandeis, D. Controlled evaluation of a neurofeedback training of slow cortical potentials in children with attention deficit/hyperactivity disorder (ADHD). Behav. Brain Funct. 2007, 3, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Moore, A.; Malinowski, P. Meditation, mindfulness and cognitive flexibility. Conscious. Cogn. 2009, 18, 176–186. [Google Scholar] [CrossRef]
- Budde, H.; Voelcker-Rehage, C.; Pietraßyk-Kendziorra, S.; Ribeiro, P.; Tidow, G. Acute coordinative exercise improves attentional performance in adolescents. Neurosci. Lett. 2008, 441, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Malmir, N.; Khaleghi, A.; Aminiorani, M. Comparison of sensorimotor rhythm (SMR) and beta training on selective attention and symptoms in children with attention deficit/hyperactivity disorder (ADHD): A trend report. Iran. J. Psychiatry 2015, 10, 165. [Google Scholar]
- Horan, R. The neuropsychological connection between creativity and meditation. Creat. Res. J. 2009, 21, 199–222. [Google Scholar] [CrossRef]
- Aftanas, L.; Reva, N.; Varlamov, A.; Pavlov, S.; Makhnev, V. Analysis of evoked EEG synchronization and desynchronization in conditions of emotional activation in humans: Temporal and topographic characteristics. Neurosci. Behav. Physiol. 2004, 34, 859–867. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, A. The cognitive neuroscience of creativity. Psychon. Bull. Rev. 2004, 11, 1011–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knight, R. Contribution of human hippocampal region to novelty detection. Nature 1996, 383, 256–259. [Google Scholar] [CrossRef] [PubMed]
- Hassabis, D.; Kumaran, D.; Maguire, E. Using imagination to understand the neural basis of episodic memory. J. Neurosci. 2007, 27, 14365–14374. [Google Scholar] [CrossRef]
- Hanslmayr, S.; Aslan, A.; Staudigl, T.; Klimesch, W.; Herrmann, C.; Bäuml, K.H. Prestimulus oscillations predict visual perception performance between and within subjects. Neuroimage 2007, 37, 1465–1473. [Google Scholar] [CrossRef]
- Pulvermüller, F.; Birbaumer, N.; Lutzenberger, W.; Mohr, B. High-frequency brain activity: Its possible role in attention, perception and language processing. Prog. Neurobiol. 1997, 52, 427–445. [Google Scholar] [CrossRef]
- Wróbel, A. Beta activity: A carrier for visual attention. Acta Neurobiol. Exp. 2000, 60, 247–260. [Google Scholar]
- Lalo, E.; Gilbertson, T.; Doyle, L.; Di Lazzaro, V.; Cioni, B.; Brown, P. Phasic increases in cortical beta activity are associated with alterations in sensory processing in the human. Exp. Brain Res. 2007, 177, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Pfurtscheller, G.; Andrew, C. Event-related changes of band power and coherence: Methodology and interpretation. J. Clin. Neurophysiol. 1999, 16, 512. [Google Scholar] [CrossRef] [PubMed]
- Polunina, A.; Davydov, D. EEG correlates of Wechsler adult intelligence scale. Int. J. Neurosci. 2006, 116, 1231–1248. [Google Scholar] [CrossRef]
- Kaiser, J.; Lutzenberger, W. Induced gamma-band activity and human brain function. Neuroscientist 2003, 9, 475–484. [Google Scholar] [CrossRef]
- Pulvermüller, F.; Keil, A.; Elbert, T. High-frequency brain activity: Perception or active memory? Trends Cogn. Sci. 1999, 3, 250–252. [Google Scholar] [CrossRef] [Green Version]
- Zabelina, D.; Ganis, G. Creativity and cognitive control: Behavioral and ERP evidence that divergent thinking, but not real-life creative achievement, relates to better cognitive control. Neuropsychologia 2018, 118, 20–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petsche, H. Approaches to verbal, visual and musical creativity by EEG coherence analysis. Int. J. Psychophysiol. 1996, 24, 145–159. [Google Scholar] [CrossRef]
- Bechtereva, N.; Nagornova, Z. Changes in EEG coherence during tests for nonverbal (figurative) creativity. Hum. Physiol. 2007, 33, 515–523. [Google Scholar] [CrossRef]
- Jung-Beeman, M.; Bowden, E.; Haberman, J.; Frymiare, J.; Arambel-Liu, S.; Greenblatt, R.; Reber, P.; Kounios, J.; Dehaene, S. Neural activity when people solve verbal problems with insight. PLoS Biol. 2004, 2, e97. [Google Scholar] [CrossRef] [Green Version]
- Ritter, S.; Abbing, J.; Van Schie, H. Eye-closure enhances creative performance on divergent and convergent creativity tasks. Front. Psychol. 2018, 9, 1315. [Google Scholar] [CrossRef]
- Sviderskaya, N. The EEG spatial pattern and psychophysiological characteristics of divergent and convergent thinking in humans. Hum. Physiol. 2011, 37, 31–38. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| M ± SD or % | |
|---|---|
| Age (M ± SD) | |
| Gender (% girls) | |
| Father educational level | |
| Mother educational level | |
| Gestational age (<37 weeks) | full-term |
| Weight at birth | (0.41) |
| Breastfeeding | |
| Exclusive | |
| Mixed | |
| Dominant hand | right-handed |
| Foreign language study | |
| Years studying a foreign language | |
| Extracurricular activities | |
| Soccer or volleyball | |
| Skate or dance school | |
| Choir combined with some sport | 20 |
| Years practicing extracurricular activities | |
| Raven (percentile) | |
| SNAP-IV | |
| Inattention | |
| Hyperactivity–impulsivity | |
| Combined ADHD | |
| d2 test | |
| Total efficiency of the task | |
| Concentration performance | |
| Fluctuation rate | |
| CREA (percentile) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mateos, D.M.; Krumm, G.; Arán Filippetti, V.; Gutierrez, M. Power Spectrum and Connectivity Analysis in EEG Recording during Attention and Creativity Performance in Children. NeuroSci 2022, 3, 347-365. https://0-doi-org.brum.beds.ac.uk/10.3390/neurosci3020025
Mateos DM, Krumm G, Arán Filippetti V, Gutierrez M. Power Spectrum and Connectivity Analysis in EEG Recording during Attention and Creativity Performance in Children. NeuroSci. 2022; 3(2):347-365. https://0-doi-org.brum.beds.ac.uk/10.3390/neurosci3020025
Chicago/Turabian StyleMateos, Diego M., Gabriela Krumm, Vanessa Arán Filippetti, and Marisel Gutierrez. 2022. "Power Spectrum and Connectivity Analysis in EEG Recording during Attention and Creativity Performance in Children" NeuroSci 3, no. 2: 347-365. https://0-doi-org.brum.beds.ac.uk/10.3390/neurosci3020025