


Abscisic Acid: Metabolism, Signaling, and Crosstalk with Other Phytohormones under Heavy Metal Stress


1
Department of Biosciences, Faculty of Science, Integral University, Lucknow 226026, India
2
Molecular Department, RML Mehrotra Pathology, Lucknow 226020, India
3
Plant Molecular Biology Laboratory, Department of Botany, Dayanand Anglo-Vedic (PG) College, Chhatrapati Shahu Ji Maharaj University, Kanpur 208001, India
4
NUS Environmental Research Institute, National University of Singapore, #02-01, T-Lab Building, 5A Engineering Drive 1, Singapore 117411, Singapore
5
Energy and Environmental Sustainability for Megacities (E2S2) Phase II, Campus for Research Excellence and Technological Enterprise (CREATE), 1 CREATE Way, Singapore 138602, Singapore
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Stresses 2023, 3(4), 665-686; https://0-doi-org.brum.beds.ac.uk/10.3390/stresses3040046
Submission received: 8 August 2023
/
Revised: 17 September 2023
/
Accepted: 18 September 2023
/
Published: 22 September 2023
(This article belongs to the Topic Effect of Heavy Metals on Plants)
Abstract
:Heavy metal (HM) stress poses a global risk to crops, ecological systems, and human health. It disrupts cellular ionic equilibrium, cell membrane integrity, metabolic balance, and the activities of enzymes and proteins, severely impacting physiological processes, plant development, and agricultural productivity. Although plants naturally activate defense mechanisms to mitigate the adverse effects of HM stress, they cannot completely prevent them. Phytohormones counter HM toxicity, aiding growth. External application and internal regulation via signaling/biosynthesis genes offer defense against HM-induced damage. A pivotal signaling molecule in plant adaptive responses to environmental stressors, including HM toxicity, is abscisic acid (ABA). Despite ABA’s role in abiotic stress responses such as drought and salinity, its function and crosstalk with other phytohormones under HM stress remain poorly understood. Nonetheless, exogenously applied ABA serves as a strategic approach to enhancing plants’ resistance to HM toxicity by promoting osmolyte accumulation and reinforcing antioxidant activity. ABA significantly regulates various plant growth and metabolic activities under diverse environmental conditions. This review highlights the effects of HM stress on plants and explores ABA involvement in production, signaling, catabolism, and transport within plant tissues. The purpose of this paper is to shed light on the complex interplay between the metabolism of ABA, its signaling, and its interactions with other phytohormones (e.g., auxins, gibberellins, and ethylene) during HM exposure. Furthermore, we delve into the function of ABA to mitigate HM stress and elucidate its interactions with other phytohormones.
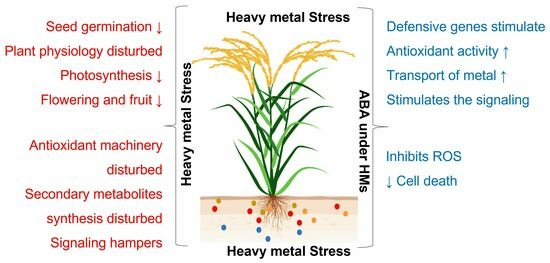
1. Introduction
Abiotic stresses, such as metal, heat, drought, cold, and salt, that have a direct impact on overall agricultural production are continually exposed to plants over a long period [1]. Heavy metal (HM) stress poses a more serious and challenging threat than pesticides and other major pollutants, including carbon dioxide and sulfur dioxide, making it a critical concern in the realm of abiotic stressors. HMs are metallic elements that are nonbiodegradable by nature and have a higher density than water [2]. The threat posed by HM contamination in soil on the worldwide agricultural system has grown significantly [3]. The buildup of these HMs and metalloids degrades soil quality, which undermines the stability of the entire food chain. The list of HMs and metalloids includes mercury (Hg), lead (Pb), chromium (Cr), iron (Fe), cadmium (Cd), zinc (Zn), copper (Cu), cobalt (Co), nickel (Ni), and arsenic (As); these are crucial for both metabolism and plant growth and development. On the other hand, these HMs may have negative impacts on the physiology as well as the biochemistry of plants when they accumulate in a higher concentration [4,5,6].
Researchers have also predicted that, in the near future, HMs will transcend solid and nuclear waste, elevating themselves to the status of one of the most hazardous pollutants with significant repercussions for humanity [7,8].
Modern agricultural practices, accelerating anthropogenic activity, and recent rapid industrialization have all contributed to a significant increase in HM accumulation in plants [9]. Key plant processes like ion distribution, photosynthesis, mineral feeding, and water uptake are negatively impacted, as the soil deposits and roots are responsible for the entry of HMs [10]. Plants use different strategies to decrease HM toxicity, including osmoprotectant biosynthesis and accumulation, compartmentalization, exclusion, the synthesis of metallothioneins and phytochelatins, and others [11,12]. However, plants get harmed due to the rise in threshold level by the accumulation of HMs, which affect the majority of biological processes [13]. Plants essentially reduce the consequences of HM stress by controlling the process of HM absorption and translocation, their chelation/compartmentation, and their transport by inducing ion homeostasis, structural modifications of different plant parts, antioxidant defense pathway activation, osmoregulation, and metal ion transporter induction [14,15]. Additionally, a number of chemical and biological techniques have been used to counteract the hazardous effects of HMs from contaminated water and soil resources. They involve stabilization or in situ fixation, soil washing, adsorption, ion exchange, excavation to remove HMs, filtration, chemical precipitation, electrochemical procedures, solvent extraction, phytoremediation, and coagulation/flocculation [16,17,18]. Recent research has also emphasized the use of biochar, compost, and nanoparticles for boosting plant development and minimizing the impacts of HM toxicity [19,20,21,22]. Furthermore, extensive effort has been made to develop crop cultivars resistant to HM stress through plant breeding, genetic engineering, and genomics methods [23].
In recent years, the application of phytohormones as an alternative environmentally benign strategy to increase plant HM stress resistance has gained traction [8]. The chemical messengers known as phytohormones are created at quite low concentrations and are essential for controlling a variety of plant growth and developmental processes [24,25]. In response to abiotic stressors, they also activate a variety of signal transduction pathways [26,27]. According to earlier studies, stress tolerance increased in plants exposed to HMs by exogenous treatment with phytohormones [8]. As a result of HM exposure, the phytohormone endogenous levels also alter, modulating a number of plant stress adaptation traits [8,11]. The characterization and identification of various pathways linked to HM stress mediated through phytohormones have been permitted by a lot of research tools like microarrays, system biology, mutant screening, expression profiling, proteomics, and bioinformatics [28]. There are few reports available as evidence that show that phytohormones are responsible for different responsive signaling components of HM stress. In the context of this review, ABA a crucial phytohormone takes center stage as we explore its multifaceted role in the context of HM stress. ABA selection as the focal point of this study is driven by its pivotal function in mediating plant responses to HM stress, including its intricate crosstalk with other phytohormones. Additionally, we delve into ABA’s significance in devising effective strategies for mitigating the adverse impacts of HMs on plants, offering insights into the intricate interplay between phytohormones and their contribution to HM stress adaptation and tolerance mechanisms.
2. Abscisic Acid
A sesquiterpene phytohormone called ABA performs crucial roles in plant growth and development. For example, it induces dormancy in seeds and buds, causes stomatal closure, and encourages flower and fruit abscission [29,30]. Although ABA was first identified in the 1960s [31], its physiological effects have not yet been thoroughly investigated. Plants have an adaptation process known as seed dormancy that allows them to quickly go into dormancy in unfavorable environmental conditions. The hormone ABA is the primary hormone that initiates and sustains seed dormancy [32], and it is one of several hormonal routes that regulate how precisely seeds react to environmental conditions.
When plants need to close their stomata, the root-produced ABA moves to the leaves. Recent research suggests that signals related to water flow can also boost ABA production in leaves [33]. Later research [34] has suggested that ABA may also be synthesized by the vascular tubes and guard cells of nutritive organs. ABA induces the stomata of guard cells to close, according to all specialists, despite disagreements among them on the location of ABA synthesis. Another point of concern among scholars is the abscission of flowers, leaves, and fruits. Primarily, it was assumed that ABA would cause plant tissues to abscise [35]. Later, scientists proposed that ethylene and ABA might possibly be involved in the process of abscission [36,37]. Other researchers, however, have also proposed that ethylene may play a direct role and that ABA’s contribution may be accidental [38]. Additionally, in plants, ABA actively participates as a natural “stress hormone” that responds to abiotic stresses [39]. Exogenous ABA injection has been demonstrated to decrease HM accumulation and enhance HM tolerance in plants [40,41,42]. Spraying ABA on Pingyi sweet tea roots reduced cell death, H2O2 and malondialdehyde buildup, leaf transpiration rate, plant Cd content, Cd2+ influx in the root system, and Cd transport from the root system to the shoots, all of which were considerably reduced by Cd stress [43]. When sprayed, the ABA synthesis inhibitor fluridone (Flu) produced the opposite effects [43], but the amount of Cd+ can be decreased by ABA treatment (Malus hupehensis Rehd. var. pingyiensis Jiang) under Cd stress [44]. Figure 1 represents an overview of the ABA mechanism under normal and HM stress conditions.
It is widely acknowledged that ABA is essential for enhancing crop plant morphological characteristics, developmental aspects, quality, yield metrics, and mitigating the negative effects of HM stress.
3. Abscisic Acid Biosynthesis
ABA is crucial for both healthy growth and development and enhances plants’ ability to adapt to environmental challenges. According to Humplik et al. [45], tissue and dose sensitivity are used to check for ABA activation or inhibition. Fine-tuning the de novo biosynthesis and catabolism of ABA allows for precise and appropriate management of its levels. Several studies have revealed that the endogenous ABA concentration increases in response to HM stress [46,47]. Under HM stress, Tamarix hispida showed enhanced expression of the ABA uncoupling enzyme β-glucosyltransferase and the ABA production pathway enzyme isoprenoid synthetase [48]. The ABA production pathway enzymes zeaxanthin epoxidase (ZEP) and ζ-carotene dehydrogenase (ZDS) interestingly showed a negative tendency under identical conditions [48]. Figure 2 depicts the catabolic and synthesis routes of ABA. Isopentenyl diphosphate (IPP) and its isomer, dimethylallyl diphosphate (DMAPP), both contain five carbon atoms and are necessary for the synthesis of ABA [49,50]. In plants, IPP is produced via methylerythritol phosphate (MEP) and mevalonate (MVA) pathways [50,51,52,53]. Both the MVA and MEP routes can create IPP through six successive steps during the IPP synthesis phase [51,52,53]. An IPP: DMAPP isomerase (IDI) interconverts IPP and DMAPP [50,51].
In the initial phase of carotenoid synthesis, IPP (isopentenyl diphosphate) undergoes a series of condensation reactions, sequentially adding one isoprene unit at a time. This process leads to the production of various compounds, including geranylgeranyl phosphate (GGPP), farnesyl pyrophosphate, and others [33]. Subsequently, two GGPP molecules are fused head-to-head by phytoene synthase, resulting in the formation of colorless phytoene [33,54]. The crimson trans-lycopene is created by a four-step dehydrogenation of phytoene [33,49]. Lycopene ε-cyclase and lycopene β-cyclase, respectively, cycle the trans-lycopene and introduce ε- or β-rings [35,54]. An ε-ring and a β-ring form α-carotene along with its derivatives [49,54]; to form β-carotene and its derivatives, two β-rings are introduced [49,54]. β-carotene hydroxylase catalyzes the hydroxylation of β-carotene to form β-cryptoxanthin, which is then further hydroxylated to form zeaxanthin [49]. Trans-violaxanthin is produced when zeaxanthin is transformed into antheraxanthin by the enzyme zeaxanthin epoxidase [54,55]. Trans-violaxanthin de-epoxidase can be converted into zeaxanthin in a reversible process known as the xanthophyll cycle, which can offer plants effective photoprotection [55]. Trans-neoxanthin can be transformed into cis-neoxanthin, trans-violet xanthin, and cis-violaxanthin [56]. Using 9-cis-epoxy carotenoid dioxygenase (NCED), cis-violet xanthin and cis-neoxanthin are split in the plastid to produce xanthoxin and the C25 metabolite [57]. Short-chain dehydrogenase (SDR) catalyzes the conversion of xanthoxin into abscisic aldehydes in the cytoplasm [56].
Abscisic aldehyde oxidase and molybdenum cofactor play a role in cleaving abscisic aldehyde, leading to the synthesis of ABA [56]. Additionally, Jia et al. [58] discovered an alternative ABA synthesis pathway independent of zeaxanthin epoxidase (ZEP). This pathway starts from β-carotene and zeaxanthin carotenoid cleavage of ROS and dioxygenases involved in producing β-carotene and zeaxanthin. Subsequently, 3-OH-β-apo-11-carotenoid aldehyde and β-apo11-carotenoid aldehyde are formed. Through the action of isomerase and hydroxylase, the former converts into 9-cis-3-β-apo-11-carotenoid aldehyde, and the latter forms 9-cis-β-apo-11-carotenoid aldehyde. Furthermore, in the presence of isomerase, 9-cis-3-OH-β-apo-11 carotenoid aldehyde can produce 9-cis-3-OH-β-apo-11-carotenal, and this compound can be further transformed into ABA when cyclooxygenase is present. This alternative pathway highlights the diverse and intricate mechanism involved in ABA synthesis from the carotenoid precursor.
The two primary mechanisms used to catabolize ABA are hydroxylation and conjugation [56,59]. Depending on where the methyl group is being oxidized, three distinct ABA hydroxylation routes are seen [59,60]. The most common hydroxylation process is 80-hydroxylation, which results in 80-OH-ABA. The subsequent independent isomerization of 80-OH-ABA results in phaseic acid (PA) [59], which is then further transformed into dihydro-phaseic acid (DPA) by PA reductase [61].
ABA levels are delicately managed by synthesis and catabolism, as previously mentioned; however, this is insufficient. The organism needs a more effective means of response than ABA production and catabolism when the external environment changes significantly. This issue is resolved by the coupling cycle’s presence. The inactive ABA-glucose ester (ABA-GE) is formed when ABA and glucose are properly linked [33]. ABA is stored and transported through ABA-glucose ester, which is restricted to the endoplasmic reticulum as well as the vacuole [33,61]. ABA-GE can be quickly transformed into ABA by BG1 (β-glucosidase 1) and BG2 (β-glucosidase 2) in response to changes in the external environment, which is crucial for responding to unfavorable conditions [61].
Therefore, under both favorable and unfavorable circumstances, ABA catabolism considerably regulates ABA levels. HM toxicity reduces endogenous ABA levels by preventing the gene expression necessary for ABA production, which halts the physiological impact mediated by ABA.
4. Transport of Abscisic Acid
ABA is chiefly synthesized in vascular tissue and plays a crucial role in regulating various physio-biochemical processes in response to diverse environmental conditions [62]. Within plants, ABA exists in both protonic (ABAH) and anionic (ABA-) forms. In its protonated state, ABA acts as a weak acid, facilitating passive diffusion across the biomembrane [63]. Active translocation of ABA across the plasma membrane is facilitated by various transporters, including toxic and di/tri-peptide or nitrate transporters [PTR/NRT1family proteins (NPF)], multidrug compound extrusion (MATE)-type/DTX transporters, ATP-binding cassette (ABC) transporter proteins, and AWPM-19 family proteins (OsPM1). These transporters enable efficient movement of ABA within the plant’s cells and tissues. Notably, the identification of these transporters originated from studies in Oryza sativa L., commonly known as rice [64,65,66]. ABC transporters in eukaryotes are divided into subfamilies ABCA through ABCH. In Arabidopsis thaliana L., 129 ABC transporters of ABA have been found [67]. It is reported that several ABCG members participate in ABA transportation. ABA is exported from vascular tissue to various plant parts by the ATP-binding cassette (ABC) transporter gene AtABCG25, which is found in the plasma membrane of the root vascular bundles and near the veins [68,69]. By restricting ABA uptake, the transporter AtABCG40, which facilitates ABA import into plant cells, slows stomatal closure when it is lost in guard cells [67]. By encouraging ABA absorption, the transporter AtABCG22 may assist in controlling stomatal function [70].
In Arabidopsis thaliana L., the import and export of ABA play crucial roles in regulating seed germination and stomatal movement. AtABCG30 and AtABCG40 are responsible for importing ABA into the embryo, which regulates seed germination. On the other hand, AtABCG31 and AtABCG25 control the export of ABA from the endosperm to various locations in the plant. As part of the ABA efflux process, AtDTX50, a DTX/MATE-type transporter, is found in guard cells and vascular tissue. When exogenous ABA is administered, it leads to an increase in the expression of the AtDTX50 gene, contributing to the regulation of ABA levels [71,72]. Recently, Shimizu et al. [73] have revealed two Arabidopsis transporters, NPF4.6 and NPF5.1, which act as ABA importers. These transporters are crucial in regulating stomatal movement by controlling the absorption of ABA in guard cells, vascular tissue, and leaf cells. According to studies, smaller concentrations of HM contamination cause ABA levels inside plant cells to rise. ABA is thus continually distributed throughout the plant, controlling its defensive reactions. However, due to obstructions in the vascular transportation pathway, ABA production and transport diminish under severe HM stress. In these circumstances, exogenous ABA administration can reduce these disturbances.
5. Signaling of Abscisic Acid
The ABA mechanism action is crucial for plants to increase their biological activity in unfavorable environmental conditions [74]. Sucrose non-fermenting (SNF1) related protein phosphatase 2C (PP2C; negative regulators), protein kinase 2 (SnRK2; positive regulators), and pyrabactin resistance (PYR)/pyrabactin resistance-like (PYL)/regulatory component ABA receptor (RCAR) are three crucial components of the ABA signaling process [75]. PYR/PYL/RCARPP2C begins to create a complex structure in the presence of ABA that inhibits the negative effects of PP2C. Additionally, SnRK2 activators promote the ABA-responsive gene expression by phosphorylating downstream proteins, such as transcription factors [76].
The ABA receptors PYL/RCAR/PYR, the protein kinase SnRK2, and the phosphatases PP2Cs make up the fundamental elements of the ABA signaling pathway. RCAR1-14 is an ABA receptor regulatory component, while PYR1 and PYL1-13 are ABA receptors [77]. Phosphatases PP2Cs are monomeric group Thr/Ser phosphatases whose activity is dependent on Mn2+ and Mg2+ [78]. There are ten types (A–J) of Arabidopsis PP2Cs, and the majority of the A-type PP2Cs are engaged in ABA signaling [79]. There are 38 SnRKs in the Arabidopsis genome, which are classified into SnRK3 (3.1–3.25) groups, SnRK2 (2.1–2.10), and SnRK1 (1.1–1.3). A family of ABA-activated protein kinases called SnRK2 is involved in the signaling of HM stress [79]. The SnRK2 family’s ten members can be further classified into three categories: 1, 2, and 3. ABA had no effect on Group 1 (SnRK2.1, SnRK2.4, SnRK2.5, and SnRK2.9), Group 2 (SnRK2.7 and SnRK2.8), and Group 3 (SnRK2.2, SnRK2.3, and SnRK2.6) [79]. The primary positive regulator of ABA signaling was found to be Group 3. One of them, SnRK2.6, is strongly related to the opening and closing of plant stomata, and Arabidopsis SnRK2.6 deletion mutants have problems with ABA-induced stomatal closure [79].
Type PP2Cs act as potent inhibitors of SnRK2 [77]. At this stage, the SnRK2 kinase remains inactive, and the transcription factors responsible for controlling ABA-responsive genes cannot trigger the pathway. However, during stressful conditions, the ABA concentration increases due to elevated ABA production and slows down the degradation or release of the conjugated ABA-GE form [8]. The activation of SnRK2 enables it to undergo autophosphorylation and subsequently activate downstream effectors, like transcription factors and ion channels [79]. These downstream effectors then initiate ABA-responsive gene transcription, contributing to the plant’s response and adaptation to the stressful environment. Plant cells close their stomata in response to HM stress, which is accomplished through ABA-mediated signal transduction. The guard cells integrate endogenous hormonal impulses and external inputs to control the stomatal pore openings [80]. A complicated signaling pathway, including secondary messengers, kinase/phosphatase, and ion channel control, is activated in guard cells by the stress hormone ABA and environmental signals (such as CO2) [80]. Stomatal aperture size has been linked to ABA via Cd2+-dependent and -independent mechanisms [61]. To close the stomata, the guard cells’ intracellular Cd2+ concentration must rise in the Cd2+-dependent pathway. ABA activates plasma-membrane-bound respiratory burst oxidase homolog (RBOH), which then catalyzes reactive oxygen species (ROS) generation by extracellular superoxide dismutases (SOD) [81]. ABA causes PYL/PYR/ RCAR to derepress PP2Cs on the protein kinase OST1/SnRK2.6 [82]. Additionally, calcineurin B subunit-like proteins (CBLs) interact with and control the activity of CBL-interacting protein kinases when they are Cd2+-bound [81]. The RBOHF protein’s N-terminal interacts with the CBL1/CBL9-CIPK26 complex, which phosphorylates RBOHF and increases the production of ROS through RBOHF [81]. Cd2+ channels can be further encouraged to open by ROS, especially H2O2, raising the Cd2+ content in the guard cells [61,81]. Numerous Cd2+ sensors detect changes in the concentration of Cd2+, such as calcium-dependent protein kinase. These channels may be phosphorylated and activated, such as slow anion channel associated 1 (SLAC1) and slow anion channel 3 [61,83,84], which ultimately causes stomatal closure. Activated SnRK2.6/OST1 interacts and phosphorylates SLAC1 and the quick activating anion channel (QUAC1) directly in the Cd2+-independent pathway [81,85]. The defense of the cell’s fast anion efflux activates SLAC1 and QUAC1 channels, depolarizing the plasma membrane and causing K+ efflux and solute release from the guard cells [85], which finally causes stomatal closure. It is simple to discover that the protein kinase SnRK2.6 does play a significant role in the process of stomatal closure by contrasting the Cd2+-dependent and -independent pathways, which is compatible with the earlier assertion. Figure 3 depicts the different phytohormone signaling genes under HM stress.
6. Regulation of ABA-Responsive Detoxification of Heavy Metals
ABA, a well-known stress hormone, controls an extensive variety of physiological functions and aids plants in overcoming the negative effects of HM stress [86]. The movement of hazardous metal from roots to shoots is controlled by ABA. It limits the long-distance transmission of HMs by closing stomata and slowing transpiration [40]. The effects of HM stress are lessened by ABA’s enhancement of plant physiological functions, biomass, osmolyte accumulation, and the antioxidant defense system. Under Cd stress, gene activation linked to ABA production raises endogenous ABA concentration in some plant species, including Oryza sativa L., Triticum aestivum L., Brassica napus L., Solanum tuberosum L., and others [40,86,87,88]. By lowering the Cd concentration and transpiration rate, exogenous ABA supplementation enhances the tolerance of Cd in the seedlings of Oryza sativa L. [87].
In Arabidopsis, IRT1 and its homologs ZIP1 and ZIP4 were expressed more frequently when endogenous ABA content was lower [89], although ABA is able to prevent Cd absorption and accumulation facilitated by IRT1, HIPP22, and HIPP44 at transcriptional levels [90]. According to Rogers et al. [91] and Vert et al. [92], IRT1 is a ZIP family member and uses substrates made of Fe, Zn, Co, Mn, and Cd. With the exception of Rhodophyta Porphyrayezoenesis and Chlodophyta Volvox carteri, homologs of ZIPs were found in almost all of the investigated algae and terrestrial plants. There are 17 ZIPs in Arabidopsis, and only angiosperms have members that resemble IRT1 [93]. For instance, four and five ZIPs were identified from P. patens and M. polymorpha, respectively. They were grouped into two subgroups: in Arabidopsis ZIP2-like subgroup carrying Zn, Fe, and Mn but not Cd, and the IRT3-like group expressing Fe/Zn transporters. While MYB49 directly controls the expression of HIPP22 and HIPP44, activating IRT1 by MYB49 necessitates Ib subgroup members of bHLHs to serve as bridge regulators [90]. Chlorophyta is where bHLHs first appeared, while HIPPs and MYBs seem to be unique to terrestrial plants.
In Arabidopsis, uptake of metalloid As facilitated by PHT1;1 can be regulated by transcriptional factor WRKY6, but MYB protein OsARM1 represses the uptake and distribution of As(III) facilitated by NIPs, and its expression is downregulated by ABA [94]. As(V) and phosphate (P), a macroelement required for all living things, are both substrates for PHTs [95]. Porphyray ezoenesis, Klebsormidium flaccidum, and Chlorophyta Ostreococcus sp. all have 4, 8, and 12 PHT orthologues, respectively, but the numbers quickly rise to 25 in the moss Physcomitrella patens, 19 in Arabidopsis, and 26 in Oryza sativa. Additionally, all terrestrial plants exhibit substantial PHT protein similarity (about 60%). With the exception of Rhodophyta, transcription factors WRKYs are often common in all the species analyzed [96,97]. Four NPF4s are ABA-importing transporters, whereas two NPF7s indirectly contribute to ABA-altered Cd distribution in shoots and roots [97,98].
The ubiquitous short peptide PCs, which contain thiols, shield cells from the toxicity of metalloids and HMs [99]. PCS catalyzes the synthesis of PCs using reduced GSH and similar thiols as substrates [100]. The ancestor of GSH genes is the streptophyte algae Klebsormidium flaccidum, which is also present in the unicellular red algae Cyanidioschyzon merolae of the phylum Rhodophyta. There is a 60% similarity between members of monocots and dicots. The liverwort Marchantia polymorpha, Spirodelapolyrhiza, Klebsormidium laccidum, and the majority of vascular plants all contain the possible PCS orthologs. However, different types of plants have different PCS behaviors and responses to different metals. In comparison to Arabidopsis PCS, basal plant PCSs generally seem to be less active [101,102]. AtZAT6 and StbZIP, which are ABA-induced ethylene transcriptional factors, might promote GSH and PCS expression, while AtWRKY12 could repress it [88,103,104]. The WRKY family may have arisen from Chlorophyta, but the ethylene member family exhibits the same evolutionary history as that of GSHs. In response to exogenous ABA, transcriptional factors from the bZIP subfamily activate PCS in Gray Poplar and Solanum tuberosum, which is consistent with the parallel evolution of PCSs and bZIPs from Streptophyte algae [88]. Additional research must be done to determine whether these regulatory modules are conserved in various plant species.
Exogenous ABA treatment also decreased internal malondialdehyde concentration and Cd accumulation while increasing fresh weight in Brassica napus L., which in turn lessened the negative consequences of Cd stress [105]. Stroinski et al. [88] found that with 0.1 mM ABA, Solanum tuberosum L. seedlings had higher levels of StPCS1, 9-cis-epoxy carotenoid dioxygenase 1, and basic leucine zipper expression, endogenous ABA, and phytochelatin synthase activity in roots. Fan et al. [106] and Pan et al. [107] found that ABA decreased Cd absorption in Arabidopsis thaliana L. by blocking the iron-regulated transporter 1 (IRT1) mechanism as well as reducing Cd-induced toxicity. By increasing antioxidant enzyme activity that scavenged H2O2 to stop Cd from entering H2O2-induced Ca2+-permeable channels, ABA reduced 100 M Cd stress in Populus euphratica L. cells [108]. In Solanum photeinocarpum L., ABA administration at 20 M L−1 increased Cd extraction and phytoremediation activity [109]. In the ABA signaling pathway and the transcription control network of MYB in plants, the JrVHAG1 gene performs the function of a Cd stress response regulator [110]. Figure 4 represents the ABA mechanism in mitigating HM stress. 10 M ABA treatment of Vitis vinifera L. seedlings decreased Zn stress by reducing Zn absorption as well as accumulation in roots and boosting the ZIP gene expression and detoxification-related genes in leaves and roots [111]. Several ABA-treated Pb transport and detoxification genes, including ABCG40, NRAMP1.4, FRD3.1, PCS1.1, and ABCC1.1, were expressed more often in Populus alba L. seedlings, according to Shi et al. [112]. Exogenous application of 10 µg L−1 ABA in Lactuca sativa L. seedlings led to a decrease in Cd absorption and alleviated Cd toxicity [113]. In Sedum alfredii L. seedlings, the exogenously added ABA (0.2 mg L−1) reduced 100 μmol/L Cd stress by boosting ABA synthetase activity and endogenous ABA content. To increase Cd resistance, it also encouraged the expression of HMA4 and NAS in shoots and HsfA4c in roots [114]. By boosting endogenous ABA concentration, ascorbate peroxidase (APX) activity, and proline accumulation and reducing lipoxygenase (LOX) activity, exogenous ABA treatment reduced the harm caused by excessive Zn [115]. Table 1 depicts the exogenous use of ABA against different concentrations of HMs and its response. According to Li and Song [115], ABA may also play a role in Arabidopsis thaliana L. upregulation of the APX gene, proline synthesis genes, and downregulation of the LOX gene. By increasing total antioxidant capacity and chlorophyll content in Hylotelephium spectabile L. and Sedum alfredii L., exogenous ABA treatment at 60 and 40 M L−1 attenuated the harmful effects of Cd and Pb [116]. By boosting glutathione levels, non-protein thiol, and cysteine synthesis, as well as phytochelatin production in plants, seed priming with ABA also decreased HM toxicity in plants [117]. By enhancing proline content and antioxidant enzyme activity in Robiniapseudo acacia L., ABA lessened Zn stress damage [118]. ABA also controlled Cd hyperaccumulation in Sedum alfredii by decreasing the expression of aquaporin in roots, the number of xylem vessels in stems, and the size and density of stomata in leaves [119]. According to Chen et al. [120], exogenously applied ABA increased the phytoremediation effectiveness of Sedum alfredii growing in soil contaminated with Cd, Pb, and Zn.
Liao et al. [121] reported that Cd stress disrupted the growth of rapeseed cultivars (Youfei 1 and Xiangyou 787), but exogenous ABA (0.5, 1, 5, or 10 μmol/L) could counteract Cd’s effects in B. napus. Introducing 5 μmol/L ABA increased biomass in Youfei 1, enhancing plant height and root length by 20.48% and 11.46%, respectively. For both cultivars Youfei 1 and Xiangyou 787, 0.5 μmol/L ABA with Cd increased chlorophyll (21.22% and 26.43%), chlorophyll b (8.76% and 10.12%), and carotenoids (20.13% and 28.80%). However, MDA levels increased at 10 μmol/L ABA. Notably, during Cd stress, MDA decreased by 20.11% in Xiangyou 787 and 36.65% in Youfei 1. ABA’s buffering included oxidative stress-induced stomatal closure. Cd stress reduced T-SOD, POD, and CAT, indicating ROS accumulation.
Similarly, Yu et al. [122] reported that exogenous ABA (10 mol/L) boosted antioxidant enzyme and plant antioxidant activity. This study revealed that by enhancing Cd accumulation, encouraging Cd adsorption on the root cell wall, and activating protective systems, ABA could lessen Cd stress. This finding might encourage the use of C. bipinnatus for the remediation of cadmium-contaminated soil.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Exogenous abscisic acid alleviates HM and metalloid toxicity in plants.
| HM and Metalloid Type | Plant Species | Growth Environment | Dose of ABA | Concentration of HMs | Response | Reference |
|---|---|---|---|---|---|---|
| As | Oryza sativa L. | Culture medium | 10 µM | 50 µM | Improved glutathione content, non-protein thiol, osmolyte concentrations, as well as phytochelatins reductase activity | [117] |
| Cd | Brassica campestris L. | Hydroponic | 5 µM | 100 µM L−1 | Cd presence in plant roots leads to increased soluble protein and chlorophyll content, as well as enhanced antioxidant defense systems. | [86] |
| Cd | Lactuca sativa L. | Soil | 5 µM | 10 µM L−1 | The plant exhibited enhancements in biomass, proline content, stomatal conductance, antioxidant enzyme activity, internal CO2 concentration, and soluble protein content. | [123] |
| Cd, Cr, Ni, Hg, Se, Pb, Sn | Fragaria × ananassa | Soil | 40 μmol L−1 | Cr (8.53), Cd (5.16), Se (6.06), Hg (4.55), Ni (2.18), Sn (1.23) Pb (32.16), in mg kg −1 | The addition of exogenous ABA led to a reduction in the translocation of Cr, Hg, Cd, and Sn into Fragaria× ananassa leaves. Moreover, the concurrent increase in antioxidant capacity significantly mitigated the harmful impact of HM stress on the chlorophyll concentration in the leaves of strawberries. | [124] |
| Cd | Brassica napus L. | Hydroponic | 10 µM | 100 µM | Decreased malondialdehyde and accumulation of Cd content and increased plant biomass | [105] |
| Cd | Populus euphratica L. | Culture medium | 5 µM | 100 µM | Antioxidant activities and cell proliferation showed improvement. | [108] |
| Cd | Vigna radiata L. | Seed tray | 10 µM | 100 µM | By inhibiting lipid peroxidation and stimulating antioxidant enzyme activity, the plant’s tolerance was affected | [125] |
| Cd | Perilla frutescens L. | Soil | 5 µM | 10 mg kg−1 | Increased plant antioxidant activities, photosynthetic pigments, and biomass | [126] |
| Co | Solanum lycopersicum L. | Hydroponic | 10 µM | 400 µM L−1 | Decreased translocation of Co from roots to shoots and improved antioxidant enzyme activities and proline content | [82] |
| Ni | Trigonella foenum-graecum L. | Soil | 40 µM | 80 mg kg−1 | Improved secondary metabolites | [127] |
| Pb | Populus alba L. | Soil | 10 µM | 3 mM | Enhanced root biomass, glutathione, ascorbate content, and photosynthetic rate | [112] |
| Zn | Arabidopsis thaliana L. | Culture medium | 15 µM | 200 mg L−1 | Increased activities of antioxidant enzyme, proline accumulation, and ABA endogenous level | [115] |
7. Abscisic Acid Crosstalk with Phytohormones
Auxin and abscisic acid govern several elements of plant development and metabolism, primarily in antagonism to one another [128]. For example, in Arabidopsis thaliana L., the induction of Auxin Response Factor 2 (ARF2) by ABA leads to increased ABA sensitivity in both germination of seeds as well as root growth [129]. The ARF2 mutants exhibit a heightened response to ABA’s effects on auxin distribution and cell division compared to the wild-type plants. When exogenous ABA is applied to the ARF2 mutant, it causes a reduction in root length [130]. ABA restricts primary root growth by inhibiting cell proliferation in the root tips [129]. Additionally, ABA hampers seedling development by enhancing auxin signaling [131]. The gene expression network involved in seed dormancy regulation, mediated by ABA and auxin, includes the transcription factor ABI3. During seed germination, auxin influences the expression of the ABI3 gene by selecting auxin response factors 10 and 16 [132]. The fact that ABA decreases seed germination, while auxin is necessary for seed dormancy, suggests that auxin and ABA interact to affect seed dormancy. By enhancing biological ABA synthesis and activating the ABA response, auxin affects ABA-modulated activities in two different ways. However, in radical protrusion tests, exogenous and endogenous indoleacetic acid (IAA) decreased seed germination, demonstrating that auxin and ABA jointly restrict seed germination [132,133]. In the absence of ABA, equal amounts of IAA did not exert any inhibitory effect on seed germination, underscoring the essential role of ABA in mediating the auxin-induced inhibition of seed germination. In addition, PIN2 (Auxin Efflux Transporter) and AUX1 (Auxin Influx Transporter) levels rose with osmotically generated ABA [134,135]. As a result, auxin and ABA interact to regulate plant growth and development through the auxin response pathway. Gibberellins (GA) and ABA interact negatively to regulate several aspects of plant development, including seed dormancy and maturity, main root growth, and blooming. In contrast to GA, which stimulates seed germination and fosters seedling growth, ABA suppresses seed germination by encouraging seed dormancy [136]. Balanced GA and ABA production and catabolism are necessary to maintain germination and dormancy [137]. Exogenous ABA supply inhibits primary root development in Arabidopsis thaliana L., but GA enhances root growth [138]; this is similar to how ABA and brassinosteroids (BR) work in opposition to one another to influence plant physiological processes. ABA hinders seed germination, while BR encourages it [139]. Additionally, ABA regulates the way BR signaling occurs. BR signaling elements, including BIN2 and BR11 receptors, interact with ABA components like AB11 and ABA12 [140]. Nonetheless, the presence of Cd inhibits the increase in phenolic compounds [141]. On the other hand, the combination of GA3 and ABA hormones with Zn and Pb shows the potential to boost the overall phenolic content. ABA’s involvement in the interaction between cytokinins (CKs) and BR is due to its inhibition of BR production during metal toxicity [142]. However, to fully comprehend the molecular mechanisms governing physiological processes in plants, additional research is necessary to establish the specific connections between ABA and GA as well as ABA and BR. Figure 5 shows the crosstalk of ABA with other phytohormones under HM stress. Table 2 represents ABA and HM crosstalk.
The stress response in plants is modulated by the interplay between CK and ABA. Within the multigenic family of cytokinin oxidase (CKX) responsible for cytokinin breakdown, various members encode the enzyme cytokinin oxidase [143,144]. In Arabidopsis thaliana L., it was observed that the expression of CKX genes is reduced by ABA. Under stressful conditions, cytokinin receptor kinases (AHK2 and AHK3) negatively regulate the gene expression that is associated with osmotic stress. Interestingly, mutants with cytokinin deficiency exhibited higher survival rates [145,146].
Together, ethylene and ABA regulate stomatal closure, with ethylene controlling the rise in ethylene levels in leaves [147] and ABA signaling pathway blockage [148,149]. The ethylene biosynthesis gene (ACS5), which encodes an enzyme for identifying the rate-limiting stage of ethylene production, is suppressed by ABA. Additionally, ABA positively regulates the behavior of the ethylene response factor 11 (ERF11) via the long hypocotyl 5 (HY5) replication factor, a crucial molecular link between ethylene and ABA production that inhibits the expression of the ACS5 gene. For ABA-controlled ethylene biosynthesis to occur, YF-AtERF11 expression must be modulated [150,151]. By raising GSH content, which led to the detoxification of metal as well as scavenging ROS generated by HM-driven ethylene formation, exogenous SA reduced Cd stress in Triticum aestivum L. In wheat seedlings under Cd stress, SA supplementation raised the levels of ABA, which was attributed to de novo ABA production. Additionally, in Triticum aestivum L. under HM stress, the protective mechanism of SA was demonstrated through ABA-controlled SA-mediated alterations in the dehydrin protein concentration [152,153].
Table 2.
Gene and protein interplay between HMs, metalloids, and phytohormones.
| Heavy Metal and Metalloid | Model Plant | Concentration of HMs | Gene Alteration | Plant Response | Reference |
|---|---|---|---|---|---|
| As | Oryza sativa L. | 50 and 25 mmol L− 1 | Proline and antioxidant biosynthesis gene, alkaline phosphatase, phosphatase, H+/ATPase, and ROS | Antioxidative biosynthesis genes upregulated and downregulated ROS biosynthesis genes and regulated H+/ATPase phosphatase | [117] |
| As | Oryza sativa L. | ABA4, PP2C4, PP2C5NCED2, NCED3, bZIP10, 12 | Increased biosynthesis and signaling of ABA in response to As exposure resulted in decreased root growth | [154] | |
| Cd | Sedum alfredii | NCED, ZEP, AAO | Increased endogenous ABA levels, alleviated Cd toxicity | [114] | |
| Cd | Malus hupehensis | 100 μmol L−1 CdSO4 | Biosynthesis genes of ROS | Downregulated MDA and H2O2 expression to decrease the Cd2+ influx rate | [43] |
| Cd | Pisum sativum | 50 µM CdCl2 | Proline biosynthesis gene, PsPDH1, PsP5CS2, PsProT1 and 2 | In the leaves of pea plants, ABA played a regulatory role in Cd toxicity by downregulating the expression of genes while upregulating proline biosynthesis genes | [155] |
| Cd | Vigna radiate | CdCl2 100 μmol L−1 and 50 μmol L−1 | Stress, ABA-responsive genes, proline, and antioxidant biosynthesis gene | Enzymatic antioxidant genes to regulate Cd toxicity | [125] |
| Cd | Sedum alfredii | Cd(NO3)2: 5 and 25 mmol L−1 | Aquaporin genes | In roots, to regulate Cd toxicity, ABA upregulated the SaPIP genes | [119] |
| Cd | Brassica campestris L. | 100 μmol L−1 | ROS-mediated biosynthesis genes for antioxidative biosynthesis, and proline gene | ABA upregulates antioxidative biosynthesis and proline gene expression to regulate the toxicity of Cd | [86] |
| Cd | Oryza sativa L. | NCED4 | Enhanced HM alleviation and ABA biosynthesis | [156] | |
| Co | Solanum lycopersicum L. | 400 μmol L−1 | CAT, APX, POD, SOD | ROS detoxification increased | [82] |
| Cu | Artemisia annua | 40 and 20 mg kg−1 | Antioxidant biosynthesis genes | Upregulated antioxidative biosynthesis gene expression, promoting the homeostasis of ROS mediated by Cu. | [157] |
| Pb | Populus x canescens | - | PCS1.1, NRAMP1.4, FRD3.1, ABCG40, ABCC1.1 | Enhanced antioxidant enzyme activity and improved uptake, transport, and detoxification of Pb | [112] |
| Pb, Cd, and Zn | Hylotelephium spectabile, Sedum alfredii | - | Antioxidative enzymatic genes, stress-responsive genes, proline, and antioxidant biosynthesis genes | antioxidative biosynthesis genes in both plants under HM toxicities and ABA upregulated antioxidative enzymatic genes | [116] |
| Zn | Vitis vinifera | - | ZIP | Regulated the uptake and accumulation of Zn | [111] |
| Zn | Vitis vinifera | - | NRAMP3, YSL, PCR2 | The co-application of Zn and ABA led to increases in the multiple detoxification-related gene expression | [111] |
The jasmonate (JA) signaling system and ABA interact in a complicated way to regulate gene expression and plant defense. The application of exogenous ABA decreases the defense gene transcription triggered by JA. In contrast, overexpression of JA-responsive genes was brought about by mutations in the ABA synthesis genes aba1 and aba2 [158]. In response to both biotic and abiotic stimuli, the regulation of stress-responsive genes involves a complex interplay between ABA and the JA–ethylene signaling pathway [158,159]. The transcription factor MYC2 plays a crucial role in controlling various elements of the JA signaling pathway in Arabidopsis thaliana L. Interestingly, genetically modified plants with overexpressed MYC2 gene exhibit heightened ABA sensitivity and increased activation of ABA-induced genes. Conversely, MYC2 mutants carrying Ds mutations show reduced gene expression and decreased ABA sensitivity [160]. Thus, MYC2 acts as a central integrative hub, managing signals from both JA and ABA in Arabidopsis thaliana L. [161]. Moreover, the collaboration between nitric oxide (NO) and ABA extends to the regulation of several physiological and developmental aspects in plants. These include seed germination, root development, stomatal movement, and antioxidant enzyme activity [162,163]. In guard cells and mesophyll, the increased expression of the 9-cis-epoxy carotenoid dioxygenase gene (SgNCED1) led to elevated levels of H2O2 and NO, along with enhanced antioxidant enzyme activity [164]. In Arabidopsis thaliana L., breaking seed dormancy resulted in rapid ABA depletion due to a swift accumulation of NO, whereas the opposite effect was observed during seed germination [165,166]. Additionally, for the regulation of stomatal movement in guard cells, ABA-induced endogenous H2O2 synthesis promoted NO production [167,168]. Moreover, in Oryza sativa L., the inhibitory effect of ABA on leaf senescence was alleviated by the application of exogenous NO [169]. By enhancing osmolyte production and the antioxidant defense system, ABA and NO alter metabolic plant processes in response to environmental stress [170,171]. NO and ABA may interact during HM stress to reduce HM stress, as toxicity was decreased by NO’s ability to inhibit the As-induced elevation of ABA content in Vicia faba L. [172]. According to Sadeghipour [173], exogenous NO treatment to Vigna unguiculata L. decreased ABA concentration during Pb stress, improving Pb tolerance. According to Wu et al. [135], Mo increased aldehyde oxidase activity, which, in turn, caused ABA production in Triticum aestivum L. Mo-induced oxidative stress was mediated by NO acting downstream of ABA, suggesting an intermediary connection between NO and ABA.
As a result, ABA plays a key role in controlling plant growth and development under stressful environmental conditions by working in concert with other phytohormones. The molecular processes behind the intricate signaling networks of ABA and other hormones require more investigation.
8. Conclusions and Future Scope
The detrimental impact of HMs on plants has garnered significant global attention among plant researchers, as it poses a threat to sustainable agricultural production. The presence of toxic HMs substantially disrupts various biochemical and physiological aspects of plants by influencing the expression of numerous enzymes and interfering with the synthesis of essential biochemical compounds. In response, plants have developed several defense mechanisms to mitigate the harmful effects of HM toxicity. This review demonstrates that ABA serves as a critical signaling hormone under HM stress conditions, affecting not only its own metabolic and signaling pathways but also the activity of other key phytohormones, such as auxins, gibberellins, and ethylene. Exogenous phytohormone treatment effectively reduces the negative impacts of HM and enhances plant morphological traits, physio-biochemical functions, and output. Exogenous ABA application improves phytochelatin synthesis, antioxidant machinery, osmolyte synthesis, and enzymatic activity in plants, which reduces the harmful effects of HMs. It is possible to utilize ABA to increase economically significant crop growth and yield by applying it to the soil as a foliar spray, seed primer, and culture medium. By enhancing plants’ ability to tolerate stress, ABA is essential in reducing the effects of HM stress. Furthermore, gaining a mechanistic understanding of how multiple phytohormones interact would offer valuable insights into comprehending the intricate complexities involved in enhancing HM stress tolerance for sustainable agriculture. The manipulation of phytohormone levels and their responses in specific tissues or organs during critical developmental stages presents an intriguing approach for cultivating HM stress-tolerant plants suitable for modern agriculture. In conclusion, ABA is much more than just a stress hormone; it is one of the key players in a complex hormonal network that allows plants to adapt to increasingly challenging environmental stresses. Looking ahead, exploring further crosstalk mechanisms among ABA and other phytohormones will undoubtedly emerge as a significant and captivating theme in the plant research field.
Author Contributions
A.B.: conceptualization; A.B., K.S. and P.S.: formal analysis and writing—original draft preparation; K.S.: writing—original draft preparation; A.B., P.S., S.P.S. and K.S.: reviewing and editing and finalizing the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Azameti, M.K.; Imoro, A.M. Nanotechnology: A Promising Field in Enhancing Abiotic Stress Tolerance in Plants. Crop Des. 2023, 100037, in press. [Google Scholar] [CrossRef]
- Raj, V.; Chauhan, M.S.; Pal, S.L. Potential of Sugarcane Bagasse in Remediation of Heavy Metals: A Review. Chemosphere 2022, 307, 135825. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Hu, W.; Tian, K.; Huang, B.; Zhao, Y.; Wang, X.; Zhou, Y.; Shi, B.; Kwon, B.O.; Choi, K.; et al. Accumulation and Ecological Risk of Heavy Metals in Soils Along the Coastal Areas of the Bohai Sea and the Yellow Sea: A Comparative Study of China and South Korea. Environ. Int. 2020, 137, 105519. [Google Scholar] [CrossRef]
- Rahman, S.U.; Nawaz, M.F.; Gul, S.; Yasin, G.; Hussain, B.; Li, Y.; Cheng, H. State-of-the-Art OMICS Strategies against Toxic Effects of Heavy Metals in Plants: A Review. Ecotoxicol. Environ. Saf. 2022, 242, 113952. [Google Scholar] [CrossRef]
- Ahmad, W.; Najeeb, U.; Zia, M.H. Soil Contamination with Metals: Sources, Types and Implications. In Soil Remediation and Plants. Prospects and Challenges; Hakeem, K.R., Sabir, M., Öztürk, M., Mermut, A.R., Eds.; Academic Press: London, UK, 2015; pp. 37–56. [Google Scholar]
- Sharma, P.; Bano, A.; Singh, S.P.; Sharma, S.; Xia, C.; Nadda, A.K.; Lam, S.S.; Tong, Y.W. Engineered Microbes as Effective Tools for the Remediation of Polyaromatic Aromatic Hydrocarbons and Heavy Metals. Chemosphere 2022, 306, 135538. [Google Scholar] [CrossRef]
- Asgari Lajayer, B.A.; Ghorbanpour, M.; Nikabadi, S. Heavy Metals in Contaminated Environment: Destiny of Secondary Metabolite Biosynthesis, Oxidative Status and Phytoextraction in Medicinal Plants. Ecotoxicol. Environ. Saf. 2017, 145, 377–390. [Google Scholar] [CrossRef]
- Sharma, P.; Bano, A.; Yadav, S.; Singh, S.P. Biocatalytic Degradation of Emerging Micropollutants. Top. Catal. 2023, 66, 676–690. [Google Scholar] [CrossRef]
- Raklami, A.; Meddich, A.; Oufdou, K.; Baslam, M. Plants—Microorganisms-Based Bioremediation for Heavy Metal Cleanup: Recent Developments, Phytoremediation Techniques, Regulation Mechanisms, and Molecular Responses. Int. J. Mol. Sci. 2022, 23, 5031. [Google Scholar] [CrossRef]
- Sikdar, A.; Jeyasundar, P.G.S.A.; Debnath, B.; Hossain, M.S.; Islam, M.A.; Ahammed, G.J. Cadmium Contamination in the Soil Environment: Impact on Plant Growth and Human Health. In Agrochemicals in Soil and Environment: Impacts and Remediation; Springer Nature: Singapore, 2022; pp. 367–408. [Google Scholar]
- Bücker-Neto, L.; Paiva, A.L.S.; Machado, R.D.; Arenhart, R.A.; Margis-Pinheiro, M. Interactions Between Plant Hormones and Heavy Metals Responses. Genet. Mol. Biol. 2017, 40 (Suppl. S1), 373–386. [Google Scholar] [CrossRef]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Heavy Metal Stress and Some Mechanisms of Plant Defense Response. Sci. World J. 2015, 2015, 756120. [Google Scholar] [CrossRef]
- Sytar, O.; Kumari, P.; Yadav, S.; Brestic, M.; Rastogi, A. Phytohormone Priming: Regulator for Heavy Metal Stress in Plants. J. Plant Growth Regul. 2019, 38, 739–752. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Adrees, M.; Rizvi, H.; Zia-Ur-Rehman, M.; Hannan, F.; Qayyum, M.F.; Hafeez, F.; Ok, Y.S. Cadmium Stress in Rice: Toxic Effects, Tolerance Mechanisms, and Management: A Critical Review. Environ. Sci. Pollut. Res. Int. 2016, 23, 17859–17879. [Google Scholar] [CrossRef] [PubMed]
- Jalmi, S.K.; Bhagat, P.K.; Verma, D.; Noryang, S.; Tayyeba, S.; Singh, K.; Sharma, D.; Sinha, A.K. Traversing the Links Between Heavy Metal Stress and Plant Signaling. Front. Plant Sci. 2018, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, W.; Song, W.; Guo, M. Remediation Techniques for Heavy Metal-Contaminated Soils: Principles and Applicability. Sci. Total Environ. 2018, 633, 206–219. [Google Scholar] [CrossRef] [PubMed]
- Muthusaravanan, S.; Sivarajasekar, N.; Vivek, J.S.; Paramasivan, T.; Naushad, M.; Prakashmaran, J.; Gayathri, V.; Al-Duaij, O.K. Phytoremediation of Heavy Metals: Mechanisms, Methods and Enhancements. Environ. Chem. Lett. 2018, 16, 1339–1359. [Google Scholar] [CrossRef]
- Sharma, G.; Naushad, M. Adsorptive Removal of Noxious Cadmium Ions from Aqueous Medium Using Activated Carbon/Zirconium Oxide Composite: Isotherm and Kinetic Modelling. J. Mol. Liq. 2020, 310, 113025. [Google Scholar] [CrossRef]
- Bashir, A.; Rizwan, M.; Zia Ur Rehman, M.Z.; Zubair, M.; Riaz, M.; Qayyum, M.F.; Alharby, H.F.; Bamagoos, A.A.; Ali, S. Application of co-Composted Farm Manure and Biochar Increased the Wheat Growth and Decreased Cadmium Accumulation in Plants under Different Water Regimes. Chemosphere 2020, 246, 125809. [Google Scholar] [CrossRef]
- Hussain, A.; Rizwan, M.; Ali, S.; Rehman, M.Z.U.; Qayyum, M.F.; Nawaz, R.; Ahmad, A.; Asrar, M.; Ahmad, S.R.; Alsahli, A.A.; et al. Combined Use of Different Nanoparticles Effectively Decreased Cadmium (Cd) Concentration in Grains of Wheat Grown in a Field Contaminated with Cd. Ecotoxicol. Environ. Saf. 2021, 215, 112139. [Google Scholar] [CrossRef]
- Khan, A.R.; Wakeel, A.; Muhammad, N.; Liu, B.; Wu, M.; Liu, Y.; Ali, I.; Zaidi, S.H.R.; Azhar, W.; Song, G.; et al. Involvement of Ethylene Signaling in Zinc Oxide Nanoparticle-Mediated Biochemical Changes in Arabidopsis thaliana Leaves. Environ. Sci. Nano 2019, 6, 341–355. [Google Scholar] [CrossRef]
- Ur Rehman, M.Z.; Waqar, M.; Bashir, S.; Rizwan, M.; Ali, S.; El Baroudy, A.A.E.F.; Khalid, H.; Ayub, M.A.; Usman, M.; Jahan, S. Effect of Biochar and Compost on Cadmium Bioavailability and Its Uptake by Wheat–Rice Cropping System Irrigated with Untreated Sewage Water: A Field Study. Arab. J. Geosci. 2021, 14, 135. [Google Scholar] [CrossRef]
- Thudi, M.; Palakurthi, R.; Schnable, J.C.; Chitikineni, A.; Dreisigacker, S.; Mace, E.; Srivastava, R.K.; Satyavathi, C.T.; Odeny, D.; Tiwari, V.K.; et al. Genomic Resources in Plant Breeding for Sustainable Agriculture. J. Plant Physiol. 2021, 257, 153351. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Asami, T. Chemical Regulators of Plant Hormones and Their Applications in Basic Research and Agriculture. Biosci. Biotechnol. Biochem. 2018, 82, 1265–1300. [Google Scholar] [CrossRef]
- Wani, S.H.; Kumar, V.; Shriram, V.; Sah, S.K. Phytohormones and Their Metabolic Engineering for Abiotic Stress Tolerance in Crop Plants. Crop J. 2016, 4, 162–176. [Google Scholar] [CrossRef]
- Ku, Y.S.; Sintaha, M.; Cheung, M.Y.; Lam, H.M. Plant Hormone Signaling Crosstalks between Biotic and Abiotic Stress Responses. Int. J. Mol. Sci. 2018, 19, 3206. [Google Scholar] [CrossRef] [PubMed]
- Rhaman, M.S.; Imran, S.; Rauf, F.; Khatun, M.; Baskin, C.C.; Murata, Y.; Hasanuzzaman, M. Seed Priming with Phytohormones: An Effective Approach for the Mitigation of Abiotic Stress. Plants 2020, 10, 37. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy Metal Tolerance in Plants: Role of Transcriptomics, Proteomics, Metabolomics, and Ionomics. Front. Plant Sci. 2015, 6, 1143. [Google Scholar] [CrossRef]
- Hossain, A.; Pamanick, B.; Venugopalan, V.K.; Ibrahimova, U.; Rahman, M.A.; Siyal, A.L.; Maitra, S.; Chatterjee, S.; Aftab, T. Emerging Roles of Plant Growth Regulators for Plants Adaptation to Abiotic Stress–Induced Oxidative Stress. In Emerging Plant Growth Regulators in Agriculture; Academic Press: Cambridge, MA, USA, 2022; pp. 1–72. [Google Scholar]
- Sonkar, S.; Sharma, L.; Singh, R.K.; Pandey, B.; Rathore, S.S.; Singh, A.K.; Porwal, P.; Singh, S.P.; Singh, S.P. Plant Stress Hormones Nanobiotechnology. In Nanobiotechnology: Mitigation of Abiotic Stress in Plants; Springer: Cham, Switzerland, 2021; pp. 349–373. [Google Scholar]
- Addicott, F.T.; Lyon, J.L. Physiology of Abscisic Acid and Related Substances. Annu. Rev. Plant Physiol. 1969, 20, 139–164. [Google Scholar] [CrossRef]
- Skubacz, A.; Daszkowska-Golec, A. Seed Dormancy: The Complex Process Regulated by Abscisic Acid, Gibberellins, and Other Phytohormones That Makes Seed Germination Work. In Phytohormones-Signaling Mechanisms and CrossTalk in Plant Development and Stress Responses; IntechOpen Limited: London, UK, 2017. [Google Scholar]
- Finkelstein, R. Abscisic Acid Synthesis and Response. Arabidopsis Book. Am. Soc. Plant Biol. 2013, 11, e0166. [Google Scholar] [CrossRef]
- Boursiac, Y.; Léran, S.; Corratgé-Faillie, C.; Gojon, A.; Krouk, G.; Lacombe, B. ABA Transport and Transporters. Trends Plant Sci. 2013, 18, 325–333. [Google Scholar] [CrossRef]
- Addicott, F.T. Plant Hormones in the Control of Abscission. Biol. Rev. 1970, 45, 485–524. [Google Scholar] [CrossRef]
- Cracker, L.E.; Abeles, F.B. Abscission: Role of Abscisic Acid. Plant Physiol. 1969, 44, 1144–1149. [Google Scholar] [CrossRef]
- Ogawa, M.; Kay, P.; Wilson, S.; Swain, S.M. Arabidopsis Dehiscence Zone Polygalacturonase1 (ADPG1), ADPG2, and QUARTET2 Are Polygalacturonases Required for Cell Separation during Reproductive Development in Arabidopsis. Plant Cell 2009, 21, 216–233. [Google Scholar] [CrossRef] [PubMed]
- Osborne, D.J.; Morgan, P.W. Abscission. Crit. Rev. Plant Sci. 1989, 8, 103–129. [Google Scholar] [CrossRef]
- Djilianov, D.L.; Dobrev, P.I.; Moyankova, D.P.; Vankova, R.; Georgieva, D.T.; Gajdošová, S.; Motyka, V. Dynamics of Endogenous Phytohormones During Desiccation and Recovery of the Resurrection Plant Species Haberlea Rhodopensis. J. Plant Growth Regul. 2013, 32, 564–574. [Google Scholar] [CrossRef]
- Hu, B.; Deng, F.; Chen, G.; Chen, X.; Gao, W.; Long, L.; Xia, J.; Chen, Z.H. Evolution of Abscisic Acid Signaling for Stress Responses to Toxic Metals and Metalloids. Front. Plant Sci. 2020, 11, 909. [Google Scholar] [CrossRef]
- Kumar, S.; Shah, S.H.; Vimala, Y.; Jatav, H.S.; Ahmad, P.; Chen, Y.; Siddique, K.H.M. Abscisic Acid: Metabolism, Transport, Crosstalk with Other Plant Growth Regulators, and Its Role in Heavy Metal Stress Mitigation. Front. Plant Sci. 2022, 13, 972856. [Google Scholar] [CrossRef]
- Gonzalez-Villagra, J.; Figueroa, C.; Luengo-Escobar, A.; Morales, M.; Inostroza-Blancheteau, C.; Reyes-Díaz, M. Abscisic Acid and Plant Response under Adverse Environmental Conditions. In Plant Performance under Environmental Stress; Springer Nature: Berlin/Heidelberg, Germany, 2021; pp. 17–47. [Google Scholar]
- Deng, B.; Zhang, W.; Yang, H. Abscisic Acid Decreases Cell Death in Malus hupehensis Rehd. Under Cd Stress by Reducing Root Cd2+ Influx and Leaf Transpiration. J. Plant Growth Regul. 2022, 41, 639–646. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, Z.; Song, J.; Yue, S. CD2+ Uptake Inhibited by MhNCED3 from Malus hupehensis Alleviates Cd-Induced Cell Death. Environ. Exp. Bot. 2019, 166, 103802. [Google Scholar] [CrossRef]
- Humplík, J.F.; Bergougnoux, V.; Van Volkenburgh, E. To Stimulate or Inhibit? That Is the Question for the Function of Abscisic Acid. Trends Plant Sci. 2017, 22, 830–841. [Google Scholar] [CrossRef]
- Asgher, M.; Rehaman, A.; Islam, S.N.U.; Arshad, M.; Khan, N.A. Appraisal of Functions and Role of Selenium in Heavy Metal Stress Adaptation in Plants. Agriculture 2023, 13, 1083. [Google Scholar] [CrossRef]
- Tao, Q.; Jupa, R.; Liu, Y.; Luo, J.; Li, J.; Kováč, J.; Li, B.; Li, Q.; Wu, K.; Liang, Y.; et al. Abscisic Acid-Mediated Modifications of Radial Apoplastic Transport Pathway Play a Key Role in Cadmium Uptake in Hyperaccumulator Sedum alfredii. Plant Cell Environ. 2019, 42, 1425–1440. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.L.; Lei, X.J.; Wang, Y.Y.; Liu, B.C.; Wang, D.N.; Liu, Z.Y.; Gao, C.Q. Transcriptomic Analysis of Cadmium Stressed Tamarix hispida Revealed Novel Transcripts and the Importance of Abscisic Acid Network. Front. Plant Sci. 2022, 13, 843725. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Zhang, J.; Nageswaran, D.; Li, L. Carotenoid Metabolism and Regulation in Horticultural Crops. Hortic. Res. 2015, 2, 15036. [Google Scholar] [CrossRef] [PubMed]
- Pu, X.; Dong, X.; Li, Q.; Chen, Z.; Liu, L. An Update on the Function and Regulation of Methylerythritol Phosphate and Mevalonate Pathways and Their Evolutionary Dynamics. J. Integr. Plant Biol. 2021, 63, 1211–1226. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Chang, W.C.; Xiao, Y.; Liu, H.W.; Liu, P. Methylerythritol Phosphate Pathway of Isoprenoid Biosynthesis. Annu. Rev. Biochem. 2013, 82, 497–530. [Google Scholar] [CrossRef] [PubMed]
- Vranová, E.; Coman, D.; Gruissem, W. Network Analysis of the MVA and MEP Pathways for Isoprenoid Synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef]
- Frank, A.; Groll, M. The Methylerythritol Phosphate Pathway to Isoprenoids. Chem. Rev. 2017, 117, 5675–5703. [Google Scholar] [CrossRef]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid Metabolism in Plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef]
- Hirschberg, J. Carotenoid Biosynthesis in Flowering Plants. Curr. Opin. Plant Biol. 2001, 4, 210–218. [Google Scholar] [CrossRef]
- Nambara, E.; Marion-Poll, A. Abscisic Acid Biosynthesis and Catabolism. Annu. Rev. Plant Biol. 2005, 56, 165–185. [Google Scholar] [CrossRef]
- Schwartz, S.H.; Qin, X.; Zeevaart, J.A.D. Elucidation of the Indirect Pathway of Abscisic Acid Biosynthesis by Mutants, Genes, and Enzymes. Plant Physiol. 2003, 131, 1591–1601. [Google Scholar] [CrossRef] [PubMed]
- Jia, K.P.; Mi, J.; Ali, S.; Ohyanagi, H.; Moreno, J.C.; Ablazov, A.; Balakrishna, A.; Berqdar, L.; Fiore, A.; Diretto, G.; et al. An Alternative, Zeaxanthin Epoxidase-Independent Abscisic Acid Biosynthetic Pathway in Plants. Mol. Plant 2022, 15, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Dong, T.; Park, Y.; Hwang, I. Abscisic Acid: Biosynthesis, Inactivation, Homoeostasis and Signalling. Essays Biochem. 2015, 58, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, M.; Todoroki, Y. ABA 8′-Hydroxylase and Its Chemical Inhibitors. Phytochem. Rev. 2006, 5, 385–404. [Google Scholar] [CrossRef]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic Acid Dynamics, Signaling, and Functions in Plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [PubMed]
- Wahab, A.; Abdi, G.; Saleem, M.H.; Ali, B.; Ullah, S.; Shah, W.; Mumtaz, S.; Yasin, G.; Muresan, C.C.; Marc, R.A. Plants’ Physio-Biochemical and Phyto-hormonal Responses to Alleviate the Adverse Effects of Drought Stress: A Comprehensive Review. Plants 2022, 11, 1620. [Google Scholar] [CrossRef]
- Kang, J. ABA Transport Assay in Plant Single-Cell System. In Abscisic Acid: Methods and Protocols; Humana: New York, NY, USA, 2022; pp. 71–84. [Google Scholar] [CrossRef]
- Kanno, Y.; Hanada, A.; Chiba, Y.; Ichikawa, T.; Nakazawa, M.; Matsui, M.; Koshiba, T.; Kamiya, Y.; Seo, M. Identification of an Abscisic Acid Transporter by Functional Screening Using the Receptor Complex as a Sensor. Proc. Natl. Acad. Sci. USA 2012, 109, 9653–9658. [Google Scholar] [CrossRef]
- Merilo, E.; Jalakas, P.; Laanemets, K.; Mohammadi, O.; Hõrak, H.; Kollist, H.; Brosché, M. Abscisic Acid Transport and Homeostasis in the Context of Stomatal Regulation. Mol. Plant. 2015, 8, 1321–1333. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, H.; Pan, Y.; Yu, Y.; Luan, S.; Li, L. A DTX/MATEtype Transporter Facilitates Abscisic Acid Efflux and Modulates ABA Sensitivity and Drought Tolerance in Arabidopsis. Mol. Plant. 2014, 7, 1522–1532. [Google Scholar] [CrossRef]
- Kang, J.; Hwang, J.U.; Lee, M.; Kim, Y.Y.; Assmann, S.M.; Martinoia, E.; Lee, Y. PDR-Type ABC Transporter Mediates Cellular Uptake of the Phytohormone Abscisic Acid. Proc. Natl. Acad. Sci. USA 2010, 107, 2355–2360. [Google Scholar] [CrossRef]
- Kuromori, T.; Miyaji, T.; Yabuuchi, H.; Shimizu, H.; Sugimoto, E.; Kamiya, A.; Moriyama, Y.; Shinozaki, K. ABC Transporter AtABCG25 Is Involved in Abscisic Acid Transport and Responses. Proc. Natl. Acad. Sci. USA 2010, 107, 2361–2366. [Google Scholar] [CrossRef]
- Andolfo, G.; Ruocco, M.; Di Donato, A.; Frusciante, L.; Lorito, M.; Scala, F.; Ercolano, M.R. Genetic Variability and Evolutionary Diversification of Membrane ABC Transporters in Plants. BMC Plant Biol. 2015, 15, 51. [Google Scholar] [CrossRef]
- Kuromori, T.; Sugimoto, E.; Shinozaki, K. Arabidopsis Mutants of AtABCG22, an ABC Transporter Gene, Increase Water Transpiration and Drought Susceptibility. Plant J. 2011, 67, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Dahuja, A.; Kumar, R.R.; Sakhare, A.; Watts, A.; Singh, B.; Goswami, S.; Sachdev, A.; Praveen, S. Role of ATP-binding cassette transporters in maintaining plant homeostasis under abiotic and biotic stresses. Physiol. Plantarum. 2021, 171, 785–801. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Fang, C.; Li, Z.; Wang, Y.; Pan, S.; Wu, Y.; An, X.; Long, Y.; Wan, X. ATP-Binding Cassette G Transporters and Their Multiple Roles Especially for Male Fertility in Arabidopsis, Rice and Maize. Int. J. Mol. Sci 2022, 23, 9304. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Kanno, Y.; Suzuki, H.; Watanabe, S.; Seo, M. Arabidopsis NPF4. 6 and NPF5. 1 Control Leaf Stomatal Aperture by Regulating Abscisic Acid Transport. Genes 2021, 12, 885. [Google Scholar] [CrossRef]
- Ali, A.; Pardo, J.M.; Yun, D.J. Desensitization of ABA-Signaling: The Swing from Activation to Degradation. Front. Plant Sci. 2020, 11, 379. [Google Scholar] [CrossRef]
- Ye, Y.; Zhou, L.; Liu, X.; Liu, H.; Li, D.; Cao, M.; Chen, H.; Xu, L.; Zhu, J.K.; Zhao, Y. A Novel Chemical Inhibitor of ABA Signaling Targets All ABA Receptors. Plant Physiol. 2017, 173, 2356–2369. [Google Scholar] [CrossRef]
- Sun, X.; Wang, S.; Tian, J.; Xiang, X.; Zheng, H.; Liu, H.; Fang, Z.; Tian, Z.; Liu, L.; Zhu, Y.; et al. Synergistic Interplay between ABA-Generating Bacteria and Biochar in the Reduction of Heavy Metal Accumulation in Radish, Pakchoi, and Tomato. Environ. Pollut. 2023, 333, 122084. [Google Scholar] [CrossRef]
- Rodriguez, P.L.; Lozano-Juste, J.; Albert, A. PYR/PYL/RCAR ABA Receptors. In Advances in Botanical Research; Elsevier Inc.: Amsterdam, The Netherlands, 2019; Volume 92, pp. 51–82. [Google Scholar]
- Yang, Q.; Liu, K.; Niu, X.; Wang, Q.; Wan, Y.; Yang, F.; Li, G.; Wang, Y.; Wang, R. Genome-Wide Identification of PP2C Genes and Their Expression Profiling in Response to Drought and Cold Stresses in Medicago truncatula. Sci. Rep. 2018, 8, 12841. [Google Scholar] [CrossRef]
- Ng, L.M.; Melcher, K.; Teh, B.T.; Xu, H.E. Abscisic Acid Perception and Signaling: Structural Mechanisms and Applications. Acta Pharmacol. Sin. 2014, 35, 567–584. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Böhmer, M.; Hu, H.; Nishimura, N.; Schroeder, J.I. Guard Cell Signal Transduction Network: Advances in Understanding Abscisic Acid, CO2, and Ca2+ Signaling. Annu. Rev. Plant Biol. 2010, 61, 561–591. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Blumwald, E. The Roles of ROS and ABA in Systemic Acquired Acclimation. Plant Cell 2015, 27, 64–70. [Google Scholar] [CrossRef]
- Kamran, M.; Danish, M.; Saleem, M.H.; Malik, Z.; Parveen, A.; Abbasi, G.H.; Jamil, M.; Ali, S.; Afzal, S.; Riaz, M.; et al. Application of Abscisic Acid and 6-Benzylaminopurine Modulated Morpho-physiological and Antioxidative Defense Responses of Tomato (Solanum lycopersicum L.) by Minimizing Cobalt Uptake. Chemosphere 2021, 263, 128169. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, Y.J.; Kim, M.H.; Kwak, J.M. MAPK Cascades in Guard Cell Signal Transduction. Front. Plant Sci. 2016, 7, 80. [Google Scholar] [CrossRef] [PubMed]
- Brandt, B.; Munemasa, S.; Wang, C.; Nguyen, D.; Yong, T.; Yang, P.G.; Poretsky, E.; Belknap, T.F.; Waadt, R.; Alemán, F.; et al. Calcium Specificity Signaling Mechanisms in Abscisic Acid Signal Transduction in Arabidopsis Guard Cells. eLife 2015, 4, e03599. [Google Scholar] [CrossRef] [PubMed]
- Pei, D.; Hua, D.; Deng, J.; Wang, Z.; Song, C.; Wang, Y.; Wang, Y.; Qi, J.; Kollist, H.; Yang, S.; et al. Phosphorylation of the Plasma Membrane H+-ATPase AHA2 by BAK1 Is Required for ABA-Induced Stomatal Closure in Arabidopsis. Plant Cell 2022, 34, 2708–2729. [Google Scholar] [CrossRef]
- Shen, G.; Niu, J.; Deng, Z. Abscisic Acid Treatment Alleviates Cadmium Toxicity in Purple Flowering Stalk (Brassica campestris L. ssp. chinensis var. purpurea Hort.) Seedlings. Plant Physiol. Biochem. 2017, 118, 471–478. [Google Scholar] [CrossRef]
- Hsu, Y.T.; Kao, C.H. Role of Abscisic Acid in Cadmium Tolerance of Rice (Oryza sativa L.) Seedlings. Plant Cell Environ. 2003, 26, 867–874. [Google Scholar] [CrossRef]
- Stroinski, A.; Giżewska, K.; Zielezińska, M. Abscisic acid is required in Transduction of Cadmium Signal to Potato Roots. Biol. Plant. 2013, 57, 121–127. [Google Scholar] [CrossRef]
- Lu, Q.; Weng, Y.; You, Y.; Xu, Q.; Li, H.; Li, Y.; Liu, H.; Du, S. Inoculation with Abscisic Acid (ABA)-Catabolizing Bacteria Can Improve Phytoextraction of Heavy Metal in Contaminated Soil. Environ. Pollut. 2020, 257, 113497. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, R.; Ju, Q.; Li, W.; Tran, L.-S.P.; Xu, J. The R2R3- MYB Transcription Factor MYB49 Regulates Cadmium Accumulation. Plant Physiol. 2019, 180, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Rogers, E.E.; Eide, D.J.; Guerinot, M.L. Altered Selectivity in an Arabidopsis Metal Transporter. Proc. Natl. Acad. Sci. USA 2000, 97, 12356–12360. [Google Scholar] [CrossRef] [PubMed]
- Vert, G.; Grotz, N.; Dédaldéchamp, F.; Gaymard, F.; Guerinot, M.L.; Briat, J.F.; Curie, C. IRT1, an Arabidopsis Transporter Essential for Iron Uptake from the Soil and for Plant Growth. Plant Cell 2002, 14, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Lo, J.C.; Tsednee, M.; Lo, Y.C.; Yang, S.C.; Hu, J.M.; Ishizaki, K.; Kohchi, T.; Lee, D.C.; Yeh, K.C. Evolutionary Analysis of Iron (Fe) Acquisition System in Marchantia polymorpha. New Phytol. 2016, 211, 569–583. [Google Scholar] [CrossRef]
- Wang, F.-Z.; Chen, M.-X.; Yu, L.-J.; Xie, L.-J.; Yuan, L.-B.; Qi, H.; Xiao, M.; Guo, W.; Chen, Z.; Yi, K.; et al. OsARM1, an R2R3 MYB transcription factor, is involved in regulation of the response to arsenic stress in rice. Front. Plant Sci. 2017, 8, 1868. [Google Scholar] [CrossRef]
- Lindsay, E.R.; Maathuis, F.J.M. New Molecular Mechanisms to Reduce Arsenic in Crops. Trends Plant Sci. 2017, 22, 1016–1026. [Google Scholar] [CrossRef]
- Pommerrenig, B.; Diehn, T.A.; Bernhardt, N.; Bienert, M.D.; Mitani-Ueno, N.; Fuge, J.; Bieber, A.; Spitzer, C.; Bräutigam, A.; Ma, J.F.; et al. Functional Evolution of Nodulin 26-Like Intrinsic Proteins: From Bacterial Arsenic Detoxification to Plant Nutrient Transport. New Phytol. 2020, 225, 1383–1396. [Google Scholar] [CrossRef]
- Léran, S.; Varala, K.; Boyer, J.C.; Chiurazzi, M.; Crawford, N.; Daniel-Vedele, F.; David, L.; Dickstein, R.; Fernandez, E.; Forde, B.; et al. A Unified Nomenclature of nitrate transporter 1/peptide transporter Family Members in Plants. Trends Plant Sci. 2014, 19, 5–9. [Google Scholar] [CrossRef]
- Meng, Y.; Huang, J.; Jing, H.; Wu, Q.; Shen, R.; Zhu, X. Exogenous abscisic acid alleviates Cd toxicity in Arabidopsis thaliana by inhibiting Cd uptake, translocation and accumulation, and promoting Cd chelation and efflux. Plant Sci. 2022, 325, 111464. [Google Scholar] [CrossRef]
- Clemens, S. Evolution and Function of Phytochelatin Synthases. J. Plant Physiol. 2006, 163, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Cobbett, C.S. A Family of Phytochelatin Synthase Genes from Plant, Fungal and Animal Species. Trends Plant Sci. 1999, 4, 335–337. [Google Scholar] [CrossRef] [PubMed]
- Degola, F.; De Benedictis, M.; Petraglia, A.; Massimi, A.; Fattorini, L.; Sorbo, S.; Basile, A.; Sanità di Toppi, L. A Cd/Fe/Zn-Responsive Phytochelatin Synthase Is Constitutively Present in the Ancient Liverwort Lunularia cruciata (L.) Dumort. Plant Cell Physiol. 2014, 55, 1884–1891. [Google Scholar] [CrossRef] [PubMed]
- Petraglia, A.; De Benedictis, M.; Degola, F.; Pastore, G.; Calcagno, M.; Ruotolo, R.; Mengoni, A.; Sanità di Toppi, L. The Capability to Synthesize Phytochelatins and the Presence of Constitutive and Functional Phytochelatin Synthases Are Ancestral (Plesiomorphic) Characters for Basal Land Plants. J. Exp. Bot. 2014, 65, 1153–1163. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Fan, T.; Zhu, X.; Wu, X.; Ouyang, J.; Jiang, L.; Cao, S. WRKY12 Represses GSH1 Expression to Negatively Regulate Cadmium Tolerance in Arabidopsis. Plant Mol. Biol. 2019, 99, 149–159. [Google Scholar] [CrossRef]
- Chen, J.; Yang, L.; Yan, X.; Liu, Y.; Wang, R.; Fan, T.; Ren, Y.; Tang, X.; Xiao, F.; Liu, Y.; et al. Zinc-Finger Transcription Factor ZAT6 Positively Regulates Cadmium Tolerance through the Glutathione-Dependent Pathway in Arabidopsis. Plant Physiol. 2016, 171, 707–719. [Google Scholar] [CrossRef]
- Meng, H.; Hua, S.; Shamsi, I.H.; Jilani, G.; Li, Y.; Jiang, L. Cadmiuminduced Stress on the Seed Germination and Seedling Growth of Brassica napus L., and Its Alleviation through Exogenous Plant Growth Regulators. Plant Growth Regul. 2009, 58, 47–59. [Google Scholar] [CrossRef]
- Fan, S.K.; Fang, X.Z.; Guan, M.Y.; Ye, Y.Q.; Lin, X.Y.; Du, S.T.; Jin, C.W. Exogenous Abscisic Acid Application Decreases Cadmium Accumulation in Arabidopsis Plants, Which Is Associated with the Inhibition of IRT1-Mediated Cadmium Uptake. Front. Plant Sci. 2014, 5, 721. [Google Scholar] [CrossRef]
- Pan, W.; You, Y.; Shentu, J.L.; Weng, Y.N.; Wang, S.T.; Xu, Q.R.; Liu, H.J.; Du, S.T. Abscisic Acid (ABA)-Importing Transporter 1 (AIT1) Contributes to the Inhibition of Cd Accumulation via Exogenous ABA Application in Arabidopsis. J. Hazard. Mater. 2020, 391, 122189. [Google Scholar] [CrossRef]
- Han, Y.; Wang, S.; Zhao, N.; Deng, S.; Zhao, C.; Li, N.; Sun, J.; Zhao, R.; Yi, H.; Shen, X.; et al. Exogenous Abscisic Acid Alleviates Cadmium Toxicity by Restricting Cd2+ Influx in Populus euphratica Cells. J. Plant Growth Regul. 2016, 35, 827–837. [Google Scholar] [CrossRef]
- Wang, J.; Lin, L.; Luo, L.; Liao, M.A.; Lv, X.; Wang, Z.; Liang, D.; Xia, H.; Wang, X.; Lai, Y.; et al. The Effects of Abscisic Acid (ABA) Addition on Cadmium Accumulation of Two Ecotypes of Solanum photeinocarpum. Environ. Monit. Assess. 2016, 188, 182. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Ge, Y.; Zhang, W.; Zhao, Y.; Yang, G. The Walnut JrVHAG1 Gene Is Involved in Cadmium Stress Response through ABA-Signal Pathway and MYB Transcription Regulation. BMC Plant Biol. 2018, 18, 19. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Yan, Y.; Rosado, A.; Zhang, Z.; Castellarin, S.D. ABA Alleviates Uptake and Accumulation of Zinc in Grapevine (Vitis vinifera L.) by Inducing Expression of ZIP and Detoxification-Related Genes. Front. Plant Sci. 2019, 10, 872. [Google Scholar] [CrossRef]
- Shi, W.G.; Liu, W.; Yu, W.; Zhang, Y.; Ding, S.; Li, H.; Mrak, T.; Kraigher, H.; Luo, Z.B. Abscisic Acid Enhances Lead Translocation from the Roots to the Leaves and Alleviates Its Toxicity in Populus × canescens. J. Hazard. Mater. 2019, 362, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Dawuda, M.M.; Liao, W.; Hu, L.; Yu, J.; Xie, J.; Calderón-Urrea, A.; Wu, Y.; Tang, Z. Foliar Application of Abscisic Acid Mitigates Cadmium Stress and Increases Food Safety of Cadmium-Sensitive Lettuce (Lactuca sativa L.) Genotype. PeerJ 2020, 8, e9270. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Chen, S.; Li, Y.; Zheng, F.; He, B.; Gu, M. Exogenous Abscisic Acid (ABA) Promotes Cadmium (Cd) Accumulation in Sedum alfredii Hance by Regulating the Expression of Cd Stress Response Genes. Environ. Sci. Pollut. Res. Int. 2020, 27, 8719–8731. [Google Scholar] [CrossRef] [PubMed]
- Li, X.S.; Song, L.L. The Role of ABA in the Responses of Wildtype and Abscisic Acid Mutants of Arabidopsis thaliana to Excess Zinc. Acta Physiol. Plant. 2020, 42, 74. [Google Scholar] [CrossRef]
- Cheng, L.; Pu, L.; Li, A.; Zhu, X.; Zhao, P.; Xu, X.; Lei, N.; Chen, J. Implication of Exogenous Abscisic Acid (ABA) Application on Phytoremediation: Plants Grown in Co-contaminated Soil. Environ. Sci. Pollut. Res. 2022, 29, 8684–8693. [Google Scholar] [CrossRef]
- Saha, I.; Hasanuzzaman, M.; Adak, M.K. Abscisic Acid Priming Regulates Arsenite Toxicity in Two Contrasting Rice (Oryza sativa L.) Genotypes through Differential Functioning of sub1A Quantitative Trait Loci. Environ. Pollut. 2021, 287, 117586. [Google Scholar] [CrossRef]
- Lou, X.; Zhang, X.; Zhang, Y.; Tang, M. The Synergy of Arbuscular Mycorrhizal Fungi and Exogenous Abscisic Acid Benefits Robinia pseudoacacia L. Growth through Altering the Distribution of Zn and Endogenous Abscisic Acid. J. Fungi 2021, 7, 671. [Google Scholar] [CrossRef]
- Tao, Q.; Jupa, R.; Dong, Q.; Yang, X.; Liu, Y.; Li, B.; Yuan, S.; Yin, J.; Xu, Q.; Li, T.; et al. Abscisic Acidmediated Modifications in Water Transport Continuum Are Involved in Cadmium Hyperaccumulation in Sedum alfredii. Chemosphere 2021, 268, 129339. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Liu, Q.; Chen, S.; Zhang, S.; Wang, M.; Mujtaba Munir, M.A.M.; Feng, Y.; He, Z.; Yang, X. Roles of Exogenous Plant Growth Regulators on Phytoextraction of Cd/Pb/Zn by Sedum alfredii Hance in Contaminated Soils. Environ. Pollut. 2022, 293, 118510. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Tang, Y.; Wang, S.; Su, H.; Chen, J.; Zhang, D.; Wu, J.; Zhou, D.; Yan, M.; Liu, L. Abscisic acid modulates differential physiological and biochemical responses to cadmium stress in Brassica napus. Environ. Pollut. Bioavailab. 2023, 35, 2168216. [Google Scholar] [CrossRef]
- Yu, X.; Yang, L.; Fan, C.; Hu, J.; Zheng, Y.; Wang, Z.; Liu, Y.; Xiao, X.; Yang, L.; Lei, T.; et al. Abscisic acid (ABA) alleviates cadmium toxicity by enhancing the adsorption of cadmium to root cell walls and inducing antioxidant defense system of Cosmos bipinnatus. Ecotoxicol. Environ. Saf. 2023, 261, 115101. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Wang, L.; Xie, Y.; Yu, X.; Li, H.; Lin, L.; Liao, M.; Wang, Z.; Sun, G.; Wang, X.; et al. Effects of Exogenous Abscisic Acid on the Growth and Cadmium Accumulation of Lettuce under Cadmium-Stress Conditions. Int. J. Environ. Anal. Chem. 2020, 100, 720–731. [Google Scholar] [CrossRef]
- Kocaman, A. Effects of Foliar Application of Abscisic Acid on Antioxidant Content, Phytohormones in Strawberry Shoots, and Translocation of Various Heavy Metals. Sci. Hortic. 2023, 314, 111943. [Google Scholar] [CrossRef]
- Leng, Y.; Li, Y.; Ma, Y.H.; He, L.F.; Li, S.W. Abscisic Acid Modulates Differential Physiological and Biochemical Responses of Roots, Stems, and Leaves in Mung Bean Seedlings to Cadmium Stress. Environ. Sci. Pollut. Res. Int. 2021, 28, 6030–6043. [Google Scholar] [CrossRef]
- Shu, J.; He, Q.; Zheng, H.; Liao, M.A.; Kuang, N.; Huang, Z. Abscisic Acid Improves the Ability of Accumulator Plant Perilla frutescens to Perform Phytoremediation in Cadmium-Contaminated Soils. Int. J. Environ. Anal. Chem. 2021, 18, 2182–2191. [Google Scholar] [CrossRef]
- Parwez, R.; Aftab, T.; Khan, M.M.A.; Naeem, M. Exogenous Abscisic Acid Fine-Tunes Heavy Metal Accumulation and Plant’s Antioxidant Defence Mechanism to Optimize Crop Performance and Secondary Metabolite Production in Trigonella foenum-Graecum L. Under Nickel Stress. Plant Sci. 2023, 332, 111703. [Google Scholar] [CrossRef]
- Parwez, R.; Aftab, T.; Gill, S.S.; Naeem, M. Abscisic Acid Signaling and Crosstalk with Phytohormones in Regulation of Environmental Stress Responses. Environ. Exp. Bot. 2022, 199, 104885. [Google Scholar] [CrossRef]
- Wang, L.; Hua, D.; He, J.; Duan, Y.; Chen, Z.; Hong, X.; Gong, Z. Auxin Response Factor2 (ARF2) and Its Regulated Homeodomain Gene HB33 Mediate Abscisic Acid Response in Arabidopsis. PLoS Genet. 2011, 7, e1002172. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, L.; Ye, T.; Lu, Y.; Chen, X.; Wu, Y. The Inhibitory Effect of ABA on Floral Transition Is Mediated by ABI5 in Arabidopsis. J. Exp. Bot. 2013, 64, 675–684. [Google Scholar] [CrossRef]
- Rock, C.D.; Sun, X. Crosstalk Between ABA and Auxin Signaling Pathways in Roots of Arabidopsis thaliana (L.) Heynh. Planta 2005, 222, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, H.; Zhao, Y.; Feng, Z.; Li, Q.; Yang, H.Q.; Luan, S.; Li, J.; He, Z.H. Auxin Controls Seed Dormancy Through Stimulation of Abscisic Acid Signaling by Inducing ARF-Mediated ABI3 Activation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2013, 110, 15485–15490. [Google Scholar] [CrossRef] [PubMed]
- Matilla, A.J. Auxin: Hormonal Signal Required for Seed Development and Dormancy. Plants 2020, 9, 705. [Google Scholar] [CrossRef]
- Xu, W.; Jia, L.; Shi, W.; Liang, J.; Zhou, F.; Li, Q.; Zhang, J. Abscisic Acid Accumulation Modulates Auxin Transport in the Root Tip to Enhance Proton Secretion for Maintaining Root Growth under Moderate Water Stress. New Phytol. 2013, 197, 139–150. [Google Scholar] [CrossRef]
- Wu, S.; Hu, C.; Tan, Q.; Zhao, X.; Xu, S.; Xia, Y.; Sun, X. Nitric Oxide Acts Downstream of Abscisic Acid in Molybdenum-Induced Oxidative Tolerance in Wheat. Plant Cell Rep. 2018, 37, 599–610. [Google Scholar] [CrossRef]
- Yuan, K.; Rashotte, A.M.; Wysocka-Diller, J.W. ABA and GA Signaling Pathways Interact and Regulate Seed Germination and Seedling Development under Salt Stress. Acta Physiol. Plant. 2011, 33, 261–271. [Google Scholar] [CrossRef]
- Nguyen, T.N.; Tuan, P.A.; Ayele, B.T. Jasmonate Regulates Seed Dormancy in Wheat via Modulating the Balance between Gibberellin and Abscisic Acid. J. Exp. Bot. 2022, 73, 2434–2453. [Google Scholar] [CrossRef]
- Muhammad Aslam, M.; Waseem, M.; Jakada, B.H.; Okal, E.J.; Lei, Z.; Saqib, H.S.A.; Yuan, W.; Xu, W.; Zhang, Q. Mechanisms of Abscisic Acid-Mediated Drought Stress Responses in Plants. Int. J. Mol. Sci. 2022, 23, 1084. [Google Scholar] [CrossRef]
- Saini, S.; Sharma, I.; Pati, P.K. Versatile Roles of Brassinosteroid in Plants in the Context of Its Homeostasis, Signaling and Crosstalks. Front. Plant Sci. 2015, 6, 950. [Google Scholar] [CrossRef]
- Yang, X.; Bai, Y.; Shang, J.; Xin, R.; Tang, W. The Antagonistic Regulation of Abscisic Acid-Inhibited Root Growth by Brassinosteroids Is Partially Mediated via Direct Suppression of Abscisic Acid Insensitive 5 Expression by Brassinazole Resistant 1. Plant Cell Environ. 2016, 39, 1994–2003. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Hadi, F.; Ali, N. Effective Phytoextraction of Cadmium (Cd) with Increasing Concentration of Total Phenolics and Free Proline in Cannabis sativa (L.) Plant Under Various Treatments of Fertilizers, Plant Growth Regulators and Sodium Salt. Int. J. Phytoremediat. 2015, 17, 56–65. [Google Scholar] [CrossRef]
- Zhang, S.; Cai, Z.; Wang, X. The Primary Signaling Outputs of Brassinosteroids Are Regulated by Abscisic Acid Signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 4543–4548. [Google Scholar] [CrossRef]
- Avalbaev, A.M.; Somov, K.A.; Yuldashev, R.A.; Shakirova, F.M. Cytokinin Oxidase Is Key Enzyme of Cytokinin Degradation. Biochemistry 2012, 77, 1354–1361. [Google Scholar] [CrossRef] [PubMed]
- Sirko, A.; Wawrzyńska, A.; Brzywczy, J.; Sieńko, M. Control of ABA Signaling and Crosstalk with Other Hormones by the Selective Degradation of Pathway Components. Int. J. Mol. Sci. 2021, 22, 4638. [Google Scholar] [CrossRef] [PubMed]
- Werner, T.; Köllmer, I.; Bartrina, I.; Holst, K.; Schmülling, T. New Insights into the Biology of Cytokinin Degradation. Plant Biol. 2006, 8, 371–381. [Google Scholar] [CrossRef]
- Tran, L.S.P.; Urao, T.; Qin, F.; Maruyama, K.; Kakimoto, T.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional Analysis of AHK1/ATHK1 and Cytokinin Receptor Histidine Kinases in Response to Abscisic Acid, Drought, and Salt Stress in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 20623–20628. [Google Scholar] [CrossRef]
- Chen, L.I.N.; Dodd, I.C.; Davies, W.J.; Wilkinson, S. Ethylene Limits Abscisic Acid-Or Soil Drying-Induced Stomatal Closure in Aged Wheat Leaves. Plant Cell Environ. 2013, 36, 1850–1859. [Google Scholar] [CrossRef]
- Harrison, M.A. Cross-Talk between Phytohormone Signaling Pathways under Both Optimal and Stressful Environmental Conditions. In Phytohormones and Abiotic Stress Tolerance in Plants; Springer: Berlin/Heidelberg, Germany, 2012; pp. 49–76. [Google Scholar]
- Arc, E.; Sechet, J.; Corbineau, F.; Rajjou, L.; Marion-Poll, A. ABA Crosstalk with Ethylene and Nitric Oxide in Seed Dormancy and Germination. Front. Plant Sci. 2013, 4, 63. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, L.; Yu, Y.; Quan, R.; Zhang, Z.; Zhang, H.; Huang, R. The Ethylene Response Factor AtERF11 That Is Transcriptionally Modulated by the bZIP Transcription Factor HY5 Is a Crucial Repressor for Ethylene Biosynthesis in Arabidopsis. Plant J. 2011, 68, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Corbineau, F.; Xia, Q.; Bailly, C.; El-Maarouf-Bouteau, H. Ethylene, a Key Factor in the Regulation of Seed Dormancy. Front. Plant Sci. 2014, 5, 539. [Google Scholar] [CrossRef] [PubMed]
- Kovács, V.; Gondor, O.K.; Szalai, G.; Darkó, E.; Majláth, I.; Janda, T.; Pál, M. Synthesis and Role of Salicylic Acid in Wheat Varieties with Different Levels of Cadmium Tolerance. J. Hazard. Mater. 2014, 280, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Shakirova, F.M.; Allagulova, C.R.; Maslennikova, D.R.; Klyuchnikova, E.O.; Avalbaev, A.M.; Bezrukova, M.V. Salicylic Acid-Induced Protection against Cadmium Toxicity in Wheat Plants. Environ. Exp. Bot. 2016, 122, 19–28. [Google Scholar] [CrossRef]
- Huang, T.L.; Nguyen, Q.T.T.; Fu, S.F.; Lin, C.Y.; Chen, Y.C.; Huang, H.J. Transcriptomic Changes and Signalling Pathways Induced by Arsenic Stress in Rice Roots. Plant Mol. Biol. 2012, 80, 587–608. [Google Scholar] [CrossRef]
- Zdunek-Zastocka, E.; Grabowska, A.; Michniewska, B.; Orzechowski, S. Proline Concentration and Its Metabolism Are Regulated in a Leaf Age Dependent Manner but Not by Abscisic Acid in Pea Plants Exposed to Cadmium Stress. Cells 2021, 10, 946. [Google Scholar] [CrossRef]
- Tan, M.; Cheng, D.; Yang, Y.; Zhang, G.; Qin, M.; Chen, J.; Chen, Y.; Jiang, M. Co-expression Network Analysis of the Transcriptomes of Rice Roots Exposed to Various Cadmium Stresses Reveals Universal Cadmium-Responsive Genes. BMC Plant Biol. 2017, 17, 194. [Google Scholar] [CrossRef]
- Zehra, A.; Choudhary, S.; Wani, K.I.; Naeem, M.; Khan, M.M.A.; Aftab, T. Exogenous Abscisic Acid Mediates ROS Homeostasis and Maintains Glandular Trichome to Enhance Artemisinin Biosynthesis in Artemisia annua under Copper Toxicity. Plant Physiol. Biochem. 2020, 156, 125–134. [Google Scholar] [CrossRef]
- Anderson, J.P.; Badruzsaufari, E.; Schenk, P.M.; Manners, J.M.; Desmond, O.J.; Ehlert, C.; Maclean, D.J.; Ebert, P.R.; Kazan, K. Antagonistic Interaction Between Abscisic Acid and Jasmonate-Ethylene Signaling Pathways Modulates Defense Gene Expression and Disease Resistance in Arabidopsis. Plant Cell 2004, 16, 3460–3479. [Google Scholar] [CrossRef]
- Proietti, S.; Caarls, L.; Coolen, S.; Van Pelt, J.A.; Van Wees, S.C.M.; Pieterse, C.M.J. Genome-Wide Association Study Reveals Novel Players in Defense Hormone Crosstalk in Arabidopsis. Plant Cell Environ. 2018, 41, 2342–2356. [Google Scholar] [CrossRef]
- Chen, Q.; Sun, J.; Zhai, Q.; Zhou, W.; Qi, L.; Xu, L.; Wang, B.; Chen, R.; Jiang, H.; Qi, J.; et al. The Basic Helixloop-Helix Transcription Factor MYC2 Directly Represses PLETHORA Expression During Jasmonate-Mediated Modulation of the Root Stem Cell Niche in Arabidopsis. Plant Cell 2011, 23, 3335–3352. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, C. Cross Talk of Signaling Pathways Between ABA and Other Phytohormones. In Abscisic Acid: Metabolism, Transport and Signaling; Springer: Dordrecht, The Netherlands, 2014; pp. 243–253. [Google Scholar]
- Hancock, J.T.; Neill, S.J.; Wilson, I.D. Nitric Oxide and ABA in the Control of Plant Function. Plant Sci. 2011, 181, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Prakash, V.; Singh, V.P.; Tripathi, D.K.; Sharma, S.; Corpas, F.J. Crosstalk Between Nitric Oxide (NO) and Abscisic Acid (ABA) Signalling Molecules in Higher Plants. Environ. Exp. Bot. 2019, 161, 41–49. [Google Scholar] [CrossRef]
- Zhang, Y.; Tan, J.; Guo, Z.; Lu, S.; He, S.; Shu, W.; Zhou, B. Increased Abscisic Acid Levels in Transgenic Tobacco Over-Expressing 9-cis-epoxycarotenoid Dioxygenase Influence H2O2 and NO Production and Antioxidant Defenses. Plant Cell Environ. 2009, 32, 509–519. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, L.; Ye, N.; Liu, R.; Jia, W.; Zhang, J. Nitric Oxideinduced Rapid Decrease of Abscisic Acid Concentration Is Required in Breaking Seed Dormancy in Arabidopsis. New Phytol. 2009, 183, 1030–1042. [Google Scholar] [CrossRef]
- Signorelli, S.; Considine, M.J. Nitric Oxide Enables Germination by a Four-Pronged Attack on ABA-Induced Seed Dormancy. Front. Plant Sci. 2018, 9, 296. [Google Scholar] [CrossRef]
- Bright, J.; Desikan, R.; Hancock, J.T.; Weir, I.S.; Neill, S.J. ABA-Induced NO Generation and Stomatal Closure in Arabidopsis Are Dependent on H2O2 Synthesis. Plant J. 2006, 45, 113–122. [Google Scholar] [CrossRef]
- Saradadevi, R.; Bramley, H.; Siddique, K.H.M.; Edwards, E.; Palta, J.A. Reprint of ‘Contrasting Stomatal Regulation and Leaf ABA Concentrations in Wheat Genotypes When Split Root Systems Were Exposed to Terminal Drought’. Field Crops Res. 2014, 165, 5–14. [Google Scholar] [CrossRef]
- Hung, K.T.; Kao, C.H. Nitric Oxide Counteracts the Senescence of Rice Leaves Induced by Abscisic Acid. J. Plant Physiol. 2003, 160, 871–879. [Google Scholar] [CrossRef]
- Iqbal, N.; Umar, S.; Khan, N.A.; Corpas, F.J. Crosstalk Between Abscisic Acid and Nitric Oxide under Heat Stress: Exploring New Vantage Points. Plant Cell Rep. 2021, 40, 1429–1450. [Google Scholar] [CrossRef]
- Iqbal, N.; Sehar, Z.; Fatma, M.; Umar, S.; Sofo, A.; Khan, N.A. Nitric Oxide and Abscisic Acid Mediate Heat Stress Tolerance through Regulation of Osmolytes and Antioxidants to Protect Photosynthesis and Growth in Wheat Plants. Antioxidants 2022, 11, 372. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, H.I.; Latif, H.H.; Hanafy, R.S. Influence of Nitric Oxide Application on Some Biochemical Aspects, Endogenous Hormones, Minerals and Phenolic Compounds of Vicia faba Plant Grown Under Arsenic Stress. Gesunde Pflanz. 2016, 68, 99–107. [Google Scholar] [CrossRef]
- Sadeghipour, O. Nitric Oxide Increases Pb Tolerance by Lowering Pb Uptake and Translocation as well as Phytohormonal Changes in Cowpea (Vigna unguiculata (L.) Walp.). Sains Malays. 2017, 46, 189–195. [Google Scholar] [CrossRef]
Figure 1.
Overview of ABA mechanism under normal stress and HM stress conditions that leads to physiological, biochemical, and molecular changes. Various transporters situated in the roots allow HMs to enter the plant and then move to various locations inside it. Excessive HM accumulation in plant tissues contributes to toxicity.
Figure 1.
Overview of ABA mechanism under normal stress and HM stress conditions that leads to physiological, biochemical, and molecular changes. Various transporters situated in the roots allow HMs to enter the plant and then move to various locations inside it. Excessive HM accumulation in plant tissues contributes to toxicity.

Figure 2.
Synthesis and catabolic routes of ABA. Where: abscisic aldehyde oxidase (AAO); zeaxanthin epoxidase (ZEP); ABA catabolic pathways (UGT71C5); PA reductase (PAR); molybdenum cofactor (MoCo); the gene encoding β-glucosyltransferase (CYP707As); a β-glucosyltransferase (GT); β-glucosidases 1/2 (BG1/2).
Figure 2.
Synthesis and catabolic routes of ABA. Where: abscisic aldehyde oxidase (AAO); zeaxanthin epoxidase (ZEP); ABA catabolic pathways (UGT71C5); PA reductase (PAR); molybdenum cofactor (MoCo); the gene encoding β-glucosyltransferase (CYP707As); a β-glucosyltransferase (GT); β-glucosidases 1/2 (BG1/2).

Figure 3.
Abscisic acid and other phytohormone signaling genes under HM stress.

Figure 4.
Mechanism of ABA in mitigating HM stress in plants.

Figure 5.
Crosstalk of ABA with other phytohormones under HM stress. Where: NO (nitric oxide); AUX (auxin); PA (polyamines); SA (salicylic acid); ETH (ethylene); PDR12 (pleiotropic drug resistance transporter 12); PIN2 (auxin polar transportation network); ethylene TFs are EIL1 and EIN3; MAPK6 is mitogen-activated protein kinase, a signaling cascade of JA and ethylene.
Figure 5.
Crosstalk of ABA with other phytohormones under HM stress. Where: NO (nitric oxide); AUX (auxin); PA (polyamines); SA (salicylic acid); ETH (ethylene); PDR12 (pleiotropic drug resistance transporter 12); PIN2 (auxin polar transportation network); ethylene TFs are EIL1 and EIN3; MAPK6 is mitogen-activated protein kinase, a signaling cascade of JA and ethylene.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bano, A.; Singh, K.; Singh, S.P.; Sharma, P. Abscisic Acid: Metabolism, Signaling, and Crosstalk with Other Phytohormones under Heavy Metal Stress. Stresses 2023, 3, 665-686. https://0-doi-org.brum.beds.ac.uk/10.3390/stresses3040046
AMA Style
Bano A, Singh K, Singh SP, Sharma P. Abscisic Acid: Metabolism, Signaling, and Crosstalk with Other Phytohormones under Heavy Metal Stress. Stresses. 2023; 3(4):665-686. https://0-doi-org.brum.beds.ac.uk/10.3390/stresses3040046
Chicago/Turabian StyleBano, Ambreen, Kratika Singh, Surendra Pratap Singh, and Pooja Sharma. 2023. "Abscisic Acid: Metabolism, Signaling, and Crosstalk with Other Phytohormones under Heavy Metal Stress" Stresses 3, no. 4: 665-686. https://0-doi-org.brum.beds.ac.uk/10.3390/stresses3040046