
Small RNAs Worm Up Transgenerational Epigenetics Research


Department of Biochemistry, BU Genome Science Institute, School of Medicine, Boston University, 72 E. Concord St. K422, Boston, MA 02118, USA
DNA 2021, 1(2), 37-48; https://0-doi-org.brum.beds.ac.uk/10.3390/dna1020005
Submission received: 18 August 2021
/
Revised: 20 September 2021
/
Accepted: 23 September 2021
/
Published: 29 September 2021
Abstract
:DNA is central to the propagation and evolution of most living organisms due to the essential process of its self-replication. Yet it also encodes factors that permit epigenetic (not included in DNA sequence) flow of information from parents to their offspring and beyond. The known mechanisms of epigenetic inheritance include chemical modifications of DNA and chromatin, as well as regulatory RNAs. All these factors can modulate gene expression programs in the ensuing generations. The nematode Caenorhabditis elegans is recognized as a pioneer organism in transgenerational epigenetic inheritance research. Recent advances in C. elegans epigenetics include the discoveries of control mechanisms that limit the duration of RNA-based epigenetic inheritance, periodic DNA motifs that counteract epigenetic silencing establishment, new mechanistic insights into epigenetic inheritance carried by sperm, and the tantalizing examples of inheritance of sensory experiences. This review aims to highlight new findings in epigenetics research in C. elegans with the main focus on transgenerational epigenetic phenomena dependent on small RNAs.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Numerous discoveries in basic science were facilitated by the model organism Caenorhabditis elegans (reviewed in [1]) (Figure 1).
C. elegans is especially rich in small regulatory RNAs that control most aspects of its development, lifespan, and reproduction. The best-known class of small RNAs, microRNA (miRNA), was discovered through genetic analyses of cell lineage in C. elegans that resulted in the identification of the founding members of the miRNA family: lin-4 in 1993 [2,3] and let-7 in 2000 [4,5], by Ambros, Ruvkun, and their colleagues. The significance of miRNAs in medicine through their use as biomarkers, drug targets, or RNA drugs is beyond dispute (reviewed in [6]), and it is hard to imagine the loss to society had miRNAs not been discovered.
One of the most fascinating areas of C. elegans research is the epigenetic inheritance of small RNAs and their potential roles in shaping genome architecture and mediating heritable adaptive responses to environmental challenges. The connection between the human diet and epigenetic adaptation to existing conditions via heritable RNA transmission in sperm (reviewed in [7]) underscores the importance of lessons learned from C. elegans.
This manuscript provides an overview of key findings that may interest a wide scientific audience, including the highlights from the Genetics Society of America 23rd International C. elegans Conference, which was held virtually on 21–24 June 2021. Detailed descriptions of the field targeting specialists can be found elsewhere, for example [8,9,10,11,12,13].
2. C. elegans Model Systems for Studying Transgenerational Epigenetic Inheritance (TEI)
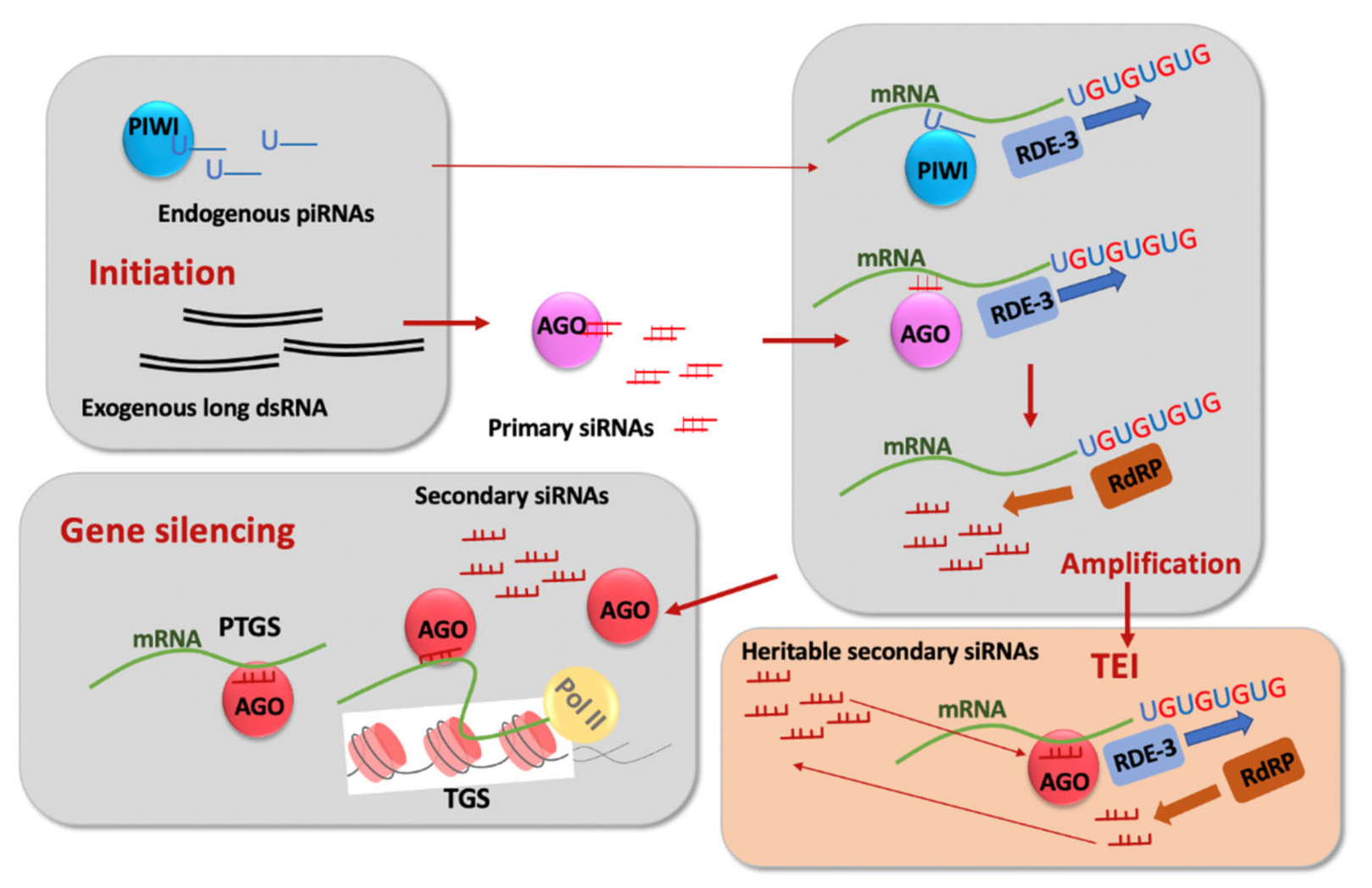
The first indication of the potent role of RNA molecules in inhibiting the expression of C. elegans genes came from applications of antisense RNA technology to nematodes [14,15]. This was followed by the seminal discovery of double-stranded RNA (dsRNA) [16] (Figure 2) as an initiator/intermediate in the silencing process termed RNA interference (RNAi) [16,17] and the identification of small interfering RNAs (siRNAs) (Figure 2) as key players in silencing induced by dsRNA in various organisms [18].
Although RNAi of genes expressed in the soma persisted in first-generation progeny (F1) of treated animals and was not inherited further [16], it was soon realized that the dsRNA-induced phenotypes mimicking mutations in germline-expressed genes could persist longer than the F1 generation and could be transmitted via factors not linked to the target DNA locus and distinct from dsRNA (likely siRNAs) [19]. Subsequent research identified the RNA-dependent RNA polymerase (RdRP)-based siRNA amplification mechanism using mRNAs as templates [20,21,22,23,24] (Figure 2). The outstanding question of how RdRPs find their appropriate target RNAs has recently been answered by the discovery of 3′ tags consisting of non-templated UG additions that attract RdRPs [25,26,27] (Figure 2). Since the deposition of UG tags required for siRNA regeneration is ultimately initiated by siRNAs, this system can lead to the potentially indefinite perpetuation of siRNA-induced gene silencing on post-transcriptional or co-transcriptional levels [26] (Figure 2).
Since most germline-expressed genes are essential for viability, their silencing compromises the production of gravid progeny, thus hampering inheritance studies. Therefore, new systems where silencing could be monitored for many generations were needed. In one such system, which is based on the silencing of the oma-1 gene containing a conditional-lethal mutation, and which monitors animal survival rather than death, the effect of the initial dsRNA exposure was shown to persist for four generations [28]. Other systems were made possible by technical advances in transgenic technology that allowed stable expression of transgenic arrays in the C. elegans germline [29]. This allowed investigators to monitor germline GFP transgene silencing in the progeny of animals exposed to dsRNA, which was shown to last up to twenty generations with selection [30,31]. When the single-copy controlled genomic integration of transgenes was developed [32,33], it further facilitated Transgenerational Epigenetic Inheritance (TEI) research [34,35,36]. Most, but not all [37], subsequent studies researching mechanisms of TEI in C. elegans utilized the oma-1 and/or germline GFP readouts.
3. RNA-Based and Chromatin-Based Epigenetic Silencing and Their Connections
In the last 10–15 years, TEI phenomena have been described in the context of gene silencing induced by endogenous Piwi-interacting RNAs (piRNAs) (Figure 2), and a broad term to describe long-lasting silencing has been introduced: RNA-induced epigenetic silencing (RNAe) [36,38,39]. The targets used to monitor the persistence of piRNA-induced silencing are usually single copy germline-expressed GFP-based sensors containing sequences complementary to some endogenous piRNAs. The persistence of piRNA-induced RNAe, similarly to dsRNA-induced RNAe, relies on the amplification cycle of siRNAs [36,38,39,40] (Figure 2). In addition to siRNA amplification, epigenetic inheritance of the compacted chromatin state is thought to play a role in various TEI phenomena [34,36,38]. In some examples of TEI, maintenance of silencing persisted apparently indefinitely [36,38]. Until very recently, the dependence of TEI on small RNAs and the silencing-associated chromatin marks [34,36,38,41,42] has been interpreted in terms of an RNA-dependent initiation step followed by a chromatin-based maintenance step [34,36,42]. However, recent studies showed that chromatin regulators may act as early as in the P0 (parents exposed to dsRNA) generation of dsRNA-induced TEI, whereas siRNA amplification sustains TEI maintenance [35,43]. Notably, dsRNA-induced silencing of repetitive transgenes expressed in somatic tissues occurs at the transcriptional level in the F1 generation of treated worms [44], and the somatic nuclear RNAi pathway is required for gene silencing in the soma of F1 larvae [45]. However, transcriptional silencing of somatic genes does not display TEI.
Both maternal and paternal inheritance of RNA-induced epigenetic silencing is usually observed in C. elegans [19,28,36,46]. A variety of RNA- and protein-rich granules, which are not bound by membranes, exist in the C. elegans germline. These granules contain mRNA and proteins implicated in the biogenesis and function of small RNAs, including those mediating TEI (reviewed in [47]). At least some of these granules, such as P-granules, are present in oocytes [48] and therefore could harbor heritable siRNA generated through the amplification cycle (see Section 2). The most recent work by Ketting and colleagues determined that mature sperm does not inherit granules that are common between the male and hermaphrodite germlines and described a novel sperm-specific condensate—PEI (paternal epigenetic inheritance) granule—named after its resident protein PEI-1 [49]. Importantly, the authors identify pei-1-like genes in humans, which points to the possible conservation of paternal epigenetic inheritance mechanisms.
4. Permanent and Limited Forms of TEI and Their Genetic Control
In C. elegans, a process of indefinite silencing of repetitive transgenes designed to be expressed in the germline has been described by Kelly and co-authors in 1997 [50]. It was shown to be related to but distinct from the dsRNA-induced gene silencing [51]. The use of single-copy integrated transgenes for TEI research is convenient but at the same time begs the question of its relevance to the understanding of the epigenetic mechanisms involved in endogenous gene regulation. The piRNA-induced RNAe in particular has been likened to epigenetic silencing of parasitic elements that must be inactivated indefinitely [36,39,40,52].
An interesting case of mating-induced silencing of a single copy transgene, which fortuitously contained DNA sequences complementary to endogenous piRNAs, has been reported recently by Jose and colleagues [53]. In this system, the transgene inherited paternally was subject to silencing by maternal piRNAs that lasted apparently indefinitely (>300 generations) and required siRNA amplification machinery in each generation for its propagation [53]. This case illustrates the efficiency of maternal piRNAs in inducing silencing of the “foreign” DNA inherited through paternal sperm. Importantly, when the mothers also expressed the transgene, mating-induced silencing did not occur [53] underscoring the discrimination between “self” and “nonself” proposed by Mello and colleagues [36]. This example of TEI in C. elegans resembles the phenomenon in Drosophila where maternal piRNAs silence paternally-inherited transposons [54].
The mutants that disrupt RNAe generally display a mortal germline phenotype (Mrt)—a progressive sterility that increases over multiple generations [55]. Both the de-repression of normally silenced repetitive elements [52] and the inappropriate silencing of essential endogenous genes, such as histones [56,57], were correlated with the Mrt phenotype. A very recent study by Fire and co-workers identified enhanced production of small RNAs antisense to rRNA as a likely cause of the Mrt phenotypes in animals lacking piRNAs [58]. Guang and colleagues characterized this new class of antisense ribosomal siRNAs (risiRNAs) earlier and had shown that they inhibit rRNA expression via the nuclear RNAi pathway (reviewed in [59]). Importantly, copy-number amplification of rDNA delayed the onset of sterility in piRNA-depleted worms supporting the causal role of impaired ribosome function in the Mrt phenotype [58].
Model systems displaying limited TEI that lasts for 6–7 generations are more suitable for studying adaptive and dynamic epigenetic phenomena. The important question in such systems is: what determines the escape from TEI after several generations? Genetic screens seeking mutants allowing longer TEI revealed the existence of control mechanisms limiting its duration [60]. This finding is conceptually significant because it demonstrates that organisms might not only allow a certain degree of environmentally-induced epigenetic change but also control its duration. The inhibitors of TEI include both chromatin regulators, such as a histone methyltransferase MET-2 [61] and a chromatin-binding protein CEC-9/HERI-1 [60], and RNA-based mechanisms [60]. Intriguingly, piRNAs are capable of both initiating TEI and suppressing its maintenance [62,63], although it is not clear whether the latter effect is direct. Notably, although the siRNA amplification system is unique to nematodes, piRNAs, as well as their RdRP-independent amplification machinery, are present in the germlines of all animals [64].
5. Coordination between Gene Silencing in the Soma and Germline; Who Is the Messenger?
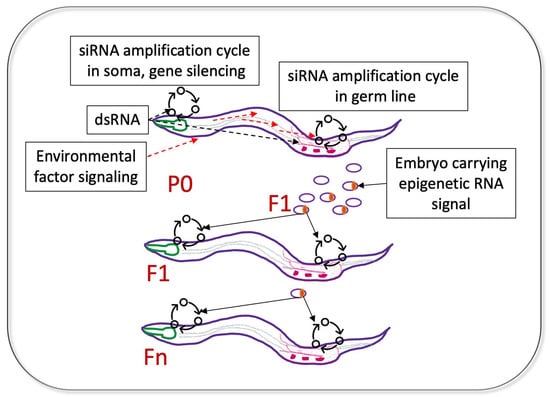
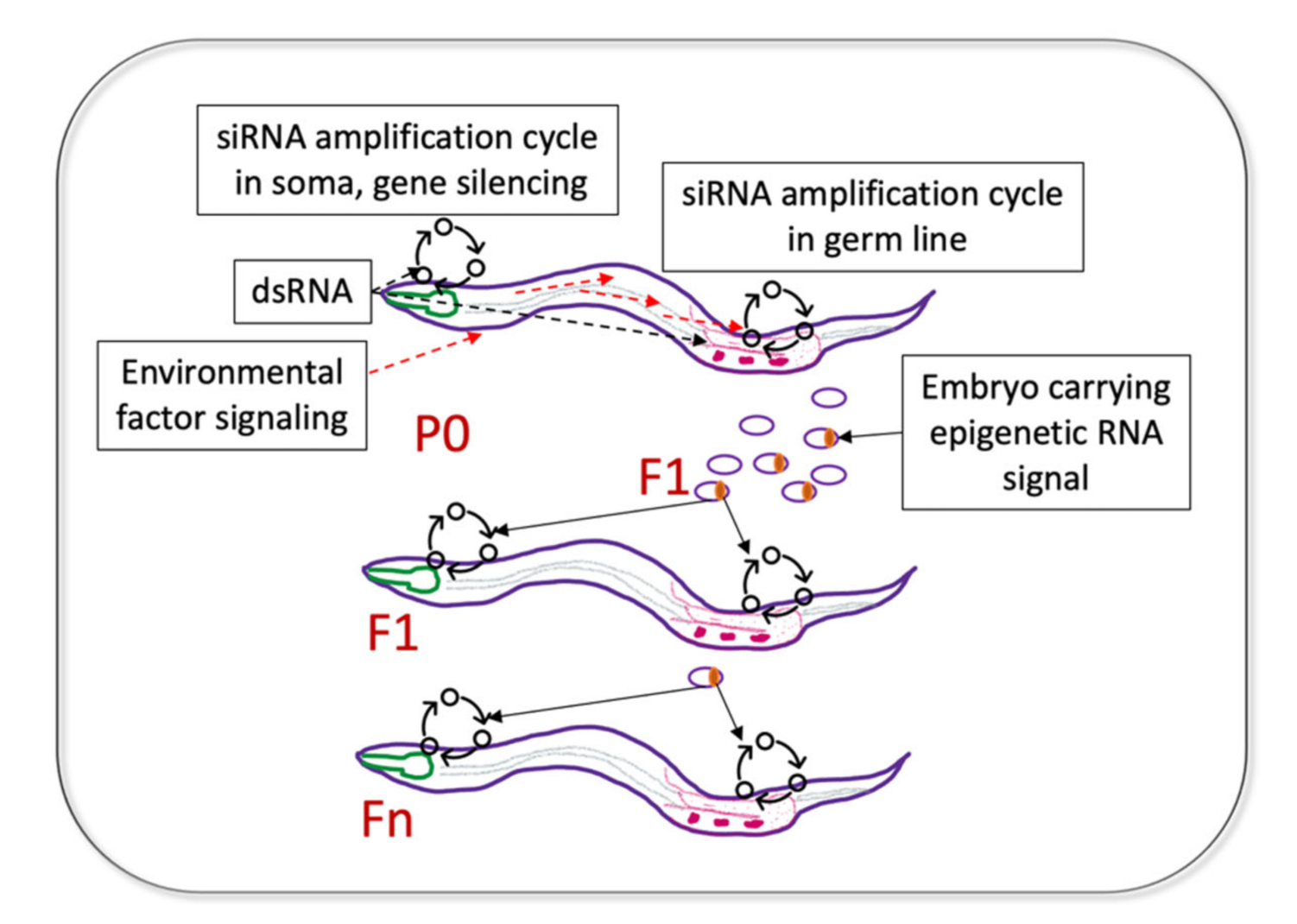
The inheritance of gene silencing by the immediate (F1) progeny of dsRNA-exposed worms was recognized early [16,19]. The F1 generation showed the strongest phenocopy of the corresponding DNA mutations in a number of germline-expressed and soma-expressed genes [16]. Remarkably, their progeny (the F2 generation) completely lost the dsRNA-induced somatic phenotypes [16]. However, subsequent work identified certain somatic genes whose silencing persisted in successive generations [30] (Figure 3).
Now, there are reasons to think that such genes are expressed in both the germline and some somatic tissues. This dual expression requirement for the RNA-induced TEI of somatic gene silencing has been postulated by Minkina and Hunter who studied TEI of the endogenous sid-1 gene [37]. In this example, both germline and somatic silencing of endogenous sid-1 were triggered by a repetitive germline-expressed transgene with homology to the sid-1 locus and were associated with the siRNAs antisense to the sid-1 coding region. Whereas sid-1 silencing in the germline persisted for 8–13 generations, somatic sid-1 silencing was observed for four generations and was dependent on the germline-expressed factors binding heritable amplified small RNAs. Thus, for genes with dual germline and soma expression, a heritable transmission of a favorable adaptive somatic phenotype (e.g., heat shock resistance, pathogen avoidance) is mechanistically possible in C. elegans [37] (Figure 3).
An important feature of the dsRNA-induced silencing in C. elegans is its systemic nature [16]. This is facilitated by the expression of the dsRNA-selective dsRNA-gated channel, SID-1 [65,66]. Thus, dsRNA species expressed through transgenes driven by tissue-specific promoters were used to silence homologous genes in the other tissues [67]. Notably, a GFP-specific dsRNA expressed in neurons triggered a persistent and self-sustaining silencing of a germline GFP transgene [68]. Therefore, it is possible that the communication between the somatic and germline tissues via endogenously produced dsRNA species exists in nematodes and contributes to adaptive TEI (Figure 3). The conservation of the SID-1 protein in mammals [65] suggests a possibility of dsRNA transport in higher organisms as well.
6. Sensory Experiences Communicated to the Germ Line and Transmitted Transgenerationally
There is a growing list of environmental conditions, including starvation [69], dauer diapause [70], and pathogen exposure [71,72,73], which elicit gene expression changes, developmental program variations, and adaptive behaviors that persist in multiple generations. Based on genetic analyses in different experimental systems, the heritable biological responses to environmental changes appear to rely both on dsRNA transport from the soma to the germline [73] and on germline TEI driven by piRNAs and/or siRNA amplification [69,70,72,73]. Most recently, Zaslaver and colleagues designed C. elegans training experiments inducing an associative memory between an odorant and starvation, which was monitored via nuclear translocation of a global stress response factor upon memory reactivation [74]. Remarkably, this association persisted in the progeny of trained worms for two generations and required both RNAi and chromatin regulators [74].
The best mechanistically understood phenomenon has been described by Murphy and colleagues [73] who investigated a specific learning behavior—avoidance of Pseudomonas aeruginosa (PA14)—that persisted for four generations [72]. Remarkably, it was found that C. elegans interpreted a specific PA14 ncRNA transcript as a pathogen signal and built an RNAi-based response to avoid it [73]. The components of this response included the dsRNA transport machinery as well as the piRNA-based germline TEI system. The authors proposed that the complementarity between a section of the bacterial P11 RNA and the mRNA coding for neuronal MACO-1 protein induced an RNAi response and caused MACO-1 downregulation. This, in turn, led to the activation of the TGF-β ligand DAF-7 in a specific neuron, which ultimately caused avoidance behavior. Interestingly, maco-1 mRNA expression has been detected in the germline [75,76], in addition to the neurons. Therefore, the biologically-relevant heritable response of C. elegans to PA14-derived ncRNA that involves maco-1 inhibition is reminiscent of the artificial TEI system where the connection between the somatic and germline silencing of sid-1 has been elucidated ([37] see Section 5) (Figure 3).
7. Epigenetics of Holocentric Centromeres
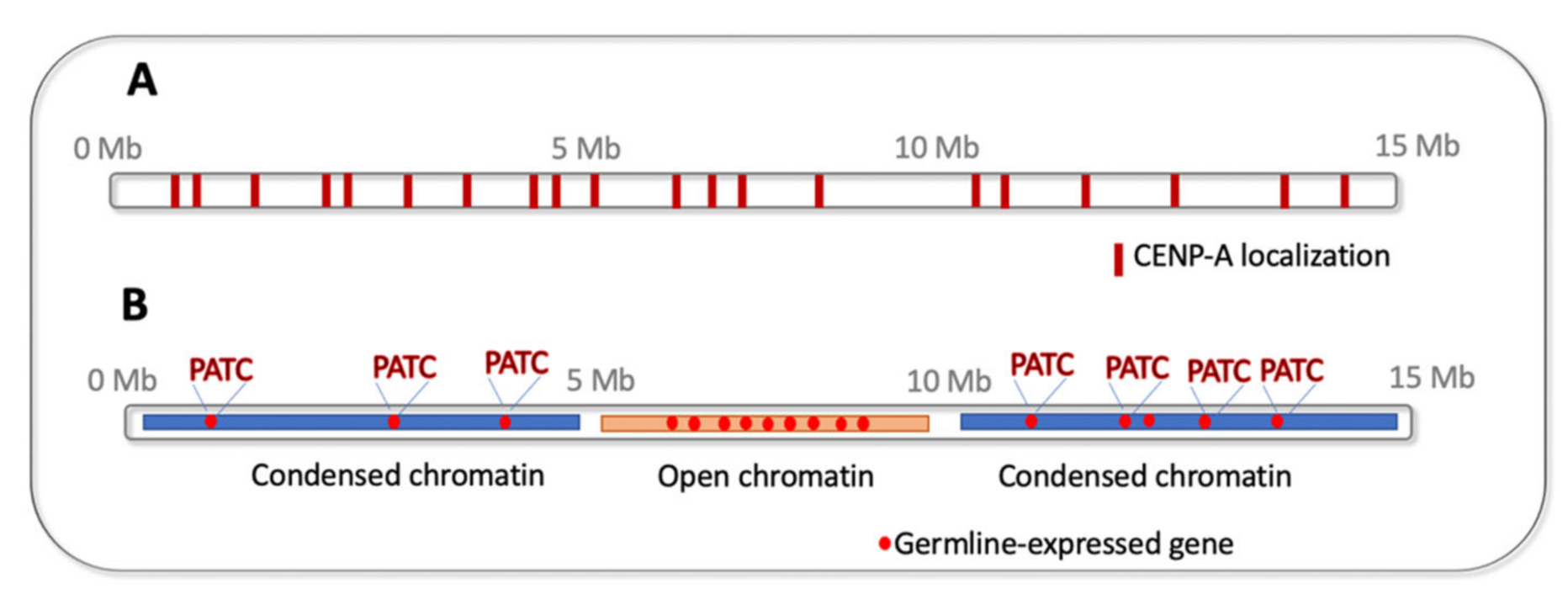
In contrast to the dynamic adaptive TEI phenomena discussed in Section 2, Section 3, Section 4, Section 5, Section 6, there are also very stable epigenetic processes ensuring proper genome maintenance across generations. These include the epigenetic inheritance of the centromeric chromatin positioning. Centromeres are specialized structures on chromosomes that are required for proper microtubule attachment and faithful chromosome segregation. They are marked by the centromere-specific histone variant protein, CENP-A. CENP-A localization on daughter DNA strands is informed by its position on the maternal templates (reviewed in [77]). In most organisms, centromeric chromatin is concentrated in one chromosomal location. The C. elegans chromosomes are holocentric, so CENP-A is distributed in numerous foci along the chromosomes (reviewed in [78]) (Figure 4A). Moreover, in the C. elegans germline there is a disruption of the templated inheritance of CENP-A during meiosis [79]. This begs the question of how the correct chromosomal locations are marked for de novo CENP-A deposition. Although the nature of the positioning mark is still not clear, Steiner and colleagues determined that the de novo placement of CENP-A occurs during a strict developmental window preceding the first embryonic division of the zygote [80]. This process is dependent on the N-terminal tail of CENP-A [80,81]. Surprisingly, the N-terminal tail is dispensable for mitotic divisions during late embryogenesis, larval development, and germline proliferation [80]. Thus, there are distinct steps of initiation and maintenance governing the epigenetic inheritance of CENP-A between generations in C. elegans.
What could serve as a signal for the de novo CENP-A placement in the zygote? Earlier studies from Desai, Strome, and colleagues suggested that the memory of germline transcription is key [82]. Importantly, there are worm antisense siRNAs produced by RdRPs on germline-expressed mRNA templates that are distinct from the silencing siRNAs [83]. They are bound to the Argonaute CSR-1, which is present both in the cytoplasm and the nucleus [84,85]. The nuclear CSR-1 complex is thought to mark genomic regions repelling the silencing chromatin marks [84,86,87], which correlate with CENP-A placement [82,86]. The mechanism of this is not clear. If this scenario is correct, then CENP-A incorporation should proceed by default. There is also the possibility of both repelling and attracting marks guiding the de novo CENP-A pattern establishment.
8. DNA “Watermarks” Allowing Gene Expression in Silenced Chromatin Environment
In C. elegans, “silencing” chromatin marks are enriched on autosome arms [88] whereas the essential genes, including most germline genes, are located at the centers of autosomes [89] (Figure 4B). Therefore, it is thought that the chromosome location (i.e., active or closed chromatin environment) determines whether the gene is expressed in the germline or not. Surprisingly, this is not the whole story. Frøkjær-Jensen and co-authors discovered periodic DNA sequence patterns, namely 10-base pair periodic An/Tn-clusters (PATCs), which are correlated with germline expression of the germline genes located on autosome arms [90] (Figure 4B). Moreover, the inclusion of such sequences on the repetitive transgenic arrays allowed their germline expression. Although the mechanism of this phenomenon is enigmatic, and the Caenorhabditis-specific PATCs periodicity is not seen in more distantly related species [90], there might be other periodic DNA marks that are biologically relevant in other organisms.
9. Concluding Remarks
The recent literature reviewed here underscores the value of basic epigenetics research in a nematode model. More importantly, recent discoveries pose tantalizing questions for future inquiries. The importance of small RNAs in transgenerational transgene silencing and in aberrant progressive silencing of endogenous genes, which underlies the Mrt phenotype, is well documented. However, the mechanistic links between small RNAs and histone-modifying complexes participating in these phenomena are poorly understood and should be investigated in the future. There is still a debate on whether the inappropriate silencing of just one category of repetitive genes, such as the rDNA repeats or histone gene loci, is responsible for the Mrt phenotype of animals lacking piRNAs. It is likely that the silencing of both types of targets contributes to the Mrt sterility. Moreover, it was shown that the Mrt phenotype can be suppressed by insulin signaling pathway mutants [91]. It is not clear whether the insulin signaling pathway regulates small RNAs matching histone genes, rDNA repeats, or something else. It is curious that insulin signaling mutant backgrounds allow a more robust response to experimental RNAi [92] further underscoring the connection to small RNA regulation.
Although the systemic nature of C. elegans RNAi has been known for a long time, and the possibility of silencing information transport from one tissue to another has been shown using transgenes, examples of endogenous RNAi transport are lacking. A recent implication of the dsRNA transport machinery in pathogen avoidance behavior [73] brings hope that the sequence of the dsRNA carrying the avoidance signal and other natural dsRNAs will be uncovered soon.
The participation of small RNAs in the memory of sensory experiences is largely concluded from the experiments with the mutants in various RNAi-related pathways. Since multiple distinct pathways are often involved, it is not easy to imagine specific mechanistic scenarios behind the phenomenology. The challenges in future work will be in dissecting the order of RNAi components’ action and in distinguishing between direct and indirect involvement of small RNAs.
The “licensing” role of small RNAs that do not cause gene silencing but rather protect from it remains enigmatic, especially with regards to nuclear phenomena, despite much progress in the field. Specifically, the mechanistic connection or cooperation between the nuclear CSR-1 pathway and the molecular marks of active chromatin must be understood. Finally, there is still the possibility that PATCs DNA sequences act through RNA intermediates, perhaps small RNAs, to facilitate germline expression of genes located in a closed chromatin environment.
Funding
This research was funded by the National Institute of General Medical Sciences of the National Institutes of Health under award number R01GM135199. The content is solely the responsibility of the author and does not necessarily represent official views of the National Institutes of Health.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Conflicts of Interest
The author declares no conflict of interest.
References
- Corsi, A.K.; Wightman, B.; Chalfie, M. A Transparent Window into Biology: A Primer on Caenorhabditis elegans. Genetics 2015, 200, 387–407. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The, C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef]
- Reinhart, B.J.; Slack, F.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 2000, 403, 901–906. [Google Scholar] [CrossRef]
- Pasquinelli, A.E.; Reinhart, B.J.; Slack, F.; Martindale, M.Q.; Kuroda, M.I.; Maller, B.; Hayward, D.C.; Ball, E.; Degnan, B.; Müller, P.; et al. Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature 2000, 408, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Donkin, I.; Barrès, R. Sperm epigenetics and influence of environmental factors. Mol. Metab. 2018, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rechavi, O.; Lev, I. Principles of Transgenerational Small RNA Inheritance in Caenorhabditis elegans. Curr. Biol. 2017, 27, R720–R730. [Google Scholar] [CrossRef] [Green Version]
- Perez, M.F.; Lehner, B. Intergenerational and transgenerational epigenetic inheritance in animals. Nat. Cell Biol. 2019, 21, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Baugh, L.R.; Day, T. Nongenetic inheritance and multigenerational plasticity in the nematode C. elegans. eLife 2020, 9, e58498. [Google Scholar] [CrossRef]
- Manterola, M.; Palominos, M.F.; Calixto, A. The Heritability of Behaviors Associated with the Host Gut Microbiota. Front. Immunol. 2021, 12, 1497. [Google Scholar] [CrossRef]
- Frolows, N.; Ashe, A. Small RNAs and chromatin in the multigenerational epigenetic landscape of Caenorhabditis elegans. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20200112. [Google Scholar] [CrossRef] [PubMed]
- Minkina, O.; Hunter, C.P. Intergenerational Transmission of Gene Regulatory Information in Caenorhabditis elegans. Trends Genet. 2017, 34, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Kemphues, K.J. par-1, a gene required for establishing polarity in C. elegans embryos, encodes a putative Ser/Thr kinase that is asymmetrically distributed. Cell 1995, 81, 611–620. [Google Scholar] [CrossRef] [Green Version]
- Fire, A.; Albertson, D.; Harrison, S.; Moerman, D. Production of antisense RNA leads to effective and specific inhibition of gene expression in C. elegans muscle. Development 1991, 113, 503–514. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Rocheleau, C.E.; Downs, W.D.; Lin, R.; Wittmann, C.; Bei, Y.; Cha, Y.-H.; Ali, M.; Priess, J.R.; Mello, C.C. Wnt Signaling and an APC-Related Gene Specify Endoderm in Early, C. elegans Embryos. Cell 1997, 90, 707–716. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, A.J. A Species of Small Antisense RNA in Posttranscriptional Gene Silencing in Plants. Science 1999, 286, 950–952. [Google Scholar] [CrossRef] [Green Version]
- Grishok, A.; Tabara, H.; Mello, C.C. Genetic Requirements for Inheritance of RNAi in C.elegans. Science 2000, 287, 2494–2497. [Google Scholar] [CrossRef] [Green Version]
- Smardon, A.; Spoerke, J.M.; Stacey, S.C.; Klein, M.E.; Mackin, N.; Maine, E.M. EGO-1 is related to RNA-directed RNA polymerase and functions in germ-line development and RNA interference in C. elegans. Curr. Biol. 2000, 10, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Grishok, A. Biology and Mechanisms of Short RNAs in Caenorhabditis elegans. Adv. Genet. 2013, 83, 1–69. [Google Scholar] [CrossRef] [PubMed]
- Pak, J.; Fire, A. Distinct Populations of Primary and Secondary Effectors During RNAi in C. elegans. Science 2006, 315, 241–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, H.-Y.; Chen, C.-C.G.; Conte, D.; Moresco, J.; Chaves, D.A.; Mitani, S.; Yates, J.R.; Tsai, M.-D.; Mello, C.C. A Ribonuclease Coordinates siRNA Amplification and mRNA Cleavage during RNAi. Cell 2015, 160, 407–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maniar, J.M.; Fire, A.Z. EGO-1, a C. elegans RdRP, Modulates Gene Expression via Production of mRNA-Templated Short Antisense RNAs. Curr. Biol. 2011, 21, 449–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-C.G.; Simard, M.; Tabara, H.; Brownell, D.R.; McCollough, J.A.; Mello, C.C. A Member of the Polymerase β Nucleotidyltransferase Superfamily Is Required for RNA Interference in C. elegans. Curr. Biol. 2005, 15, 378–383. [Google Scholar] [CrossRef] [Green Version]
- Shukla, A.; Yan, J.; Pagano, D.J.; Dodson, A.E.; Fei, Y.; Gorham, J.; Seidman, J.G.; Wickens, M.; Kennedy, S. poly (UG)-tailed RNAs in genome protection and epigenetic inheritance. Nature 2020, 582, 283–288. [Google Scholar] [CrossRef]
- Preston, M.; Porter, D.F.; Chen, F.; Buter, N.; Lapointe, C.P.; Keles, S.; Kimble, J.; Wickens, M. Unbiased screen of RNA tailing activities reveals a poly (UG) polymerase. Nat. Methods 2019, 16, 437–445. [Google Scholar] [CrossRef]
- Alcazar, R.M.; Lin, R.; Fire, A.Z. Transmission Dynamics of Heritable Silencing Induced by Double-Stranded RNA in Caenorhabditis elegans. Genetics 2008, 180, 1275–1288. [Google Scholar] [CrossRef] [Green Version]
- Praitis, V.; Casey, E.; Collar, D.; Austin, J. Creation of Low-Copy Integrated Transgenic Lines in Caenorhabditis elegans. Genetics 2001, 157, 1217–1226. [Google Scholar] [CrossRef]
- Vastenhouw, N.L.; Brunschwig, K.; Okihara, K.L.; Müller, F.; Tijsterman, M.; Plasterk, R.H.A. Long-term gene silencing by RNAi. Nature 2006, 442, 882. [Google Scholar] [CrossRef]
- Buckley, B.A.; Burkhart, K.B.; Gu, S.G.; Spracklin, G.; Kershner, A.; Fritz, H.; Kimble, J.; Fire, A.; Kennedy, S. A nuclear Argonaute promotes multigenerational epigenetic inheritance and germline immortality. Nature 2012, 489, 447–451. [Google Scholar] [CrossRef] [Green Version]
- Frøkjær-Jensen, C.; Davis, M.W.; Hopkins, C.E.; Newman, B.J.; Thummel, J.M.; Olesen, S.-P.; Grunnet, M.; Jorgensen, E.M. Single-copy insertion of transgenes in Caenorhabditis elegans. Nat. Genet. 2008, 40, 1375–1383. [Google Scholar] [CrossRef] [Green Version]
- Frøkjær-Jensen, C.; Davis, M.; Ailion, M.; Jorgensen, E.M. Improved Mos1-mediated transgenesis in C. elegans. Nat. Methods 2012, 9, 117–118. [Google Scholar] [CrossRef]
- Ashe, A.; Sapetschnig, A.; Weick, E.-M.; Mitchell, J.; Bagijn, M.P.; Cording, A.C.; Doebley, A.-L.; Goldstein, L.D.; Lehrbach, N.; Le Pen, J.; et al. piRNAs Can Trigger a Multigenerational Epigenetic Memory in the Germline of C. elegans. Cell 2012, 150, 88–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodhouse, R.; Buchmann, G.; Hoe, M.; Harney, D.J.; Low, J.K.; Larance, M.; Boag, P.; Ashe, A. Chromatin Modifiers SET-25 and SET-32 Are Required for Establishment but Not Long-Term Maintenance of Transgenerational Epigenetic Inheritance. Cell Rep. 2018, 25, 2259–2272.e5. [Google Scholar] [CrossRef] [Green Version]
- Shirayama, M.; Seth, M.; Lee, H.-C.; Gu, W.; Ishidate, T.; Conte, D.; Mello, C.C. piRNAs Initiate an Epigenetic Memory of Nonself RNA in the C. elegans Germline. Cell 2012, 150, 65–77. [Google Scholar] [CrossRef] [Green Version]
- Minkina, O.; Hunter, C.P. Stable Heritable Germline Silencing Directs Somatic Silencing at an Endogenous Locus. Mol. Cell 2017, 65, 659–670.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luteijn, M.J.; Van Bergeijk, P.; Kaaij, L.J.T.; Almeida, M.V.; Roovers, E.F.; Berezikov, E.; Ketting, R.F. Extremely stable Piwi-induced gene silencing in Caenorhabditis elegans. EMBO J. 2012, 31, 3422–3430. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-C.; Gu, W.; Shirayama, M.; Youngman, E.; Conte, D.; Mello, C.C. C. elegans piRNAs Mediate the Genome-wide Surveillance of Germline Transcripts. Cell 2012, 150, 78–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagijn, M.P.; Goldstein, L.D.; Sapetschnig, A.; Weick, E.-M.; Bouasker, S.; Lehrbach, N.; Simard, M.; Miska, E.A. Function, Targets, and Evolution of Caenorhabditis elegans piRNAs. Science 2012, 337, 574–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guang, S.; Bochner, A.F.; Burkhart, K.B.; Burton, N.; Pavelec, D.M.; Kennedy, S. Small regulatory RNAs inhibit RNA polymerase II during the elongation phase of transcription. Nature 2010, 465, 1097–1101. [Google Scholar] [CrossRef] [Green Version]
- Gu, S.G.; Pak, J.; Guang, S.; Maniar, J.M.; Kennedy, S.; Fire, A. Amplification of siRNA in Caenorhabditis elegans generates a transgenerational sequence-targeted histone H3 lysine 9 methylation footprint. Nat. Genet. 2012, 44, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Kalinava, N.; Ni, J.Z.; Gajic, Z.; Kim, M.; Ushakov, H.; Gu, S.G. C. elegans Heterochromatin Factor SET-32 Plays an Essential Role in Transgenerational Establishment of Nuclear RNAi-Mediated Epigenetic Silencing. Cell Rep. 2018, 25, 2273–2284.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grishok, A.; Sinskey, J.L.; Sharp, P.A. Transcriptional silencing of a transgene by RNAi in the soma of C. elegans. Genes Dev. 2005, 19, 683–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, N.; Burkhart, K.B.; Kennedy, S. Nuclear RNAi maintains heritable gene silencing in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2011, 108, 19683–19688. [Google Scholar] [CrossRef] [Green Version]
- Rechavi, O.; Minevich, G.; Hobert, O. Transgenerational Inheritance of an Acquired Small RNA-Based Antiviral Response in C. elegans. Cell 2011, 147, 1248–1256. [Google Scholar] [CrossRef] [Green Version]
- Sundby, A.E.; Molnar, R.I.; Claycomb, J.M. Connecting the Dots: Linking Caenorhabditis elegans Small RNA Pathways and Germ Granules. Trends Cell Biol. 2021, 31, 387–401. [Google Scholar] [CrossRef]
- Pitt, J.N.; Schisa, J.A.; Priess, J.R. P Granules in the Germ Cells of Caenorhabditis elegans Adults Are Associated with Clusters of Nuclear Pores and Contain RNA. Dev. Biol. 2000, 219, 315–333. [Google Scholar] [CrossRef] [Green Version]
- Schreier, J.; Dietz, S.; de Jesus Domingues, A.M.; Seistrup, A.-S.; Nguyen, D.A.H.; Gleason, E.J.; Ling, H.; L’Hernault, S.W.; Phillips, C.M.; Butter, F.; et al. A Membrane-Associated Condensate Drives Paternal Epigenetic Inheritance in C. Elegans. bioRxiv 2020. [Google Scholar] [CrossRef]
- Kelly, W.G.; Xu, S.; Montgomery, M.K.; Fire, A. Distinct Requirements for Somatic and Germline Expression of a Generally Expressed Caernorhabditis elegans Gene. Genetics 1997, 146, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Tabara, H.; Sarkissian, M.; Kelly, W.G.; Fleenor, J.; Grishok, A.; Timmons, L.; Fire, A.; Mello, C.C. The rde-1 Gene, RNA Interference, and Transposon Silencing in C. elegans. Cell 1999, 99, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Sakaguchi, A.; Sarkies, P.; Simon, M.; Doebley, A.-L.; Goldstein, L.D.; Hedges, A.; Ikegami, K.; Alvares, S.M.; Yang, L.; LaRocque, J.; et al. Caenorhabditis elegans RSD-2 and RSD-6 promote germ cell immortality by maintaining small interfering RNA populations. Proc. Natl. Acad. Sci. USA 2014, 111, E4323–E4331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devanapally, S.; Raman, P.; Chey, M.; Allgood, S.; Ettefa, F.; Diop, M.; Lin, Y.; Cho, Y.E.; Jose, A.M. Mating can initiate stable RNA silencing that overcomes epigenetic recovery. Nat. Commun. 2021, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Brennecke, J.; Malone, C.D.; Aravin, A.A.; Sachidanandam, R.; Stark, A.; Hannon, G.J. An Epigenetic Role for Maternally Inherited piRNAs in Transposon Silencing. Science 2008, 322, 1387–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, S.; Hodgkin, J. MRT-2 checkpoint protein is required for germline immortality and telomere replication in C. elegans. Nature 2000, 403, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Barucci, G.; Cornes, E.; Singh, M.; Li, B.; Ugolini, M.; Samolygo, A.; Didier, C.; Dingli, F.; Loew, D.; Quarato, P.; et al. Small-RNA-mediated transgenerational silencing of histone genes impairs fertility in piRNA mutants. Nature 2020, 22, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Reed, K.J.; Svendsen, J.M.; Brown, K.C.; Montgomery, E.B.; Marks, T.N.; Vijayasarathy, T.; Parker, D.M.; Nishimura, E.O.; Updike, D.L.; Montgomery, A.T. Widespread roles for piRNAs and WAGO-class siRNAs in shaping the germline transcriptome of Caenorhabditis elegans. Nucleic Acids Res. 2019, 48, 1811–1827. [Google Scholar] [CrossRef] [Green Version]
- Wahba, L.; Hansen, L.; Fire, A.Z. An essential role for the piRNA pathway in regulating the ribosomal RNA pool in C. elegans. Dev. Cell 2021, 56, 2295–2312.e6. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, X.; Wang, Y.; Feng, X.; Guang, S. A new layer of rRNA regulation by small interference RNAs and the nuclear RNAi pathway. RNA Biol. 2017, 14, 1492–1498. [Google Scholar] [CrossRef] [Green Version]
- Perales, R.; Pagano, D.; Wan, G.; Fields, B.D.; Saltzman, A.L.; Kennedy, S.G. Transgenerational Epigenetic Inheritance Is Negatively Regulated by the HERI-1 Chromodomain Protein. Genetics 2018, 210, 1287–1299. [Google Scholar] [CrossRef] [Green Version]
- Lev, I.; Seroussi, U.; Gingold, H.; Bril, R.; Anava, S.; Rechavi, O. MET-2-Dependent H3K9 Methylation Suppresses Transgenerational Small RNA Inheritance. Curr. Biol. 2017, 27, 1138–1147. [Google Scholar] [CrossRef] [Green Version]
- Shukla, A.; Perales, R.; Kennedy, S. PiRNAs Coordinate Poly (UG) Tailing to Prevent Aberrant and Permanent Gene Silencing. bioRxiv 2021. [Google Scholar] [CrossRef]
- Frøkjær-Jensen, C.; (King Abdullah University of Science and Technology, Thuwal, Saudi Arabia). Personal communication, 2021.
- Klattenhoff, C.; Theurkauf, W. Biogenesis and germline functions of piRNAs. Development 2008, 135, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Winston, W.M.; Molodowitch, C.; Hunter, C.P. Systemic RNAi in C. elegans Requires the Putative Transmembrane Protein SID-1. Science 2002, 295, 2456–2459. [Google Scholar] [CrossRef] [Green Version]
- Shih, J.D.; Hunter, C.P. SID-1 is a dsRNA-selective dsRNA-gated channel. RNA 2011, 17, 1057–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jose, A.M.; Smith, J.J.; Hunter, C.P. Export of RNA silencing from C. elegans tissues does not require the RNA channel SID-1. Proc. Natl. Acad. Sci. USA 2009, 106, 2283–2288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devanapally, S.; Ravikumar, S.; Jose, A.M. Double-stranded RNA made in C. elegans neurons can enter the germline and cause transgenerational gene silencing. Proc. Natl. Acad. Sci. USA 2015, 112, 2133–2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rechavi, O.; Houri-Ze’Evi, L.; Anava, S.; Goh, S.; Kerk, S.Y.; Hannon, G.J.; Hobert, O. Starvation-Induced Transgenerational Inheritance of Small RNAs in C. elegans. Cell 2014, 158, 277–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewe, C.K.; Cleuren, Y.N.T.; Flowers, S.E.; Alok, G.; Snell, R.G.; Rothman, J.H. Natural cryptic variation in epigenetic modulation of an embryonic gene regulatory network. Proc. Natl. Acad. Sci. USA 2020, 117, 13637–13646. [Google Scholar] [CrossRef]
- Palominos, M.F.; Verdugo, L.; Gabaldon, C.; Pollak, B.; Ortíz-Severín, J.; Varas, M.A.; Chávez, F.P.; Calixto, A. Transgenerational Diapause as an Avoidance Strategy against Bacterial Pathogens in Caenorhabditis elegans. mBio 2017, 8, e01234-17. [Google Scholar] [CrossRef] [Green Version]
- Moore, R.S.; Kaletsky, R.; Murphy, C.T. Piwi/PRG-1 Argonaute and TGF-β Mediate Transgenerational Learned Pathogenic Avoidance. Cell 2019, 177, 1827–1841.e12. [Google Scholar] [CrossRef]
- Kaletsky, R.; Moore, R.S.; Vrla, G.D.; Parsons, L.R.; Gitai, Z.; Murphy, C.T. C. elegans interprets bacterial non-coding RNAs to learn pathogenic avoidance. Nature 2020, 586, 445–451. [Google Scholar] [CrossRef]
- Deshe, N.; Eliezer, Y.; Hoch, L.; Itskovits, E.; Ben-Ezra, S.; Zaslaver, A. Inheritance of Associative Memories in C. Elegans Nematodes. bioRxiv 2020. [Google Scholar] [CrossRef]
- Han, S.; Schroeder, E.A.; Silva-García, C.G.; Hebestreit, K.; Mair, W.B.; Brunet, A. Mono-unsaturated fatty acids link H3K4me3 modifiers to C. elegans lifespan. Nature 2017, 544, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Grün, D.; Kirchner, M.; Thierfelder, N.; Stoeckius, M.; Selbach, M.; Rajewsky, N. Conservation of mRNA and Protein Expression during Development of C. elegans. Cell Rep. 2014, 6, 565–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stellfox, M.E.; Bailey, A.O.; Foltz, D.R. Putting CENP-A in its place. Experientia 2012, 70, 387–406. [Google Scholar] [CrossRef] [Green Version]
- Maddox, P.S.; Oegema, K.; Desai, A.; Cheeseman, I.M. Holoer than thou: Chromosome segregation and kinetochore function in C. elegans. Chromosom. Res. 2004, 12, 641–653. [Google Scholar] [CrossRef]
- Monen, J.; Maddox, P.S.; Hyndman, F.; Oegema, K.; Desai, A. Differential role of CENP-A in the segregation of holocentric C. elegans chromosomes during meiosis and mitosis. Nature 2005, 7, 1248–1255. [Google Scholar] [CrossRef]
- Prosée, R.F.; Wenda, J.M.; Özdemir, I.; Gabus, C.; Delaney, K.; Schwager, F.; Gotta, M.; Steiner, F.A. Transgenerational inheritance of centromere identity requires the CENP-A N-terminal tail in the C. elegans maternal germ line. PLoS Biol. 2021, 19, e3000968. [Google Scholar] [CrossRef]
- De Groot, C.; Houston, J.; Davis, B.; Gerson-Gurwitz, A.; Monen, J.; Lara-Gonzalez, P.; Oegema, K.; Shiau, A.K.; Desai, A. The N-terminal Tail of C. elegans CENP-A Interacts with KNL-2 and is Essential for Centromeric Chromatin Assembly. Mol. Biol. Cell 2021, 32, mbc.E20–12. [Google Scholar] [CrossRef]
- Gassmann, R.; Rechtsteiner, A.; Yuen, K.W.; Muroyama, A.; Egelhofer, T.; Gaydos, L.; Barron, F.; Maddox, P.; Essex, A.; Monen, J.; et al. An inverse relationship to germline transcription defines centromeric chromatin in C. elegans. Nature 2012, 484, 534–537. [Google Scholar] [CrossRef] [Green Version]
- Gu, W.; Shirayama, M.; Conte, D.; Vasale, J.; Batista, P.J.; Claycomb, J.; Moresco, J.; Youngman, E.M.; Keys, J.; Stoltz, M.J.; et al. Distinct Argonaute-Mediated 22G-RNA Pathways Direct Genome Surveillance in the C. elegans Germline. Mol. Cell 2009, 36, 231–244. [Google Scholar] [CrossRef] [Green Version]
- Claycomb, J.; Batista, P.J.; Pang, K.M.; Gu, W.; Vasale, J.J.; van Wolfswinkel, J.C.; Chaves, D.A.; Shirayama, M.; Mitani, S.; Ketting, R.F.; et al. The Argonaute CSR-1 and Its 22G-RNA Cofactors Are Required for Holocentric Chromosome Segregation. Cell 2009, 139, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Avgousti, D.C.; Palani, S.; Sherman, Y.; Grishok, A. CSR-1 RNAi pathway positively regulates histone expression in C. elegans. EMBO J. 2012, 31, 3821–3832. [Google Scholar] [CrossRef] [Green Version]
- Cecere, G.; Hoersch, S.; O’Keeffe, S.; Sachidanandam, R.; Grishok, A. Global effects of the CSR-1 RNA interference pathway on the transcriptional landscape. Nat. Struct. Mol. Biol. 2014, 21, 358–365. [Google Scholar] [CrossRef] [Green Version]
- Gushchanskaia, E.S.; Esse, R.; Ma, Q.; Lau, N.C.; Grishok, A. Interplay between small RNA pathways shapes chromatin landscapes in C. elegans. Nucleic Acids Res. 2019, 47, 5603–5616. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Rechtsteiner, A.; Egelhofer, T.A.; Vielle, A.; Latorre, I.; Cheung, M.-S.; Ercan, S.; Ikegami, K.; Jensen, M.; Kolasinska-Zwierz, P.; et al. Broad chromosomal domains of histone modification patterns in C. elegans. Genome Res. 2010, 21, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Campos, T.L.; Korhonen, P.K.; Sternberg, P.W.; Gasser, R.B.; Young, N.D. Predicting gene essentiality in Caenorhabditis elegans by feature engineering and machine-learning. Comput. Struct. Biotechnol. J. 2020, 18, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Frøkjær-Jensen, C.; Jain, N.; Hansen, L.; Davis, M.W.; Li, Y.; Zhao, D.; Rebora, K.; Millet, J.; Liu, X.; Kim, S.K.; et al. An Abundant Class of Non-coding DNA Can Prevent Stochastic Gene Silencing in the C. elegans Germline. Cell 2016, 166, 343–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, M.; Sarkies, P.; Ikegami, K.; Doebley, A.-L.; Goldstein, L.D.; Mitchell, J.; Sakaguchi, A.; Miska, E.A.; Ahmed, S. Reduced Insulin/IGF-1 Signaling Restores Germ Cell Immortality to Caenorhabditis elegans Piwi Mutants. Cell Rep. 2014, 7, 762–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Ruvkun, G. Regulation of Caenorhabditis elegans RNA Interference by the daf-2 Insulin Stress and Longevity Signaling Pathway. Cold Spring Harb. Symp. Quant. Biol. 2004, 69, 429–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Artistic representation of the adult Caenorhabditis elegans hermaphrodite and a young larva crawling on a plate. Image of the oil painting “Red” by Blanca Craven-Bartle (2015).
Figure 1.
Artistic representation of the adult Caenorhabditis elegans hermaphrodite and a young larva crawling on a plate. Image of the oil painting “Red” by Blanca Craven-Bartle (2015).

Figure 2.
Illustration of the transgenerational epigenetic inheritance steps in the C. elegans germline: initiation via double-stranded RNA (dsRNA), which is processed by the Dicer complex into primary small interfering RNAs (siRNAs) bound by Argonaute (AGO) or via Piwi-interacting RNA (piRNA) bound to PIWI; amplification of siRNAs on mRNA templates bound by AGO or PIWI, which includes non-templated UGn addition by the nucleotidyltransferase RDE-3 followed by secondary siRNA production by RNA-dependent RNA polymerase (RdRP). Secondary siRNAs are bound by AGO that are distinct from those binding primary RNAs. Secondary siRNA/AGO complexes induce either post-transcriptional (PTGS) or transcriptional (TGS) gene silencing. Secondary siRNAs are thought to be heritable and capable of initiating the siRNA amplification cycle in the germline of progeny.
Figure 2.
Illustration of the transgenerational epigenetic inheritance steps in the C. elegans germline: initiation via double-stranded RNA (dsRNA), which is processed by the Dicer complex into primary small interfering RNAs (siRNAs) bound by Argonaute (AGO) or via Piwi-interacting RNA (piRNA) bound to PIWI; amplification of siRNAs on mRNA templates bound by AGO or PIWI, which includes non-templated UGn addition by the nucleotidyltransferase RDE-3 followed by secondary siRNA production by RNA-dependent RNA polymerase (RdRP). Secondary siRNAs are bound by AGO that are distinct from those binding primary RNAs. Secondary siRNA/AGO complexes induce either post-transcriptional (PTGS) or transcriptional (TGS) gene silencing. Secondary siRNAs are thought to be heritable and capable of initiating the siRNA amplification cycle in the germline of progeny.

Figure 3.
Possible mechanisms of heritable transmission of adaptive responses for genes with dual germline and soma expression, based on published work. dsRNA segments found in bacterial ncRNA can initiate a physiological response in tissues expressing mRNAs complementary to dsRNA (siRNA amplification in soma, gene silencing). At the same time, the ingested dsRNA can travel to the germline and initiate an siRNA amplification cycle on the same mRNAs. The heritable epigenetic RNA signal (likely secondary siRNA) is deposited in the F1 embryos; when these siRNAs are distributed to tissues expressing their mRNA targets, they initiate new siRNA amplification cycles. The initiation of gene silencing in the target somatic tissue (e.g., neuron) leads to the phenotypic manifestation of the adaptive response in F1 (e.g., pathogen avoidance), whereas amplification of siRNAs on the same mRNAs expressed in the germline leads to propagation of the adaptive responses through generations. Note that the environmental signal sensed by the P0 worms does not need to be dsRNA, but this signal has to ultimately induce the siRNA amplification cycle in the germline.
Figure 3.
Possible mechanisms of heritable transmission of adaptive responses for genes with dual germline and soma expression, based on published work. dsRNA segments found in bacterial ncRNA can initiate a physiological response in tissues expressing mRNAs complementary to dsRNA (siRNA amplification in soma, gene silencing). At the same time, the ingested dsRNA can travel to the germline and initiate an siRNA amplification cycle on the same mRNAs. The heritable epigenetic RNA signal (likely secondary siRNA) is deposited in the F1 embryos; when these siRNAs are distributed to tissues expressing their mRNA targets, they initiate new siRNA amplification cycles. The initiation of gene silencing in the target somatic tissue (e.g., neuron) leads to the phenotypic manifestation of the adaptive response in F1 (e.g., pathogen avoidance), whereas amplification of siRNAs on the same mRNAs expressed in the germline leads to propagation of the adaptive responses through generations. Note that the environmental signal sensed by the P0 worms does not need to be dsRNA, but this signal has to ultimately induce the siRNA amplification cycle in the germline.

Figure 4.
Epigenetic features of C. elegans chromosomes (an autosome schematic is shown): (A) distribution of CENP-A; (B) open chromatin on autosome centers and condensed chromatin on autosome arms, germline-expressed genes located on autosome arms contain 10-base pair periodic An/Tn-clusters (PATCs) that facilitate their activation.
Figure 4.
Epigenetic features of C. elegans chromosomes (an autosome schematic is shown): (A) distribution of CENP-A; (B) open chromatin on autosome centers and condensed chromatin on autosome arms, germline-expressed genes located on autosome arms contain 10-base pair periodic An/Tn-clusters (PATCs) that facilitate their activation.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Grishok, A. Small RNAs Worm Up Transgenerational Epigenetics Research. DNA 2021, 1, 37-48. https://0-doi-org.brum.beds.ac.uk/10.3390/dna1020005
AMA Style
Grishok A. Small RNAs Worm Up Transgenerational Epigenetics Research. DNA. 2021; 1(2):37-48. https://0-doi-org.brum.beds.ac.uk/10.3390/dna1020005
Chicago/Turabian StyleGrishok, Alla. 2021. "Small RNAs Worm Up Transgenerational Epigenetics Research" DNA 1, no. 2: 37-48. https://0-doi-org.brum.beds.ac.uk/10.3390/dna1020005