
Assessment of Genetic Diversity of Rice in Registered Cultivars and Farmers’ Fields in Burkina Faso

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
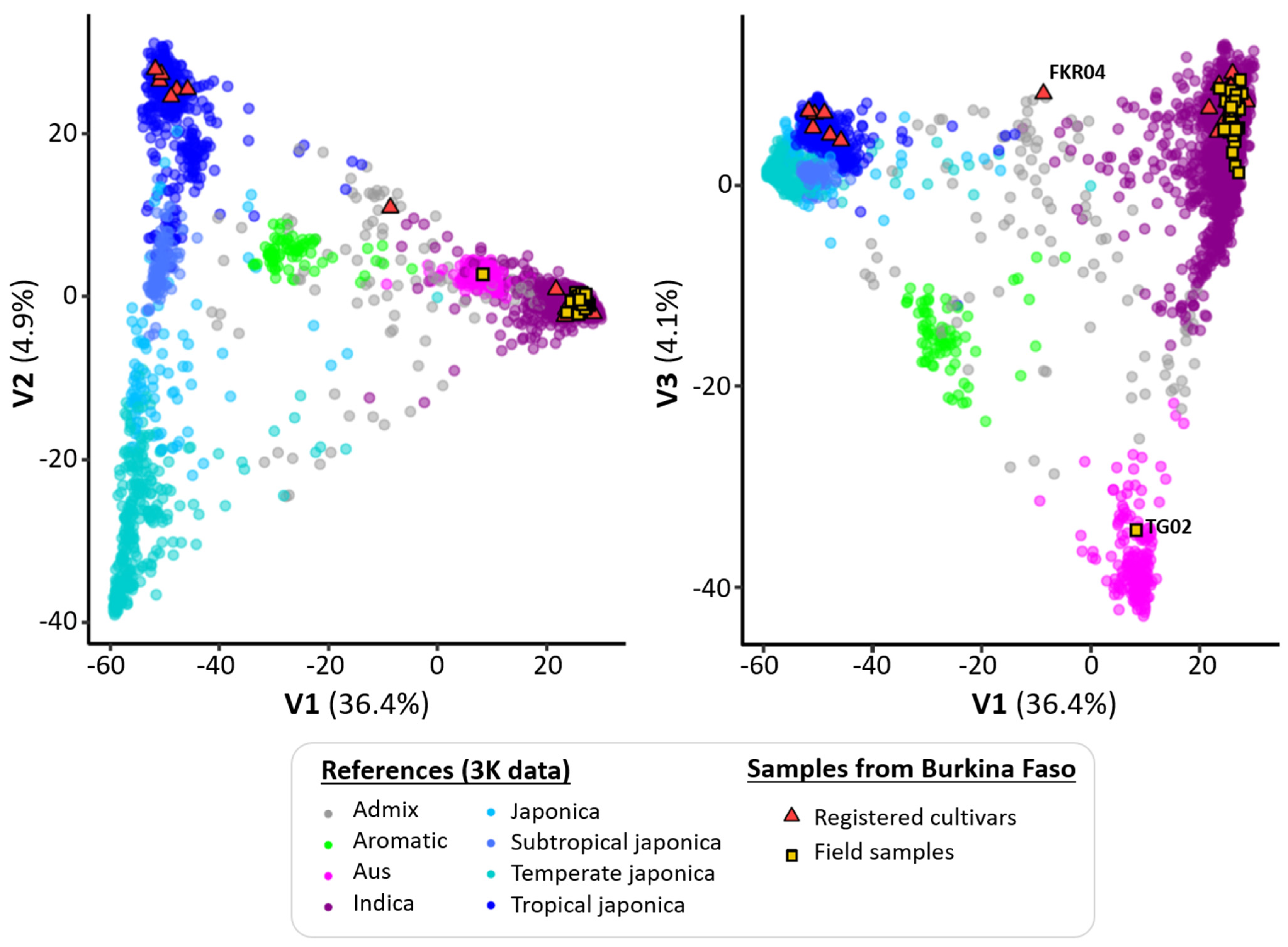
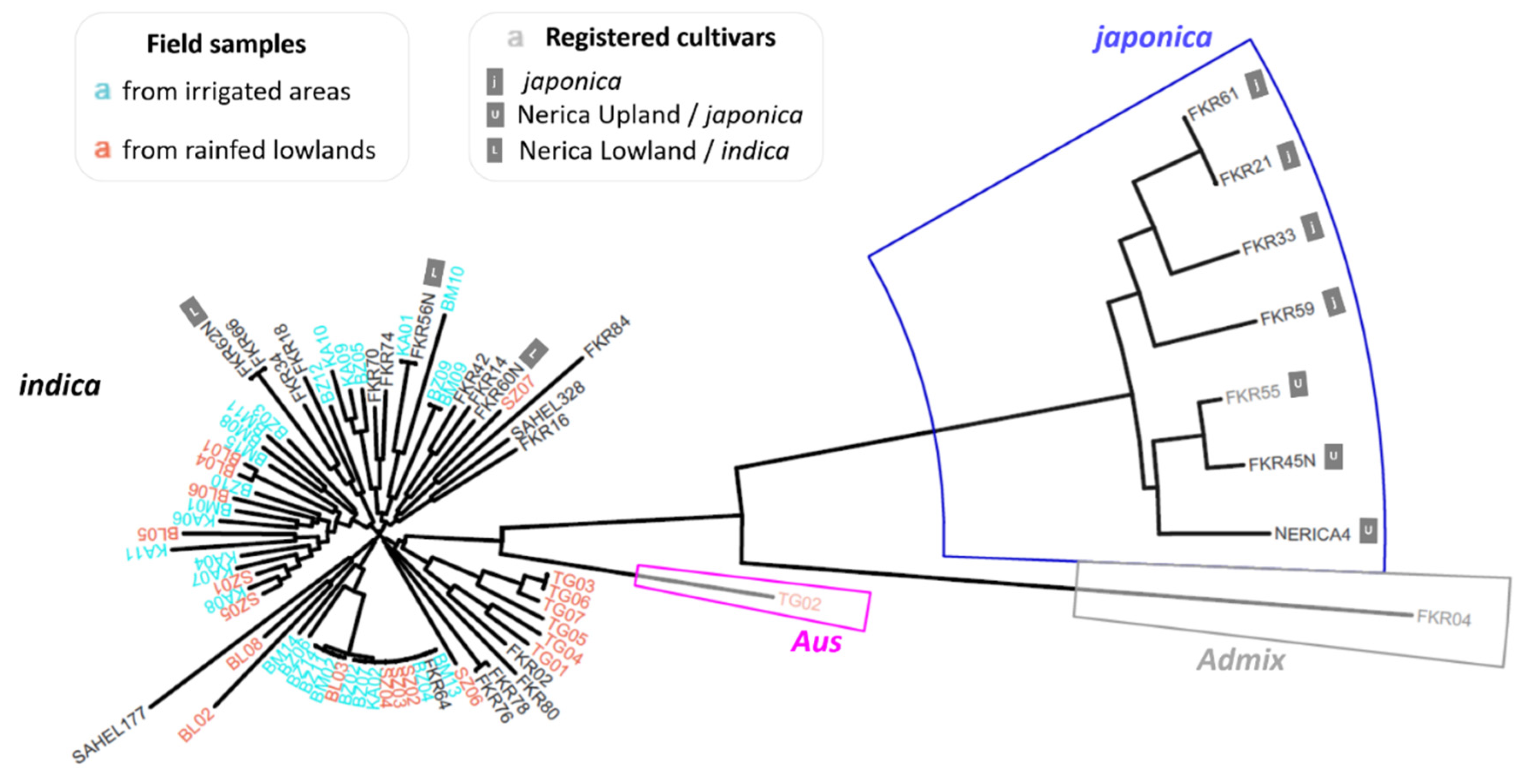
3. Results
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zimmerer, K.S.; de Haan, S.; Jones, A.D.; Creed-Kanashiro, H.; Tello, M.; Carrasco, M.; Meza, K.; Plasencia Amaya, F.; Cruz-Garcia, G.S.; Tubbeh, R.; et al. The biodiversity of food and agriculture (Agrobiodiversity) in the anthropocene: Research advances and conceptual framework. Anthropocene 2019, 25, 100192. [Google Scholar] [CrossRef]
- Pironon, S.; Etherington, T.R.; Borrell, J.S.; Kühn, N.; Macias-Fauria, M.; Ondo, I.; Tovar, C.; Wilkin, P.; Willis, K.J. Potential adaptive strategies for 29 sub-Saharan crops under future climate change. Nat. Clim. Chang. 2019, 9, 758–763. [Google Scholar] [CrossRef]
- Soullier, G.; Demont, M.; Arouna, A.; Lancon, F.; del Villar, P.M. The state of rice value chain upgrading in West Africa. Glob. Food Secur.-Agric. Policy Econ. Environ. 2020, 25, 100365. [Google Scholar] [CrossRef]
- Linares, O.F. African rice (Oryza glaberrima): History and future potential. Proc. Natl. Acad. Sci. USA 2002, 99, 16360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarla, N.; Swamy, B.P.M. Oryza glaberrima: A source for the improvement of Oryza sativa. Curr. Sci. 2005, 89, 955–963. [Google Scholar]
- Ndjiondjop, M.N.; Semagn, K.; Sow, M.; Manneh, B.; Gouda, A.C.; Kpeki, S.B.; Pegalepo, E.; Wambugu, P.; Sie, M.; Warburton, M.L. Assessment of Genetic Variation and Population Structure of Diverse Rice Genotypes Adapted to Lowland and Upland Ecologies in Africa Using SNPs. Front. Plant Sci. 2018, 9, 446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; He, W.; Nassirou, T.Y.; Nsabiyumva, A.; Dong, X.; Adedze, Y.M.N.; Jin, D. Molecular characterization and genetic diversity of different genotypes of Oryza sativa and Oryza glaberrima. Electron. J. Biotechnol. 2017, 30, 48–57. [Google Scholar] [CrossRef]
- Luther, Z.; Akromah, R.; Nyadanu, D.; Tokpah, D.; Page, Z.; Voor, V.; Kwaloe, A. Evaluation of genetic diversity in rice (Oryza sativa and Oryza glaberrima) germplasm from Liberia and Ghana using simple sequence repeat (SSR) markers. Afr. J. Biotechnol. 2017, 16, 1990–1996. [Google Scholar]
- Yelome, O.I.; Audenaert, K.; Landschoot, S.; Dansi, A.; Vanhove, W.; Silue, D.; Van Damme, P.; Haesaert, G. Analysis of population structure and genetic diversity reveals gene flow and geographic patterns in cultivated rice (O. sativa and O. glaberrima) in West Africa. Euphytica 2018, 214, 215. [Google Scholar] [CrossRef]
- Diop, B.; Wang, D.R.; Drame, K.N.; Gracen, V.; Tongoona, P.; Dzidzienyo, D.; Nartey, E.; Greenberg, A.J.; Djiba, S.; Danquah, E.Y.; et al. Bridging old and new: Diversity and evaluation of high iron-associated stress response of rice cultivated in West Africa. J. Exp. Bot. 2020, 71, 4188–4200. [Google Scholar] [CrossRef] [Green Version]
- Sié, M.; Zongo, J.D.; Dona Dakouo, D. Prospection des cultivars traditionnels de riz du Burkina Faso. Rev. CAMES Sci. Médecine 1998, 00, 21–27. [Google Scholar]
- Sié, M.; Ghesquière, A.; Miezan, K. Structure génétique des variétés traditionnelles de riz (Oryza sp.) du Burkina Faso. Agron. Afr. 1999, 11, 57–71. [Google Scholar]
- Kam, H.; Ndjiondjop, M.; Sanou, J.; Sow, M.; Ouédraogo, I.; Laing, M. Diversity assessment and geographical variability of rice landraces collected from Burkina Faso. Int. J. Dev. Sustain. 2017, 6, 479–496. [Google Scholar]
- Kam, H.; Ndjiondjop, M.; Laing, M.; Ahmadi, N. Molecular characterization and diversity analysis of Burkina Faso rice landraces using 23 microsatellite markers and establishment of a core collection. Int. J. Curr. Res. 2017, 9, 56222–56232. [Google Scholar]
- Demont, M.; Ndour, M.; Zossou, E. Can local African rice be competitive? An analysis of quality-based competitiveness through experimental auctions. Cah. Agric. 2013, 22, 345–352. [Google Scholar] [CrossRef]
- Cotonou, B. Boosting Africa’s Rice Sector: A Research for Development Strategy 2011–2020; AfricaRice: Bouaké, Cote D’Ivoire, 2011; p. 77. [Google Scholar]
- Katic, P.G.; Namara, R.E.; Hope, L.; Owusu, E.; Fujii, H. Rice and irrigation in West Africa: Achieving food security with agricultural water management strategies. Water Resour. Econ. 2013, 1, 75–92. [Google Scholar] [CrossRef]
- MAHRH—Ministère de l’Agriculture, de l’Hydraulique et des Ressources Halieutiques (Ministry of Agriculture, Hydraulics and Fisheries Resources in Burkina Faso). Burkina Faso National Rice Development Strategy (NRDS). 2011. Available online: https://riceforafrica.net/images/stories/PDF/burkina_faso_en.pdf (accessed on 30 October 2021).
- DGPER—Direction Générale de la Promotion de l’Economie Rurale (General Direction for the Promotion of Rural Economy in Burkina Faso). Résultats Définitifs de la Campagne Agricole et de la Situation Alimentaire et Nutritionnelle 2009/2010. 2010. Available online: http://cns.bf/IMG/pdf/masa_rdepa_2009-2010.pdf (accessed on 30 October 2021).
- McCouch, S.; Wing, R.; Semon, M.; Venuprasad, R.; Atlin, G.; Sorrells, M.; Jannink, J. Making Rice Genomics Work for Africa. In Realizing Africa’s Rice Promise; Wopereis, M., David Johnson, D., Ahmadi, N., Tollens, E., Jalloh, A., Eds.; CABI: Wallingford, UK, 2013; pp. 108–129. [Google Scholar]
- Semagn, K.; Ndjiondjop, M.N.; Cissoko, M. Microsatellites and agronomic traits for assessing genetic relationships among 18 New Rice for Africa (NERICA) varieties. Afr. J. Biotechnol. 2006, 5, 800–810. [Google Scholar]
- Bocco, R.; Lorieux, M.; Seck, P.A.; Futakuchi, K.; Manneh, B.; Baimey, H.; Ndjiondjop, M.N. Agro-morphological characterization of a population of introgression lines derived from crosses between IR 64 (Oryza sativa indica) and TOG 5681 (Oryza glaberrima) for drought tolerance. Plant Sci. 2012, 183, 65–76. [Google Scholar] [CrossRef]
- Barro, M.; Kassankogno, A.I.; Wonni, I.; Sereme, D.; Somda, I.; Kaboré, K.H.; Bena, G.; Brugidou, C.; Tharreau, D.; Tollenaere, C. Spatiotemporal Survey of Multiple Rice Diseases in Irrigated Areas Compared to Rainfed Lowlands in the Western Burkina Faso. Plant Dis. 2021. [Google Scholar] [CrossRef]
- Thomson, M.J. High-Throughput SNP Genotyping to Accelerate Crop Improvement. Plant Breed. Biotechnol. 2014, 2, 195–212. [Google Scholar] [CrossRef]
- Kretzschmar, T.; Mbanjo, E.G.N.; Magalit, G.A.; Dwiyanti, M.S.; Habib, M.A.; Diaz, M.G.; Hernandez, J.; Huelgas, Z.; Malabayabas, M.L.; Das, S.K.; et al. DNA fingerprinting at farm level maps rice biodiversity across Bangladesh and reveals regional varietal preferences. Sci. Rep. 2018, 8, 14920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.A.M.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. GigaScience 2015, 4, 7. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. GigaScience 2021, 10, giab008. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: www.r-project.org/about.html (accessed on 30 October 2021).
- Gain, C.; François, O. LEA 3: Factor models in population genetics and ecological genomics with R. Mol. Ecol. Resour. 2021, 1–11. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. Available online: https://ggplot2.tidyverse.org (accessed on 21 April 2021).
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- Goudet, J.; Jombart, T. Hierfstat: Estimation and Tests of Hierarchical F-Statistics. R Package Version 0.5-7. 2020. Available online: https://CRAN.R-project.org/package=hierfstat (accessed on 21 July 2021).
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef] [PubMed]
- Yu, G. Using ggtree to Visualize Data on Tree-Like Structures. Curr. Protoc. Bioinform. 2020, 69, e96. [Google Scholar] [CrossRef] [PubMed]
- Dray, S.; Dufour, A.-B. The ade4 Package: Implementing the Duality Diagram for Ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Ndjiondjop, M.N.; Semagn, K.; Zhang, J.W.; Gouda, A.C.; Kpeki, S.B.; Goungoulou, A.; Wambugu, P.; Drame, K.N.; Bimpong, I.K.; Zhao, D.L. Development of species diagnostic SNP markers for quality control genotyping in four rice (Oryza L.) species. Mol. Breed. 2018, 38, 131. [Google Scholar] [CrossRef] [Green Version]
- Loko, Y.L.E.; Ewedje, E.E.; Orobiyi, A.; Djedatin, G.; Toffa, J.; Gbemavo, C.D.S.J.; Tchakpa, C.; Gavoedo, D.; Sedah, P.; Sabot, F. On-farm management of rice diversity, varietal preference criteria, and farmers’ perceptions of the African (Oryza glaberrima Steud.) versus Asian rice (Oryza sativa L.) in the Republic of Benin (West Africa): Implications for breeding and conservation. Econ. Bot. 2021, 75, 1–29. [Google Scholar] [CrossRef]
- Arouna, A.; Lokossou, J.C.; Wopereis, M.C.S.; Bruce-Oliver, S.; Roy-Macauley, H. Contribution of improved rice varieties to poverty reduction and food security in sub-Saharan Africa. Glob. Food Secur. 2017, 14, 54–60. [Google Scholar] [CrossRef]
- Mokuwa, A.; Nuijten, E.; Okry, F.; Teeken, B.; Maat, H.; Richards, P.; Struik, P.C. Robustness and Strategies of Adaptation among Farmer Varieties of African Rice (Oryza glaberrima) and Asian Rice (Oryza sativa) across West Africa. PLoS ONE 2013, 8, e34801. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Synonym | Origin | Introduction Date | Time to Maturity | Group |
|---|---|---|---|---|---|
| FKR02 | GAMBIAKA | Gambia | 1970 | 145 | indica |
| FKR04 | SINTANE DIOFOR | Senegal (Casamance) | 1960 | 120 | - |
| FKR14 | 4418 | Inde | 1976 | 125 | indica |
| FKR16 | 4456 | Inde | 1976 | 120 | indica |
| FKR18 | SC 27 | Burkina Faso | 1980 | 135 | indica |
| FKR21 | ITA 257 | IITA/Nigeria | 1987 | 98 | japonica |
| FKR33 | 1195-5-2 | Burkina Faso | 1982 | 98 | japonica |
| FKR34 | RP 1125-1526-2 | Inde | 1984 | 129 | indica |
| FKR42 | IR64 | IRRI/Philippines | 1989 | 123 | indica |
| FKR45N | WAB880-1-38 NERICA 12 | AfricaRice | 1999 | 95 | japonica (NERICA upland) |
| FKR55 | WAB450-I-BL-1-736-HB NERICA8 | AfricaRice/INERA | - | 75–95 | japonica (NERICA upland) |
| FKR56N | WSA 161-B-9-3 NERICA L-41 | AfricaRice | 1999 | 116 | indica (NERICA, lowland) |
| FKR59 | WAB9984 | AfricaRice | 2009 | 90 | japonica |
| FKR60N | WAS122-IDSA-1 NERICA L-20 | AfricaRice | 1999 | 115 | Indica (NERICA, lowland) |
| FKR61 | WAB C1 65 | AfricaRice | 2009 | 90 | japonica |
| FKR62N | WAS 122-IDSA-1 NERICA L-19 | AfricaRice | 1999 | 118 | Indica (NERICA, lowland) |
| FKR64 | TS2 | Taiwan DGPV/INERA | - | 120 | indica |
| FKR66 | WAT1046-B43 | AfricaRice | 2010 | 125 | indica |
| FKR70 | IR 75-884-12-12 | AfricaRice | 2009 | 130 | indica |
| FKR74 | WAB 2094-WAC2-TGR2-B ARICA1 | AfricaRice/INERA | - | 101 | indica (ARICA lowland) |
| FKR76 | F6-36 | DGPV/INERA | 2012 | 90 | indica |
| FKR78 | F6-41 | DGPV/INERA | 2012 | 97 | indica |
| FKR80 | F6-49 | DGPV/INERA | 2012 | 98 | indica |
| FKR84 | ORYLUX 6 | AfricaRice/INERA | - | 100–105 | indica |
| NERICA4 | WAB450-I-B-P-91-HB | AfricaRice/INERA | - | 95–100 | japonica (NERICA upland) |
| SAHEL177 | WAS 197-B-6-3-11 | AfricaRice/Saint-Louis | - | 122 | indica |
| SAHEL328 | WAS 197-B-4-1-5 | ISRA/AfricaRice | - | 116 | indica |
| Geographical Zone | Site | Rice Production System | Latitude | Longitude | N Samples | Gene Diversity | |
|---|---|---|---|---|---|---|---|
| Bama | Badala (BL) | RL | 11.368 | −4.373 | 7 | 0.125 | 0.117 |
| Bama (BM) | IR | 11.390 | −4.410 | 9 | 0.131 | ||
| Banzon | Senzon (SZ) | RL | 11.288 | −4.829 | 7 | 0.113 | 0.124 |
| Banzon (BZ) | IR | 11.335 | −4.796 | 11 | 0.108 | ||
| Karfiguela | Tengrela (TG) | RL | 10.648 | −4.838 | 7 | 0.155 | 0.132 |
| Karfiguela (KA) | IR | 10.678 | −4.813 | 9 | 0.130 | ||
| Average | 0.131 | 0.124 | |||||
| Global | 50 | 0.137 | |||||
| Bama | Banzon | Karfiguela | |||||
|---|---|---|---|---|---|---|---|
| Geographical Zone | Study Site | Badala (BL) | Bama (BM) | Senzon (SZ) | Banzon (BZ) | Tengrela (TG) | Karfiguela (KA) |
| Bama | Badala (BL) | - | |||||
| Bama (BM) | 0.0018 | - | |||||
| Banzon | Senzon (SZ) | 0.0123 | −0.0194 | - | |||
| Banzon (BZ) | 0.0563 | −0.0129 | −0.0194 | - | |||
| Karfiguela | Tengrela (TG) | 0.2949 | 0.2531 | 0.2834 | 0.3288 | - | |
| Karfiguela (KA) | 0.0310 | 0.0118 | 0.0408 | 0.1074 | 0.2615 | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barro, M.; Konate, K.A.; Wonni, I.; Kassankogno, A.I.; Sabot, F.; Albar, L.; Somda, I.; Béna, G.; Ghesquière, A.; Kam, H.; et al. Assessment of Genetic Diversity of Rice in Registered Cultivars and Farmers’ Fields in Burkina Faso. Crops 2021, 1, 129-140. https://0-doi-org.brum.beds.ac.uk/10.3390/crops1030013
Barro M, Konate KA, Wonni I, Kassankogno AI, Sabot F, Albar L, Somda I, Béna G, Ghesquière A, Kam H, et al. Assessment of Genetic Diversity of Rice in Registered Cultivars and Farmers’ Fields in Burkina Faso. Crops. 2021; 1(3):129-140. https://0-doi-org.brum.beds.ac.uk/10.3390/crops1030013
Chicago/Turabian StyleBarro, Mariam, Kadougoudiou Abdourasmane Konate, Issa Wonni, Abalo Itolou Kassankogno, François Sabot, Laurence Albar, Irénée Somda, Gilles Béna, Alain Ghesquière, Honoré Kam, and et al. 2021. "Assessment of Genetic Diversity of Rice in Registered Cultivars and Farmers’ Fields in Burkina Faso" Crops 1, no. 3: 129-140. https://0-doi-org.brum.beds.ac.uk/10.3390/crops1030013