
Immunomodulatory and Antiviral Effects of Macroalgae Sulphated Polysaccharides: Case Studies Extend Knowledge on Their Importance in Enhancing Shellfish Health, and the Control of a Global Viral Pathogen Ostreid Herpesvirus-1 microVar

 ,
,  ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Effects of (a) Intact Seaweed and (b) Extracted Derivatives on Pacific Oyster Crassostrea Gigas Performance and Pathogen Development
2.2. Immunological Effects of Naturally Extracted Sulphated Polysaccharides from a Red Seaweed Gracilaria Fisheri on Shrimp (European Common Prawn) Palaemon Serratus
2.3. Statistics
2.3.1. Crassostrea gigas Exposed to Intact Seaweed
2.3.2. Crassostrea gigas Exposed to Extracted Derivatives
2.3.3. Palaemon serratus Exposed to Extracted Sulphated Galactans
3. Results and Discussion
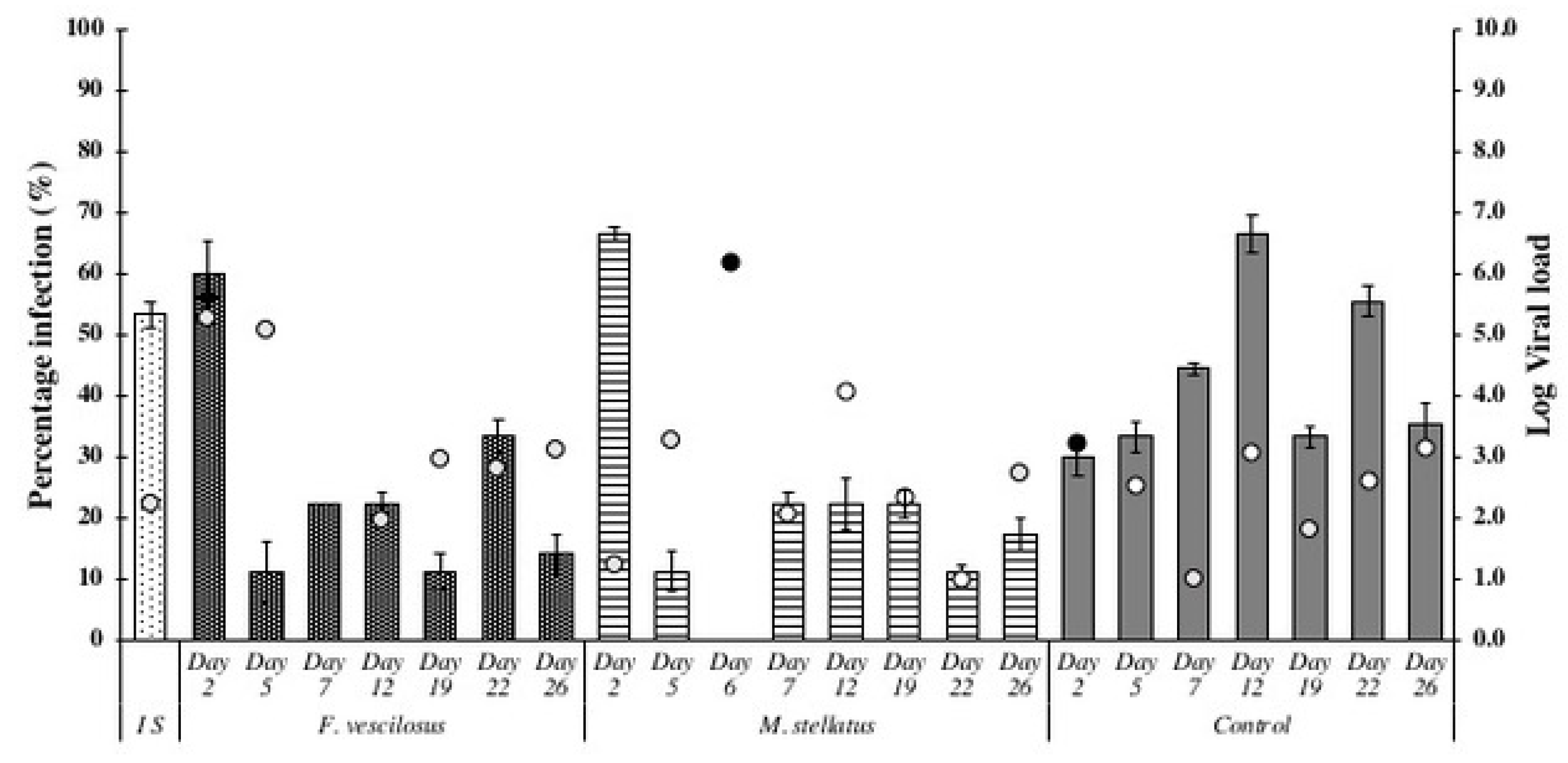
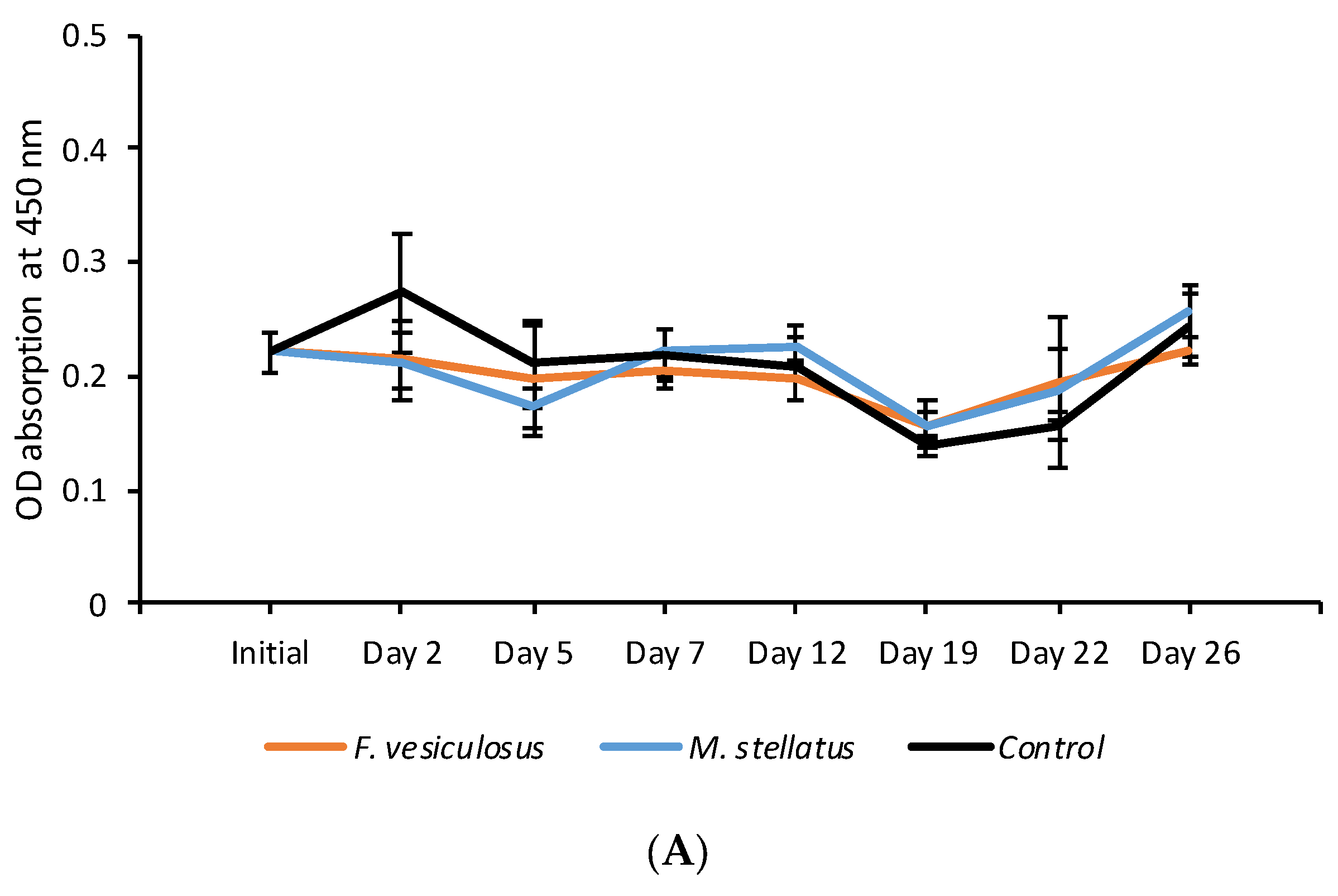
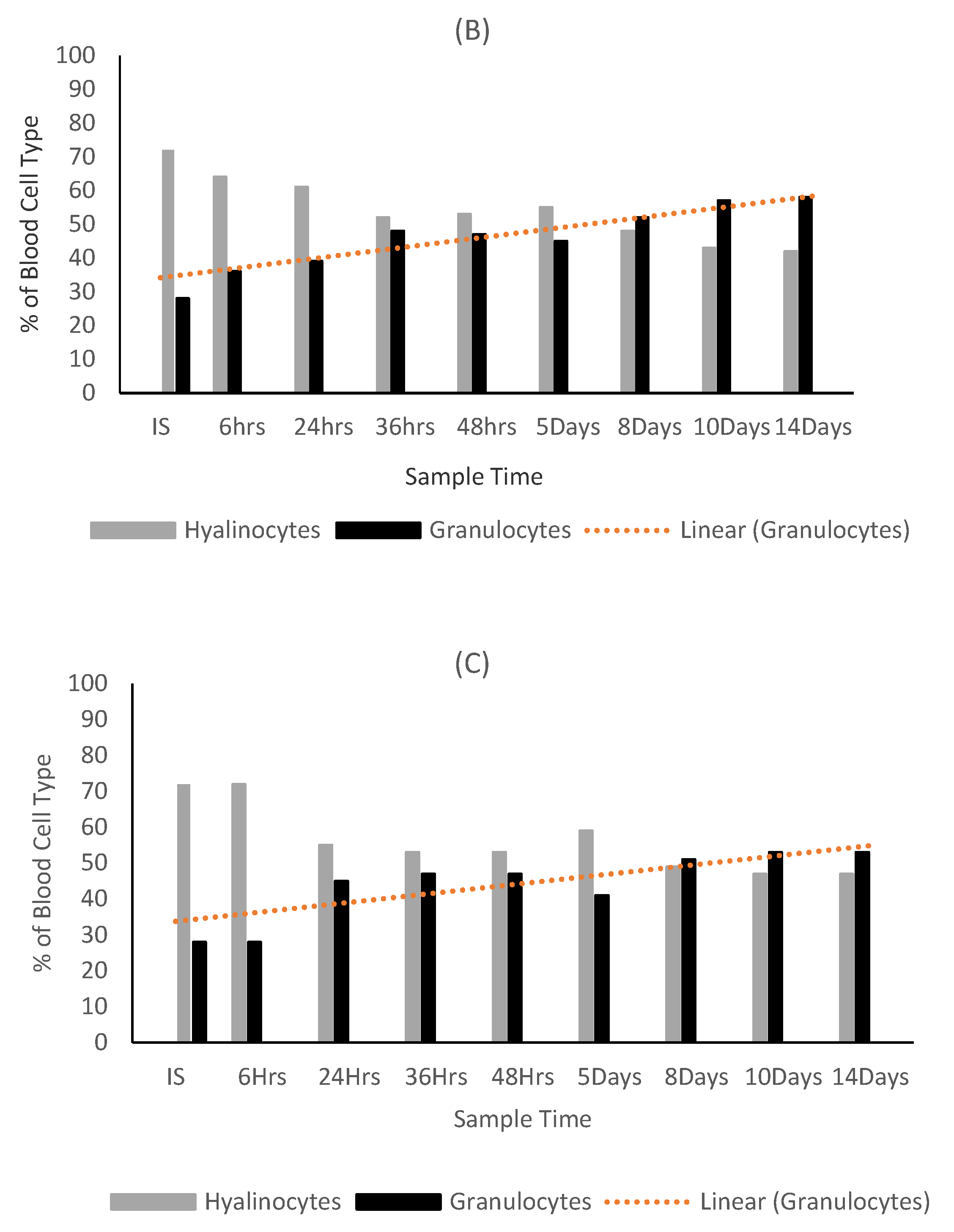
3.1. Exposure of Crassostrea Gigas to Intact Seaweed Species Fucus Vesiculosus and Mastocarpus Stellatus
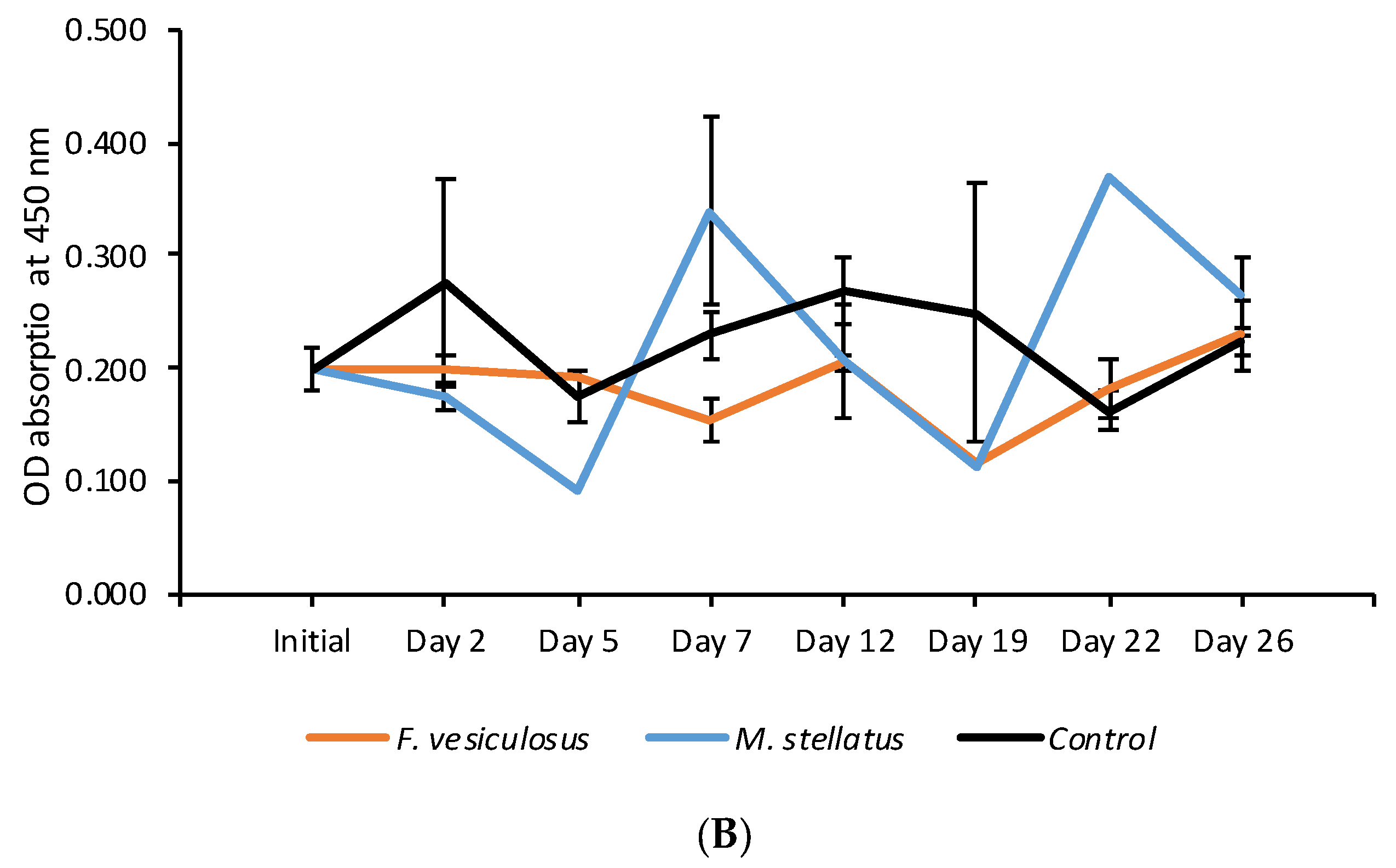
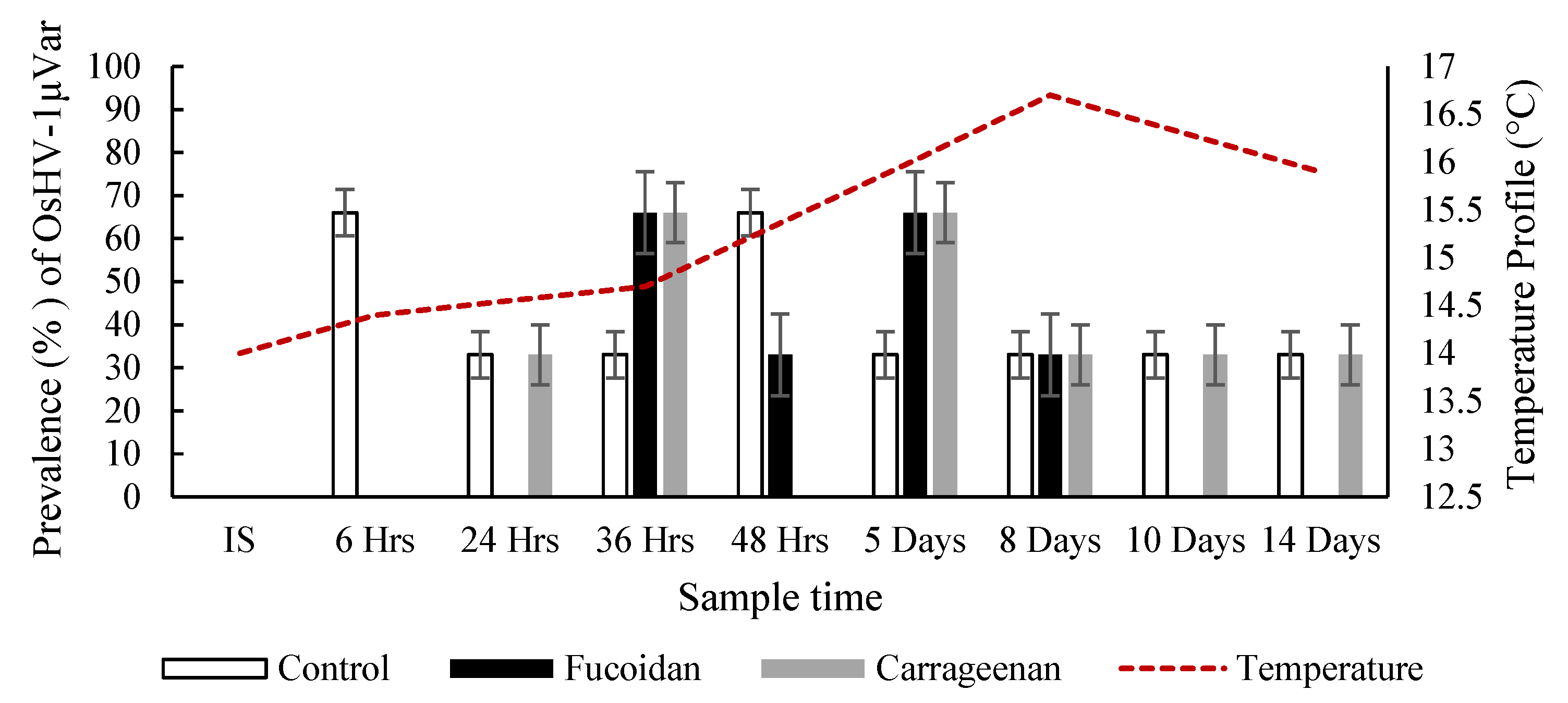
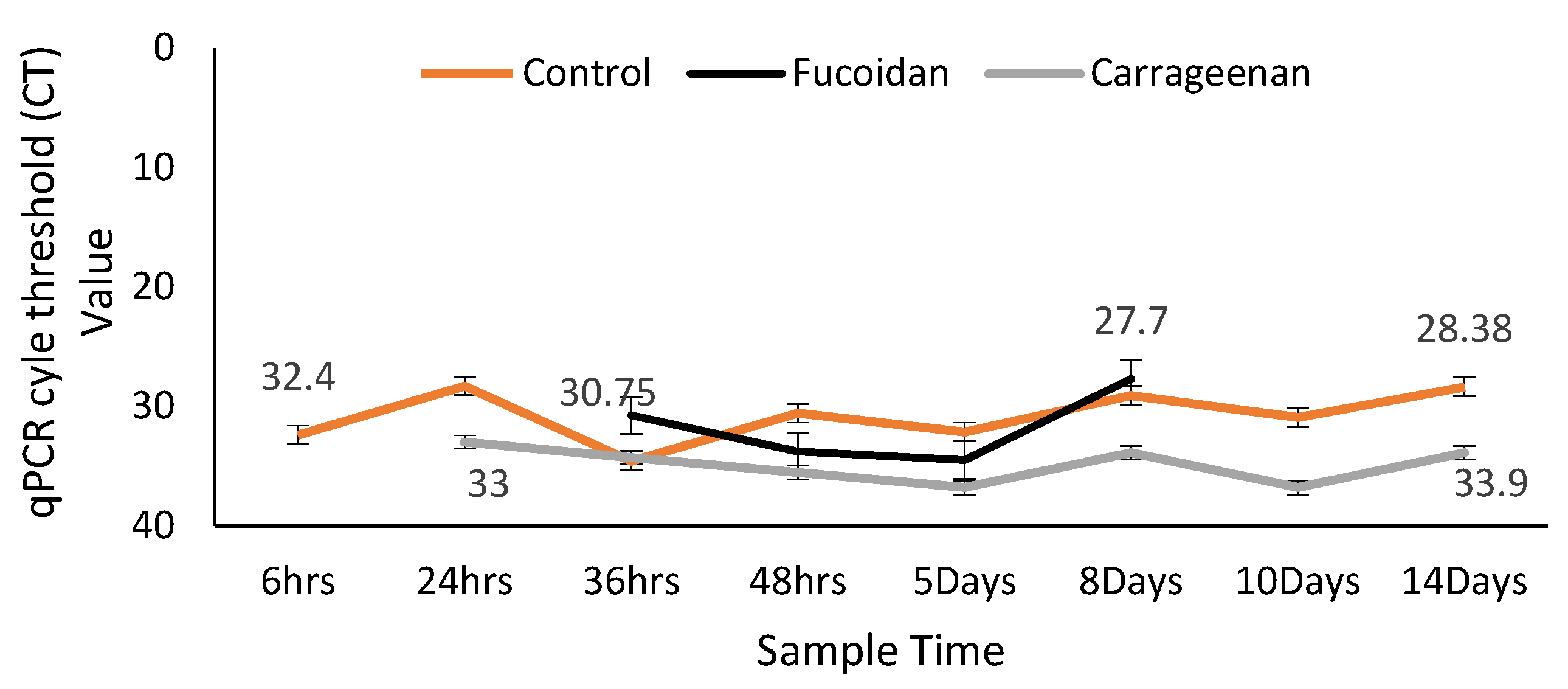
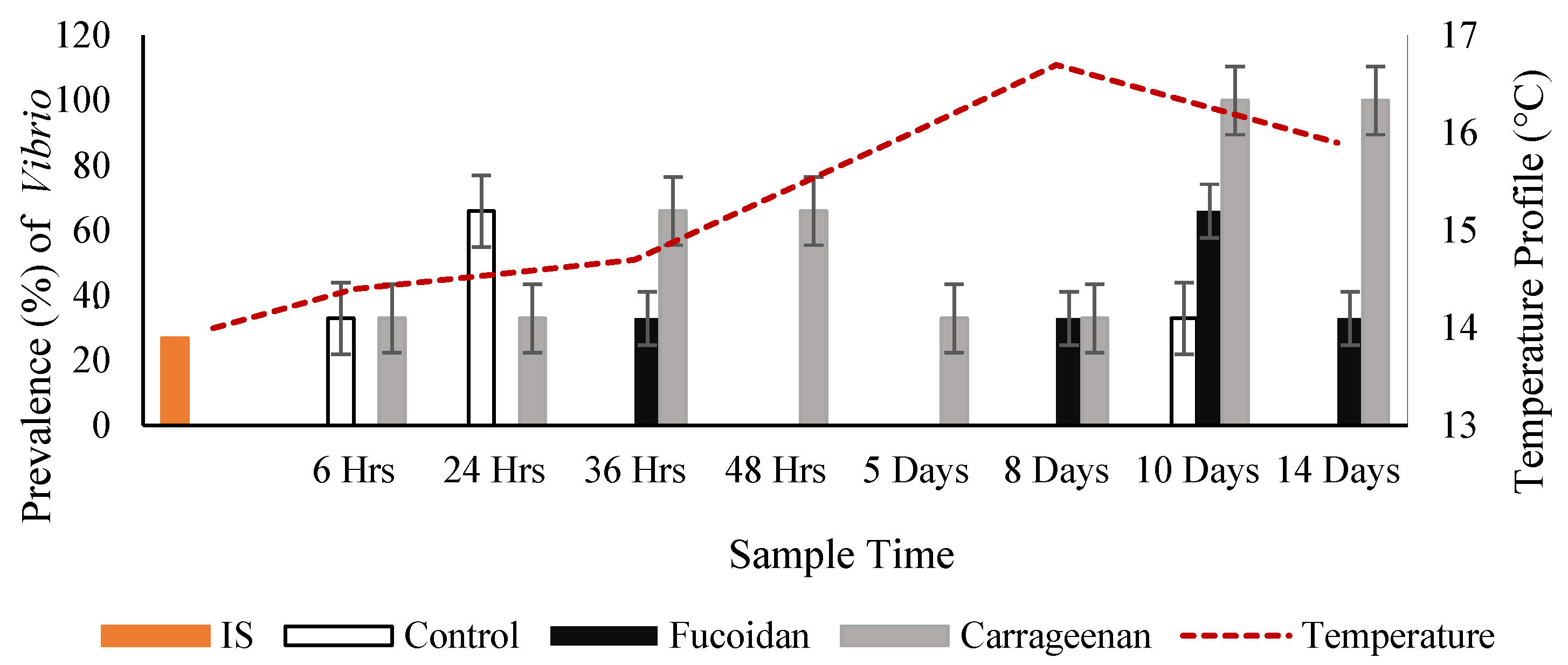
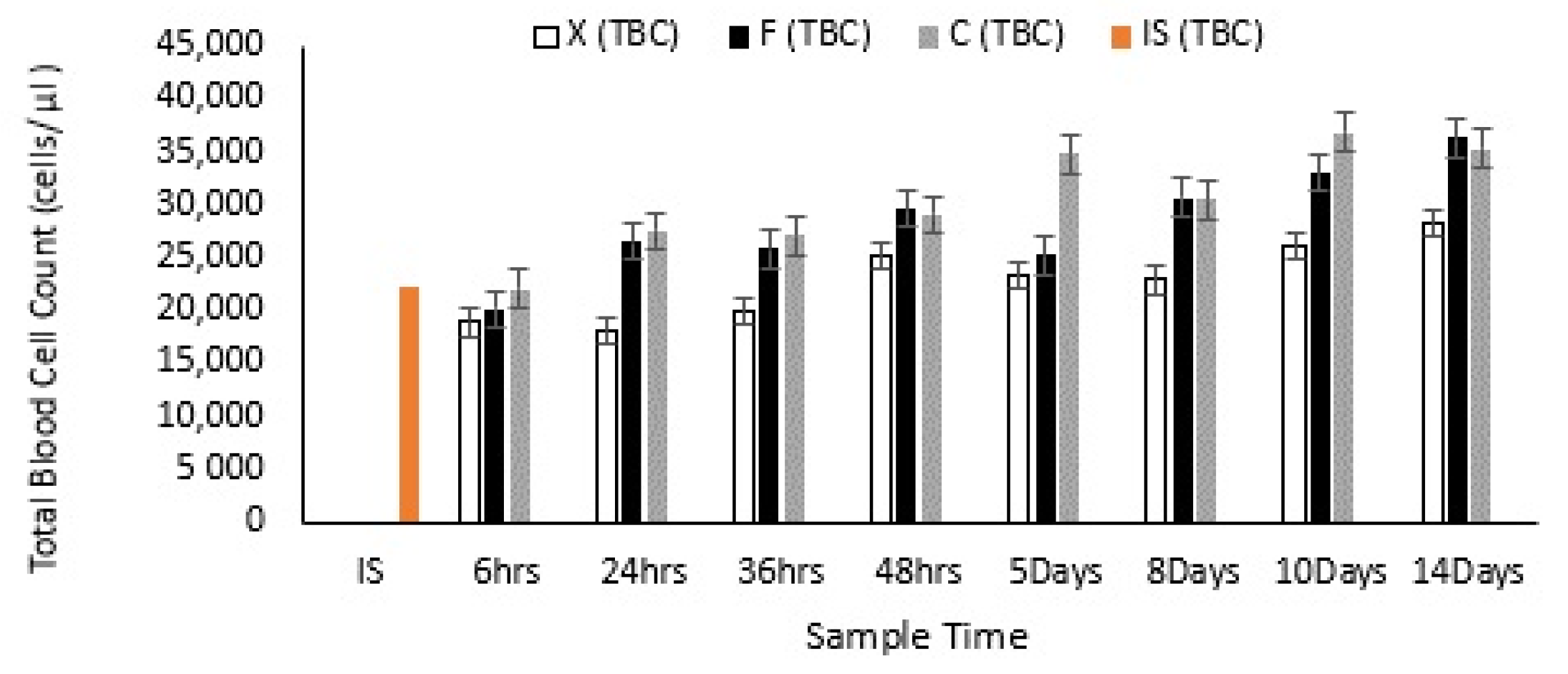
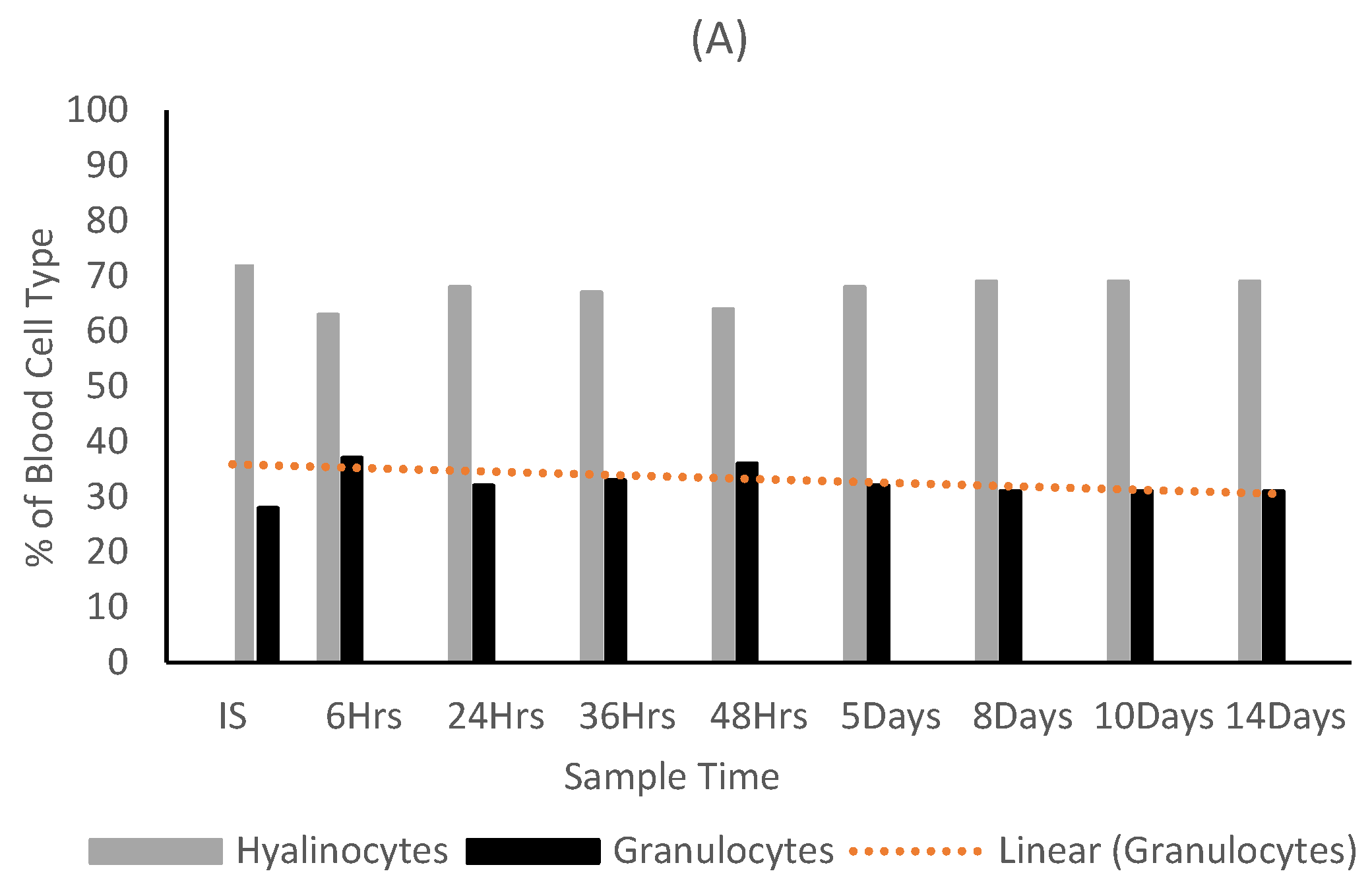
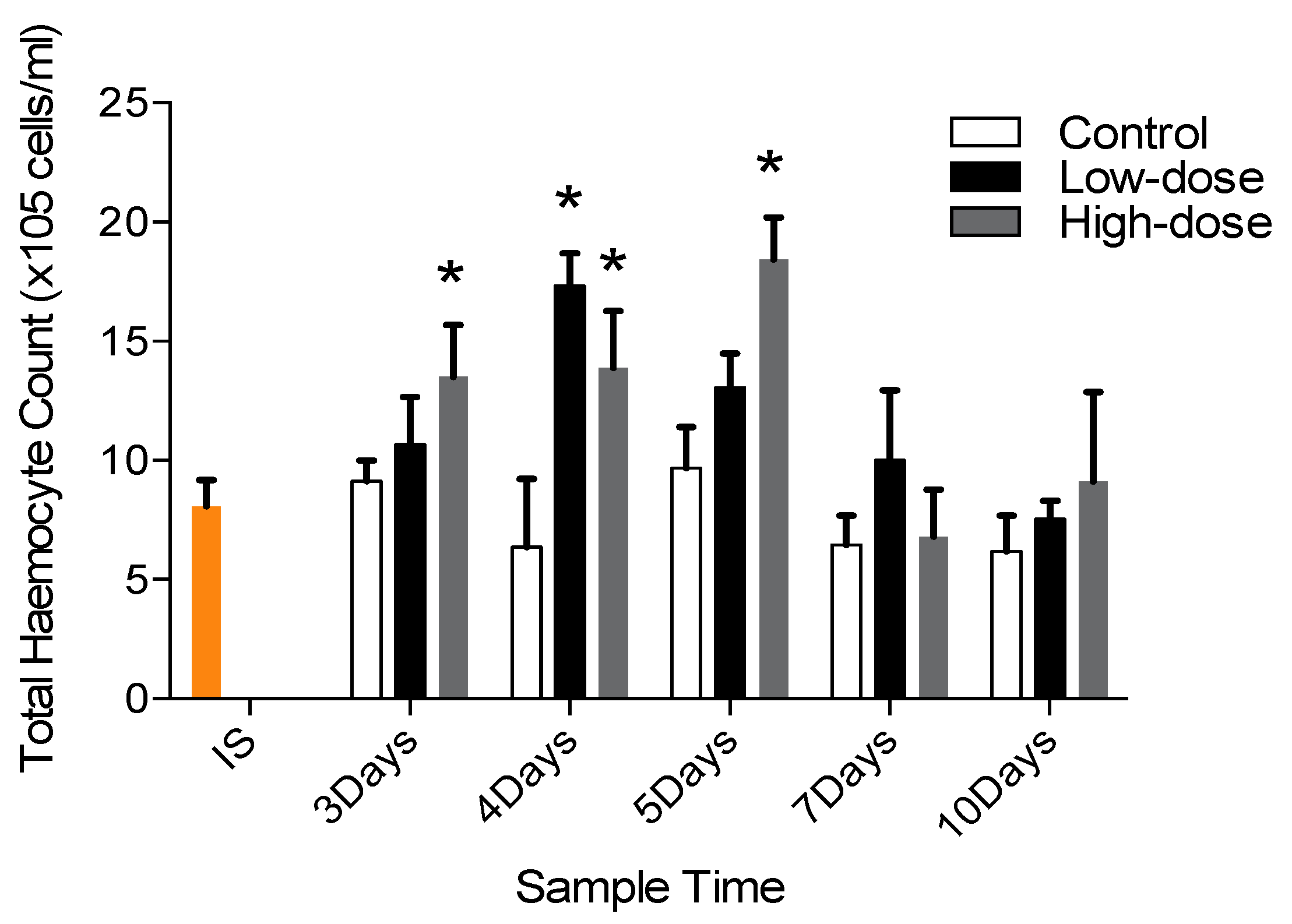
3.2. Effects of Fucoidan and Kappa (κ) Carrageenan on Crassostrea Gigas and Pathogens (OsHV-1 μVar and Vibrio spp (V. splendidus)
3.3. Effects of Sulphated Galactans on Palaemon Serratus
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Løvstad Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- De Almeida, C.L.F.; Falcão, H.D.S.; Lima, G.R.D.M.; Montenegro, C.D.A.; Lira, N.S.; De Athayde-Filho, P.F.; Rodrigues, L.C.; Souza, M.D.F.V.D.; Barbosa-Filho, J.M.; Batista, L.M. Bioactivities from Marine Algae of the Genus Gracilaria. Int. J. Mol. Sci. 2011, 12, 4550–4573. [Google Scholar] [CrossRef]
- Kesarcodi-Watson, A.; Miner, P.; Nicolas, J.-L.; Robert, R. Protective effect of four potential probiotics against pathogen-challenge of the larvae of three bivalves: Pacific oyster (Crassostrea gigas), flat oyster (Ostrea edulis) and scallop (Pecten maximus). Aquaculture 2012, 344–349, 29–34. [Google Scholar] [CrossRef]
- Kawashima, T.; Murakami, K.; Nishimura, I.; Nakano, T.; Obata, A. A sulfated polysaccharide, fucoidan, enhances the immunomodulatory effects of lactic acid bacteria. Int. J. Mol. Med. 2011, 29, 447–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isnansetyo, A.; Fikriyah, A.; Kasanah, N. Murwantoko Non-specific immune potentiating activity of fucoidan from a tropical brown algae (Phaeophyceae), Sargassum cristaefolium in tilapia (Oreochromis niloticus). Aquac. Int. 2016, 24, 465–477. [Google Scholar] [CrossRef]
- Craigie, J. Cell walls. In Biology of the Red Algae; Cole, K., Sheath, R., Eds.; Cambridge Univ. Press: Cambridge, UK, 1990; pp. 221–257. [Google Scholar]
- McCandless, E.L.; Craigie, J.S. Sulfated Polysaccharides in Red and Brown Algae. Annu. Rev. Plant Physiol. 1979, 30, 41–53. [Google Scholar] [CrossRef]
- Ehresmann, D.W.; Deig, E.F.; Hatch, M.T.; Disalvo, L.H.; Vedros, N.A. Antiviral substances from california marine algae1. J. Phycol. 1977, 13, 37–40. [Google Scholar] [CrossRef]
- Richards, J.T.; Kern, E.R.; Glasgow, L.A.; Overall, J.C.; Deign, E.F.; Hatch, M.T. Antiviral Activity of Extracts from Marine Algae. Antimicrob. Agents Chemother. 1978, 14, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Laille, M.; Gerald, F.; Debitus, C. In vitro antiviral activity on dengue virus of marine natural products. Cell. Mol. Life Sci. 1998, 54, 167–170. [Google Scholar] [CrossRef]
- Ghosh, P.; Adhikari, V.; Ghosal, P.K.; Pujol, C.A.; Carlucci, M.J.; Damonte, E.B.; Ray, B. In vitro anti-herpetic activity of sulfated polysaccharide fractions from Caulerpa racemosa. Phytochemistry 2004, 65, 3151–3157. [Google Scholar] [CrossRef]
- Hidari, K.I.; Takahashi, N.; Arihara, M.; Nagaoka, M.; Morita, K.; Suzuki, T. Structure and anti-dengue virus activity of sulfated polysaccharide from a marine alga. Biochem. Biophys. Res. Commun. 2008, 376, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Catarino, M.D.; Silva, A.M.S.; Cardoso, S.M. Phycochemical consitutents and biological activities of Fucus spp. Mar. Drugs. 2018, 16, 249. [Google Scholar] [CrossRef] [Green Version]
- Jönsson, M.; Allahgholi, L.; Sardari, R.R.R.; Hreggviðsson, G.O.; Karlsson, E.N. Extraction and modification of macroalgal polysaccharies fro current and next-generation applications. Molecules 2020, 25, 930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, L. Population Studies and Carrageenan Properties in Eight Gigartinales (Rhodophyta) from Western Coast of Portugal. Sci. World J. 2013, 2013, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Important Determinants for Fucoidan Bioactivity: A Critical Review of Structure-Function Relations and Extraction Methods for Fucose-Containing Sulfated Polysaccharides from Brown Seaweeds. Mar. Drugs 2011, 9, 2106–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, J.A.; Newell, R.I. Temporal and spatial variations in the composition of seston available to the suspension feeder Crassostrea virginica. Estuar. Coast. Shelf Sci. 1986, 23, 375–386. [Google Scholar] [CrossRef]
- Lynch, S.A.; Dillane, E.; Carlsson, J.; Culloty, S.C. Development and assessment of a sensitive and cost-effective polymerase chain reaction to detect ostreid herpesvirus 1 and variants. J. Shellfish. Res. 2013, 32, 1–8. [Google Scholar] [CrossRef]
- Schmitt, P.; Duperthuy, M.; Montagnani, C.; Bachère, E.; Destoumieux-Garzón, D. Immune Responses in the Pacific Oyster Crassostrea gigas an Overview with Focus on Summer Mortalities in Oyster’s Physiology, Ecological Distribution and Mortality; Qin, J., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2012. [Google Scholar]
- Kakehi, S.; Kamiyama, T.; Abe, H.; Hanawa, S.; Oota, H.; Matsuura, R.; Oshino, A. Mechanisms leading to the decline in Pacific oyster Crassostrea gigas seedlings in Matsushima Bay, Japan. Fish. Sci. 2016, 82, 499–508. [Google Scholar] [CrossRef]
- Whittington, R.J.; Paul-Pont, I.; Evans, O.; Hick, P.; Dhand, N.K. Counting the dead to determine the source and transmission of the marine herpesvirus OsHV-1 in Crassostrea gigas. Veter-Res. 2018, 49, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, H.J.; Williams, K.F.; Kaiser, M.J. A Review of the Palaemon Serratus Fishery: Biology, Ecology and Management, Fisheries and Conservation Report 2014 No. 38; Bangor University: Wales, UK, 2014. [Google Scholar]
- Galloway, T.S.; Depledge, M.H. Immunotoxicity in invertebrates: Measurement and ecotoxicological relevance. Ecotoxicology 2001, 10, 5–23. [Google Scholar] [CrossRef]
- Mydlarz, L.D.; Jones, L.E.; Harvell, C.D. Innate Immunity, Environmental Drivers, and Disease Ecology of Marine and Freshwater Invertebrates. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 251–288. [Google Scholar] [CrossRef] [Green Version]
- Allam, B.; Raftos, D. Immune responses to infectious diseases in bivalves. J. Invertebr. Pathol. 2015, 131, 121–136. [Google Scholar] [CrossRef] [Green Version]
- Medzhitov, R.; Janeway, C.A. Innate immunity: Impact on the adaptive immune response. Curr. Opin. Immunol. 1997, 9, 4–9. [Google Scholar] [CrossRef]
- Li, C.; Weng, S.; He, J. WSSV–host interaction: Host response and immune evasion. Fish Shellfish. Immunol. 2019, 84, 558–571. [Google Scholar] [CrossRef] [PubMed]
- Bachere, E.; Gueguen, Y.; Gonzalez, M.; De Lorgeril, J.; Garnier, J.; Romestand, B. Insights into the anti-microbial defense of marine invertebrates: The penaeid shrimps and the oyster Crassostrea gigas. Immunol. Rev. 2004, 198, 149–168. [Google Scholar] [CrossRef]
- Gestal, C.; Roch, P.; Renault, T.; Pallavicini, A.; Paillard, C.; Novoa, B.; Oubella, R.; Venier, P.; Figueras, A. Study of diseases and the immune system of bivalves using molecular biology and genomics. Rev. Fish. Sci. Aquac. 2008, 16, 131–154. [Google Scholar] [CrossRef] [Green Version]
- Gagnaire, B.; Frouin, H.; Moreau, K.; Thomas-Guyon, H.; Renault, T. Effects of temperature and salinity on haemocyte activities of the Pacific oyster, Crassostrea gigas (Thunberg). Fish Shellfish. Immunol. 2006, 20, 536–547. [Google Scholar] [CrossRef] [Green Version]
- Chu, F.-L.E.; La Peyre, J.F. Effect of environmental factors and parasitism on hemolymph lysozyme and protein of American oysters (Crassostrea virginica). J. Invertebr. Pathol. 1989, 54, 224–232. [Google Scholar] [CrossRef]
- Cronin, M.; Culloty, S.; Mulcahy, M. Lysozyme activity and protein concentration in the haemolymph of the flat oyster Ostrea edulis (L.). Fish Shellfish. Immunol. 2001, 11, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Prado-Alvarez, M.; Darmody, G.; Hutton, S.; O’Reilly, A.; Lynch, S.A.; Culloty, S.C. Occurrence of OsHV-1 in Crassostrea gigas Cultured in Ireland during an Exceptionally Warm Summer. Selection of Less Susceptible Oysters. Front. Physiol. 2016, 7, 492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholz, U.; Diaz, G.G.; Ricque, D.; Suarez, L.C.; Albores, F.V.; Latchford, J. Enhancement of vibriosis resistance in juvenile Penaeus vannamei by supplementation of diets with different yeast products. Aquaculture 1999, 176, 271–283. [Google Scholar] [CrossRef]
- Ganz, T. The Role of Antimicrobial Peptides in Innate Immunity. Integr. Comp. Biol. 2003, 43, 300–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Hao, R.; Deng, Y.; Liao, Y.; Wang, Q.; Sun, R.; Jiao, Y.; Du, X. Effects of protein sources on growth, immunity and antioxidant capacity of juvenile pearl oyster Pinctada fucata martensii. Fish Shellfish. Immunol. 2017, 67, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Carboni, S.; Clegg, S.H.; Hughes, A.D. The use of biorefinery by-products and natural detritus as feed sources for oysters (Crassostrea gigas) juveniles. Aquaculture 2016, 464, 392–398. [Google Scholar] [CrossRef]
- Tanyaros, S.; Chuseingjaw, S. A partial substitution of microalgae with single cell detritus produced from seaweed (Porphyra haitanensis) for the nursery culture of tropical oyster (Crassostrea belcheri). Aquac. Res. 2014, 47, 2080–2088. [Google Scholar] [CrossRef]
- Rudtanatip, T.; Lynch, S.A.; Wongprasert, K.; Culloty, S.C. Assessment of the effects of sulfated polysaccharides extracted from the red seaweed Irish moss Chondrus crispus on the immune-stimulant activity in mussels Mytilus spp. Fish Shellfish. Immunol. 2018, 75, 284–290. [Google Scholar] [CrossRef]
- Wongprasert, K.; Rudtanatip, T.; Praiboon, J. Immunostimulatory activity of sulfated galactans isolated from the red seaweed Gracilaria fisheri and development of resistance against white spot syndrome virus (WSSV) in shrimp. Fish Shellfish. Immunol. 2014, 36, 52–60. [Google Scholar] [CrossRef]
- Sinurat, E.; Saepudin, E.; Peranginangin, R.; Hudiyono, S. Immunostimulatory activity of brown seaweed-derived fucoidans at different molecular weights and purity levels towards white spot syndrome virus (WSSV) in shrimp Litopenaeus vannamei. J. Appl. Pharm. Sci. 2016, 6, 082–091. [Google Scholar] [CrossRef] [Green Version]
- Cantelli, L.; Goncalves, P.; Guertler, C.; Kayser, M.; Pilotto, M.R.; Barracco, M.A.; Perazzolo, L.M. Dietary supplementation with sulfated polysaccharides from Gracilaria birdiae promotes a delayed immunostimulation in marine shrimp challenged by the white spot syndrome virus. Aquac. Int. 2018, 27, 349–367. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Sim, S.S.; Chiew, S.L.; Yeh, S.-T.; Liou, C.-H.; Chen, J.-C. Dietary administration of a Gracilaria tenuistipitata extract produces protective immunity of white shrimp Litopenaeus vannamei in response to ammonia stress. Aquaculture 2012, 370–371, 26–31. [Google Scholar] [CrossRef]
- Cunha, L.; Grenha, A. Sulfated Seaweed Polysaccharides as Multifunctional Materials in Drug Delivery Applications. Mar. Drugs 2016, 14, 42. [Google Scholar] [CrossRef]
- Manilal, A.; Selvin, J.; Sugathan, S. Immuno-Modulatory Efficacy of Indian Red Algae, Asparagopsis taxiformis, in Penaeus monodon. J. Appl. Aquac. 2013, 25, 81–93. [Google Scholar] [CrossRef]
- Jasmanindar, Y.; Sukenda, S.; Zairin, M., Jr.; Alimuddin, A.; Utomo, N.B. Dietary administration of Gracilaria verrucosa extract on Litopenaeus vannamei immune response, growth, and resistance to Vibrio harveyi. Aquacult. Aquar. Conserv. Legis. 2018, 11, 1069–1080. [Google Scholar]
- Manilal, A.; Selvin, J.; George, S. In vivo therapeutic potentiality of red seaweed, Asparagopsis (Bonnemaisoniales, Rhodophyta) in the treatment of Vibriosis in Penaeus monodon Fabricius. Saudi J. Biol. Sci. 2012, 19, 165–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steentoft, M.; Farnharn, W.F. Northern distribution boundaries and thermal requirements of Gracilaria and Gracilariopsis (Gracilariales, Rhodophyta) in Atlantic Europe and Scandinavia. Nord. J. Bot. 1997, 17, 87–94. [Google Scholar] [CrossRef]
- Hardy, F.G.; Guiry, M.D. A Check-List and Atlas of the Seaweeds of Britain and Ireland; British Phycological Society: London, UK, 2003; ISBN 0-9527115-1-6. [Google Scholar]
- Ruperez, P. Mineral content of edible marine seaweeds. Food Chem. 2002, 79, 23–26. [Google Scholar] [CrossRef]
- Gómez-Ordóñez, E.; Jiménez-Escrig, A.; Rupérez, P. Bioactivity of sulfated polysaccharides from the edible red seaweed Mastocarpus stellatus. Bioact. Carbohydr. Diet. Fibre 2014, 3, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Walsh, P.S.; Metzger, D.A.; Higuchi, R. Chelex 100 as a Medium for Simple Extraction of DNA for PCR-Based Typing from Forensic Material. Biotechniques 2013, 54, 134–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, S.C.; Fidler, A.; Renault, T. Primers for PCR-based detection of ostreid herpes virus-1 (OsHV-1): Application in a survey of New Zealand molluscs. Aquaculture 2007, 272, 126–139. [Google Scholar] [CrossRef]
- Carballal, M.J.; Lopez, C.; Azevedo, C.; Vilalba, A. Enzymes involved in defence function of hemocytes of mussel Mytilus galloprovincialis. J. Invertebr. Pathol. 1997, 70, 96–105. [Google Scholar] [CrossRef]
- McCleary, S.; Henshilwood, K. Novel quantitative TaqMan® MGB real-time PCR for sensitive detection of Vibrio aestuarianus in Crassostrea gigas. Dis. Aquat. Org. 2015, 114, 239–248. [Google Scholar] [CrossRef]
- Kett, G.F.; Culloty, S.C.; Jansen, M.A.K.; Lynch, S.A. Development of a Sensitive Polymerase Chain Reaction (PCR) and DIG-Labelled in situ Hybridisation (ISH) for the Detection of Vibrio Bacteria; University College Cork: Cork, Ireland, 2021; Unpublished. [Google Scholar]
- Notaro, D.A.; Culloty, S.C.; Lynch, S.A. A pilot study investigating the potential of antimicrobial photodynamic therapy (aPDT) to control Vibrio spp. development in microalgae and seawater. Aquac. Int. 2021, 29, 355–372. [Google Scholar] [CrossRef]
- Walberg, J. White blood cell counting techniques in birds. Semin. Avian Exot. Pet Med. 2001, 10, 72–76. [Google Scholar] [CrossRef]
- Carisch, L.; Stirn, M.; Hatt, J.M.; Federer, K.; Hofmann-Lehmann, R.; Riond, B. White blood cell count in birds: Evaluation of a commercially available method. BMC Veter-Res. 2019, 15, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Wang, L.; Zhou, Z.; Sun, Y.; Wang, M.; Wang, H.; Hou, Z.; Gao, D.; Gao, Q.; Song, L. The simple neuroendocrine-immune regulatory network in oyster Crassostrea gigas mediates complex functions. Sci. Rep. 2016, 6, 26396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerdol, M.; Gomez-Chiarri, M.; Castillo, M.G.; Figueras, A.; Fiorito, G.; Moreira, R.; Novoa, B.; Pallavicini, A.; Ponte, G.; Roumbedakis, K.; et al. Immunity in Molluscs: Recognition and Effector Mechanisms, with a Focus on Bivalvia. In Advances in Comparative Immunology; Metzler, J.B., Ed.; Springer: New York City, NY, USA, 2018; pp. 225–341. [Google Scholar]
- Usov, A.I. Polyscaccharides of the red algae. Adv. Carb. Chem. Biochem. 2011, 65, 115–217. [Google Scholar]
- Sritunyalucksana, K.; Gangnonngiw, W.; Archakunakorn, S.; Fegan, D.; Flegel, T.W. Bacterial clearance rate and a new differential hemocyte staining method to assess immunostimulant activity in shrimp. Dis. Aquat. Org. 2005, 63, 89–94. [Google Scholar] [CrossRef]
- Rudtanatip, T.; Withyachumnarnkul, B.; Wongprasert, K. Sulfated galactans from Gracilaria fisheri bind to shrimp haemocyte membrane proteins and stimulate the expression of immune genes. Fish Shellfish. Immunol. 2015, 47, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Wargacki, A.J.; Leonard, E.; Win, M.N.; Regitsky, D.D.; Santos, C.N.S.; Kim, P.B.; Cooper, S.R.; Raisner, R.M.; Herman, A.; Sivitz, A.B.; et al. An Engineered Microbial Platform for Direct Biofuel Production from Brown Macroalgae. Science 2012, 335, 308–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitikiew, S.; Chen, J.-C.; Putra, D.F.; Lin, Y.-C.; Yeh, S.-T.; Liou, C.-H. Fucoidan effectively provokes the innate immunity of white shrimp Litopenaeus vannamei and its resistance against experimental Vibrio alginolyticus infection. Fish Shellfish. Immunol. 2013, 34, 280–290. [Google Scholar] [CrossRef]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lynch, S.A.; Breslin, R.; Bookelaar, B.; Rudtanatip, T.; Wongprasert, K.; Culloty, S.C. Immunomodulatory and Antiviral Effects of Macroalgae Sulphated Polysaccharides: Case Studies Extend Knowledge on Their Importance in Enhancing Shellfish Health, and the Control of a Global Viral Pathogen Ostreid Herpesvirus-1 microVar. Polysaccharides 2021, 2, 202-217. https://0-doi-org.brum.beds.ac.uk/10.3390/polysaccharides2020014
Lynch SA, Breslin R, Bookelaar B, Rudtanatip T, Wongprasert K, Culloty SC. Immunomodulatory and Antiviral Effects of Macroalgae Sulphated Polysaccharides: Case Studies Extend Knowledge on Their Importance in Enhancing Shellfish Health, and the Control of a Global Viral Pathogen Ostreid Herpesvirus-1 microVar. Polysaccharides. 2021; 2(2):202-217. https://0-doi-org.brum.beds.ac.uk/10.3390/polysaccharides2020014
Chicago/Turabian StyleLynch, Sharon A., Rachel Breslin, Babette Bookelaar, Tawut Rudtanatip, Kanokpan Wongprasert, and Sarah C. Culloty. 2021. "Immunomodulatory and Antiviral Effects of Macroalgae Sulphated Polysaccharides: Case Studies Extend Knowledge on Their Importance in Enhancing Shellfish Health, and the Control of a Global Viral Pathogen Ostreid Herpesvirus-1 microVar" Polysaccharides 2, no. 2: 202-217. https://0-doi-org.brum.beds.ac.uk/10.3390/polysaccharides2020014