
The Impact of Exercise Serum on Selected Parameters of CD4+ T Cell Metabolism

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Exercise Intervention and Blood Sampling
2.3. Isolation, Activation, and Cell Culture Experiments
2.4. Real-Time Measurement of Oxygen Concentration
2.5. Analysis of Initial Cell Culture Condition and Supernatants
2.6. RNA Extraction and Real-Time Reverse Transcription PCR
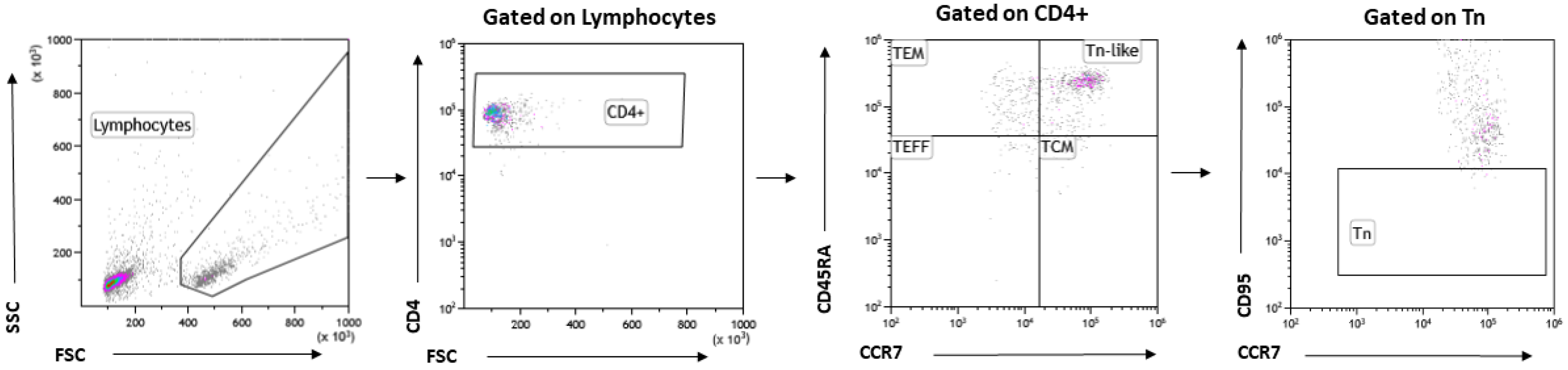
2.7. Cell Phenotyping by Flow Cytometry
2.8. Cell Count and Cell Growth
2.9. Statistical Analysis
3. Results
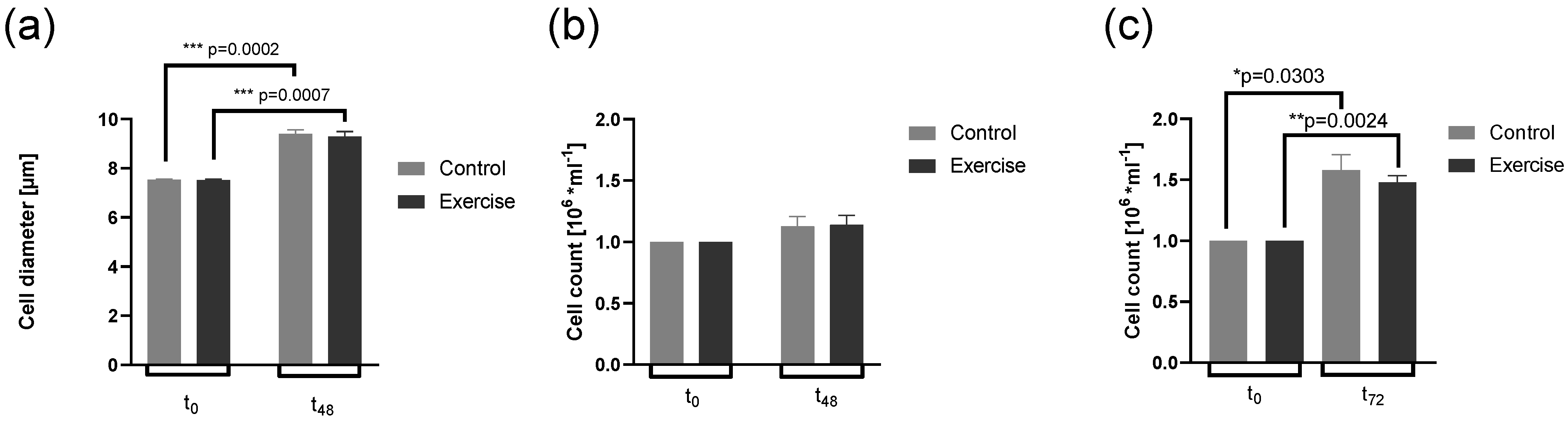
3.1. Cell Growth and Cell Counts Are Not Altered by Exercise Serum
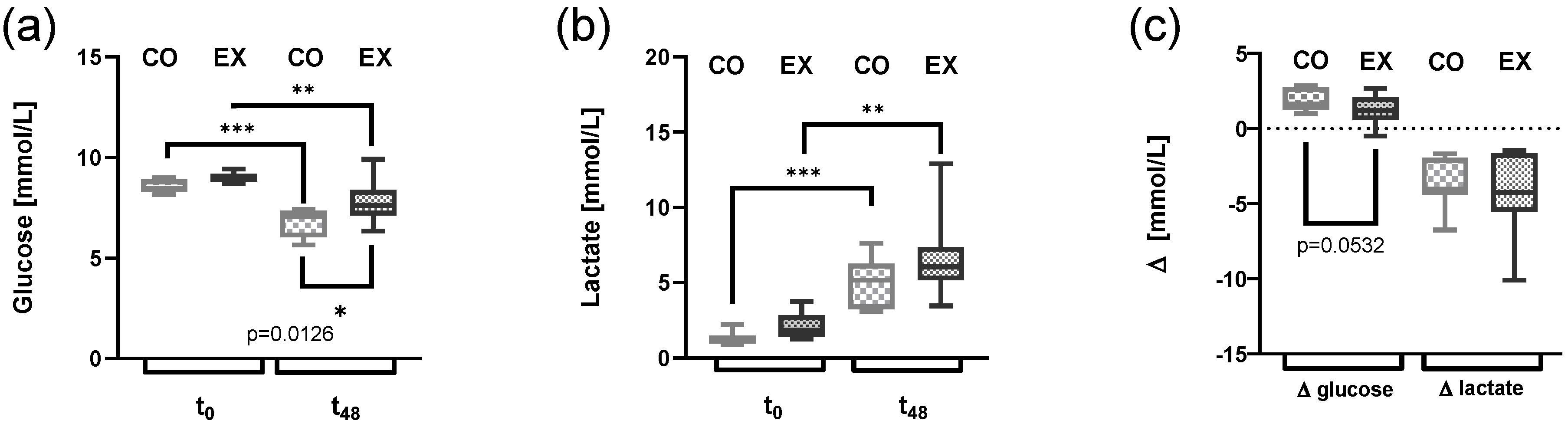
3.2. Glucose Consumption Is Altered by Exercise Serum
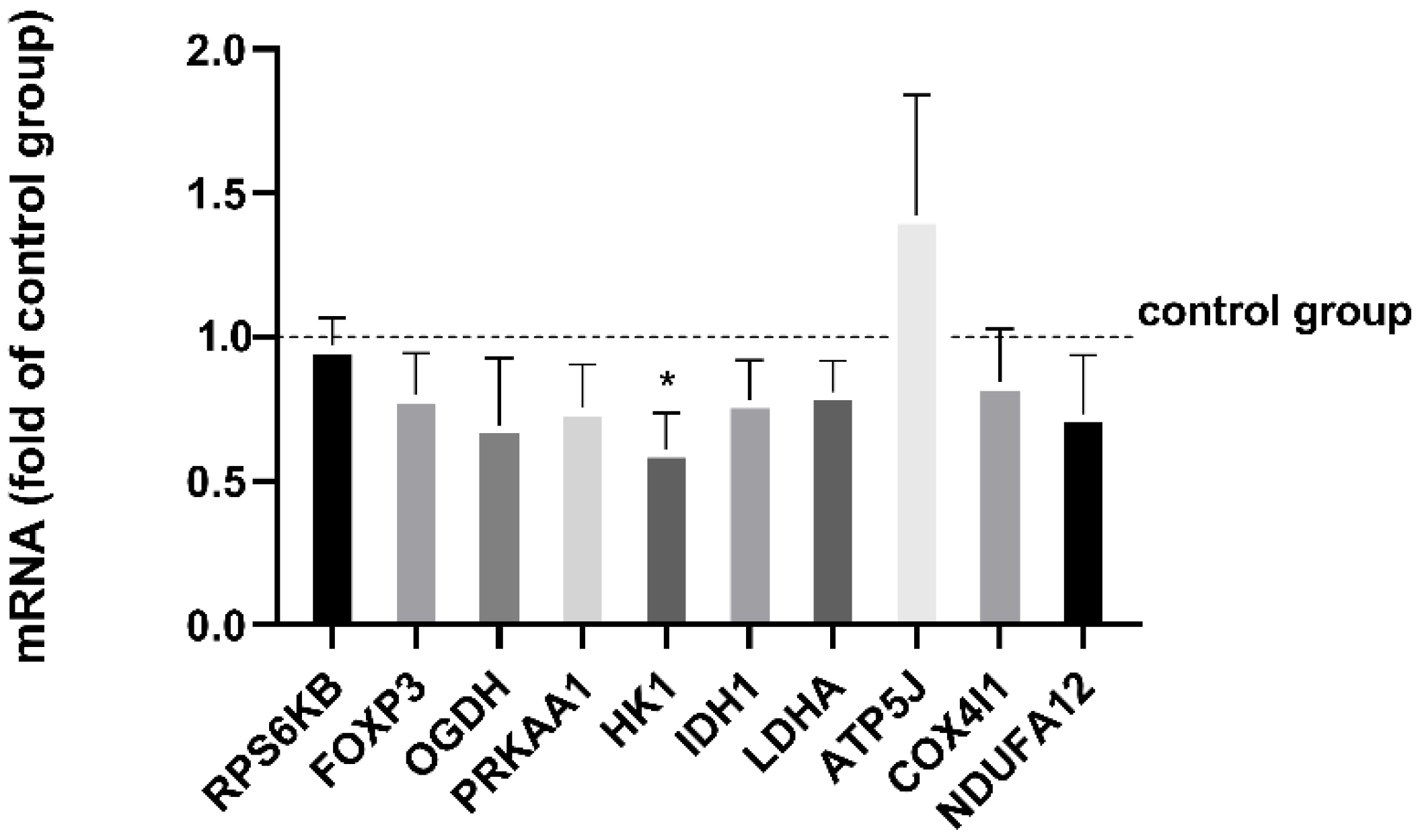
3.3. mRNA Expression of HK1 Is Reduced by Exercise Serum
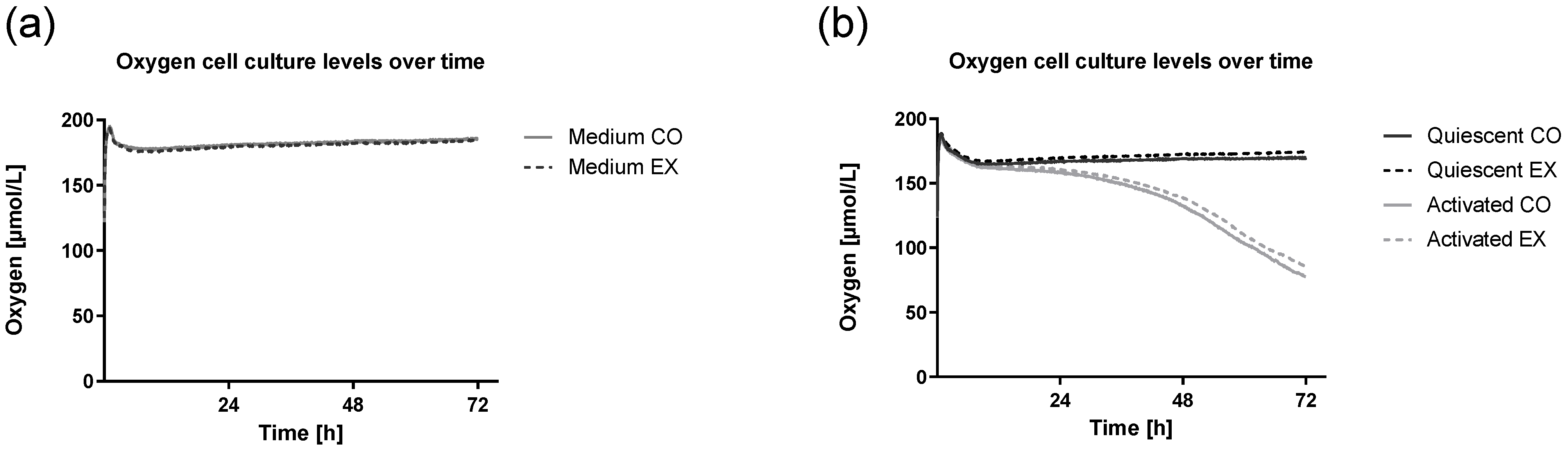
3.4. Cell Culture Oxygen Levels Following Activation Are Not Altered by Exercise Serum
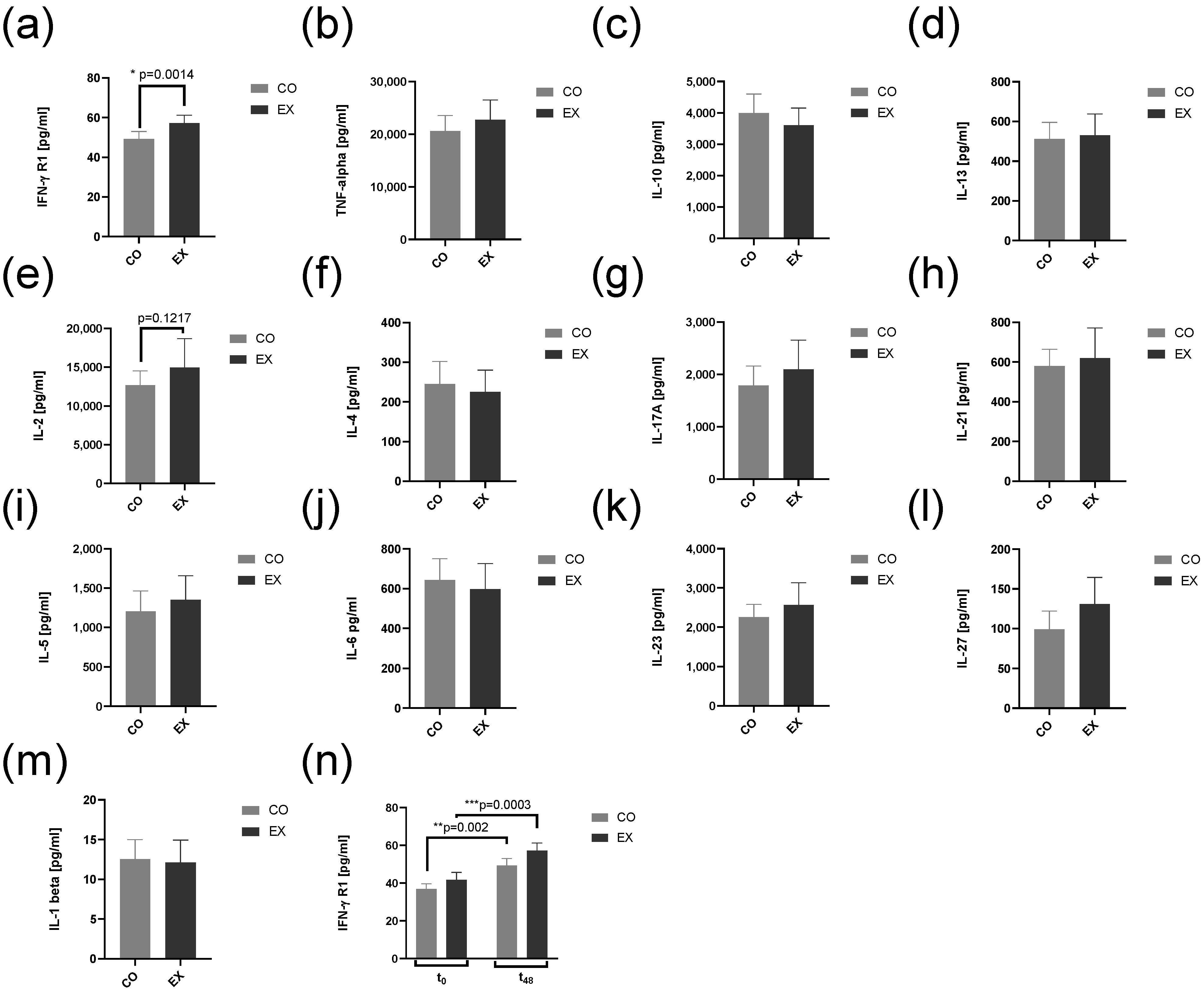
3.5. IFN-Gamma-R1 Levels Are Increased by Exercise Condition
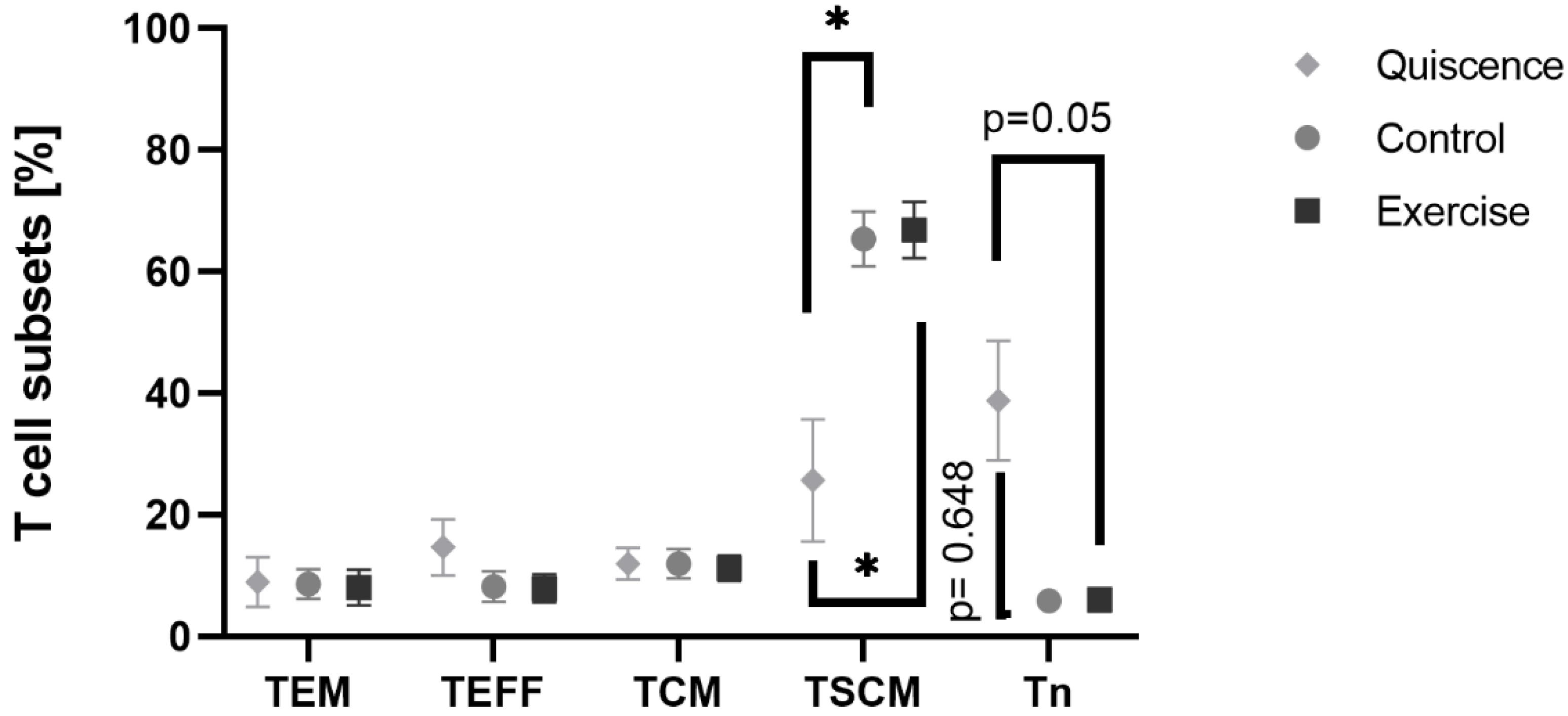
3.6. Exercise Serum Does Not Influence T Cell Phenotype and Activation Markers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mobasheri, A.; Rayman, M.P.; Gualillo, O.; Sellam, J.; van der Kraan, P.; Fearon, U. The role of metabolism in the pathogenesis of osteoarthritis. Nat. Rev. Rheumatol. 2017, 13, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Kolan, S.S.; Li, G.; Wik, J.A.; Malachin, G.; Guo, S.; Kolan, P.; Skålhegg, B.S. Cellular metabolism dictates T cell effector function in health and disease. Scand. J. Immunol. 2020, 92, e12956. [Google Scholar] [CrossRef] [PubMed]
- Bantug, G.R.; Galluzzi, L.; Kroemer, G.; Hess, C. The spectrum of T cell metabolism in health and disease. Nat. Rev. Immunol. 2018, 18, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Patel, C.H.; Leone, R.D.; Horton, M.R.; Powell, J.D. Targeting metabolism to regulate immune responses in autoimmunity and cancer. Nat. Rev. Drug Discov. 2019, 18, 669–688. [Google Scholar] [CrossRef]
- Coyle, E.F. Physical activity as a metabolic stressor. Am. J. Clin. Nutr. 2000, 72, 512s–520s. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Saltin, B. Exercise as medicine—Evidence for prescribing exercise as therapy in 26 different chronic diseases. Scand. J. Med. Sci. Sports 2015, 25, 1–72. [Google Scholar] [CrossRef] [Green Version]
- Thompson, P.D.; Crouse, S.F.; Goodpaster, B.; Kelley, D.; Moyna, N.; Pescatello, L. The acute versus the chronic response to exercise. Med. Sci. Sports Exerc. 2001, 33, S438–S445. [Google Scholar] [CrossRef]
- Phan, A.T.; Goldrath, A.W. Hypoxia-inducible factors regulate T cell metabolism and function. Mol. Immunol. 2015, 68, 527–535. [Google Scholar] [CrossRef] [Green Version]
- Pucino, V.; Certo, M.; Bulusu, V.; Cucchi, D.; Goldmann, K.; Pontarini, E.; Haas, R.; Smith, J.; Headland, S.E.; Blighe, K.; et al. Lactate Buildup at the Site of Chronic Inflammation Promotes Disease by Inducing CD4(+) T Cell Metabolic Rewiring. Cell Metab. 2019, 30, 1055–1074.e8. [Google Scholar] [CrossRef] [Green Version]
- Thirupathi, A.; Pinho, R.A.; Ugbolue, U.C.; He, Y.; Meng, Y.; Gu, Y. Effect of Running Exercise on Oxidative Stress Biomarkers: A Systematic Review. Front. Physiol. 2020, 11, 610112. [Google Scholar] [CrossRef]
- Nalbandian, M.; Takeda, M. Lactate as a Signaling Molecule That Regulates Exercise-Induced Adaptations. Biology 2016, 5, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, L.A.; Kishton, R.J.; Rathmell, J. A guide to immunometabolism for immunologists. Nat. Rev. Immunol. 2016, 16, 553–565. [Google Scholar] [CrossRef] [Green Version]
- Pearce, E.L.; Poffenberger, M.C.; Chang, C.H.; Jones, R.G. Fueling immunity: Insights into metabolism and lymphocyte function. Science 2013, 342, 1242454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cretenet, G.; Clerc, I.; Matias, M.; Loisel, S.; Craveiro, M.; Oburoglu, L.; Kinet, S.; Mongellaz, C.; Dardalhon, V.; Taylor, N. Cell surface Glut1 levels distinguish human CD4 and CD8 T lymphocyte subsets with distinct effector functions. Sci. Rep. 2016, 6, 24129. [Google Scholar] [CrossRef]
- Jones, N.; Vincent, E.E.; Cronin, J.G.; Panetti, S.; Chambers, M.; Holm, S.R.; Owens, S.E.; Francis, N.J.; Finlay, D.K.; Thornton, C.A. Akt and STAT5 mediate naïve human CD4+ T-cell early metabolic response to TCR stimulation. Nat. Commun. 2019, 10, 2042. [Google Scholar] [CrossRef] [Green Version]
- Renner, K.; Geiselhöringer, A.-L.; Fante, M.; Bruss, C.; Färber, S.; Schönhammer, G.; Peter, K.; Singer, K.; Andreesen, R.; Hoffmann, P.; et al. Metabolic plasticity of human T cells: Preserved cytokine production under glucose deprivation or mitochondrial restriction, but 2-deoxy-glucose affects effector functions. Eur. J. Immunol. 2015, 45, 2504–2516. [Google Scholar] [CrossRef]
- Ricciardi, S.; Manfrini, N.; Alfieri, R.; Calamita, P.; Crosti, M.C.; Gallo, S.; Müller, R.; Pagani, M.; Abrignani, S.; Biffo, S. The Translational Machinery of Human CD4(+) T Cells Is Poised for Activation and Controls the Switch from Quiescence to Metabolic Remodeling. Cell Metab. 2018, 28, 895–906.e5. [Google Scholar] [CrossRef] [Green Version]
- Siedlik, J.A.; Benedict, S.H.; Landes, E.J.; Weir, J.P.; Vardiman, J.P.; Gallagher, P.M. Acute bouts of exercise induce a suppressive effect on lymphocyte proliferation in human subjects: A meta-analysis. Brain Behav. Immun. 2016, 56, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Simpson, R.J.; Campbell, J.P.; Gleeson, M.; Krüger, K.; Nieman, D.C.; Pyne, D.B.; Turner, J.E.; Walsh, N.P. Can exercise affect immune function to increase susceptibility to infection? Exerc. Immunol. Rev. 2020, 26, 8–22. [Google Scholar] [PubMed]
- Siedlik, J.A.; Deckert, J.A.; Benedict, S.H.; Bhatta, A.; Dunbar, A.J.; Vardiman, J.P.; Gallagher, P.M. T cell activation and proliferation following acute exercise in human subjects is altered by storage conditions and mitogen selection. J. Immunol. Methods 2017, 446, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Holbrook, A.K.; Peterson, H.D.; Bianchi, S.A.; Macdonald, B.W.; Bredahl, E.C.; Belshan, M.; Siedlik, J.A. CD4(+) T cell activation and associated susceptibility to HIV-1 infection in vitro increased following acute resistance exercise in human subjects. Physiol. Rep. 2019, 7, e14234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contrepois, K.; Wu, S.; Moneghetti, K.J.; Hornburg, D.; Ahadi, S.; Tsai, M.S.; Metwally, A.A.; Wei, E.; Lee-McMullen, B.; Quijada, J.V.; et al. Molecular Choreography of Acute Exercise. Cell 2020, 181, 1112–1130. [Google Scholar] [CrossRef] [PubMed]
- Renner, K.; Singer, K.; Koehl, G.E.; Geissler, E.K.; Peter, K.; Siska, P.J.; Kreutz, M. Metabolic Hallmarks of Tumor and Immune Cells in the Tumor Microenvironment. Front. Immunol. 2017, 8, 248. [Google Scholar] [CrossRef] [Green Version]
- Perry, C.; Pick, M.; Bdolach, N.; Hazan-Halevi, I.; Kay, S.; Berr, I.; Reches, A.; Harishanu, Y.; Grisaru, D. Endurance exercise diverts the balance between Th17 cells and regulatory T cells. PLoS ONE 2013, 8, e74722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorneles, G.P.; da Silva, I.M.; Santos, M.A.; Elsner, V.R.; Fonseca, S.G.; Peres, A.; Romão, P.R.T. Immunoregulation induced by autologous serum collected after acute exercise in obese men: A randomized cross-over trial. Sci. Rep. 2020, 10, 21735. [Google Scholar] [CrossRef]
- Takenaka, M.C.; Robson, S.; Quintana, F.J. Regulation of the T Cell Response by CD39. Trend. Immunol. 2016, 37, 427–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieman, D.C.; Henson, D.A.; Gross, S.J.; Jenkins, D.P.; Davis, J.M.; Murphy, E.A.; Carmichael, M.D.; Dumke, C.L.; Utter, A.C.; McAnulty, S.R.; et al. Quercetin reduces illness but not immune perturbations after intensive exercise. Med. Sci. Sports Exerc. 2007, 39, 1561–1569. [Google Scholar] [CrossRef] [Green Version]
- Chiappisi, E.; Ringseis, R.; Eder, K.; Gessner, D.K. Effect of endoplasmic reticulum stress on metabolic and stress signaling and kidney-specific functions in Madin-Darby bovine kidney cells. J. Dairy Sci. 2017, 100, 6689–6706. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, Research0034. [Google Scholar] [CrossRef] [Green Version]
- Mousset, C.M.; Hobo, W.; Woestenenk, R.; Preijers, F.; Dolstra, H.; van der Waart, A.B. Comprehensive Phenotyping of T Cells Using Flow Cytometry. Cytometry A 2019, 95, 647–654. [Google Scholar] [CrossRef]
- Sinclair, L.V.; Barthelemy, C.; Cantrell, D.A. Single Cell Glucose Uptake Assays: A Cautionary Tale. Immunometabolism 2020, 2, e200029. [Google Scholar] [CrossRef] [PubMed]
- Al-Ani, A.; Toms, D.; Kondro, D.; Thundathil, J.; Yu, Y.; Ungrin, M. Oxygenation in cell culture: Critical parameters for reproducibility are routinely not reported. PLoS ONE 2018, 13, e0204269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radom-Aizik, S.; Leu, S.Y.; Cooper, D.M.; Zaldivar, F., Jr. Serum from exercising humans suppresses t-cell cytokine production. Cytokine 2007, 40, 75–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skrenta, H.; Yang, Y.; Pestka, S.; Fathman, C.G. Ligand-independent down-regulation of IFN-gamma receptor 1 following TCR engagement. J. Immunol. 2000, 164, 3506–3511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alack, K.; Weiss, A.; Krüger, K.; Höret, M.; Schermuly, R.; Frech, T.; Eggert, M.; Mooren, F.C. Profiling of human lymphocytes reveals a specific network of protein kinases modulated by endurance training status. Sci. Rep. 2020, 10, 888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, R.; Cucchi, D.; Smith, J.; Pucino, V.; Macdougall, C.E.; Mauro, C. Intermediates of Metabolism: From Bystanders to Signalling Molecules. Trends Biochem. Sci. 2016, 41, 460–471. [Google Scholar] [CrossRef]
- Hall, M.M.; Rajasekaran, S.; Thomsen, T.W.; Peterson, A.R. Lactate: Friend or Foe. PM R 2016, 8, S8–S15. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Steensberg, A.; Fischer, C.; Keller, C.; Keller, P.; Plomgaard, P.; Wolsk-Petersen, E.; Febbraio, M. The metabolic role of IL-6 produced during exercise: Is IL-6 an exercise factor? Proc. Nutr. Soc. 2004, 63, 263–267. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (from 5′ to 3′) Reverse Primer (from 5′ to 3′) | PCR Product Size (bp) | NCBI GenBank Accession Number | Primer Efficiency |
|---|---|---|---|---|
| Reference genes | ||||
| ATP5B | TCGCGTGCCATTGCTGAGCT CGTGCACGGGACACGGTCAA | 218 | NM_001686 | 1.72 |
| SDHA | CCAAGCCCATCCAGGGGCAAC TCCAGAGTGACCTTCCCAGTGCCAA | 100 | NM_004168 | 1.91 |
| YWHAZ | TGGGGACTACGACGTCCCTCAA CATATCGCTCAGCCTGCTCGG | 115 | NM_003406 | 2.01 |
| Target genes | ||||
| ATP5J | TCGGGACTGAGTGCAAGAAT AACACCAATGTTCCTCCGCA | 105 | NM_001003703 | 1.89 |
| COX4I1 | GTGGCGGCAGAATGTTGG GGCAGACAGGTGCTTGACAT | 182 | NM_001861 | 2.05 |
| FOXP3 | CACTTACAGGCACTCCTCCA CAGCCGAAAGGGTGCTGTC | 286 | NM_014009 | 1.78 |
| HK1 | GCCTATTACTTCACGGAGCTGA TGAAATCTCCCTTTTCAGAGCCA | 220 | NM_000188 | 1.95 |
| IDH1 | TACTTCAGAAGCGGAGGCAC GTAGTTTATCGCCTGCCGGG | 129 | NM_005896 | 2.04 |
| LDHA | ACGTGCATTCCCGATTCCTT GGAAAAGGCTGCCATGTTGG | 251 | NM_005566 | 2.07 |
| NDUFA12 | TAGTGCAGGTCCTGAAACGC CCATCGGTGACGGCCAAAAA | 176 | NM_018838 | 2.12 |
| ODGH | CTGTGCTTGGCTGGAAAACC CATGGTGCCCTCGTATCTGA | 245 | NM_002541 | 2.36 |
| PRKAA1 | CGGCAAAGTGAAGGTTGGC CTGTTGGAACAGACGCCGA | 286 | NM_006251 | 1.98 |
| RPS6KB1 | CTGAGGACATGGCAGGAGTG ACAATGTTCCATGCCAAGTTCA | 146 | NM_003161 | 1.93 |
| Antibody | Fluorochrome | Vendor/Cat. No./Clone | Isotype |
|---|---|---|---|
| CD95 | PE | Biolegend/305608/DX2 | Mouse IgG1, κ |
| CD45RA | APC | Biolegend/304111/HI100 | Mouse IgG2b, κ |
| CD197/CCR7 | BV421 | Biolegend/353207/G043H7 | Mouse IgG2a, κ |
| CD4 | Alexa Fluor 700 | Biolegend/344621/SK3 | Mouse IgG1, κ |
| Live/Dead | Zombie Aqua | Biolegend/423101/- | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palmowski, J.; Gebhardt, K.; Reichel, T.; Frech, T.; Ringseis, R.; Eder, K.; Renner-Sattler, K.; Krüger, K. The Impact of Exercise Serum on Selected Parameters of CD4+ T Cell Metabolism. Immuno 2021, 1, 119-131. https://0-doi-org.brum.beds.ac.uk/10.3390/immuno1030008
Palmowski J, Gebhardt K, Reichel T, Frech T, Ringseis R, Eder K, Renner-Sattler K, Krüger K. The Impact of Exercise Serum on Selected Parameters of CD4+ T Cell Metabolism. Immuno. 2021; 1(3):119-131. https://0-doi-org.brum.beds.ac.uk/10.3390/immuno1030008
Chicago/Turabian StylePalmowski, Jana, Kristina Gebhardt, Thomas Reichel, Torsten Frech, Robert Ringseis, Klaus Eder, Kathrin Renner-Sattler, and Karsten Krüger. 2021. "The Impact of Exercise Serum on Selected Parameters of CD4+ T Cell Metabolism" Immuno 1, no. 3: 119-131. https://0-doi-org.brum.beds.ac.uk/10.3390/immuno1030008