
Host Choice and Feeding Behaviours of Glossina morsitans Offspring Whose Parents Were Fed on Different Host Species

Abstract
:1. Introduction
2. Materials and Methods
2.1. Rearing the Tsetse Flies
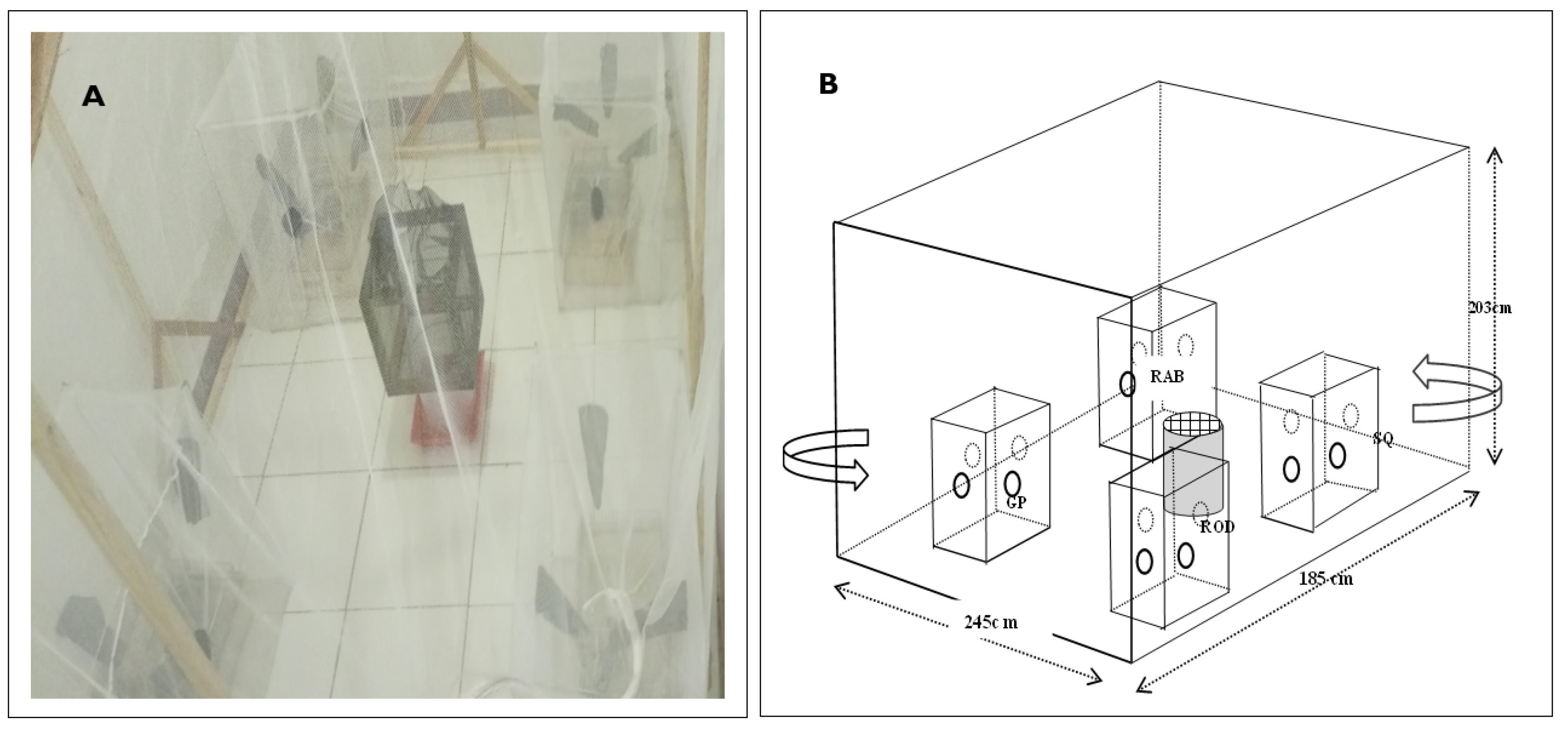
2.2. Experimental Setup
2.3. Assessing the Host Choice and Feeding Success of Offspring Whose Mothers Were Fed on Different Host Species
2.4. Determination of Haemoglobin (Hb) Concentrations and Total Plasma Protein of Blood Samples from the Experimental Host Species
2.5. Data Analysis
2.6. Ethical Approval
3. Results
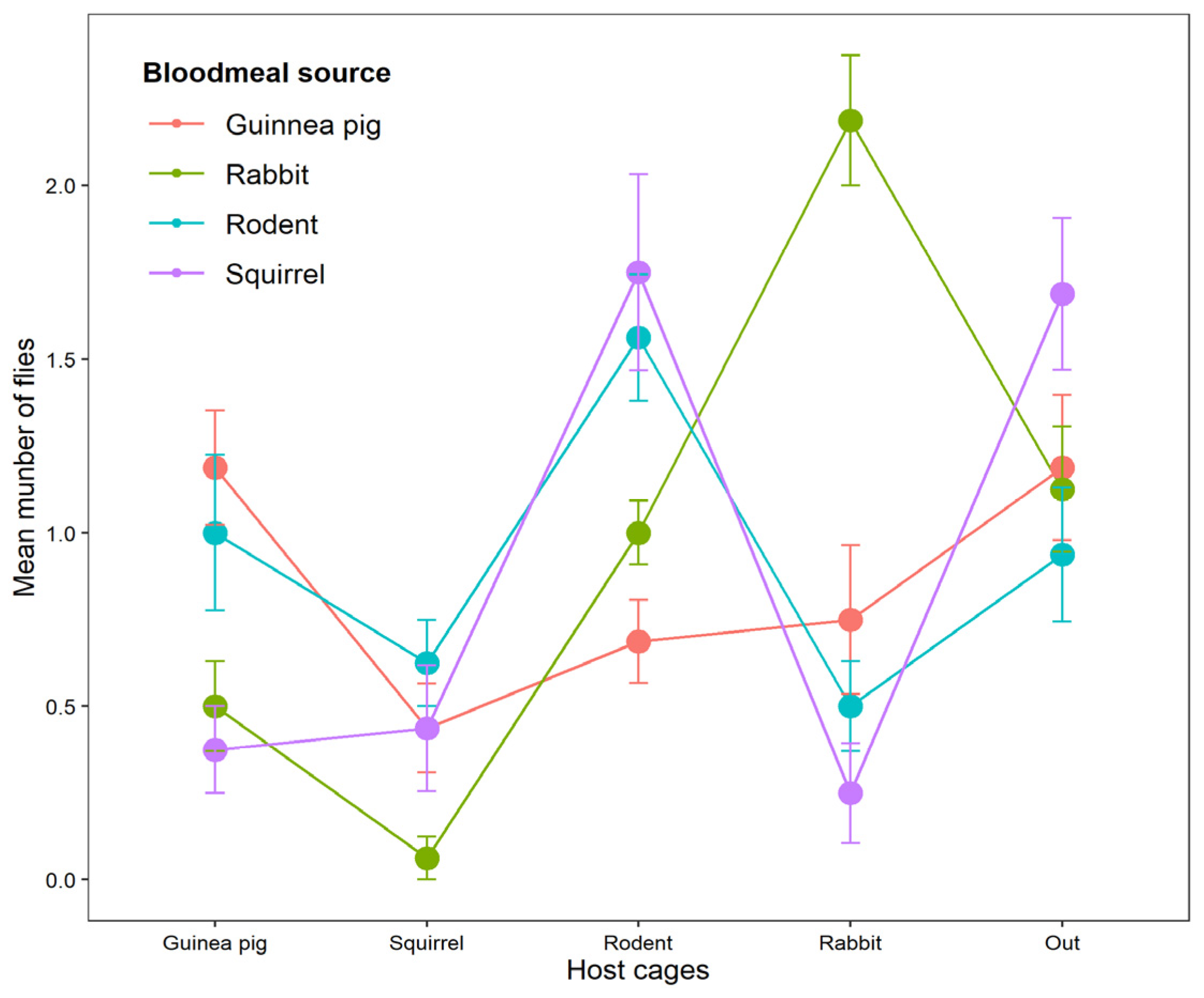
3.1. Choice of Adult Tsetse Fly, Glossina Morsitans, on Different Host Species
3.2. Feeding Success of Tsetse Flies Attracted to Different Hosts
3.3. Haemoglobin Concentration (Hb) and Total Plasma Protein in Different Hosts
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vreysen, M.J.B.; Seck, M.T.; Sall, B.; Bouyer, J. Tsetse flies: Their biology and control using area-wide integrated pest management approaches. J. Invertebr. Pathol. 2013, 112 (Suppl. S1), S15–S25. [Google Scholar] [CrossRef] [PubMed]
- Gashururu, R.S.; Githigia, S.M.; Gasana, M.N.; Habimana, R.; Maingi, N.; Cecchi, G.; Paone, M.; Zhao, W.; Masiga, D.K.; Gashumba, J. An update on the distribution of Glossina (tsetse flies) at the wildlife-human-livestock interface of Akagera National Park, Rwanda. Parasites Vectors 2021, 14, 294. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Control and Surveillance of Human African Trypanosomiasis; World Health Organization: Geneva, Switzerland, 2013; pp. 1–237. [Google Scholar]
- Gooding, R.H.; Krafsur, E.S. Tsetse genetics: Contributions to biology, systematics, and control of tsetse flies. Annu. Rev. Entomol. 2005, 50, 101–123. [Google Scholar] [CrossRef]
- Cecchi, G.; Mattioli, R.C.; Slingenbergh, J.; De La Rocque, S. Land cover and tsetse fly distributions in sub-Saharan Africa. Med. Vet. Entomol. 2008, 22, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Bouteille, B. Human African Trypanosomiasis. In Encyclopedia of the Neurological Sciences, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 2, pp. 618–624. [Google Scholar] [CrossRef]
- Moloo, S.K. Distribution of Glossina species in Africa. Acta Trop. 1985, 42, 275–281. [Google Scholar]
- Malele, A.I.I.; Kinung, S.M.; Nyingilili, H.S.; Matemba, L.E.; Sahani, J.K.; Mlengeya, T.D.K.; Wambura, M.; Kibona, S.N. Glossina dynamics in and around the sleeping sickness endemic Serengeti ecosystem of northwestern Tanzania Glossina dynamics in and around the sleeping sickness endemic Serengeti ecosystem of northwestern Tanzania. J. Vector Ecol. 2007, 32, 263–268. [Google Scholar] [CrossRef]
- Kweka, E.J.; Kimaro, E.E.; Kweka, J.; Kimaro, E.; Kimaro, E.G.; Nagagi, Y.P.; Malele, I.I. Major Disease Vectors in Tanzania: Distribution, Major Disease Vectors in Tanzania: Distribution, Control and Challenges Control and Challenges. In Biological Control of Pest and Vector Insects; IntechOpen: London, UK, 2017. [Google Scholar]
- Malele, I.; Craske, L.; Knight, C.; Ferris, V.; Njiru, Z.; Hamilton, P.; Lehane, S.; Lehane, M.; Gibson, W. The use of specific and generic primers to identify trypanosome infections of wild tsetse flies in Tanzania by PCR. Infect. Genet. Evol. 2003, 3, 271–279. [Google Scholar] [CrossRef]
- Malele, I.I.; Magwisha, H.B.; Nyingilili, H.S.; Mamiro, K.A.; Rukambile, E.J.; Daffa, J.W.; Lyaruu, E.A.; Kapange, L.A.; Kasilagila, G.K.; Lwitiko, N.K. Multiple Trypanosoma infections are common amongst Glossina species in the new farming areas of Rufiji district, Tanzania. Parasites Vectors 2011, 4, 217. [Google Scholar] [CrossRef]
- Makhulu, E.E.; Villinger, J.; Adunga, V.O.; Jeneby, M.M.; Kimathi, E.M.; Mararo, E.; Oundo, J.W.; Musa, A.A.; Wambua, L. Tsetse blood-meal sources, endosymbionts and trypanosome-associations in the Maasai mara national reserve, a wildlife-human-livestock interface. PLoS Negl. Trop. Dis. 2021, 15, e0008267. [Google Scholar] [CrossRef]
- Jaenike, J. Host specialization in phytophagous insects. Annu. Rev. Ecol. Syst. 1990, 21, 243–273. [Google Scholar] [CrossRef]
- Vinauger, C.; Lahondère, C.; Cohuet, A.; Lazzari, C.R.; Riffell, J.A. Learning and Memory in Disease Vector Insects. Trends Parasitol. 2016, 32, 761–771. [Google Scholar] [CrossRef] [PubMed]
- McCall, P.J.; Kelly, D.W. Learning and memory in disease vectors. Trends Parasitol. 2002, 18, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Campbell-Lendrum, D.H.; Brandão-Filho, S.P.; Ready, P.D.; Davies, C.R. Host and/or site loyalty of Lutzomyia whitmani (Diptera: Psychodidae) in Brazil. Med. Vet. Entomol. 1999, 13, 209–211. [Google Scholar] [CrossRef] [PubMed]
- Bouyer, J.; Pruvot, M.; Bengaly, Z.; Guerin, P.M.; Lancelot, R. Learning influences host choice in tsetse. Biol. Lett. 2007, 3, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Barron, A.B. The Life and Death of Hopkins’ Host-Selection Principle. J. Insect Behav. 2001, 14, 725–737. [Google Scholar] [CrossRef]
- Weitz, B. The feeding habits of Glossina. Bull. World Health Organ. 1963, 28, 711–729. [Google Scholar]
- Onyiah, J.A. Mechanism of host selection by tsetse flies. Int. J. Trop. Insect Sci. 1980, 1, 31–34. [Google Scholar] [CrossRef]
- Clausen, P.H.; Adeyemi, I.; Bauer, B.; Breloeer, M.; Salchow, F.; Staak, C. Host preferences of tsetse (Diptera: Glossinidae) based on bloodmeal identifications. Med. Vet. Entomol. 1998, 12, 169–180. [Google Scholar] [CrossRef]
- Barrozo, R.B. Food recognition in hematophagous insects. Curr. Opin. Insect Sci. 2019, 34, 55–60. [Google Scholar] [CrossRef]
- Camara, K.; Ilboudo, K.; Salou, E.W.; Gimonneau, G. Evaluation of different blood-feeding frequencies on Glossina palpalis gambiensis performance in a mass-rearing insectary. Parasites Vectors 2021, 14, 46. [Google Scholar] [CrossRef]
- Opiyo, E.; Parker, A.G.; Mohammed, A.H. Standard Operating Procedures for Mass Rearing Tsetse Flies; FAO/IAEA: Vienna, Austria, 2006; Volume 239. [Google Scholar]
- Posner, L.P.; Erb, H.N.; Gleed, R.D. The HemoCue® for point-of-care hemoglobin measurement and packed cell volume estimation in cats. J. Vet. Emerg. Crit. Care 2005, 15, 22–25. [Google Scholar] [CrossRef]
- Nagagi, Y.P.; Silayo, R.S.; Kweka, E.J. Advancements in bait technology to control Glossina swynnertoni Austen, the species of limited distribution in Kenya and Tanzania border: A review. J. Vector Borne Dis. 2017, 54, 16–24. [Google Scholar] [PubMed]
- Boyt, W.P.; Mackenzie, P.K.I.; Pilson, R.D. The relative attractiveness of donkeys, cattle, sheep and goats to Glossina morsitans morsitans Westwood and G. pallidipes Austen (Diptera: Glos-sinidae) in a middle-veld area of Rhodesia. Bull. Entomol. Res. 1978, 68, 497–500. [Google Scholar] [CrossRef]
- Bouyer, J.; Cuisance, D.; Messad, S.; Guérin, P.M. Learning Affects Host Preference in Tsetse Flies. Rev. D’elevage Médecine Vétérinaire Pays Trop. 2005, 58, 27–29. [Google Scholar] [CrossRef]
- Green, C.H. The use of two-coloured screens for catching Glossina palpalis palpalis. Bull. Entomol. Res. 1989, 79, 81–93. [Google Scholar] [CrossRef]
- Gibson, G.; Torr, S.J. Visual and olfactory responses of haematophagous Diptera to host stimuli. Med. Vet. Entomol. 1999, 13, 2–23. [Google Scholar] [CrossRef]
- Torr, S.J. The host-orientated behaviour of tsetse flies (Glossina): The interaction of visual and olfactory stimuli. Physiol. Entomol. 1989, 14, 325–340. [Google Scholar] [CrossRef]
- Mwandawiro, C.; Boots, M.; Tuna, N.; Suwonkerd, W. Transactions of the Royal Society of Tropical Medicine and Hygiene; Royal Society of Tropical Medicine and Hygiene: London, UK, 2000; Volume 94, pp. 238–242. [Google Scholar]
- Phasomkusolsil, S.; Tawong, J.; Monkanna, N.; Pantuwatana, K.; Damdangdee, N.; Khongtak, W.; Kertmanee, Y.; Evans, B.P.; Schuster, A.L. Maintenance of mosquito vectors: Effects of blood source on feeding, survival, fecundity, and egg hatching rates. J. Vector Ecol. 2013, 38, 38–45. [Google Scholar] [CrossRef]
- Al-rashidi, H.S.; Alghamdi, K.M.; Al-otaibi, W.M.; Al-Solami, H.M.; Mahyoub, J.A. Saudi Journal of Biological Sciences Effects of blood meal sources on the biological characteristics of Aedes aegypti and Culex pipiens (Diptera: Culicidae). Saudi J. Biol. Sci. 2022, 29, 103448. [Google Scholar] [CrossRef]
- Khan, S.A.; Kassim, N.F.A.; Webb, C.E.; Aqueel, M.A.; Ahmad, S.; Malik, S.; Hussain, T. Human blood type influences the host-seeking behavior and fecundity of the Asian malaria vector Anopheles stephensi. Sci. Rep. 2021, 11, 24298. [Google Scholar] [CrossRef]
- Hawkey, C.M.; Bennett, P.M.; Gascoyne, S.C.; Hart, M.G.; Kirkwood, J.K. Erythrocyte size, number and haemoglobin content in vertebrates. Br. J. Haematol. 1991, 77, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Walker, E.D.; Edman, J.D. The influence of host defensive behavior on mosquito (diptera: Culicidae) biting persistence. J. Med. Entomol. 1985, 22, 370–372. [Google Scholar] [CrossRef] [PubMed]
- Takken, W.; Smallegange, R.C.; Vigneau, A.J.; Johnston, V.; Brown, M.; Mordue-Luntz, A.J.; Billingsley, P.F. Larval nutrition differentially affects adult fitness and Plasmodium development in the malaria vectors Anopheles gambiae and Anopheles stephensi. Parasites Vectors 2013, 6, 345. [Google Scholar] [CrossRef]
- Schofield, S.; Torr, S.J. A comparison of the feeding behaviour of tsetse and stable flies. Med. Vet. Entomol. 2002, 16, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Torr, S.J.; Mangwiro, T.N.C. Interactions between cattle and biting flies: Effects on the feeding rate of tsetse. Med. Vet. Entomol. 2000, 14, 400–409. [Google Scholar] [CrossRef]
- Lyimo, I.N.; Ferguson, H.M. Ecological and evolutionary determinants of host species choice in mosquito vectors. Trends Parasitol. 2009, 25, 189–196. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Bloodmeal Source | Host Cages | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Guinea Pig | Rabbit | Rodent | Squirrel | Out | ||||||
| n | % | n | % | n | % | n | % | n | % | |
| Guinea pig | 19 | 38.78 | 12 | 20.34 | 11 | 13.75 | 7 | 28.00 | 19 | 24.05 |
| Rabbit | 8 | 16.33 | 35 | 59.32 | 16 | 20.00 | 1 | 4.00 | 18 | 22.78 |
| Rodent | 16 | 32.65 | 8 | 13.56 | 25 | 31.25 | 10 | 40.00 | 15 | 18.99 |
| Squirrel | 6 | 12.24 | 4 | 6.78 | 28 | 35.00 | 7 | 28.00 | 27 | 34.18 |
| TOTAL | 49 | 100.00 | 59 | 100.00 | 80 | 100.00 | 25 | 100.00 | 79 | 100.00 |
| Host Cages | Blood Meal Sources | ||||
|---|---|---|---|---|---|
| Guinea Pig | Rabbit | Rodent | Squirrel | Total | |
| Guinea pig | 1 | 0 | 7 | 2 | 10 |
| Rabbit | 1 | 3 | 1 | 1 | 6 |
| Rodent | 0 | 1 | 9 | 13 | 23 |
| Squirrel | 0 | 0 | 0 | 0 | 0 |
| Total | 2 | 4 | 17 | 16 | 39 |
| Host Type | Hb Concentration (g/dL) | Total Plasma Protein (g/dL) | Number of Flies (n, %) | |||
|---|---|---|---|---|---|---|
| Mean | Std. Error | Mean | Std. Error | Attracted | Bloodfed | |
| Guinea pig | 14.730 | 0.071 | 52.695 | 0.033 | 49 (23.00%) | 10 (25.64%) |
| Rabbit | 14.515 | 0.050 | 65.225 | 0.003 | 59 (27.7%) | 6 (15.38%) |
| Rodent | 17.335 | 0.244 | 75.170 | 0.497 | 80 (37.56%) | 23 (58.97%) |
| Squirrel | 19.320 | 0.505 | 7.756 | 0.028 | 25 (11.74%) | 0 (0.000%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mdee, F.E.; Lyatuu, J.; Mafie, E.; Mnyone, L.L. Host Choice and Feeding Behaviours of Glossina morsitans Offspring Whose Parents Were Fed on Different Host Species. Parasitologia 2024, 4, 38-46. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia4010003
Mdee FE, Lyatuu J, Mafie E, Mnyone LL. Host Choice and Feeding Behaviours of Glossina morsitans Offspring Whose Parents Were Fed on Different Host Species. Parasitologia. 2024; 4(1):38-46. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia4010003
Chicago/Turabian StyleMdee, Filbert E., Jeremiah Lyatuu, Eliakunda Mafie, and Ladslaus L. Mnyone. 2024. "Host Choice and Feeding Behaviours of Glossina morsitans Offspring Whose Parents Were Fed on Different Host Species" Parasitologia 4, no. 1: 38-46. https://0-doi-org.brum.beds.ac.uk/10.3390/parasitologia4010003