
Melanocortins and Their Potential for the Treatment, Prevention and Amelioration of Complications of Diabetes

Department of Diabetes, Central Clinical School, Monash University, Melbourne, VIC 3004, Australia
*
Author to whom correspondence should be addressed.
Diabetology 2024, 5(1), 69-84; https://0-doi-org.brum.beds.ac.uk/10.3390/diabetology5010006
Submission received: 7 December 2023
/
Revised: 9 January 2024
/
Accepted: 29 January 2024
/
Published: 4 February 2024
(This article belongs to the Special Issue Exclusive Papers Collection of Editorial Board Members in Diabetology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Diabetes mellitus (type I and II) is an advancing global health problem, concerningly increasing in prevalence in most of the developed and developing world. Current therapies, such as the subcutaneous injection of insulin, are invasive and require a close monitoring of blood glucose levels to prevent hypo- or hyperglycaemia from occurring. Despite an inexorable search for a cure since Banting and Best discovered and purified insulin in 1921, insulin remains a solitary insula, still the gold standard for treatment of type I and late-stage type II diabetes mellitus. Apropos of complications, diabetes causes a myriad of secondary maladies, ranging from diabetic kidney disease, diabetic retinopathy and diabetic neuropathy to erectile dysfunction and peripheral vascular disease. While scientists continue to interminably tinker with perfecting mechanical insulin pumps or dampening the immune response to pancreatic beta cells, an important aspect of the aetiology of diabetes should not be neglected, that of the metabolism. At its heart, diabetes can arguably be considered a metabolic disease, and this review suggests a return to focusing on preventing and treating diabetes by focussing on its metabolic causes. This narrative review summarises the potential of a recent class of synthetic peptides, the melanocortins, to help prevent and treat the complications of diabetes mellitus. The review summarises recent work showing the potential benefits of the melanocortins in treating diabetic complications through various pathways.
Keywords:
diabetes mellitus; melanocortins; obesity; metabolism; atherosclerosis; retinopathy; nephropathy; neuropathy; POMC; α-MSH1. Introduction
Approximately 537 million people suffer from diabetes (type I and II) around the world, and approximately 6.7 million people died from diabetes and its complications in 2021 [1]. It is therefore pellucid that diabetes is one of the major health concerns of the 21st century. Although the major two types of diabetes have different aetiologies, they both have similar late-stage complications. These devastating complications include diabetic kidney disease, diabetic retinopathy and cardiovascular disease.
This review brings attention to a class of recently developed synthetic peptide analogues, the melanocortins, currently used in rare sunlight disorders, sexual dysfunction treatment and obesity, that have so far been overlooked in terms of their potential to prevent type II diabetes and treat the complications of both type I and type II diabetes. The majority of this review focusses on the ability of melanocortins to influence the metabolism, neurologically and peripherally, to ameliorate the obesity and insulin resistance often seen in those prone to developing type II diabetes. Indeed, insulin resistance is often an indication of an impending diagnosis of type II diabetes. Its inimical effects lead to a vicious cycle of increased hunger and subsequent weight gain which, in turn, worsens the insulin resistance and exhausts the ability of pancreatic beta cells to produce insulin. This is especially relevant considering 541 million people around the world live with impaired glucose tolerance [1]. Melanocortins have the ability to break this cycle and lead to the prevention or amelioration of type II diabetes. The rest of the review focusses on the ability of melanocortins to act as anti-inflammatories to treat the complications of both type I and type II diabetes.
2. Melanocortins and the POMC System
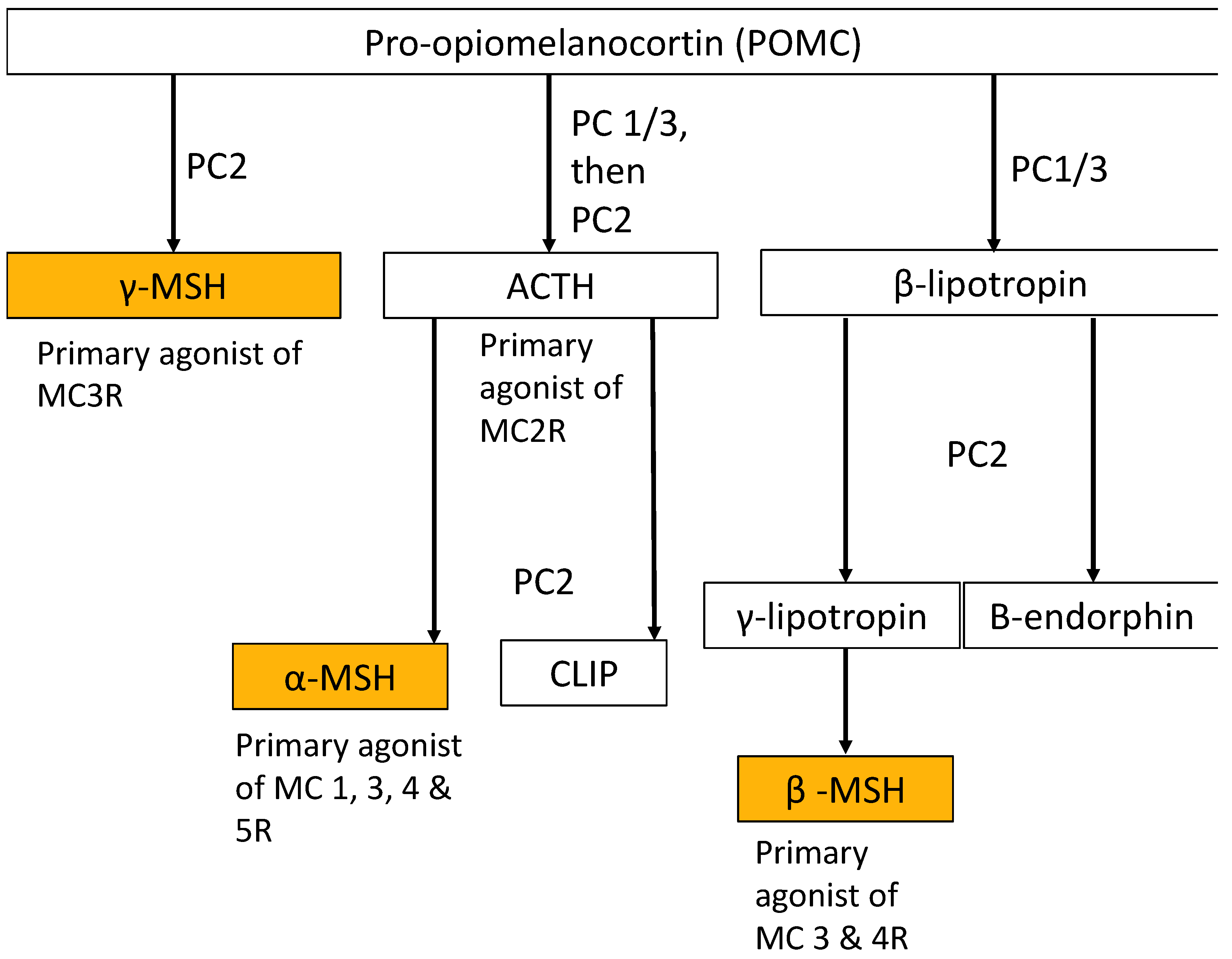
Melanocortins, a group of cleavage peptide products of pro-opiomelanocortin (POMC), activate melanocortin receptors on the surface of a diverse range of cell types, leading to different biological actions. They are so named because of their melanotropic activity, that is, the ability of melanocortins to increase pigmentation in melanocytes in the skin and hair follicles, increase concentrations of eumelanin and prevent an increase in photosensitive pheomelanin [2]. Melanocortins are produced by POMC neurons in the pars intermedia of the pituitary gland, the hypothalamic arcuate nucleus and the dorsal medullary nucleus of the solitary tract. They can be distinguished by the presence of an invariant amino acid sequence in each melanocortin peptide, His-Phe-Arg-Trp [3]. The melanocortins produced in humans include alpha-melanocyte stimulating hormone (α-MSH), beta-melanocyte stimulating hormone (β-MSH), gamma-melanocyte stimulating hormone (γ-MSH) and adrenocorticotropic hormone (ACTH).
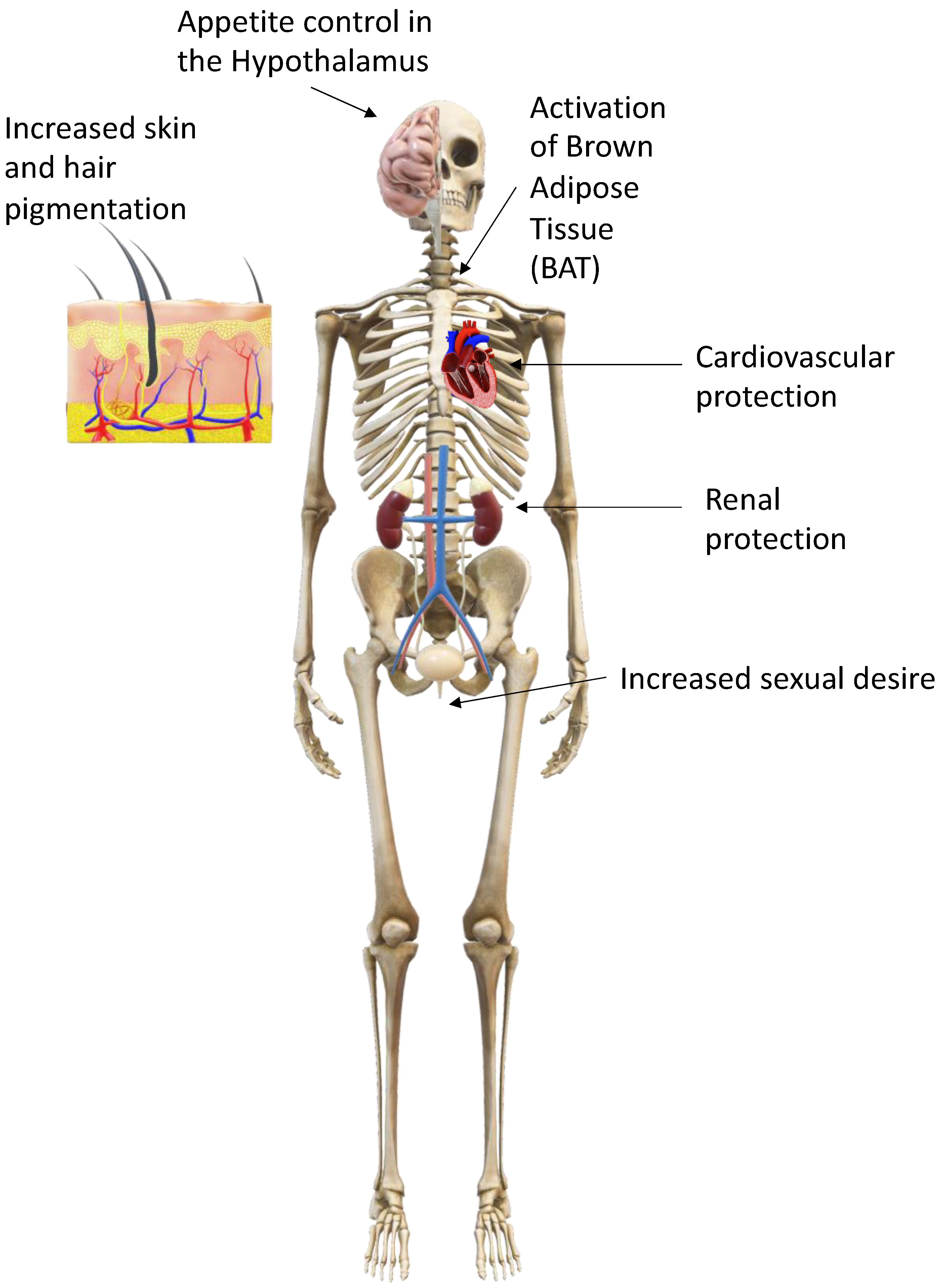
The POMC system is a neuroendocrine system involved in the regulation of appetite, energy homeostasis, sexual behaviour, DNA repair, cognitive function, memory, neuronal regeneration, pain, blood pressure, skin and hair pigmentation, inflammation and steroidogenesis [4]. POMC, a 28.9 kDa precursor protein produced in the brain, anterior pituitary gland, skin and other distal locations, is proteolytically cleaved into various peptide effector molecules (termed melanocortins, corticotrophins and opioids; see Figure 1) by prohormone convertases (PCs), which activate receptors known as the melanocortin, corticotrophin and opioid receptors in order to affect a diverse range of physiological functions both centrally and in the periphery (see Figure 2) [5].
The POMC gene, located on chromosome 2p23, contains three exons which produce a protein consisting of three main segments: N-POMC at the amino terminus, adrenocorticotropic hormone (ACTH) in the middle and beta-lipotropin hormone (β-LPH) at the carboxyl terminus. β-LPH can be further cleaved into gamma-lipotropin (γ-LPH), beta- melanocyte stimulating hormone (β-MSH) and beta-endorphin (β-END). Each segment produces one form of MSH. Mutations in POMC have been shown to cause severe, early-onset obesity, and all of the major melanocortins (ACTH, α-MSH, β-MSH and γ-MSH) have been shown to ameliorate obesity in animal models [6,7,8,9,10,11,12,13,14]. Melanocortins activate a group of five G-coupled protein receptors (termed melanocortin receptors, MCR) on the cell surface leading to a signal transduction cascade in the cell.
3. Melanocortins Protect against Diabetes Caused by Obesity and Insulin Resistance
3.1. Melanocortins and the Control of Neural Energy Homeostasis
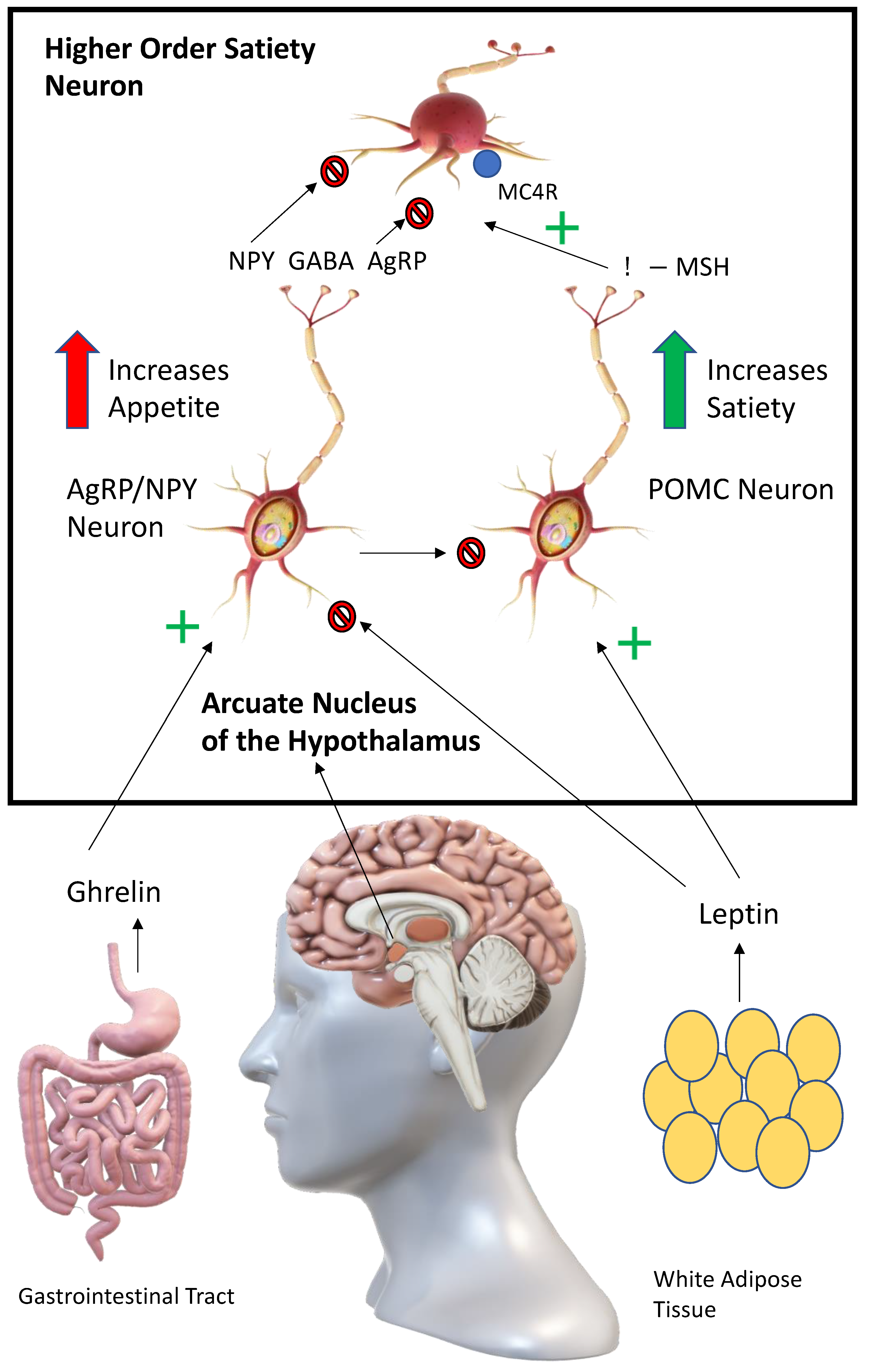
Perhaps the most obvious link between melanocortins and metabolism is their effect on energy homeostasis via hunger reward pathways in the brain (see Figure 3). Appetite and food intake is controlled in the arcuate nucleus of the hypothalamus by the competing actions of pro-opiomelanocortin (POMC) and agouti-related peptide/neuropeptide Y (AgRP/NPY) neurons [15]. Activation of POMC neurons leads to anorexigenic effects, while activation of AgRP/NPY neurons leads to orexigenic effects. The way this system functions after a meal is as follows: leptin, which is produced by adipocytes and enterocytes, activates POMC neurons in the arcuate nucleus [16]. This leads to the release of melanocortins (predominantly α-MSH) from POMC neurons which activates melanocortin receptors (predominantly MC4R) on higher order satiety neurons. Satiety and the desire to perform physical activity are increased. In post-prandial periods, the AgRP/NPY neurons act as antipodes of POMC neurons, leading to unpleasant hunger pains. Ghrelin, a hormone produced by the enteroendocrine cells, activates AgRP/NPY neurons in the arcuate nucleus [17]. These neurons, in turn, release AgRP, NPY and GABA (gamma-aminobutyric acid) to antagonise, directly and indirectly, higher-order satiety neurons and their receptors (e.g., MCR4), leading to hyperphagia. AgRP/NPY neurons also directly inhibit the release of melanocortins from POMC neurons during post-prandial periods. Agouti-related protein/peptide (AgRP) is a neuropeptide synthesised by the cell bodies of AgRP/NPY neurons in the ventromedial arcuate nucleus in the brain. Agouti protein is a paracrine competitive antagonist of MC1R and MC4R. The effect of this blockage is hyperphagia, obesity and eventual type II diabetes [18]. Therefore, providing exogeneous melanocortins is likely to correct hyperphagia and restore satiety.
3.2. Melanocortins and Leptin: A Potential Downstream Advantage
Since the first ob/ob mouse was discovered by chance in 1949 at the Jackson Laboratory, scientists have been intrigued by the possible use of leptin as an obesity therapy to prevent type II diabetes [19]. Ob/ob have a recessive gene mutation that prevents them from producing leptin [20]. This leads to uncontrollable eating and high blood glucose levels, creating phenotypically corpulent mice. While leptin has been found to have some positive effects in insulin-deficient type I diabetes, it has not seen the same success in treatment for obesity, metabolic syndrome and type II diabetes [21]. This is due to a key difference between ob/ob mice and human type II diabetics. Leptin mutations in humans are vanishingly rare and therefore, most type II diabetics have adequate leptin levels. Unfortunately, in type II diabetics, there seems to be a so-called leptin insensitivity, which is comparable to insulin resistance seen in type II diabetics and patients with metabolic syndrome. As explained in the previous section, melanocortins are downstream from leptin signalling and there is no known resistance to the effects of the melanocortins. Administering melanocortins potentially bypasses the leptin pathway by cozening the appetite centres in the brain into thinking a large calorie-dense meal has just been consumed.
A level of extended satiety can thus be reached, allowing for an amelioration of obesity and its consequent conditions, such as type II diabetes and its associated insulin resistance. The administration of various synthetic melanocortin peptides has been successfully used in this manner in multiple animal and human studies [6,7,8,9,11,12,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41].
3.3. An Additional Benefit: The Effects of Melanocortins in the Periphery
The metabolic benefits of melanocortins do not cease at the neuroendocrine level. Melanocortins also directly act on adipocytes via MC4R in the periphery to increase lipolysis and activation of brown adipose tissue (BAT).
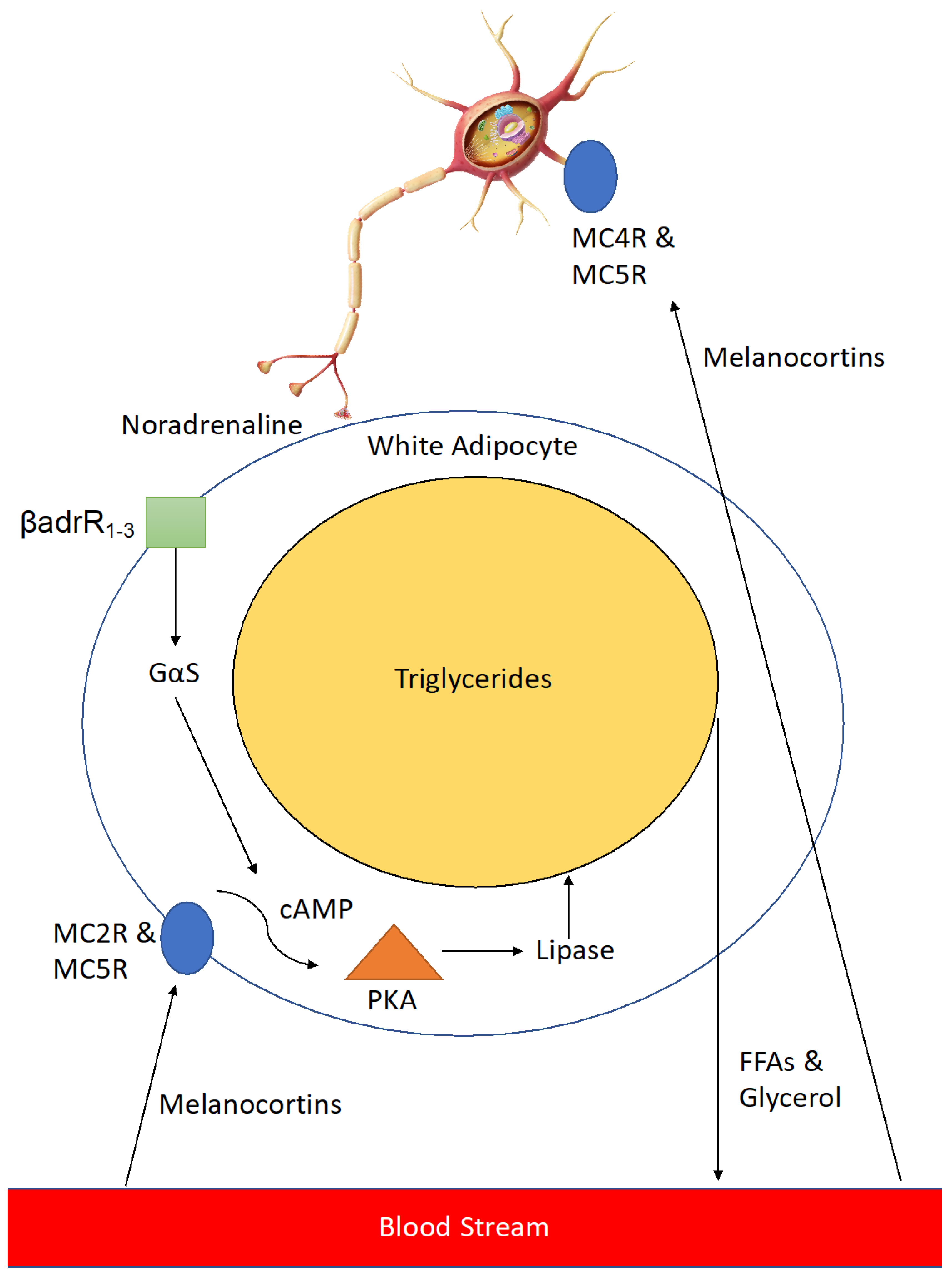
Melanocortins activate neurons innervating adipose tissue via MC4R and MC5R to release noradrenaline which activates β 1–3 adrenergic receptors on the cell surface, leading to a downstream signalling cascade which results in expression of lipase enzyme that breaks down triglycerides in the large lipid droplets found in adipocytes into free-fatty acids and glycerol (Figure 4). This leads to reduced lipid volume in the adipocytes and thus a reduction in their size, resulting in a loss of adipose tissue mass. In addition, melanocortins help to counteract adipogenesis caused by enhanced levels of Agouti-signalling protein (ASIP) seen in type II diabetics [42].
BAT, differentiated from white adipose tissue (WAT), is a metabolically active form of adipocyte found in small quantities in human adults. It is primarily found around cervical, supraclavicular, axillary periaortic, para-vertebral and suprarenal areas. BAT undergoes a process of non-shivering thermogenesis via activation of the mitochondrial protein, uncoupling protein 1 (UCP-1), which acts to decrease the proton gradient across the mitochondrial inner membrane formed by oxidative phosphorylation (see Figure 5). This scuppers the ability of the mitochondria to produce adenosine triphosphate (ATP) and instead produces copious amounts of heat. BAT is normally activated following a large meal or exposure to cold temperatures. In the early part of the twentieth century an explosive used in munitions factories during World War I, 2,4-dinitrophenol, was repurposed as a weight loss drug due to its mitochondrial uncoupling effects. Unfortunately, whilst being highly efficacious in reducing obesity and insulin resistance, uncoupling drugs tread a very thin line between a therapeutic and a fatal dose [43]. They have been banned by the FDA for use in humans since 1938 [44]. Consequently, a direct upregulation or activation of UCP-1 or other uncoupling proteins is considered unwise. Indeed, as this review has expounded on, homeostatic mechanisms control the majority of metabolic processes. Therefore, it is crucial for an effective therapy to work within these confines and to only supplement where a rate-limiting step exists. Melanocortins function within these bounds, failing to cause the dangerous cardiovascular side effects of uncoupling drugs while providing a similar metabolic benefit.
3.4. Setmelanotide: Emerging Evidence for the Use of a Clinical Melanocortin Analogue in Metabolic Disease
The practicality of using melanocortins as therapeutics was initially uncertain as bioidentical melanocortin peptides have vanishingly short half-lives and are promiscuous agonists of all melanocortin receptors [45]. Formulation work commenced and biochemists utilised non-canonical amino acid substitutions to prolong half-life and to make the synthetic peptides more specific for certain melanocortin receptors, although some would argue more work needs to be conducted on increasing specificity as every synthetic melanocortin to date is still somewhat promiscuous [45]. Setmelanotide is an example of a designed synthetic selective peptide agonist of MC4R currently being developed by Rhythm Pharmaceuticals as a subcutaneous injectable anti-obesity treatment for Prader–Willi syndrome (PWS) (a genetic cause of MC4R deficiency), leptin receptor deficiency (LEPR), POMC deficiency and lifestyle-induced obesity. Setmelanotide has completed phase III trials for POMC deficiency and LEPR deficiency and was approved by the FDA as first-in-class medication in November 2020. It has also been provisionally approved this year in France for lesional hypothalamic obesity and has been approved in Canada for Bardet-Biedl syndrome. The trials showed significant weight loss (an average of 10%) after one year of setmelanotide therapy [10,11,46]. The therapy was shown to achieve this 10% average weight loss in 80% of patients with POMC deficiency and 45.5% of patients with a LEPR deficiency [10,46,47]. Furthermore, it was shown that setmelanotide induced at least a 25% reduction in hunger in 72.7% of patients with LEPR deficiency and in 50% of patients with POMC deficiency [10,46]. The average weight loss of patients with POMC and LEPR deficiency was 31.9 kg and 16.7 kg, respectively [10,46]. BMI measurements were also significantly lowered by an average of between 22.3 and 49.2% in patients with POMC deficiency and by an average of 10.6% in patients with LEPR deficiency [10,46]. Follow up unpublished studies on these patients reported at a recent French paediatrics conference showed sustained weight loss over a period of 3 years. Additional successful phase III trials in other forms of genetic obesity such as Bardet–Biedl and Alström syndromes are currently under priority review with the FDA [47,48,49]. The FDA approved setmelanotide for Bardet–Biedl syndrome in June 2022. A range of phase II and III trials (titled DAYBREAK and EMANATE) in different rare genetic forms of obesity including MC4R deficiency are currently underway [7]. The mechanism of action of setmelanotide is via suppression of appetite, an increase in satiety and an increase in thermogenesis, leading to a reduction in excess feeding behaviours and a higher resting energy expenditure rate [7,11,46,47,48,49]. Setmelanotide exerts its effects by binding to MC4R in the PVN of the hypothalamus and the lateral hypothalamic area (LHA). Some have raised concerns of negative neuropsychiatric effects of setmelanotide such as an increased suicidal ideation seen in some patients in follow up studies of patients from the initial phase III study [49]. However, it should be noted that many patients with obesity report low self-esteem, poor body image, depression and anxiety, with depression being a major risk factor for developing obesity [48,50]. Therefore, it is unclear whether pre-existing depression is causing the suicidal ideation or if it is worsened by the drug. The manufacturer denies that these cases were a result of treatment, and further follow-up studies report improved quality of life (QOL) assessments in the majority of patients treated [48,50]. However, further studies should focus on this potential concerning side effect.
Similar to how GLP-1 analogues are currently used to treat both type II diabetes and obesity (although this review does not intend to make a direct comparison between the two with regard to either efficacy or side effect profile), melanocortins with similar actions to setmelanotide have potential for treating diabetic patients in the future and for preventing many more patients suffering from obesity, PCOS and metabolic syndrome from developing type II diabetes. Although setmelanotide and other synthetic melanocortins have been developed for genetic forms of obesity, melanocortins (melanotan I and II and bremelanotide) also work to decrease body fat in both healthy non-obese and diet-induced obese individuals [51,52,53]. A truncated form of MSH caused an average approximate body fat loss of 2 kg over a 6-week period using twice daily nasal administration in healthy, non-obese subjects [51]. These are the only human trials to date using melanocortins in non-genetic forms of obesity. More long-term human studies are needed to determine the safety and utility of melanocortins for treating diet-induced obesity. Various diet-induced obese animal models have shown substantial weight loss after administration of various bioidentical and synthetic melanocortin peptides [27,30,54,55]. This melanocortin-induced weight loss has also been seen even in leptin resistant animals [56,57]. With the paradigm-shifting success of GLP-1 agonists, the question may be asked as to what further benefit could melanocortins provide in the obesity treatment milieu. Although they provide a sustained weight loss (up to 15% average of total body weight in long-term large-scale trials), according to studies, 70% of patients taking GLP-1 agonists discontinue use after 24 months (it is still a matter of speculation as to why this is so, but some have postulated that severe nausea and other gastrointestinal side effects could be major contributors, as well as cost and ease-of-access issues) [58,59,60]. Melanocortins can cause skin darkening, flushing, increased libido, spontaneous erections and mild nausea but are generally well-tolerated, some of these side effects may be seen as additional benefits (although they may not be desirable for all people), hence the colloquial moniker of the melanocortin class as being “Barbie drugs” as they cause skin tanning, weight loss and sexual arousal [61,62]. Spurred on by weight loss results seen in healthy obese women using the FDA-approved female sexual dysfunction melanocortin drug bremelanotide (an average loss of 1.3 kg bodyweight over 16 days and a reduction of an average of 400 kcal in food consumption per day in the first phase I trial and an average loss of 1.7 kg bodyweight over 12 days in the second phase I trial; side effects in the trial were classed as mostly mild, whilst a smaller subset of subjects had side effects classed as moderate; skin darkening was reported as a side effect in 15–63.3% of the bremelanotide-treated group across both studies; and five patients had to withdraw due to nausea, dizziness and hypertension), Palatin Pharmaceuticals is developing a combination treatment of bremelanotide and a low-dose GLP-1 agonist to treat obesity [52]. They hope to have improved tolerability and patient compliance with similar or enhanced weight loss compared to GLP-1 agonist monotherapy. Therefore, rather than replacing the already efficacious GLP-1 agonists, in the future, melanocortin/GLP-1 agonist dual therapy may be a way of maximising the potential benefits of both drugs whilst allowing far lower dosages and thus decreasing the chances of side effects and increasing patient adherence.
4. Melanocortins and Their Potential in Treating Diabetic Complications
4.1. Atherosclerosis and the Melanocortins
Atherosclerosis, a chronic inflammatory disease affecting the aorta, is a complex process whereby inflammation and oxidative stress result in endothelial dysfunction, lipid accumulation in the vessel walls, monocyte infiltration and foam cell formation, all of which leads to the formation of a lesion or plaque. If these plaques become unstable, they can rupture. Rupture of an atherosclerotic plaque can lead to myocardial infarction or stroke, often resulting in sudden death. Diabetes accelerates atherosclerotic plaque formation.
So what evidence is available for the role of melanocortin receptors in atherosclerosis? Previous studies have shown that the MC4R is involved in cardiovascular disease, reviewed in Copperi et al. [63]. A recent study on Korean subjects showed that the MC4R single-nucleotide polymorphism (SNP) rs17782313 was found to be associated with a higher risk of diabetes and cardiovascular disease in lean men [64]. Although the C allele of this gene in humans increases BMI, it did not increase cardiovascular disease risk [65]. Knocking out MC4R in mice resulted in increased body weight and the development of late-onset hyperglycaemia and atherosclerotic plaque accumulation when fed a semisynthetic diet of 0.02% cholesterol [66]. MC4R knockout mice fed a western diet and infused with ANGII are susceptible to abdominal aortic aneurism, and MC4R mice, on an ApoE ko background, have exacerbated atherosclerosis [67].
Deficiency in MC1R (on an ApoE background) in mice fed a high-fat diet exacerbates atherosclerosis by increasing monocyte accumulation in the vessels along with elevated total plasma cholesterol [68]. Another study examining leukocyte-specific MCR1 deficiency showed an increase in circulating Ly6Chigh monocytes, B and CD4+ T cells, though this did not translate to changes in atherosclerotic plaque accumulation after HFD as there was a preference of these cells to migrate to the spleen [69].
Mice deficient in MC3R after ischemia/reperfusion injury showed increased cell adhesion and emigration as well as increased chemokines [70]. Activating MC3R using the agonist [D-Trp8]-y-MSH in APOE KO mice on a high-fat diet has also been shown to reduce plasma proinflammatory cytokines and leukocytes in both the circulation and in the aorta [71]. However, this intervention model did not result in changes in the expression of aortic adhesion molecules or changes in plaque accumulation and stability [71].
Is there also evidence for the role of the melanocortins in atherosclerosis? The expression of melanocortins and associated proteins have been examined in the blood vessels of both rodents and humans. In studies by Rinne, POMC was shown to be expressed in human plaques and was associated with the presence of macrophages [72,73]. Expression of α-MSH was detected in healthy and atherosclerotic human and murine vessels and decreased expression was seen in advanced atherosclerosis in ApoE KO mice on an HFD [74]. In contrast to declining α-MSH in advanced lesions, an increase in POMC was detected in unstable plaques in humans [72]. This paradox is explained by a decrease in the POMC processing enzymes, carboxypeptidase E (CPE) and α-amidating monooxygenase (PAM), and an increase in prolylcarboxypeptidase (PRCP), the enzyme that inactivates α-MSH [72]. The increase in POMC expression in plaques is most likely a compensatory mechanism responding to the inactivation of α-MSH or the decrease in expression of POMC processing enzymes. Western-diet-fed LDLR knockout mice overexpressing α-MSH and γ-MSH showed improved glucose tolerance, reduced plaque formation and reduced pro-inflammatory cytokines [75]. A recent study on T2DM patients showed that α-MSH is reduced in the circulation of diabetic patients compared to non-diabetic controls [76].
Despite some evidence that the melanocortins are involved in inflammation and vascular tone and that increasing α-MSH may be a beneficial treatment strategy, few studies have examined melanocortins in type 1 diabetic models. Whether the melanocortins are effective in ameliorating atherosclerosis in various diabetic models needs further investigation.
4.2. Diabetic Nephropathy and the Melanocortins
Chronic kidney disease (CKD) is the irreversible alteration of kidney structure and function over an extended period of time [77]. Diabetic nephropathy is the main cause of CKD, comprising up to 30–50% of all CKD, a condition without a definitive cure [61,77]. Very few studies to date have looked at the melanocortins in diabetic nephropathy despite the need for new therapeutics for CKD. ACTH was used for many years to stimulate the release of cortisol to treat conditions such as rheumatoid arthritis, lupus, multiple sclerosis and nephrotic syndrome but was soon after replaced by glucocorticoid steroids due to their lower cost [78]. Corticosteroids are commonly used in the treatment of kidney disease; however, patients can have side effects including an increased risk of developing diabetes mellitus [79]. ACTH was later found to have steroid-independent effects through binding to the MC3R and a superior side effect profile [61,80]. ACTH treatment improved proteinuria and glomerular filtration rate in patients with membranous glomerular nephropathy [81]. The use of ACTH was also examined in patients with diabetic nephropathy and showed stabilisation of renal function and reduction in urinary protein excretion [82]. The non-steroidal melanocortins, α-MSH, γ-MSH and β-MSH, can stimulate the melanocortin receptors and are anti-inflammatory and immunomodulatory. As a result, the melanocortins should have some effect on the kidney, especially as their receptors are expressed on many kidney cells [61,83]. MSH has been shown to be renoprotective in mouse models of human nephrotic syndrome, but no studies have been performed in models of diabetic nephropathy to date [84]. Therefore, due to their mechanistic potential which has been partially evidenced in studies to date, the melanocortins deserve further investigation into their ability to treat kidney disease, including diabetic nephropathy.
4.3. Diabetic Retinopathy and the Melanocortins
Diabetic retinopathy is the only cause of blindness that has increased in prevalence in the past 20 years [85]. Diabetic retinopathy is either non-proliferative or proliferative, where the latter involves the growth of newly formed vessels, and the former involves microvascular alterations in the retina [86]. Diabetic macular edema, a severe complication of diabetic retinopathy, is more likely to occur as diabetic retinopathy progresses. As with other diabetic complications, the initiator is hyperglycemia but other processes such as inflammation, oxidative stress and epigenetic modifications are involved in diabetic retinopathy [87].
Controlling blood glucose, blood pressure and serum cholesterol is critical to slowing the progression of diabetic retinopathy [88], though this can be difficult to achieve. The current treatments that inhibit the progression of DR are laser-induced photocoagulation and intravitreal injections of anti-VEGF or corticosteroids. These therapies are not ideal. Photocoagulation can destroy parts of the retina, while intravitreal injections require frequent visits to the doctor for transitory beneficial effects in only 50% of patients [87]. The increased risk of cataract formation and intraocular pressure limits the use of corticosteroids. Therefore, many studies have been investigating the use of other therapies including melanocortins to treat diabetic retinopathy. In 2011, it was shown that human type 2 diabetic patients with proliferative diabetic retinopathy have lower serum concentrations of ACTH than diabetic patients with non-proliferative disease, indicating that increasing melanocortins may be beneficial [89].
The role of the melanocortin receptors, in particular MC1, MC4 and MC5, detected in the retina, have been investigated. The expression of MC1 and MC5 receptors increases in retinal cells after exposure to high-glucose [90]. Cells treated with glucose in the presence of agonists for these receptors have been shown to restore antioxidant enzymes to normal levels. In another study, blocking MC1 and MC5 receptors in an STZ-induced diabetic retinopathy mouse model caused an increase in an array of cytokines, while receptor agonists returned these levels back to normal, indicating a protective role for these receptors in DR [91]. Fingolimod, a drug used to treat multiple sclerosis, was also examined for its effects in the retina as there is an increased risk of macular oedema in these patients [92]. However, a study in diabetic rats showed that fingolimod protected against retinal inflammation [93]. This drug was also predicted to bind to the melanocortin receptors using in silico modelling [91]. In an STZ mouse model of retinopathy, it was demonstrated that Fingolimod acts as an MCR1 and MCR5 agonist to reduce retinal angiogenesis and reduce retinal VEGF [91]. This drug was less effective in the presence of MC1R and MC5R antagonists [91].
Studies have also examined the effect of melanocortin treatment on diabetic retinopathy. In studies looking at injecting α-MSH intravitreally in STZ-induced diabetic rats showed that α-MSH prevented the breakdown of the blood–retina barrier by blocking the aberrant expression of tight junction genes, pro-inflammatory genes and oxidative stress [94,95]. In a model of retinal ischemia/reperfusion injury in akita mice, reduced retinal swelling and oxidative stress and increases in IL-10 were observed following intraperitoneal injection of α-MSH [96]. During ischemia, glutamate is elevated leading to the generation of ROS through ionotropic and metabotrophic receptors, leading to cell death. A study demonstrated the ability of α-MSH to prevent glutamate-induced cell death in chicken retinas in vitro (treated explants) and in vivo (intravitreal injections) [97]. This study showed that cell death is prevented through α-MSH blocking the upregulation of the pro-apoptotic micro RNA 194 [97]. One limitation of α-MSH is its short serum half-life. Therefore, the generation of analogs has been undertaken for use in DR. One such analog, PL-8331, was used in a mouse model of diabetic retinopathy [98]. Here, it was shown that PL-8331 was anti-inflammatory and enhanced the survival of retinal cells in vivo [98].
To date, research has focused on using α-MSH or MCR agonists; however, it is clear that further research into melanocortins is warranted for the treatment of diabetic retinopathy.
5. Conclusions
Melanocortins have shown promise in early animal and human studies in treating a variety of conditions inculpated in the aetiology and complications of diabetes. They have been found to be well-tolerated in large human trials and now, a number of melanocortins have made their way to market for various dermatological, sexual and metabolic disorders, although potential side effects such as nausea and skin flushing should be considered. Therefore, it is an opportune time for endocrinologists and scientists in the field to consider the benefits of melanocortins in assisting their patients to prevent or treat diabetes. The conclusion of this review is simple: melanocortins have a number of tangible positive effects on the development, progression and complications of diabetes. Further formulation work to ensure the specificity of melanocortin receptor activation and reformulations to maximise serum half-life should also be considerations for the further development of melanocortins in treating diabetes and its complications. However, if these and other potential hurdles are cleared, melanocortins may become a vital tool in the treatment of this burgeoning disease in the ensuing decades; thus, they warrant a more thorough and pronounced investigation in fields outside of dermatology.
Author Contributions
Conceptualization, G.N.R. and R.J.P.; writing—original draft preparation, G.N.R. and R.J.P.; writing—review and editing, G.N.R. and R.J.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Raelene Pickering is a Global Moderna Research Fellow. Gardner Robinson is supported by an Australian Government Research Training Program Scholarship.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- International Diabetes Federation. IDF Diabetes Atlas; International Diabetes Federation: Brussels, Belgium, 2021. [Google Scholar]
- Slominski, A.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef]
- Winget, M.D.; Ericson, M.D.; Freeman, K.T.; Haskell-Luevano, C. Single Nucleotide Polymorphisms in the Melanocortin His-Phe-Arg-Trp Sequences Decrease Tetrapeptide Potency and Efficacy. ACS Med. Chem. Lett. 2020, 11, 272–277. [Google Scholar] [CrossRef]
- Hadley, M.E.; Haskell-Luevano, C. The Proopiomelanocortin System. Ann. N. Y. Acad. Sci. 1999, 885, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Harno, E.; Ramamoorthy, T.G.; Coll, A.P.; White, A. POMC: The Physiological Power of Hormone Processing. Physiol. Rev. 2018, 98, 2381–2430. [Google Scholar] [CrossRef] [PubMed]
- Biebermann, H.; Castañeda, T.R.; van Landeghem, F.; von Deimling, A.; Escher, F.; Brabant, G.; Hebebrand, J.; Hinney, A.; Tschöp, M.H.; Grüters, A.; et al. A role for beta-melanocyte-stimulating hormone in human body-weight regulation. Cell Metab. 2006, 3, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Collet, T.-H.; Dubern, B.; Mokrosinski, J.; Connors, H.; Keogh, J.; Oliveira, E.; Henning, E.; Poitou Bernert, C.; Oppert, J.-M.; Tounian, P.; et al. Evaluation of a Melanocortin-4 Receptor (MC4R) agonist (Setmelanotide) in MC4R deficiency. Mol. Metab. 2017, 6, 1321–1329. [Google Scholar] [CrossRef] [PubMed]
- De Jonghe, B.C.; Hayes, M.R.; Zimmer, D.J.; Kanoski, S.E.; Grill, H.J.; Bence, K.K. Food intake reductions and increases in energetic responses by hindbrain leptin and melanotan II are enhanced in mice with POMC-specific PTP1B deficiency. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E644–E651. [Google Scholar] [CrossRef] [PubMed]
- Eerola, K.; Virtanen, S.; Vähätalo, L.; Ailanen, L.; Cai, M.; Hruby, V.; Savontaus, M.; Savontaus, E. Hypothalamic γ-melanocyte stimulating hormone gene delivery reduces fat mass in male mice. J. Endocrinol. 2018, 239, 19–31. [Google Scholar] [CrossRef]
- Kühnen, P.; Clément, K.; Wiegand, S.; Blankenstein, O.; Gottesdiener, K.; Martini, L.L.; Mai, K.; Blume-Peytavi, U.; Grüters, A.; Krude, H. Proopiomelanocortin Deficiency Treated with a Melanocortin-4 Receptor Agonist. N. Engl. J. Med. 2016, 375, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Kühnen, P.; Krude, H.; Biebermann, H. Melanocortin-4 Receptor Signalling: Importance for Weight Regulation and Obesity Treatment. Trends Mol. Med. 2019, 25, 136–148. [Google Scholar] [CrossRef]
- Lee, Y.S.; Challis, B.G.; Thompson, D.A.; Yeo, G.S.; Keogh, J.M.; Madonna, M.E.; Wraight, V.; Sims, M.; Vatin, V.; Meyre, D.; et al. A POMC variant implicates beta-melanocyte-stimulating hormone in the control of human energy balance. Cell Metab. 2006, 3, 135–140. [Google Scholar] [CrossRef]
- Quiñones, M.; Al-Massadi, O.; Folgueira, C.; Bremser, S.; Gallego, R.; Torres-Leal, L.; Haddad-Tóvolli, R.; García-Caceres, C.; Hernandez-Bautista, R.; Lam, B.Y.H.; et al. p53 in AgRP neurons is required for protection against diet-induced obesity via JNK1. Nat. Commun. 2018, 9, 3432. [Google Scholar] [CrossRef]
- van den Heuvel, J.K.; Eggels, L.; van Rozen, A.J.; Fliers, E.; Kalsbeek, A.; Adan, R.A.; la Fleur, S.E. Inhibitory Effect of the Melanocortin Receptor Agonist Melanotan-II (MTII) on Feeding Depends on Dietary Fat Content and not Obesity in Rats on Free-Choice Diets. Front. Behav. Neurosci. 2015, 9, 358. [Google Scholar] [CrossRef]
- Cui, H.; Sohn, J.W.; Gautron, L.; Funahashi, H.; Williams, K.W.; Elmquist, J.K.; Lutter, M. Neuroanatomy of melanocortin-4 receptor pathway in the lateral hypothalamic area. J. Comp. Neurol. 2012, 520, 4168–4183. [Google Scholar] [CrossRef]
- Caron, A.; Dungan Lemko, H.M.; Castorena, C.M.; Fujikawa, T.; Lee, S.; Lord, C.C.; Ahmed, N.; Lee, C.E.; Holland, W.L.; Liu, C.; et al. POMC neurons expressing leptin receptors coordinate metabolic responses to fasting via suppression of leptin levels. eLife 2018, 7, e33710. [Google Scholar] [CrossRef]
- Meneguetti, B.T.; Cardoso, M.H.; Ribeiro, C.F.A.; Felicio, M.R.; Pinto, I.B.; Santos, N.C.; Carvalho, C.M.E.; Franco, O.L. Neuropeptide receptors as potential pharmacological targets for obesity. Pharmacol. Ther. 2019, 196, 59–78. [Google Scholar] [CrossRef]
- Zemel, M.B. Agouti/melanocortin interactions with leptin pathways in obesity. Nutr. Rev. 1998, 56, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Ingalls, A.M.; Dickie, M.M.; Snell, G.D. Obese, a new mutation in the house mouse. J. Hered. 1950, 41, 317–318. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Cummings, B.P. Leptin therapy in type 2 diabetes. Diabetes Obes. Metab. 2013, 15, 607–612. [Google Scholar] [CrossRef]
- Backholer, K.; Smith, J.; Clarke, I.J. Melanocortins may stimulate reproduction by activating orexin neurons in the dorsomedial hypothalamus and kisspeptin neurons in the preoptic area of the ewe. Endocrinology 2009, 150, 5488–5497. [Google Scholar] [CrossRef]
- Boghossian, S.; Park, M.; York, D.A. Melanocortin activity in the amygdala controls appetite for dietary fat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R385–R393. [Google Scholar] [CrossRef]
- Campos, C.A.; Shiina, H.; Ritter, R.C. Central vagal afferent endings mediate reduction of food intake by melanocortin-3/4 receptor agonist. J. Neurosci. Off. J. Soc. Neurosci. 2014, 34, 12636–12645. [Google Scholar] [CrossRef]
- Chai, B.; Li, J.Y.; Zhang, W.; Wang, H.; Mulholland, M.W. Melanocortin-4 receptor activation inhibits c-Jun N-terminal kinase activity and promotes insulin signaling. Peptides 2009, 30, 1098–1104. [Google Scholar] [CrossRef] [PubMed]
- Cote, I.; Green, S.M.; Morgan, D.; Carter, C.S.; Tumer, N.; Scarpace, P.J. Activation of the central melanocortin system in rats persistently reduces body and fat mass independently of caloric reduction. Can. J. Physiol. Pharmacol. 2018, 96, 308–312. [Google Scholar] [CrossRef]
- Cote, I.; Sakarya, Y.; Kirichenko, N.; Morgan, D.; Carter, C.S.; Tumer, N.; Scarpace, P.J. Activation of the central melanocortin system chronically reduces body mass without the necessity of long-term caloric restriction. Can. J. Physiol. Pharmacol. 2017, 95, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.A.; Freeman, J.N.; Hall, J.E.; do Carmo, J.M. Control of appetite, blood glucose, and blood pressure during melanocortin-4 receptor activation in normoglycemic and diabetic NPY-deficient mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 314, R533–R539. [Google Scholar] [CrossRef] [PubMed]
- Do Carmo, J.M.; da Silva, A.A.; Rushing, J.S.; Pace, B.; Hall, J.E. Differential control of metabolic and cardiovascular functions by melanocortin-4 receptors in proopiomelanocortin neurons. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R359–R368. [Google Scholar] [CrossRef] [PubMed]
- Fani, L.; Bak, S.; Delhanty, P.; van Rossum, E.F.; van den Akker, E.L. The melanocortin-4 receptor as target for obesity treatment: A systematic review of emerging pharmacological therapeutic options. Int. J. Obes. 2014, 38, 163–169. [Google Scholar] [CrossRef]
- Gavini, C.K.; Jones, W.C., 2nd; Novak, C.M. Ventromedial hypothalamic melanocortin receptor activation: Regulation of activity energy expenditure and skeletal muscle thermogenesis. J. Physiol. 2016, 594, 5285–5301. [Google Scholar] [CrossRef] [PubMed]
- Monge-Roffarello, B.; Labbe, S.M.; Lenglos, C.; Caron, A.; Lanfray, D.; Samson, P.; Richard, D. The medial preoptic nucleus as a site of the thermogenic and metabolic actions of melanotan II in male rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R158–R166. [Google Scholar] [CrossRef] [PubMed]
- Monge-Roffarello, B.; Labbe, S.M.; Roy, M.C.; Lemay, M.L.; Coneggo, E.; Samson, P.; Lanfray, D.; Richard, D. The PVH as a site of CB1-mediated stimulation of thermogenesis by MC4R agonism in male rats. Endocrinology 2014, 155, 3448–3458. [Google Scholar] [CrossRef] [PubMed]
- Mul, J.D.; van Boxtel, R.; Bergen, D.J.; Brans, M.A.; Brakkee, J.H.; Toonen, P.W.; Garner, K.M.; Adan, R.A.; Cuppen, E. Melanocortin receptor 4 deficiency affects body weight regulation, grooming behavior, and substrate preference in the rat. Obesity 2012, 20, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, Y.B.; Vaughan, C.H.; Smith, B.J., Jr.; Song, C.K.; Baro, D.J.; Bartness, T.J. Central melanocortin stimulation increases phosphorylated perilipin A and hormone-sensitive lipase in adipose tissues. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R140–R149. [Google Scholar] [CrossRef] [PubMed]
- Song, C.K.; Vaughan, C.H.; Keen-Rhinehart, E.; Harris, R.B.; Richard, D.; Bartness, T.J. Melanocortin-4 receptor mRNA expressed in sympathetic outflow neurons to brown adipose tissue: Neuroanatomical and functional evidence. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R417–R428. [Google Scholar] [CrossRef] [PubMed]
- Toda, C.; Shiuchi, T.; Lee, S.; Yamato-Esaki, M.; Fujino, Y.; Suzuki, A.; Okamoto, S.; Minokoshi, Y. Distinct effects of leptin and a melanocortin receptor agonist injected into medial hypothalamic nuclei on glucose uptake in peripheral tissues. Diabetes 2009, 58, 2757–2765. [Google Scholar] [CrossRef]
- Wang, J.; Ling, S.; Usami, T.; Murata, T.; Narita, K.; Higuchi, T. Effects of ghrelin, corticotrophin-releasing hormone, and melanotan-II on food intake in rats with paraventricular nucleus lesions. Exp. Clin. Endocrinol. Diabetes Off. J. Ger. Soc. Endocrinol. Ger. Diabetes Assoc. 2007, 115, 669–673. [Google Scholar] [CrossRef]
- Xu, Y.; Wu, Z.; Sun, H.; Zhu, Y.; Kim, E.R.; Lowell, B.B.; Arenkiel, B.R.; Xu, Y.; Tong, Q. Glutamate mediates the function of melanocortin receptor 4 on Sim1 neurons in body weight regulation. Cell Metab. 2013, 18, 860–870. [Google Scholar] [CrossRef]
- Ye, Z.Y.; Li, D.P. Activation of the melanocortin-4 receptor causes enhanced excitation in presympathetic paraventricular neurons in obese Zucker rats. Regul. Pept. 2011, 166, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Collazo, R.; Gao, Y.; Li, G.; Scarpace, P.J. Intermittent MTII application evokes repeated anorexia and robust fat and weight loss. Peptides 2010, 31, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Albrecht, E.; Schering, L.; Kuehn, C.; Yang, R.; Zhao, Z.; Maak, S. Agouti Signaling Protein and Its Receptors as Potential Molecular Markers for Intramuscular and Body Fat Deposition in Cattle. Front. Physiol. 2018, 9, 172. [Google Scholar] [CrossRef]
- Grundlingh, J.; Dargan, P.I.; El-Zanfaly, M.; Wood, D.M. 2,4-dinitrophenol (DNP): A weight loss agent with significant acute toxicity and risk of death. J. Med. Toxicol. 2011, 7, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Colman, E. Dinitrophenol and obesity: An early twentieth-century regulatory dilemma. Regul. Toxicol. Pharmacol. 2007, 48, 115–117. [Google Scholar] [CrossRef]
- Laiho, L.; Murray, J.F. The Multifaceted Melanocortin Receptors. Endocrinology 2022, 163, bqac083. [Google Scholar] [CrossRef] [PubMed]
- Clément, K.; Biebermann, H.; Farooqi, I.S.; Van der Ploeg, L.; Wolters, B.; Poitou, C.; Puder, L.; Fiedorek, F.; Gottesdiener, K.; Kleinau, G.; et al. MC4R agonism promotes durable weight loss in patients with leptin receptor deficiency. Nat. Med. 2018, 24, 551–555. [Google Scholar] [CrossRef]
- Haws, R.; Brady, S.; Davis, E.; Fletty, K.; Yuan, G.; Gordon, G.; Stewart, M.; Yanovski, J. Effect of setmelanotide, a melanocortin-4 receptor agonist, on obesity in Bardet-Biedl syndrome. Diabetes Obes. Metab. 2020, 22, 2133–2140. [Google Scholar] [CrossRef]
- Forsythe, E.; Haws, R.M.; Argente, J.; Beales, P.; Martos-Moreno, G.; Dollfus, H.; Chirila, C.; Gnanasakthy, A.; Buckley, B.C.; Mallya, U.G.; et al. Quality of life improvements following one year of setmelanotide in children and adult patients with Bardet-Biedl syndrome: Phase 3 trial results. Orphanet J. Rare Dis. 2023, 18, 12. [Google Scholar] [CrossRef] [PubMed]
- Haqq, A.M.; Chung, W.K.; Dollfus, H.; Haws, R.M.; Martos-Moreno, G.; Poitou, C.; Yanovski, J.A.; Mittleman, R.S.; Yuan, G.; Forsythe, E.; et al. Efficacy and safety of setmelanotide, a melanocortin-4 receptor agonist, in patients with Bardet-Biedl syndrome and Alström syndrome: A multicentre, randomised, double-blind, placebo-controlled, phase 3 trial with an open-label period. Lancet Diabetes Endocrinol. 2022, 10, 859–868. [Google Scholar] [CrossRef]
- Kühnen, P.; Wabitsch, M.; von Schnurbein, J.; Chirila, C.; Mallya, U.G.; Callahan, P.; Gnanasakthy, A.; Poitou, C.; Krabusch, P.M.; Stewart, M.; et al. Quality of life outcomes in two phase 3 trials of setmelanotide in patients with obesity due to LEPR or POMC deficiency. Orphanet J. Rare Dis. 2022, 17, 38. [Google Scholar] [CrossRef]
- Fehm, H.L.; Smolnik, R.; Kern, W.; McGregor, G.P.; Bickel, U.; Born, J. The melanocortin melanocyte-stimulating hormone/adrenocorticotropin(4–10) decreases body fat in humans. J. Clin. Endocrinol. Metab. 2001, 86, 1144–1148. [Google Scholar]
- Spana, C.; Jordan, R.; Fischkoff, S. Effect of bremelanotide on body weight of obese women: Data from two phase 1 randomized controlled trials. Diabetes Obes. Metab. 2022, 24, 1084–1093. [Google Scholar] [CrossRef]
- Chen, K.Y.; Muniyappa, R.; Abel, B.S.; Mullins, K.P.; Staker, P.; Brychta, R.J.; Zhao, X.; Ring, M.; Psota, T.L.; Cone, R.D.; et al. RM-493, a melanocortin-4 receptor (MC4R) agonist, increases resting energy expenditure in obese individuals. J. Clin. Endocrinol. Metab. 2015, 100, 1639–1645. [Google Scholar] [CrossRef]
- Pierroz, D.D.; Ziotopoulou, M.; Ungsunan, L.; Moschos, S.; Flier, J.S.; Mantzoros, C.S. Effects of acute and chronic administration of the melanocortin agonist MTII in mice with diet-induced obesity. Diabetes 2002, 51, 1337–1345. [Google Scholar] [CrossRef]
- Kievit, P.; Halem, H.; Marks, D.L.; Dong, J.Z.; Glavas, M.M.; Sinnayah, P.; Pranger, L.; Cowley, M.A.; Grove, K.L.; Culler, M.D. Chronic treatment with a melanocortin-4 receptor agonist causes weight loss, reduces insulin resistance, and improves cardiovascular function in diet-induced obese rhesus macaques. Diabetes 2013, 62, 490–497. [Google Scholar] [CrossRef]
- Zhang, Y.; Matheny, M.; Tümer, N.; Scarpace, P.J. Aged-obese rats exhibit robust responses to a melanocortin agonist and antagonist despite leptin resistance. Neurobiol. Aging 2004, 25, 1349–1360. [Google Scholar] [CrossRef]
- Blüher, S.; Ziotopoulou, M.; Bullen, J.W., Jr.; Moschos, S.J.; Ungsunan, L.; Kokkotou, E.; Maratos-Flier, E.; Mantzoros, C.S. Responsiveness to peripherally administered melanocortins in lean and obese mice. Diabetes 2004, 53, 82–90. [Google Scholar] [CrossRef]
- Weiss, T.; Carr, R.D.; Pal, S.; Yang, L.; Sawhney, B.; Boggs, R.; Rajpathak, S.; Iglay, K. Real-World Adherence and Discontinuation of Glucagon-like Peptide-1 Receptor Agonists Therapy in Type 2 Diabetes Mellitus Patients in the United States. Patient Prefer. Adherence 2020, 14, 2337–2345. [Google Scholar] [CrossRef]
- Mannucci, E.; Dicembrini, I.; Nreu, B.; Monami, M. Glucagon-like peptide-1 receptor agonists and cardiovascular outcomes in patients with and without prior cardiovascular events: An updated meta-analysis and subgroup analysis of randomized controlled trials. Diabetes Obes. Metab. 2019, 22, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.; Cuthbertson, D.J.; Wilding, J.P. Newer GLP-1 receptor agonists and obesity-diabetes. Peptides 2018, 100, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.; Chen, B.; Shaffner, J.; Dworkin, L.D.; Gong, R. Melanocortin System in Kidney Homeostasis and Disease: Novel Therapeutic Opportunities. Front. Physiol. 2021, 12, 651236. [Google Scholar] [CrossRef] [PubMed]
- Langan, E.A.; Nie, Z.; Rhodes, L.E. Melanotropic peptides: More than just ‘Barbie drugs’ and ‘sun-tan jabs’? Br. J. Dermatol. 2010, 163, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Copperi, F.; Kim, J.D.; Diano, S. Role of the Melanocortin System in the Central Regulation of Cardiovascular Functions. Front. Physiol. 2021, 12, 725709. [Google Scholar] [CrossRef] [PubMed]
- Sull, J.W.; Kim, G.; Jee, S.H. Association of MC4R (rs17782313) with diabetes and cardiovascular disease in Korean men and women. BMC Med. Genet. 2020, 21, 160. [Google Scholar] [CrossRef] [PubMed]
- Blauw, L.L.; Noordam, R.; van der Laan, S.W.; Trompet, S.; Kooijman, S.; van Heemst, D.; Jukema, J.W.; van Setten, J.; de Borst, G.J.; Tybjærg-Hansen, A.; et al. Common Genetic Variation in MC4R Does Not Affect Atherosclerotic Plaque Phenotypes and Cardiovascular Disease Outcomes. J. Clin. Med. 2021, 10, 932. [Google Scholar] [CrossRef]
- Lede, V.; Franke, C.; Meusel, A.; Teupser, D.; Ricken, A.; Thiery, J.; Schiller, J.; Huster, D.; Schöneberg, T.; Schulz, A. Severe Atherosclerosis and Hypercholesterolemia in Mice Lacking Both the Melanocortin Type 4 Receptor and Low Density Lipoprotein Receptor. PLoS ONE 2016, 11, e0167888. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Okuma, H.; Nakamura, S.; Uchinuma, H.; Kaga, S.; Nakajima, H.; Ogawa, Y.; Tsuchiya, K. Melanocortin-4 receptor in macrophages attenuated angiotensin II-induced abdominal aortic aneurysm in mice. Sci. Rep. 2023, 13, 19768. [Google Scholar] [CrossRef]
- Rinne, P.; Kadiri, J.J.; Velasco-Delgado, M.; Nuutinen, S.; Viitala, M.; Hollmén, M.; Rami, M.; Savontaus, E.; Steffens, S. Melanocortin 1 Receptor Deficiency Promotes Atherosclerosis in Apolipoprotein E(−/−) Mice. Arter. Thromb. Vasc. Biol. 2018, 38, 313–323. [Google Scholar] [CrossRef]
- Kadiri, J.J.; Tadayon, S.; Thapa, K.; Suominen, A.; Hollmén, M.; Rinne, P. Melanocortin 1 Receptor Deficiency in Hematopoietic Cells Promotes the Expansion of Inflammatory Leukocytes in Atherosclerotic Mice. Front. Immunol. 2021, 12, 774013. [Google Scholar] [CrossRef]
- Leoni, G.; Patel, H.B.; Sampaio, A.L.; Gavins, F.N.; Murray, J.F.; Grieco, P.; Getting, S.J.; Perretti, M. Inflamed phenotype of the mesenteric microcirculation of melanocortin type 3 receptor-null mice after ischemia-reperfusion. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2008, 22, 4228–4238. [Google Scholar] [CrossRef]
- Kadiri, J.J.; Thapa, K.; Kaipio, K.; Cai, M.; Hruby, V.J.; Rinne, P. Melanocortin 3 receptor activation with [D-Trp8]-γ-MSH suppresses inflammation in apolipoprotein E deficient mice. Eur. J. Pharmacol. 2020, 880, 173186. [Google Scholar] [CrossRef]
- Rinne, P.; Lyytikäinen, L.-P.; Raitoharju, E.; Kadiri, J.J.; Kholova, I.; Kähönen, M.; Lehtimäki, T.; Oksala, N. Pro-opiomelanocortin and its Processing Enzymes Associate with Plaque Stability in Human Atherosclerosis—Tampere Vascular Study. Sci. Rep. 2018, 8, 15078. [Google Scholar] [CrossRef]
- Rinne, P.; Silvola, J.M.; Hellberg, S.; Stahle, M.; Liljenback, H.; Salomaki, H.; Koskinen, E.; Nuutinen, S.; Saukko, P.; Knuuti, J.; et al. Pharmacological activation of the melanocortin system limits plaque inflammation and ameliorates vascular dysfunction in atherosclerotic mice. Arter. Thromb. Vasc. Biol. 2014, 34, 1346–1354. [Google Scholar] [CrossRef]
- Rinne, P.; Ahola-Olli, A.; Nuutinen, S.; Koskinen, E.; Kaipio, K.; Eerola, K.; Juonala, M.; Kähönen, M.; Lehtimäki, T.; Raitakari, O.T.; et al. Deficiency in Melanocortin 1 Receptor Signaling Predisposes to Vascular Endothelial Dysfunction and Increased Arterial Stiffness in Mice and Humans. Arter. Thromb. Vasc. Biol. 2015, 35, 1678–1686. [Google Scholar] [CrossRef] [PubMed]
- Nuutinen, S.; Ailanen, L.; Savontaus, E.; Rinne, P. Melanocortin overexpression limits diet-induced inflammation and atherosclerosis in LDLR(−/−) mice. J. Endocrinol. 2018, 236, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Ma, S.; Guo, S.; Li, N.; Liu, D.; Wang, H.; Zhai, N.; Zhang, Y. Altered features of neurotransmitters: NPY, α-MSH, and AgRP in type 2 diabetic patients with hypertension. J. Int. Med. Res. 2020, 48, 300060520919580. [Google Scholar] [CrossRef] [PubMed]
- Webster, A.C.; Nagler, E.V.; Morton, R.L.; Masson, P. Chronic Kidney Disease. Lancet 2017, 389, 1238–1252. [Google Scholar] [CrossRef] [PubMed]
- Montero-Melendez, T. ACTH: The forgotten therapy. Semin. Immunol. 2015, 27, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-X.; Cummins, C.L. Fresh insights into glucocorticoid-induced diabetes mellitus and new therapeutic directions. Nat. Rev. Endocrinol. 2022, 18, 540–557. [Google Scholar] [CrossRef] [PubMed]
- Getting, S.J.; Christian, H.C.; Flower, R.J.; Perretti, M. Activation of melanocortin type 3 receptor as a molecular mechanism for adrenocorticotropic hormone efficacy in gouty arthritis. Arthritis Rheum. 2002, 46, 2765–2775. [Google Scholar] [CrossRef]
- Berg, A.L.; Nilsson-Ehle, P.; Arnadottir, M. Beneficial effects of ACTH on the serum lipoprotein profile and glomerular function in patients with membranous nephropathy. Kidney Int. 1999, 56, 1534–1543. [Google Scholar] [CrossRef]
- Tumlin, J.A.; Galphin, C.M.; Rovin, B.H. Advanced diabetic nephropathy with nephrotic range proteinuria: A pilot study of the long-term efficacy of subcutaneous ACTH gel on proteinuria, progression of CKD, and urinary levels of VEGF and MCP-1. J. Diabetes Res. 2013, 2013, 489869. [Google Scholar] [CrossRef] [PubMed]
- Bergwall, L.; Wallentin, H.; Elvin, J.; Liu, P.; Boi, R.; Sihlbom, C.; Hayes, K.; Wright, D.; Haraldsson, B.; Nyström, J.; et al. Amplification of the Melanocortin-1 Receptor in Nephrotic Syndrome Identifies a Target for Podocyte Cytoskeleton Stabilization. Sci. Rep. 2018, 8, 15731. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Wang, P.; Chang, M.; Chen, B.; Ge, Y.; Malhotra, D.K.; Dworkin, L.D.; Gong, R. Melanocortin therapy ameliorates podocytopathy and proteinuria in experimental focal segmental glomerulosclerosis involving a podocyte specific non-MC1R-mediated melanocortinergic signaling. Clin. Sci. 2020, 134, 695–710. [Google Scholar] [CrossRef]
- Burton, M.J.; Ramke, J.; Marques, A.P.; Bourne, R.R.A.; Congdon, N.; Jones, I.; Ah Tong, B.A.M.; Arunga, S.; Bachani, D.; Bascaran, C.; et al. The Lancet Global Health Commission on Global Eye Health: Vision beyond 2020. Lancet Glob. Health 2021, 9, e489–e551. [Google Scholar] [CrossRef]
- Fung, T.H.; Patel, B.; Wilmot, E.G.; Amoaku, W.M. Diabetic retinopathy for the non-ophthalmologist. Clin. Med. 2022, 22, 112–116. [Google Scholar] [CrossRef]
- Roy, S.; Kern, T.S.; Song, B.; Stuebe, C. Mechanistic Insights into Pathological Changes in the Diabetic Retina: Implications for Targeting Diabetic Retinopathy. Am. J. Pathol. 2017, 187, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, R.; Harris, A.; Ciulla, T.A.; Kheradiya, N.; Winston, D.M.; Wirostko, B. Diabetic macular oedema: Physical, physiological and molecular factors contribute to this pathological process. Acta Ophthalmol. 2010, 88, 279–291. [Google Scholar] [CrossRef]
- Obulkasim, M.; Turdi, A.; Amat, N.; Haxim, M.; Eziz, R.; Haji, H.; Li, L.; Chen, X.; Upur, H.; Ren, J. Neuroendocrine–immune disorder in type 2 diabetic patients with retinopathy. Clin. Exp. Pharmacol. Physiol. 2011, 38, 229–232. [Google Scholar] [CrossRef]
- Maisto, R.; Gesualdo, C.; Trotta, M.; Grieco, P.; Testa, F.; Simonelli, F.; Barcia, J.; Amico, M.; Filippo, C.; Rossi, S. Melanocortin receptor agonists MCR 1-5 protect photoreceptors from high-glucose damage and restore antioxidant enzymes in primary retinal cell culture. J. Cell. Mol. Med. 2016, 21, 968–974. [Google Scholar] [CrossRef]
- Rossi, S.; Maisto, R.; Gesualdo, C.; Trotta, M.C.; Ferraraccio, F.; Kaneva, M.K.; Getting, S.J.; Surace, E.; Testa, F.; Simonelli, F.; et al. Activation of Melanocortin Receptors MC 1 and MC 5 Attenuates Retinal Damage in Experimental Diabetic Retinopathy. Mediat. Inflamm. 2016, 2016, 7368389. [Google Scholar] [CrossRef]
- Mandal, P.; Gupta, A.; Fusi-Rubiano, W.; Keane, P.A.; Yang, Y. Fingolimod: Therapeutic mechanisms and ocular adverse effects. Eye 2017, 31, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Yan, H. FTY720 Attenuates Retinal Inflammation and Protects Blood–Retinal Barrier in Diabetic Rats. Investig. Ophthalmol. Vis. Sci. 2016, 57, 1254–1263. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Yang, Q.; Hou, M.; Han, Q.; Zhang, H.; Wang, J.; Qi, C.; Bo, Q.; Ru, Y.; Yang, W.; et al. A-Melanocyte-Stimulating Hormone Protects Early Diabetic Retina from Blood-Retinal Barrier Breakdown and Vascular Leakage via MC4R. Cell. Physiol. Biochem. 2018, 45, 505–522. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.B.; Cheng, L.; Bi, H.E.; Zhang, Z.Q.; Yao, J.; Zhou, X.Z.; Jiang, Q. Alpha-melanocyte stimulating hormone protects retinal pigment epithelium cells from oxidative stress through activation of melanocortin 1 receptor-Akt-mTOR signaling. Biochem. Biophys. Res. Commun. 2014, 443, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Goit, R.K.; Taylor, A.W.; Lo, A.C.Y. Anti-inflammatory α-Melanocyte-Stimulating Hormone Protects Retina After Ischemia/Reperfusion Injury in Type I Diabetes. Front. Neurosci. 2022, 16, 799739. [Google Scholar] [CrossRef]
- Zhang, Y.; Bo, Q.; Wu, W.; Xu, C.; Yu, G.; Ma, S.; Yang, Q.; Cao, Y.; Han, Q.; Ru, Y.; et al. α-Melanocyte-stimulating hormone prevents glutamate excitotoxicity in developing chicken retina via MC4R-mediated down-regulation of microRNA-194. Sci. Rep. 2015, 5, 15812. [Google Scholar] [CrossRef]
- Ng, T.F.; Taylor, A.W. Stimulating the Melanocortin System in Uveitis and Diabetes Preserves the Structure and Anti-Inflammatory Activity of the Retina. Int. J. Mol. Sci. 2023, 24, 6928. [Google Scholar] [CrossRef]
Figure 1.
Processing of POMC by Prohormone Convertases (PCs) leading to the production of melanocortins and other neurohormones. The short length melanocortins most applicable to clinical use in the settings of diabetes and obesity are highlighted. ACTH: Adrenocorticotropic Hormone. CLIP: Corticotropin-like Intermediate [Lobe] Peptide.
Figure 1.
Processing of POMC by Prohormone Convertases (PCs) leading to the production of melanocortins and other neurohormones. The short length melanocortins most applicable to clinical use in the settings of diabetes and obesity are highlighted. ACTH: Adrenocorticotropic Hormone. CLIP: Corticotropin-like Intermediate [Lobe] Peptide.

Figure 2.
General effects of the melanocortins on the human body.

Figure 3.
Control of appetite by opposing actions of POMC and AgRP/NPY neurons in the arcuate nucleus. Signalling controlling appetite and food intake in the arcuate nucleus. Following stimulation by ghrelin, AgRP/NPY neurons release AgRP, NPY (Neuropeptide Y) and GABA (Gamma-Aminobutyric acid) to directly and indirectly antagonise downstream satiety neurons and their receptors, including MC4R, increasing feeding. AgRP/NPY neurons also directly inhibit POMC neurons. Following stimulation by leptin, POMC neurons release melanocortins (including α-MSH) to act at MC4 receptors on downstream satiety neurons, increasing satiety.
Figure 3.
Control of appetite by opposing actions of POMC and AgRP/NPY neurons in the arcuate nucleus. Signalling controlling appetite and food intake in the arcuate nucleus. Following stimulation by ghrelin, AgRP/NPY neurons release AgRP, NPY (Neuropeptide Y) and GABA (Gamma-Aminobutyric acid) to directly and indirectly antagonise downstream satiety neurons and their receptors, including MC4R, increasing feeding. AgRP/NPY neurons also directly inhibit POMC neurons. Following stimulation by leptin, POMC neurons release melanocortins (including α-MSH) to act at MC4 receptors on downstream satiety neurons, increasing satiety.

Figure 4.
Stimulation of Lipolysis in white adipose tissue (WAT) by melanocortins. Melanocortins stimulate lipolysis in peripheral white adipose tissue through two related, yet distinct processes. In the first process, following stimulation by melanocortins (including α-MSH) at cell surface G-protein coupled receptors (MC2R & MC5R), an intracellular signalling pathway is mediated through cAMP and protein kinase (PKA), leading to the release of the lipolytic enzyme lipase, which in turn breaks down triglycerides into free fatty acids (FFAs) and glycerol which are then exported into the bloodstream to be metabolised. In the second process neurons innervating WAT are activated by melanocortins through MC4R & MC5R to release noradrenaline. Noradrenaline activates β 1-3-adrenergic receptors (βadrR1-3) leading to a similar intracellular signalling cascade as the first process.
Figure 4.
Stimulation of Lipolysis in white adipose tissue (WAT) by melanocortins. Melanocortins stimulate lipolysis in peripheral white adipose tissue through two related, yet distinct processes. In the first process, following stimulation by melanocortins (including α-MSH) at cell surface G-protein coupled receptors (MC2R & MC5R), an intracellular signalling pathway is mediated through cAMP and protein kinase (PKA), leading to the release of the lipolytic enzyme lipase, which in turn breaks down triglycerides into free fatty acids (FFAs) and glycerol which are then exported into the bloodstream to be metabolised. In the second process neurons innervating WAT are activated by melanocortins through MC4R & MC5R to release noradrenaline. Noradrenaline activates β 1-3-adrenergic receptors (βadrR1-3) leading to a similar intracellular signalling cascade as the first process.

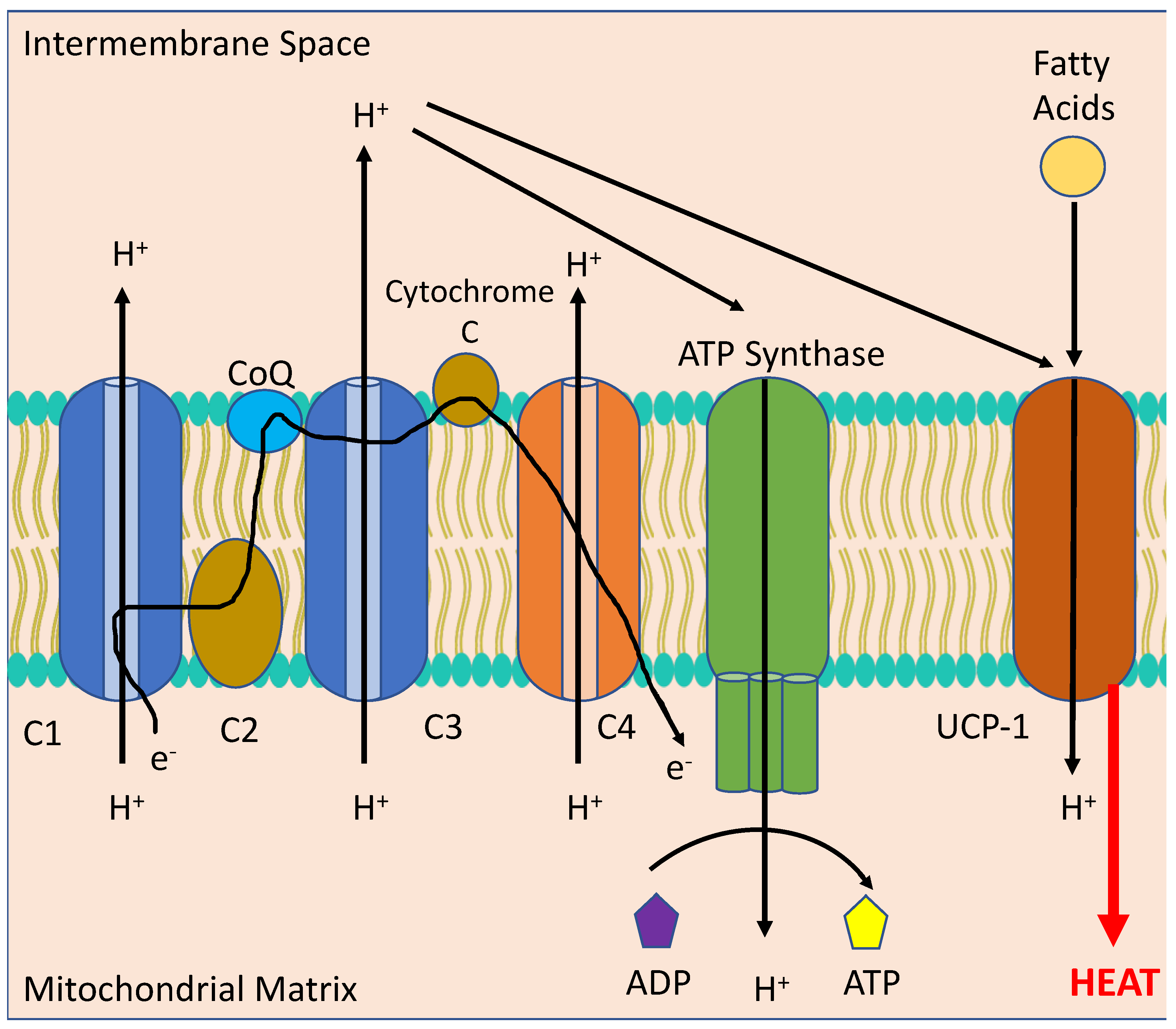
Figure 5.
Thermogenesis in the mitochondrion of a Brown Adipocyte. Melanocortins stimulate the activation of uncoupling protein 1 (UCP-1) via sympathetic stimulation of neurons innervating Brown Adipose Tissue (BAT) to uncouple the electron transport chain (consisting of C1, C2, Co-enzyme Q, C3, Cytochrome C and C4) from converting adenosine diphosphate (ADP) to adenosine triphosphate (ATP) through ATP Synthase to instead producing heat from fatty acids, leading to reduced body weight.
Figure 5.
Thermogenesis in the mitochondrion of a Brown Adipocyte. Melanocortins stimulate the activation of uncoupling protein 1 (UCP-1) via sympathetic stimulation of neurons innervating Brown Adipose Tissue (BAT) to uncouple the electron transport chain (consisting of C1, C2, Co-enzyme Q, C3, Cytochrome C and C4) from converting adenosine diphosphate (ADP) to adenosine triphosphate (ATP) through ATP Synthase to instead producing heat from fatty acids, leading to reduced body weight.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Robinson, G.N.; Pickering, R.J. Melanocortins and Their Potential for the Treatment, Prevention and Amelioration of Complications of Diabetes. Diabetology 2024, 5, 69-84. https://0-doi-org.brum.beds.ac.uk/10.3390/diabetology5010006
AMA Style
Robinson GN, Pickering RJ. Melanocortins and Their Potential for the Treatment, Prevention and Amelioration of Complications of Diabetes. Diabetology. 2024; 5(1):69-84. https://0-doi-org.brum.beds.ac.uk/10.3390/diabetology5010006
Chicago/Turabian StyleRobinson, Gardner N., and Raelene J. Pickering. 2024. "Melanocortins and Their Potential for the Treatment, Prevention and Amelioration of Complications of Diabetes" Diabetology 5, no. 1: 69-84. https://0-doi-org.brum.beds.ac.uk/10.3390/diabetology5010006