
Associative Genetic Diversity of Sarcocystis levinei Isolates across the Globe

1
Department of Parasitology, U. P. Pandit Deen Dayal Upadhyaya Pashu Chikitsa Vigyan Vishwavidyalaya Evam Go Anusandhan Sansthan (DUVASU), Mathura 281001, Uttar Pradesh, India
2
Department of Veterinary Parasitology, College of Veterinary Science (CoVS), Guru Angad Dev Veterinary and Animal Sciences University (GADVASU), Rampura Phul 151103, Punjab, India
3
College of Biotechnology, U. P. Pandit Deen Dayal Upadhyaya Pashu Chikitsa Vigyan Vishwavidyalaya Evam Go Anusandhan Sansthan (DUVASU), Mathura 281001, Uttar Pradesh, India
4
Department of Veterinary Pathology, U. P. Pandit Deen Dayal Upadhyaya Pashu Chikitsa Vigyan Vishwavidyalaya Evam Go Anusandhan Sansthan (DUVASU), Mathura 281001, Uttar Pradesh, India
*
Author to whom correspondence should be addressed.
Parasitologia 2023, 3(3), 231-240; https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3030024
Submission received: 9 February 2023
/
Revised: 21 June 2023
/
Accepted: 29 June 2023
/
Published: 4 July 2023
(This article belongs to the Special Issue Sarcocystis in Domestic and Wildlife Animals)
Abstract
:Bubaline sarcocystosis, caused by Sarcocystis levinei, is worldwide in distribution. Yet, very limited reports are available across the globe on the phylogenetic aspects of this particular Sarcocystis spp. Virtually no literature is available on its molecular aspects from India. Thus, the present study was designed to phylogenetically characterize the various isolates of S. levinei based on 18S rRNA and cox 1 mitochondrial genes. A total of 11 isolates of S. levinei from Northern India were characterized for 18S rRNA (MG957189-MG957199) and cox 1 (MH255771-MH255781) genes. PCR products were cloned, sequenced, and compared with other sequences across the world. A phylogenetic tree was constructed based on the maximum likelihood (ML) method. Phylogenetic analysis placed S. levinei and S. cruzi into two monophyletic sister groups. An appreciable amount of genetic variability was noticed between various S. levinei sequences for both the gene loci. A total of three haplotypes were noticed for each gene. The generated sequences showed 99.8–100.0% and 99.7–100.0% nucleotide homologies within themselves with regard to the 18S rRNA and cox 1 genes, respectively. Few of the studied sequences showed marked similarity and closeness with the Egyptian sequences than their Indian counterparts. This is the first report of molecular characterization and sequence phylogenetic study of S. levinei from India.
1. Introduction
Bubaline sarcocystosis is caused by four species of Sarcocystis genus viz., S. fusiformis, and S. buffalonis (forming macroscopic sarcocysts), alongside S. levinei and S. sinensis/S. dubeyi (forming microscopic sarcocysts) [1,2]. There is no valid record of human illness from consumption of Sarcocystis spp.-infected buffalo meat; however, reports of human illness in two Chinese volunteers after ingesting Sarcocystis-infected meat suggest possible public heath significance [1]. Ample work has been reported across the world on characterization studies of Sarcocystis spp. affecting different hosts [3,4,5,6]. The nuclear (18S ribosomal RNA, 28S ribosomal RNA and Internal Transcribed Spacer-ITS) and mitochondrial cytochrome (cox 1) genes have been used for the genetic characterization of various Sarcocystis spp. [7,8,9,10].
Sarcocystis levinei is widely reported in the cardiac muscles of buffaloes across the globe [2,11,12]. The parasite has a high prevalence rate of 75% in China, 83% in Iran, 73% in Italy, 74% in Vietnam, and 68% in India [2,12]. But, in spite of its global distribution, very limited work has been conducted on the phylogenetic characterization aspects of S. levinei [5]. Even that limited work indicates appreciable genetic variability within its isolates. Although there are ample reports of the distribution of S. levinei across India [12,13,14,15], there is no record of phylogenetic characterization of S. levinei from India. Of late, the parasite has been thoroughly redescribed [16]. The present study was designed to characterize the Indian isolates of S. levinei based on 18SrRNA and cox 1 genes. In addition, the evolutionary phylogeny of S. levinei, with regard to other Sarcocystis spp. affecting different hosts, was also delineated. Subsequently, the phylogenetic analysis was carried out to determine the placement of Indian isolates in terms of the available S. levinei sequences.
2. Results
The sarcocysts were identified as those of S. levinei (Figure 1) based on the previously described morphological features [2,16]. The sarcocysts were thin-walled and elongated in shape. A total of 11 sarcocysts were characterized for 18S and cox 1 genes, resulting into 22 sequences (11 apiece). Upon submission to
NCBI, accession numbers MG957189-MG957199 were obtained for the 18S gene, while the accession numbers for cox 1 were MH255771-MH255781.
2.1. Phylogenetic Analysis of 18S rRNA and Cox 1 Gene Sequences of Various Sarcocystis spp.
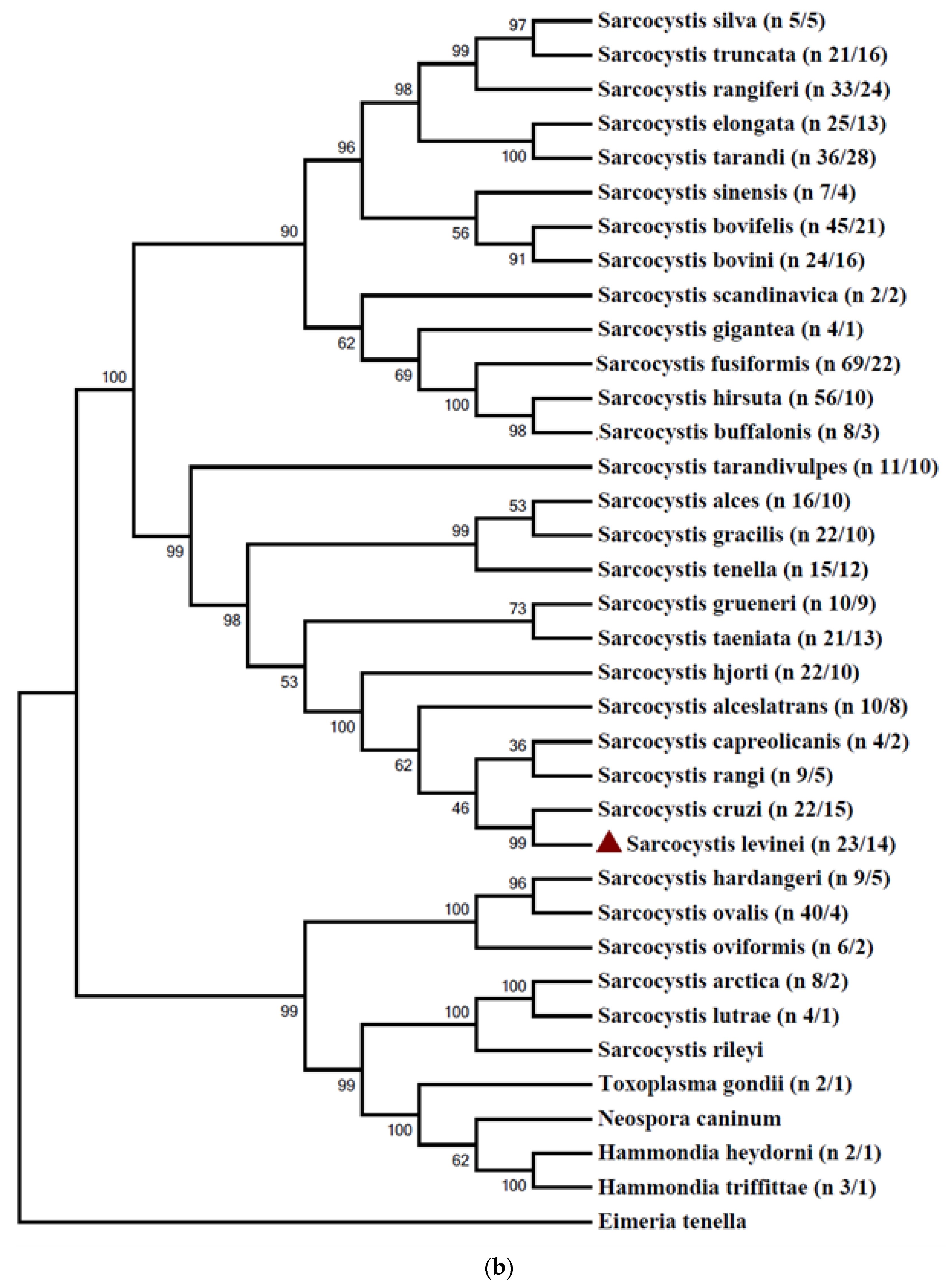
A total of 103 sequences belonging to 50 taxa of Sarcocystis spp. were used for 18S rRNA gene analysis, including 11 sequences of S. levinei generated from the present study. Since the main objective of our analysis was to compare the phylogenetic placement of various Sarocystis spp. based on the 18S rRNA gene, all sequences/haplotypes of a species were grouped under the name of the respective species. Hence, a total of 45 sequences representing 45 Sarocystis species and 7 sequences of other closely related parasitic species were used. Sequences of 18S rRNA genes were trimmed at both ends so that most sequences started and ended at the homologous nucleotide position, corresponding to positions 161 and 985, respectively, of the S. bovifelis sequence. The gene bank sequence of E. tenella was used as an out-group species to root the tree for the 18S rRNA gene (Figure 2a).
Likewise, 570 sequences originating from 37 taxa of Sarcocystis spp. were used in the analysis of the cox 1 gene, including 11sequences of the S. levinei generated from the present study. However, in order to reduce the computation time, identical superfluous sequences were removed so that each haplotype was only represented by a single sequence. Hence, for the final analysis, a total of 300 sequences (haplotypes) were used. Sub-trees formed by two or more sequences/haplotypes of the same species were collapsed, but the quantities of included sequences/haplotypes are given behind the taxon names. The sequences were trimmed at both ends so that most sequences started and ended at the same (homologous) nucleotide positions, corresponding to positions 13 and 1016, respectively, of gene sequence KC209690 of S. bovifelis. GeneBank sequence HQ702484 of E. tenella was used as an out-group species to root the tree for the cox 1 gene (Figure 2b).
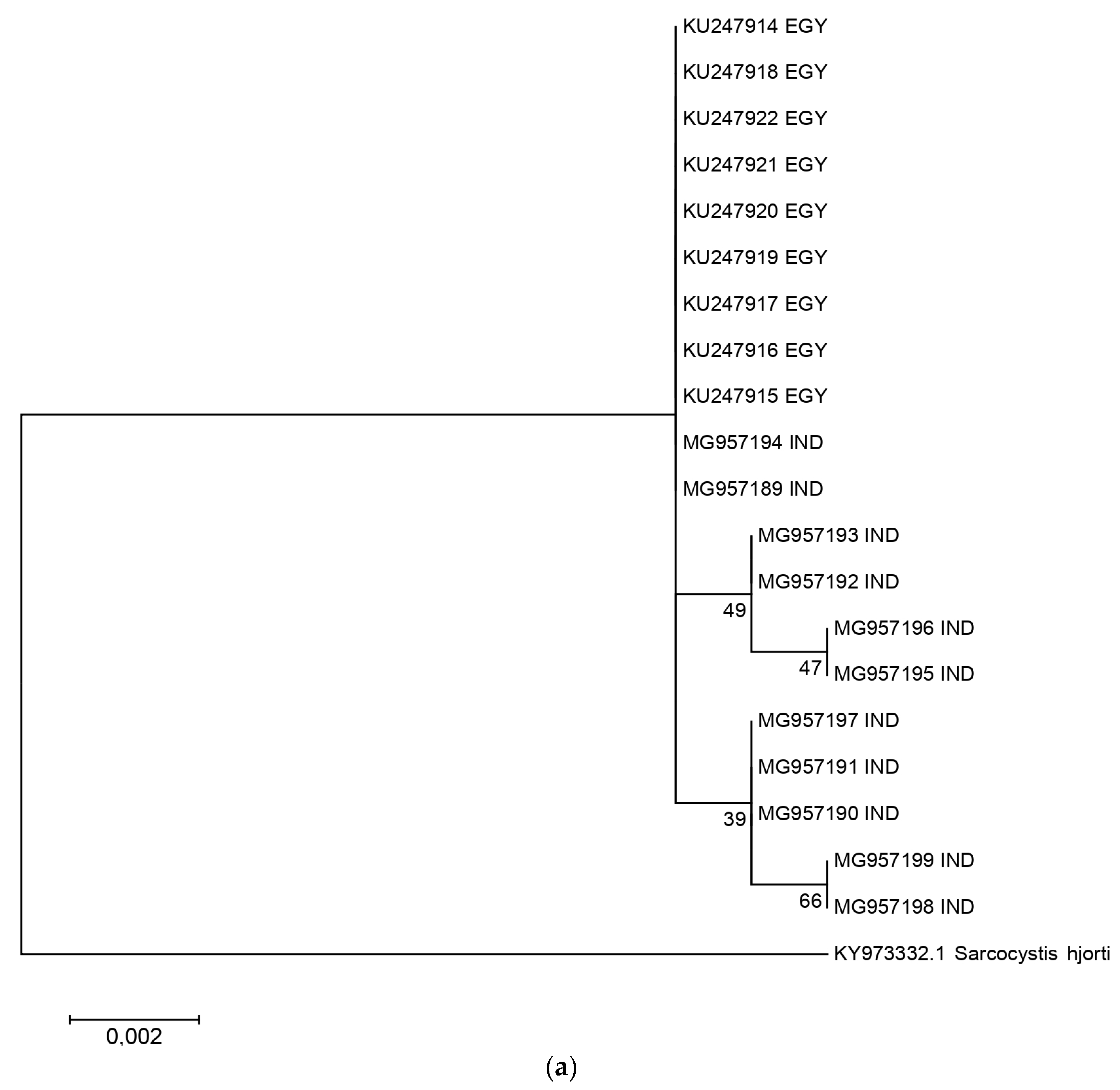
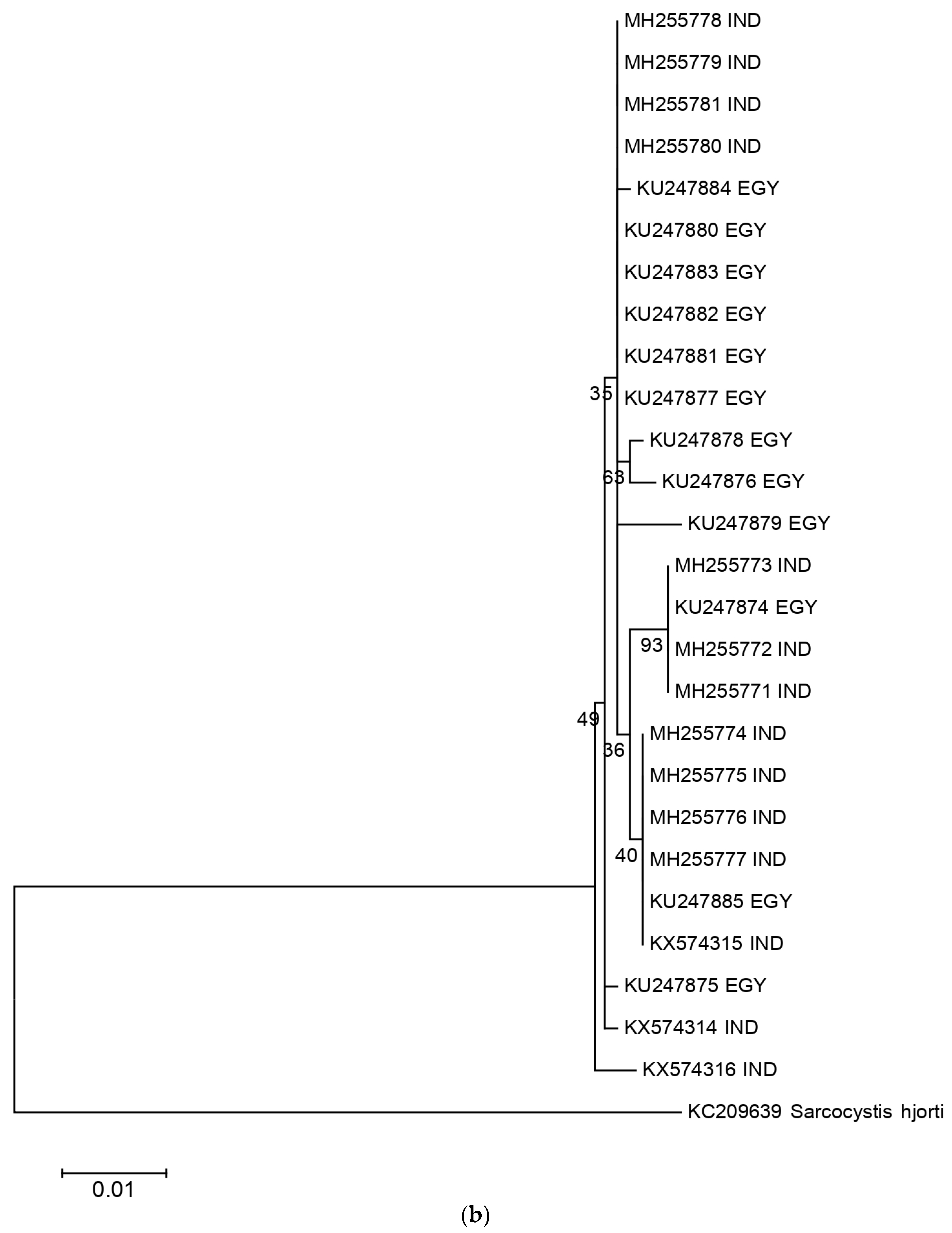
2.2. Phylogenetic Analysis and Nucleotide Identity of 18S rRNA and Cox 1 Gene Sequences of the Present Isolates with Other Isolates of S. levinei
A total of 11 sarcocysts of S. levinei were characterized for the 18S ribosomal gene. These sequences were aligned with other sequences of S. levinei available in NCBI, and a phylogenetic tree was drawn using Mega 6 software (Figure 3a). For 18S rRNA, the length of multiple alignment was 876 bp, and the Tamura 3 parameter was chosen as the best nucleotide evolution model. For cox 1, the length of multiple alignment was 822 bp and the Kimura 2 parameter was chosen as the best nucleotide evolution model. S. hjorti was used as the outgroup for both analyses. Only the sequences of S. levinei from Egypt were available in NCBI for the comparison and construction of the phylogenetic tree. Likewise, the nucleotide identity of the present sequences with other sequences of S. levinei available from India and Egypt was also calculated. A total of five haplotypes were noticed within the studied S. levinei sequences. The presented Indian sequences showed 99.7–100.0% genetic identity within themselves. On the other hand, Egyptian isolates of S. levinei (KU247914-KU247922) comprised only a single haplotype, which was identical to two isolates of S. levinei obtained in the present study (MG957189, MG957194). Eleven cox 1 sequences of S. levinei generated in the present study were compared with those of S. levinei from Egypt. Overall, 10 haplotypes of S. levinei were identified with the cox 1 gene. The presented Indian isolates of the S. levinei species numbered 99. A sequence identity of 5–100% was found, whereas, when comparing all available isolates of this Sarcocystis species, 98.8–100% sequence identity was calculated. Details of the haplotype number, haplotype diversity, nucleotide diversity, number of segregating sites, and average number of nucleotide differences are represented in tabular form in Table 1.
3. Discussion
Contrary to the general belief that Sarcocystis spp. are highly specific for intermediate hosts, a few 18S rRNA gene sequences suggest a considerably high nucleotide identity between Sarcocystis spp. affecting cattle with those affecting water buffaloes, and between the species affecting water buffaloes with those affecting African buffaloes [9]. The extensive sequence variation at the 5-end of 18S gene rRNA [17] often leads to interpretations that the morphologically similar Sarcocystis spp. from cattle and buffalo can be considered to be the same species [18] which is basically false [5,8,17,19]. Besides cattle and buffaloes, this appreciable amount of intraspecific sequence variations at 5-end of the 18S gene has been reported in many Sarcocystis spp. affecting a wide range of hosts [6,20,21]. Furthermore, Sarcocystis spp.’s heterogeneity in 18S is also seen in apicomplexan parasites—Plasmodium, Crytosporidium, Babesia, Theileria, and Eimeria [22,23,24]. It is much easier to study 18S-linked intra-isolate variability in Sarcocystis spp., owing to the availability of fairly large amount of DNA from a single sarcocyst compared to these apicomplexan parasites [17]; hence, multiple genes can be easily explored from a single source. This intra-isolate heterogeneity involves indels that necessitate the cloning of PCR products before sequencing, as it is very confusing to assign the various sequence variants to a particular species unambiguously until it is confirmed that all of them have originated from a single sarcocyst [17]. Hence, many ambiguities have been reported by researchers owing to the direct sequencing of PCR products involving Sarcocystis spp. [4,25,26].
A phylogenetic analysis involving different Sarcocystis spp. placed S. levinei and S. cruzi into a monophyletic group. Certain sequences of S. levinei could not be distinguished from those of S. cruzi, either at the 18S or 28S rRNA genes [5]. However, the cox 1 gene was found to be superior to the 18S and 28S rRNA genes with regard to the ability to unambiguously delimit the species [5]. Gjerde et al. [5] reported nucleotide identity cox 1 sequences of S. levinei and S. cruzi to be around 92.9–94.0 %. cox 1 is considered to be superior to the 18S rRNA gene for its ability to separate closely related (recently diverged) and morphologically indistinguishable Sarcocystis spp. in ruminants.
The government legislations on slaughter, owing to religious sentiments of the general public in India, limit the number of licensed slaughter houses for large ruminants. Hence, it is not possible to obtain a large amount of material to be collected and transferred to faraway areas for research proposes. Hence, the heart was chosen as the material of choice for the collection of specimens, as S. levinei is the only species reported in the cardiac tissue of buffaloes from India [12]. A total of 20 S. levinei sequences, including 11 (MG957189-MG957199) generated in the present study, were compared based on the 18S rRNA gene locus. Amongst the present Indian isolates, three haplotypes were noticed. The present Indian isolates showed 99.7–100.0% nucleotide identity, both within themselves and with other isolates available from Egypt. The first molecular characterization of S. levinei-like sarcocysts from water buffaloes was performed by Yang et al. [18], and it was confused, at that time, with S. cruzi of cattle due to the much higher identity based on the 18S gene. However, Gjerde et al. [5] clarified these differences by comparing multiple genes of both these species. A substantial amount of intra- and/or inter-isolate sequence variation in the 18S rRNA gene has been recorded in several Sarcocystis spp. specimens. This is often attributed to such variations and subsequently leads to confusions about the species involved. Thus, the 18S gene alone cannot be used for the genetic characterization of Sarcocystis spp. [5,16].
The cox 1 gene is a promising genetic marker for the differentiation of various Sarcocystis spp. Species affecting ruminants [8,11]. This gene seems to be more suitable than the nuclear 18S rRNA gene for the separation of closely related species of Sarcocystis spp. Which affect a wide range of ruminants. Amongst the 11 sequences generated from the present study, three haplotypes were noticed. Again, the present isolates formed three clades on the phylogenetic tree, wherein two clades were closer to the sequences of S. levinei earlier reported from India (KX574315). However, the third clade was phylogenetically closer to the Egyptian isolates than their Indian counterparts. These sequences shared 99.7–100.0% and 98.8–100.0% nucleotide identity within themselves and with other isolates across the globe, respectively.
4. Materials and Methods
4.1. Sample Collection
Cardiac tissues were collected from buffaloes slaughtered at abattoirs of Mathura, Unnao, and Aligarh, alongside from dead animals brought for post mortem examination at the Department of Pathology, DUVASU, Mathura, India. Selection of the heart for collection of sarcocysts was based on the proven fact that only S. levinei has a preference for cardiac tissue, thereby minimizing the chances of picking up any kind of mixed infection. The respective cardiac tissue was observed under a microscope for the presence of sarcocysts following the standard muscle squash technique with methylene blue addition [12]. The sarcocysts were identified based on the morphological features reported in the literature [2,16]. Thereafter, the individual sarcocysts (one per animal) were stored at −20 °C.
4.2. DNA Isolation, Primer Selection, and Cloning of 18S rRNA and Cox 1 Genes of S. levinei
DNA was isolated from individual sarcocysts using a commercial DNA isolation kit (Qiagen, Germany), following the manufacturer’s protocol. The primers for the 18S rRNA and cox 1 genes of Sarcocystis spp. were custom-synthesized using the primer sequence described previously [19]. The PCR conditions for both the genes were followed as described previously [6,19]. The amplified products were visualized in ethidium bromide incorporated into 1.25% agarose gel. Water and known positive DNA of S. tenella (accession number: MH523439) were used as negative and positive templates (for both the genes, as the primers for both of them were genus-specific).Thereafter, PCR products were purified using gel purification and a DNA clean-up kit (Thermo Fisher Scientific, Waltham, MA, USA), following the manufacturer’s protocol. Subsequently, the purified products were cloned into CloneJET PCR Cloning Vector (Fermentas, Thermo Fisher Scientific). The recombinant plasmids were harvested (Plasmid purification kit, Fermentas, Thermo Fisher Scientific), and the purified plasmids were outsourced for bidirectional DNA sequencing to Invitrogen Bio Sciences Pvt. Ltd., Gurugram, India, using pJET1.2 universal primers (specific for the CloneJET cloning vector).
4.3. Sequence Analysis and Construction of Phylogenetic Trees
The sequences were submitted into NCBI, and the accession number corresponding to each sequence was obtained. Thereafter, the sequences were analyzed using Mega 6 and Gene tool software. The alignments of the DNA sequences were obtained with the Clustal W algorithm incorporated into the MEGA6 [27]. A gap opening penalty of 10 and gap extension penalties of 0.1 and 0.2 were used for the pairwise and multiple alignments, respectively. The phylogenetic relationship of S. levinei with other species of Sarcocystis, affecting different hosts, was commutated based on the available nucleotide sequences of the 18S rRNA and cox 1 genes in the NCBI (Figure 2a,b). The respective phylogenetic trees for various isolates of S. levinei were constructed using maximum likelihood (ML) (Figure 2a,b). Thereafter, the nucleotide identity between Indian sequences of S. levinei, alongside sequences across the world, was also calculated (Figure 3a,b). Phylogeny was tested at 1000 bootstrap replications. For 18S rRNA, the length of multiple alignment was 876 bp and the Tamura 3 parameter was chosen as the best nucleotide evolution model. For cox 1, the length of multiple alignment was 822 bp and the Kimura 2 parameter was chosen as the best nucleotide evolution model. Sarcocystis hjorti was used as the outgroup for both analyses. Furthermore, the parameters of intraspecific variation within both analyzed genes are presented in Table 1.
5. Conclusions
The present study is the first detailed report on the phylogenetic characterization of S. levinei from India. Before this study, the sequences of S. levinei were available only from Egypt for comparison. Phylogenetics positioned S. levinei and S. cruzi into two monophyletic sister groups. An appreciable amount of genetic variability was noticed between various S. levinei sequences, both at the 18S and cox 1 gene loci. On the basis of the 18S gene, three haplotypes with– 99.7–100.0% nucleotide identity were observed. Higher intraspecific variation of S. levinei was noticed within cox 1, as 10 haplotypes were identified and the sequence identity was 98.8–100%.The study generated some vital phylogenetic data on this erstwhile-understudied parasite.
Author Contributions
Conceptualization, D.S. and V.S.; methodology, V.S. and R.K.; validation, V.S. and S.P.; investigation, V.S.; data curation, V.S. and S.P.; writing—original draft preparation, V.S.; writing—review and editing, V.S. and S.P.; supervision, D.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The tissues were collected from the animals in compliance with the ethical standards and guidelines of the Institutional Animal Ethics Committee (IAEC), and permission was granted via. order no. IAEC/17/24.
Informed Consent Statement
Not applicable.
Data Availability Statement
The sequences generated in the present study are available at NCBI portal.
Acknowledgments
The authors are highly thankful to the Director of Research, Dean, and Vice Chancellor, DUVASU, for the facilities provided.
Conflicts of Interest
The authors declare that they have no competing interests.
Abbreviations
| 18S rRNA | 18S ribosomal RNA |
| 28S rRNA | 28S ribosomal RNA |
| ITS | Internal Transcribed Spacer |
| cox 1 | mitochondrial cytochrome 1 |
| DNA | Deoxyrobose Nucleic Acid |
| PCR | Polymerase Chain Reaction |
| NCBI | National Center for Biotechnology Information |
| MEGA | Molecular Evolutionary Genetics Analysis |
References
- Chen, X.; Zuo, Y.; Rosenthal, B.M.; He, Y.; Cui, L.; Yang, Z. Sarcocystis sinensis is an ultrastructurally distinct parasite of water buffalo that can cause foodborne illness but cannot complete its life-cycle in human beings. Vet. Parasitol. 2011, 178, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Calero-Bernal, R.; Rosenthal, B.M.; Speer, C.A.; Fayer, R. Sarcocystosis of Animals and Humans, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 243–248. [Google Scholar]
- Ashmawy, K.L.; Somaia, S.; Akkada, A.; Gashir, M.B. Prevalence and molecular characterization of Sarcocystis spp. in water buffaloes (Bubalis bubalis) in Egypt. Trop. Anim. Health Prod. 2014, 46, 1351–1356. [Google Scholar] [CrossRef] [PubMed]
- Oryan, A.; Sharifiyazdi, H.; Khordadmehr, M.; Larki, S. Characterization of Sarcocystis fusiformis based on sequencing and PCR-RFLP in water buffalo (Bubalus bubalis) in Iran. Parasitol. Res. 2011, 109, 1563–1570. [Google Scholar] [CrossRef] [PubMed]
- Gjerde, B.; Hilali, M.; Abbas, I.E. Molecular differentiation of Sarcocystis buffalonis and Sarcocystis levinei in water buffaloes (Bubalus bubalis) from Sarcocystis hirsuta and Sarcocystis cruzi in cattle (Bos taurus). Parasitol. Res. 2016, 115, 2459–2471. [Google Scholar] [CrossRef] [PubMed]
- Sudan, V.; Kumar, R.; Shanker, D.; Paliwal, S. First report of molecular characterization and phylogenetic analysis of Sarcocystis tenella from India. Parasitol. Res. 2019, 118, 1429–1434. [Google Scholar] [CrossRef]
- Sudan, V.; Shanker, D.; Paliwal, S.; Kumar, R.; Singh, A. Phylogenetics of Sarcocystis fusiformis isolates based on 18S rRNA and cox 1 genes. Microb. Pathog. 2021, 159, 105144. [Google Scholar] [CrossRef] [PubMed]
- Gjerde, B. Phylogenetic relationships among Sarcocystis species in cervids, cattle and sheep inferred from the mitochondrial cytochrome c oxidase subunit I gene. Int. J. Parasitol. 2013, 43, 579–591. [Google Scholar] [CrossRef]
- Jehle, C.; Dinkel, A.; Sander, A.; Morent, M.; Romig, T.; Luc, P.; De, T.; Thai, V.; Mackenstedt, U. Diagnosis of Sarcocystis spp. in cattle (Bos taurus) and water buffalo (Bubalus bubalis) in Northern Vietnam. Vet. Parasitol. 2009, 166, 314–320. [Google Scholar] [CrossRef] [Green Version]
- Xiang, Z.; He, Y.; Zhao, H.; Rosenthal, B.M.; Dunams, D.B.; Li, X.; Zuo, Y.; Feng, G.; Cui, L.; Yang, Z. Sarcocystis cruzi: Comparative studies confirm natural infections of buffaloes. Exp. Parasitol. 2011, 127, 460–466. [Google Scholar] [CrossRef]
- Sudan, V.; Shanker, D. Redescription of Sarcocystis Species Affecting Buffaloes in the Wake of Advancement in Molecular Biology in Parasitology: Camera Lucida to OMICS; Himanshu Publishers: Udaipur, India, 2018; pp. 133–141. [Google Scholar]
- Sudan, V.; Shanker, D.; Kumar, R.; Sachan, A. Pathological studies on bubaline tissues naturally infected with Sarcocystis spp. J. Vet. Parasitol. 2019, 33, 8–11. [Google Scholar] [CrossRef]
- Ghoshal, S.B.; Joshi, S.C.; Shah, H.L. A note on the natural occurrence of Sarcocystis in buffaloes (Bubalus bubalis) in Jabalpur region, M.P. Indian Vet. J. 1986, 63, 165–166. [Google Scholar]
- Chhabra, M.B.; Samantaray, S. Sarcocystis and sarcocystosis in India: Status and emerging perspectives. J. Parasit. Dis. 2012, 37, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudan, V.; Kumar, R.; Sachan, D. Molecular identification of Sarcocystis cruzi and S. hirsuta sarcocysts in Mathura, Uttar Pradesh. J. Vet. Parasitol. 2020, 32, 8–11. [Google Scholar]
- Huong, L.T.T.; Dubey, J.P.; Uggla, A. Redescription of Sarcocystis levinei Dissanaike and Kan, 1978 (Protozoa: Sarcocystidae) of the water buffalo (Bubalus bubalis). J. Parasitol. 1997, 83, 1148–1152. [Google Scholar] [CrossRef] [PubMed]
- Gjerde, B.; Hilali, M.; Mawgood, S.A. Molecular characterization of three regions of the nuclear ribosomal DNA unit and the mitochondrial cox 1 gene of Sarcocystis fusiformis from water buffaloes (Bubalus bubalis) in Egypt. Parasitol. Res. 2015, 114, 3401–3413. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.Q.; Zuo, Y.X.; Ding, B.; Chen, X.W.; Luo, J.; Zhang, Y.P. Identification of Sarcocystis hominis-like (Protozoa:Sarcocystidae) cyst in water buffalo (Bubalus bubalis) based on 18S rRNA gene sequences. J. Parasitol. 2001, 87, 934–937. [Google Scholar] [CrossRef]
- Gjerde, B. Molecular characterisation of Sarcocystis bovifelis, Sarcocystis bovini n. sp., Sarcocystis hirsuta and Sarcocystis cruzi from cattle (Bos taurus) and Sarcocystis sinensis from water buffaloes (Bubalus bubalis). Parasitol. Res. 2015, 115, 1473–1492. [Google Scholar] [CrossRef]
- Sudan, V.; Kumar, R.; Shanker, D.; Singh, A. Sequence phylogenetic analysis and associative genetic diversity of Sarcocystis hirsuta based on 18S rRNA gene. Beni Suef Univ. J. Basic Appl. Sci. 2021, 10, 22. [Google Scholar] [CrossRef]
- Rosenthal, B.M.; Dunams, D.B.; Pritt, B. Restricted genetic diversity in the ubiquitous cattle parasite, Sarcocystis cruzi. Infect. Genet. Evol. 2008, 8, 588–592. [Google Scholar] [CrossRef]
- Bhoora, R.; Franssen, L.; Oosthuizen, M.C.; Guthrie, A.J.; Zweygarth, E.; Penzhorn, B.L.; Jongejan, F.; Collins, N.E. Sequence heterogeneity in the 18S rRNA gene within Theileria equi and Babesia caballi from horses in South Africa. Vet. Parasitol. 2009, 159, 112–120. [Google Scholar] [CrossRef] [Green Version]
- El-Sherry, S.; Ogedengbe, M.E.; Hafeez, M.A.; Barta, J.R. Divergent nuclear 18S rDNA paralogs in a turkey coccidium, Eimeria meleagrimitis, complicate molecular systematics and identification. Int. J. Parasitol. 2013, 43, 679–685. [Google Scholar] [CrossRef] [PubMed]
- Vrba, V.; Poplstein, M.; Pakandl, M. The discovery of the two types of small subunit ribosomal RNA gene in Eimeria mitis contests the existence of E. mivati as an independent species. Vet. Parasitol. 2011, 183, 47–53. [Google Scholar] [CrossRef] [PubMed]
- El-Seify, M.; El-Morsey, A.; Hilali, M.A.; Zayed, A.; El-Dakhly, K.; Haridy, M.; Sakai, H.; Yanai, T.; Seify, E. Molecular characterization of Sarcocytis fusiformis and Sarcocystis buffalonis infecting water buffaloes (Bubalus bubalis) from Egypt. Am. J. Anim. Vet. Sci. 2014, 9, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Holmdahl, O.J.; Mattsson, J.G.; Uggla, A.; Johansson, K.E. The phylogeny of Neospora caninum and Toxoplasma gondii based on ribosomal RNA sequences. FEMS Microbiol. Lett. 1994, 119, 187–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Squashed and methylene-blue-stained preparation of heart tissue showing characteristic elongated sarcocysts.
Figure 1.
Squashed and methylene-blue-stained preparation of heart tissue showing characteristic elongated sarcocysts.

Figure 2.
Phylogenetic tree for selected members of Sarcocystidae, inferred using the maximum parsimony method (a) based on partial sequences of 18S and (b) based on partial sequences of cox 1. The numbers represent the boot strap values. Sub-trees formed by two or more sequences/haplotypes of the same species were collapsed, but the quantities of included sequences/haplotypes are given behind the taxon names.
Figure 2.
Phylogenetic tree for selected members of Sarcocystidae, inferred using the maximum parsimony method (a) based on partial sequences of 18S and (b) based on partial sequences of cox 1. The numbers represent the boot strap values. Sub-trees formed by two or more sequences/haplotypes of the same species were collapsed, but the quantities of included sequences/haplotypes are given behind the taxon names.


Figure 3.
Phylogenetic relationship of S. levinei Indian isolates with other isolates across the world, based on the (a) 18S and (b) cox 1 genes. All accession numbers correspond to different S. levinei isolates, followed by their country of origin.
Figure 3.
Phylogenetic relationship of S. levinei Indian isolates with other isolates across the world, based on the (a) 18S and (b) cox 1 genes. All accession numbers correspond to different S. levinei isolates, followed by their country of origin.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Comparative study on the sequences of S. levinei based on both genes (Nh: number of haplotypes; Hd: haplotype diversity; Pi: nucleotide diversity; S: number of segregating sites; K: average number of nucleotide differences).
Table 1.
Comparative study on the sequences of S. levinei based on both genes (Nh: number of haplotypes; Hd: haplotype diversity; Pi: nucleotide diversity; S: number of segregating sites; K: average number of nucleotide differences).
| Marker | Nh | Hd | Pi | S | K |
|---|---|---|---|---|---|
| Cox 1 | 10 | 0.825 | 0.00336 | 19 | 2.76308 |
| 18S | 5 | 0.679 | 0.00118 | 3 | 1.00526 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sudan, V.; Shanker, D.; Paliwal, S.; Kumar, R. Associative Genetic Diversity of Sarcocystis levinei Isolates across the Globe. Parasitologia 2023, 3, 231-240. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3030024
AMA Style
Sudan V, Shanker D, Paliwal S, Kumar R. Associative Genetic Diversity of Sarcocystis levinei Isolates across the Globe. Parasitologia. 2023; 3(3):231-240. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3030024
Chicago/Turabian StyleSudan, Vikrant, Daya Shanker, Sanjhi Paliwal, and Rahul Kumar. 2023. "Associative Genetic Diversity of Sarcocystis levinei Isolates across the Globe" Parasitologia 3, no. 3: 231-240. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia3030024