Frequency of Occurrence and Ingested Biomass of Different Prey of the Barn Owl Tyto alba in an Island Ecosystem





Abstract
:Simple Summary
Abstract
1. Introduction
2. Experimental Section
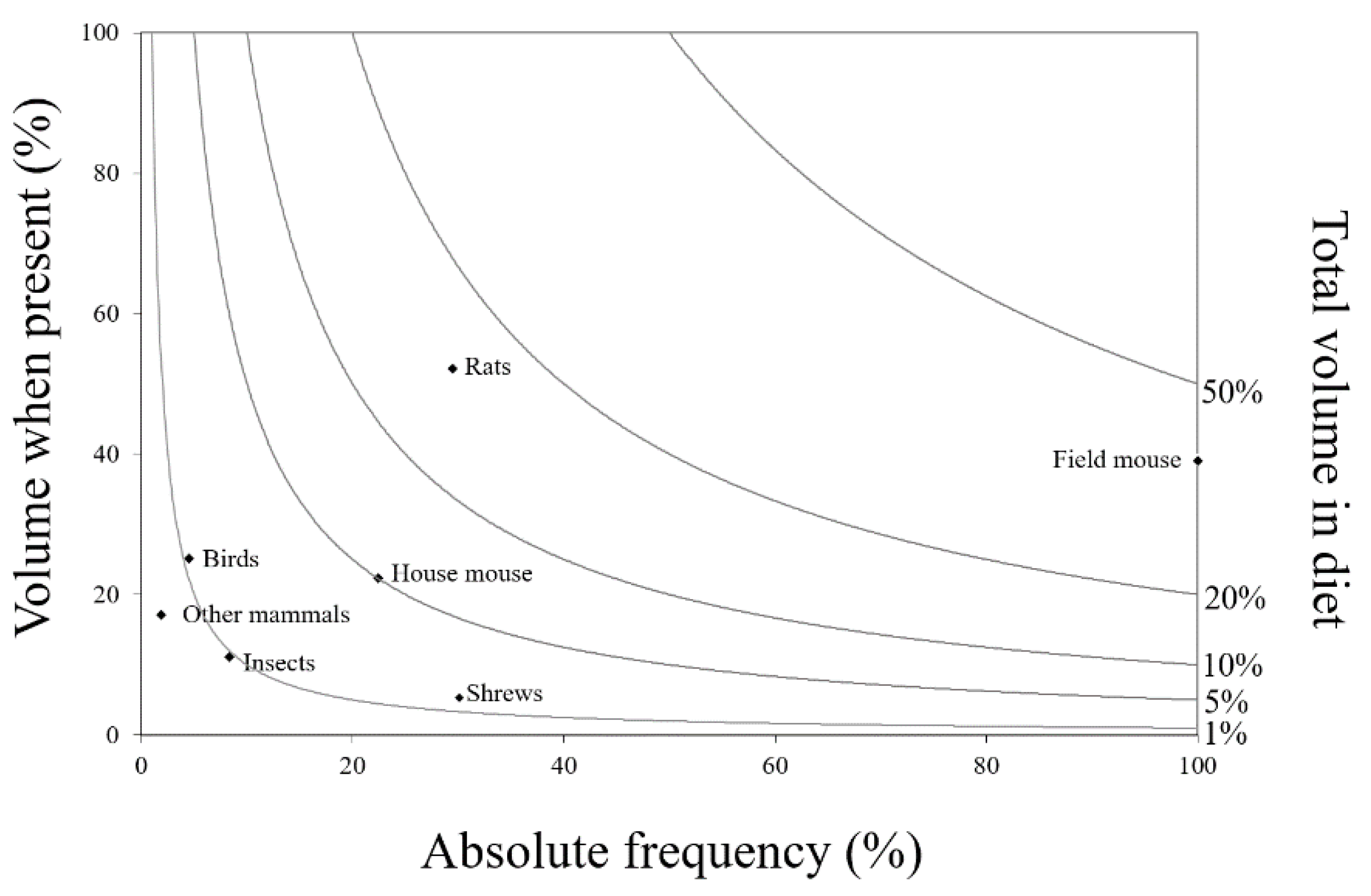
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bai, M.L.; Schmidt, D.; Gottschalk, E.; Mühlenberg, M. Distribution pattern of an expanding Osprey (Pandion haliaetus) population in a changing environment. J. Ornithol. 2009, 150, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Bozinovic, F.; Calosi, P.; Spicer, J.I. Physiological correlates of geographic range in animals. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 155–179. [Google Scholar] [CrossRef] [Green Version]
- Hilderbrand, G.V.; Gustine, D.D.; Mangipane, B.; Joly, K.; Leacock, W.; Mangipane, L.; Erlenbach, J.; Sorum, M.S.; Cameron, M.D.; Belant, J.L.; et al. Plasticity in physiological condition of female brown bears across diverse ecosystems. Polar Biol. 2018, 41, 773–780. [Google Scholar] [CrossRef]
- Podgórski, T.; Baś, G.; Jędrzejewska, B.; Sönnichsen, L.; Śnieżko, S.; Jędrzejewski, W.; Okarma, H. Spatiotemporal behavioral plasticity of wild boar (Sus scrofa) under contrasting conditions of human pressure: Primeval forest and metropolitan area. J. Mammal. 2013, 94, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Newsome, T.M.; Boitani, L.; Chapron, G.; Ciucci, P.; Dickman, C.R.; Dellinger, J.A.; Lopez-Bao, J.V.; Peterson, R.O.; Shores, C.R.; Wirsing, A.J.; et al. Food habits of the world’s grey wolves. Mammal. Rev. 2016, 46, 255–269. [Google Scholar] [CrossRef]
- Soe, E.; Davison, J.; Süld, K.; Valdmann, H.; Laurimaa, L.; Saarma, U. Europe-wide biogeographical patterns in the diet of an ecologically and epidemiologically important mesopredator, the red fox Vulpes vulpes: A quantitative review. Mammal. Rev. 2017, 47, 198–211. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Monographs in Population Biology; Princeton University Press: Princeton, NJ, USA, 1967; Volume 1. [Google Scholar]
- Jiménez, J.E.; Marquet, P.A.; Medel, R.G.; Jaksic, F.M. Comparative ecology of Darwin’s fox (Pseudalopex fulvipes) in mainland and island settings of southern Chile. Rev. Chil. Hist. Nat. 1991, 63, 177–186. [Google Scholar]
- Clevenger, A.P. Pine marten (Martes martes Linné, 1758) comparative feeding ecology in an island and mainland population of Spain. Zeitschr. Saugetierk. 1993, 58, 212–224. [Google Scholar]
- Luiselli, L.; Filippi, E.; Capula, M. Geographic variation in diet composition of the grass snake (Natrix natrix) along the mainland and an island of Italy: The effects of habitat type and interference with potential competitors. Herp. J. 2005, 15, 221–230. [Google Scholar]
- Stronen, A.V.; Navid, E.L.; Quinn, M.S.; Paquet, P.C.; Bryan, H.M.; Darimont, C.T. Population genetic structure of gray wolves (Canis lupus) in a marine archipelago suggests island-mainland differentiation consistent with dietary niche. BMC Ecol. 2014, 14, 11. [Google Scholar] [CrossRef] [Green Version]
- BirdLife International. Tyto alba (Amended Version of 2016 Assessment). The IUCN Red List of Threatened Species 2019. 2019. Available online: https://www.iucnredlist.org/species/22688504/155542941 (accessed on 26 October 2020).
- Romano, A.; Séchaud, R.; Roulin, A. Global biogeographical patterns in the diet of a cosmopolitan avian predator. J. Biogeogr. 2020, 47, 1467–1481. [Google Scholar] [CrossRef]
- Love, R.A.; Webon, C.; Glue, D.E.; Harris, S.; Harris, S. Changes in the food of British Barn Owls (Tyto alba) between 1974 and 1997. Mammal. Rev. 2000, 30, 107–129. [Google Scholar] [CrossRef]
- Milana, G.; Lai, M.; Maiorano, L.; Luiselli, L.; Amori, G. Geographic patterns of predator niche breadth and prey species richness. Ecol. Res. 2016, 31, 111–115. [Google Scholar] [CrossRef]
- Milana, G.; Luiselli, L.; Amori, G. Forty years of dietary studies on Barn Owl (Tyto alba) reveal long term trends in diversity metrics of small mammal prey. Anim. Biol. 2018, 68, 129–146. [Google Scholar] [CrossRef]
- Jaksic, F.M.; Seib, R.L.; Herrera, C.M. Predation by the Barn Owl (Tyto alba) in Mediterranean habitats of Chile, Spain and California: A comparative approach. Am. Midl. Nat. 1982, 107, 151–162. [Google Scholar] [CrossRef]
- Bellocq, M.I. A review of the trophic ecology of the Barn Owl in Argentina. J. Raptor Res. 2000, 34, 108–119. [Google Scholar]
- Cagnacci, F.; Cardini, A.; Ciucci, P.; Ferrari, N.; Mortelliti, A.; Preatoni, D.G.; Russo, D.; Scandura, M.; Wauters, L.A.; Amori, G. Less is more: Researcher survival guide in times of economic crisis. Hystrix 2012, 23, 1–7. [Google Scholar]
- Roulin, A.; Dubey, S. The occurrence of reptiles in Barn Owl diet in Europe. Bird Study 2012, 59, 504–508. [Google Scholar] [CrossRef] [Green Version]
- Teta, P.; Hercolini, C.; Cueto, G. Variation in the diet of Western Barn Owls (Tyto alba) along an urban-rural gradient. Wilson J. Ornit. 2012, 124, 589–596. [Google Scholar] [CrossRef]
- Roulin, A.; Christe, P. Geographic and temporal variation in the consumption of bats by European Barn Owls. Bird Study 2013, 60, 561–569. [Google Scholar] [CrossRef] [Green Version]
- Roulin, A. Shrews and moles are less often captured by European Barn Owls Tyto alba nowadays than 150 years ago. Bird Study 2016, 63, 559–563. [Google Scholar] [CrossRef]
- Roulin, A. Strong decline in the consumption of invertebrates by Barn Owls from 1860 to 2012 in Europe. Bird Study 2016, 63, 146–147. [Google Scholar] [CrossRef] [Green Version]
- Amori, G.; Rizzo Pinna, V.; Sammuri, G.; Luiselli, L. Diversity of small mammal communities of the Tuscan Archipelago: Testing the effects of island size, distance from mainland and human density. Folia Zool. 2015, 64, 161–166. [Google Scholar] [CrossRef]
- Guerra, C.; García, D.; Alcover, J.A. Unusual foraging patterns of the Barn Owl, Tyto alba (Strigiformes: Tytonidae), on small islets from the Pityusic archipelago (Western Mediterranean Sea). Folia Zool. 2014, 63, 180–187. [Google Scholar] [CrossRef]
- Janžekovič, F.; Klenovšek, T. The biogeography of diet diversity of Barn Owls on Mediterranean islands. J. Biogeogr. 2020, 47, 2353–2361. [Google Scholar] [CrossRef]
- Johnston, D.W.; Hill, J.M. Prey selection of common Barn Owls on islands and mainland sites. J. Raptor Res. 1987, 21, 3–7. [Google Scholar]
- Szpunar, G.; Aloise, G.; Filippucci, M.G. Suncus etruscus (Soricomorpha, Soricidae): A new species for Elba island (Tuscan Archipelago, Italy). Ital. J. Zool. 2008, 75, 445–447. [Google Scholar] [CrossRef] [Green Version]
- Contoli, L.; Aloise, G.; Filippucci, M.G. Sulla nicchia trofica di Tyto alba ed Athene noctua nell’isola d’Elba (Toscana) in rapporto all’Italia tirrenica. Bull. Ecol. 1988, 19, 367–373. [Google Scholar]
- Martínez, J.A.; Zuberogoitia, I. Habitat preferences and causes of population decline for Barn Owls Tyto alba: A multi-scale approach. Ardeola 2004, 51, 303–317. [Google Scholar]
- Roulin, A. Spatial variation in the decline of European birds as shown by the Barn Owl Tyto alba diet. Bird Study 2015, 62, 271–275. [Google Scholar] [CrossRef] [Green Version]
- Cassola, F.; Lovari, S. Food habits of rollers during the nesting season. Boll. Zool. 1979, 46, 87–89. [Google Scholar] [CrossRef] [Green Version]
- Nappi, A. L’analisi delle borre degli uccelli: Metodiche, applicazioni e informazioni. Un lavoro monografico. Picus 2011, 37, 106–120. [Google Scholar]
- Nappi, A. I Micromammiferi d’Italia; Simoni, Ed.; Simoni: Naples, Italy, 2001. [Google Scholar]
- Gaggi, A.; Paci, A.M. Atlante degli Erinaceomorfi dei Soricomorfi e dei Piccoli Roditori dell’Umbria; Dimensione Grafica Snc.: Spello (Perugia), Italy, 2014. [Google Scholar]
- Di Carlo, E.A. Avifauna delle Isole dell’Arcipelago Toscano. Le forme ornitiche insulari. Biogeographia 1976, 5, 845–878. [Google Scholar] [CrossRef] [Green Version]
- Vaccaro, A.; Turrisi, G.F. Ritrovamento di Zamenis longissimus (Laurenti, 1768) (Reptilia, Colubridae) sull’Isola d’Elba (Toscana, Italia). Acta Herpetol. 2007, 2, 59–63. [Google Scholar]
- Amori, G.; Contoli, L.; Nappi, A. Mammalia II: Erinaceomorpha, Soricomorpha, Lagomorpha, Rodentia Il Sole 24 Ore; Edagricole Calderini: Bologna, Italy, 2008. [Google Scholar]
- Mori, E.; Iacucci, A.; Castiglia, R.; Santini, L. Sexual-size dimorphism in two synanthropic rat species: Comparison and eco-evolutionary perspectives. Mammal. Biol. 2017, 83, 78–80. [Google Scholar] [CrossRef]
- Mori, E.; Brambilla, M.; Ramazzotti, F.; Ancillotto, L.; Mazza, G.; Russo, D.; Amori, G.; Galimberti, A. In or out of the checklist? DNA barcoding and distribution modelling unveil a new species of Crocidura shrew for Italy. Animals 2020, 12, 380. [Google Scholar] [CrossRef]
- Hutson, A.M.; Spitzenberger, F.; Aulagnier, S.; Nagy, Z. Myotis emarginatus. IUCN RedList Data. 2008. Available online: https://www.iucnredlist.org/species/14129/22051191 (accessed on 3 November 2020).
- Levins, R. Evolution in Changing Environments; Princeton University Press: Princeton, NJ, USA, 1968. [Google Scholar]
- Kruuk, H. The Social Badger: Ecology and Behaviour of Group Living Carnivore (Meles meles); Oxford University Press: Oxford, UK, 1989. [Google Scholar]
- Glen, A.S.; Dickman, C.R. Diet of the spotted-tailed quoll (Dasyurus maculatus) in Eastern Australia: Effects of season, sex and size. J. Zool. 2006, 269, 241–248. [Google Scholar] [CrossRef]
- Taylor, I.R. Barn Owls: Predator-Prey Relationships and Conservation; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar]
- Mori, E.; Ferretti, F.; Fattorini, N. Alien war: Ectoparasite load, diet and temporal niche partitioning in a multi-species assembly of small rodents. Biol. Invasions 2019, 21, 3305–3318. [Google Scholar] [CrossRef]
- Saufi, S.; Ravindran, S.; Hamid, N.H.; Abidin, C.M.R.Z.; Ahmad, H.; Ahmad, A.H.; Salim, H. Diet composition of introduced Barn Owls (Tyto alba javanica) in urban area in comparison with agricultural settings. J. Urban Ecol. 2020, 1, 1–8. [Google Scholar]
- Ruprecht, A.L. Bats (Chiroptera) as constituents of the food of Barn Owls Tyto alba in Poland. Ibis 1979, 121, 489–494. [Google Scholar] [CrossRef]
- Lewis, S.E. Low roost-site fidelity in pallid bats: Associated factors and effect on group stability. Behav. Ecol. Sociobiol. 1996, 39, 335–344. [Google Scholar] [CrossRef]
- Lesiński, G.; Kasprzyk, K.; Gryz, J. Bats taken by the tawny owl in relation to its roosting site. North-West J. Zool. 2012, 8, 247–251. [Google Scholar]
- Kahmann, H.; Goerner, P. Les chiroptères de Corse. Mammalia 1956, 20, 333–389. [Google Scholar] [CrossRef]
- Vergari, S.; Dondini, G. La Chirotterofauna dell’Arcipelago Toscano; WWF Toscana, Ed.; WWF Toscana: Roma, Italy, 1998. [Google Scholar]
- Fattorini, N.; Burrini, L.; Morao, G.; Ferretti, F.; Romeo, G.; Mori, E. Splitting hairs: How to tell hair of hares apart for predator diet studies. Mammal. Biol. 2018, 85, 84–89. [Google Scholar] [CrossRef]
- Obuch, J.; Khaleghizadeh, A. Spatial variation in the diet of the Barn Owl Tyto alba in Iran. Podoces 2011, 6, 103–116. [Google Scholar]


{kind=link}
| Species | Body Mass (Grams, Mean ± SD) | Reference |
|---|---|---|
| Apodemus sylvaticus | 28 ± 6 | [39] |
| Mus domesticus | 25 ± 9 | [39] |
| Rattus rattus | 195 ± 21 | [40] |
| Rattus norvegicus | 364 ± 62 | [40] |
| Crocidura suaveolens | 10 ± 6 | [41] |
| Myotis emarginatus | 12 ± 3 | [42] |
| Suncus etruscus | 3 ± 1 | [39] |
| Lepus sp.-newborns | 110 ± 13 | [39] |
| Passer italiae | 22 ± 4 | EM, unpublished data |
| Decticus albifrons | 2 ± 1 | University of Turin, unpublished data |
| Species | Min N° Ind. | RF (%) | AF (%) | Biomass (%) |
|---|---|---|---|---|
| Apodemus sylvaticus | 248 | 62.16 | 100 | 39.06 |
| Rattus rattus | 45 | 11.28 | 27.95 | 50.12 |
| Rattus norvegicus | 1 | 0.25 | 0.62 | 2.05 |
| Mus domesticus | 35 | 8.77 | 21.74 | 4.92 |
| Suncus etruscus | 28 | 7.02 | 17.39 | 0.47 |
| Crocidura suaveolens | 19 | 4.76 | 11.80 | 1.07 |
| Lepus sp. | 2 | 0.50 | 1.24 | 1.24 |
| Myotis emarginatus | 1 | 0.25 | 0.62 | 0.07 |
| Passer italiae | 5 | 1.25 | 3.11 | 0.62 |
| Sturnus vulgaris | 1 | 0.25 | 0.62 | 0.45 |
| Sylviidae | 1 | 0.25 | 0.62 | 0.08 |
| Decticus albifrons | 10 | 2.51 | 6.21 | 0.10 |
| Scarabaeidae | 1 | 0.25 | 0.62 | 0.01 |
| Melolonthidae | 1 | 0.25 | 0.62 | 0.01 |
| Forficula auricularia | 1 | 0.25 | 0.62 | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viganò, M.; Ancillotto, L.; Agnelli, P.; Ficetola, G.F.; Mori, E. Frequency of Occurrence and Ingested Biomass of Different Prey of the Barn Owl Tyto alba in an Island Ecosystem. Birds 2020, 1, 5-12. https://0-doi-org.brum.beds.ac.uk/10.3390/birds1010002
Viganò M, Ancillotto L, Agnelli P, Ficetola GF, Mori E. Frequency of Occurrence and Ingested Biomass of Different Prey of the Barn Owl Tyto alba in an Island Ecosystem. Birds. 2020; 1(1):5-12. https://0-doi-org.brum.beds.ac.uk/10.3390/birds1010002
Chicago/Turabian StyleViganò, Miriam, Leonardo Ancillotto, Paolo Agnelli, Gentile Francesco Ficetola, and Emiliano Mori. 2020. "Frequency of Occurrence and Ingested Biomass of Different Prey of the Barn Owl Tyto alba in an Island Ecosystem" Birds 1, no. 1: 5-12. https://0-doi-org.brum.beds.ac.uk/10.3390/birds1010002