
Evaluation and Development of a Nutrition Model to Predict Intake and Growth of Suckling Calves

Abstract
:1. Introduction
2. Materials and Methods
2.1. Base Model
2.2. Model Evaluation Data
2.2.1. Dairy Calf Intake and Body Weight Dataset
2.2.2. Beef Calf Intake and Body Weight Dataset
2.3. Model Adjustment Data
2.3.1. Calf Growth
2.3.2. Forage Digestibility
2.3.3. Milk Composition
2.4. Model Evaluation
2.4.1. Evaluation Metrics
2.4.2. Milk Intake
2.4.3. Forage Intake
2.4.4. Body Weight
3. Results
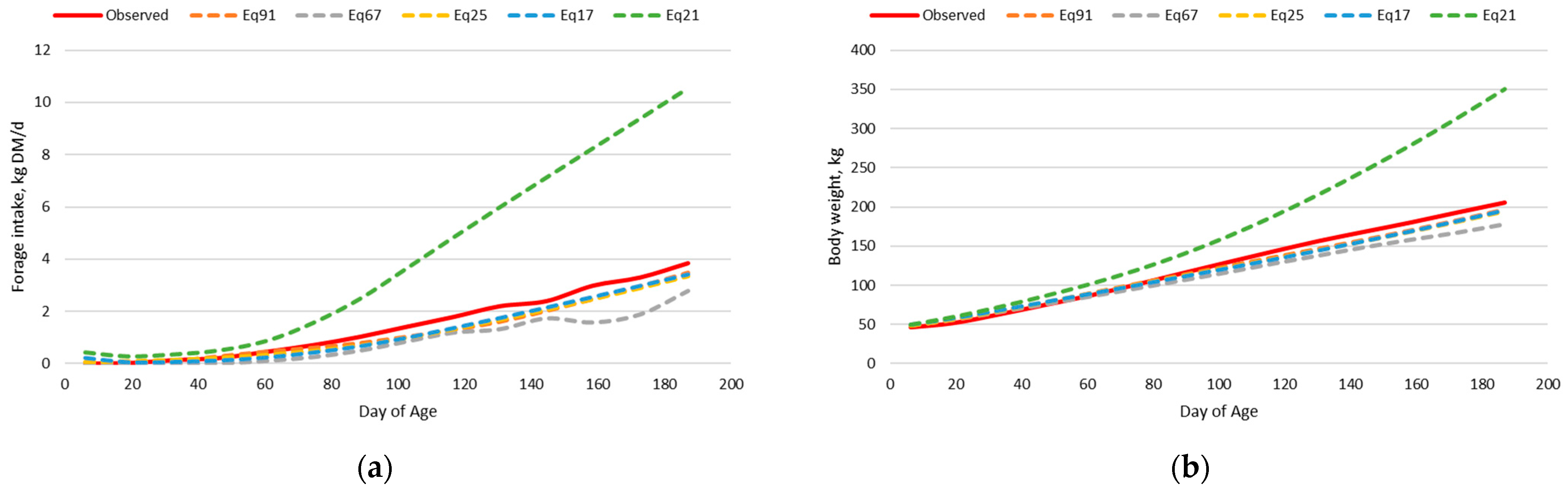
3.1. Dairy Calf Intake and Body Weight Dataset
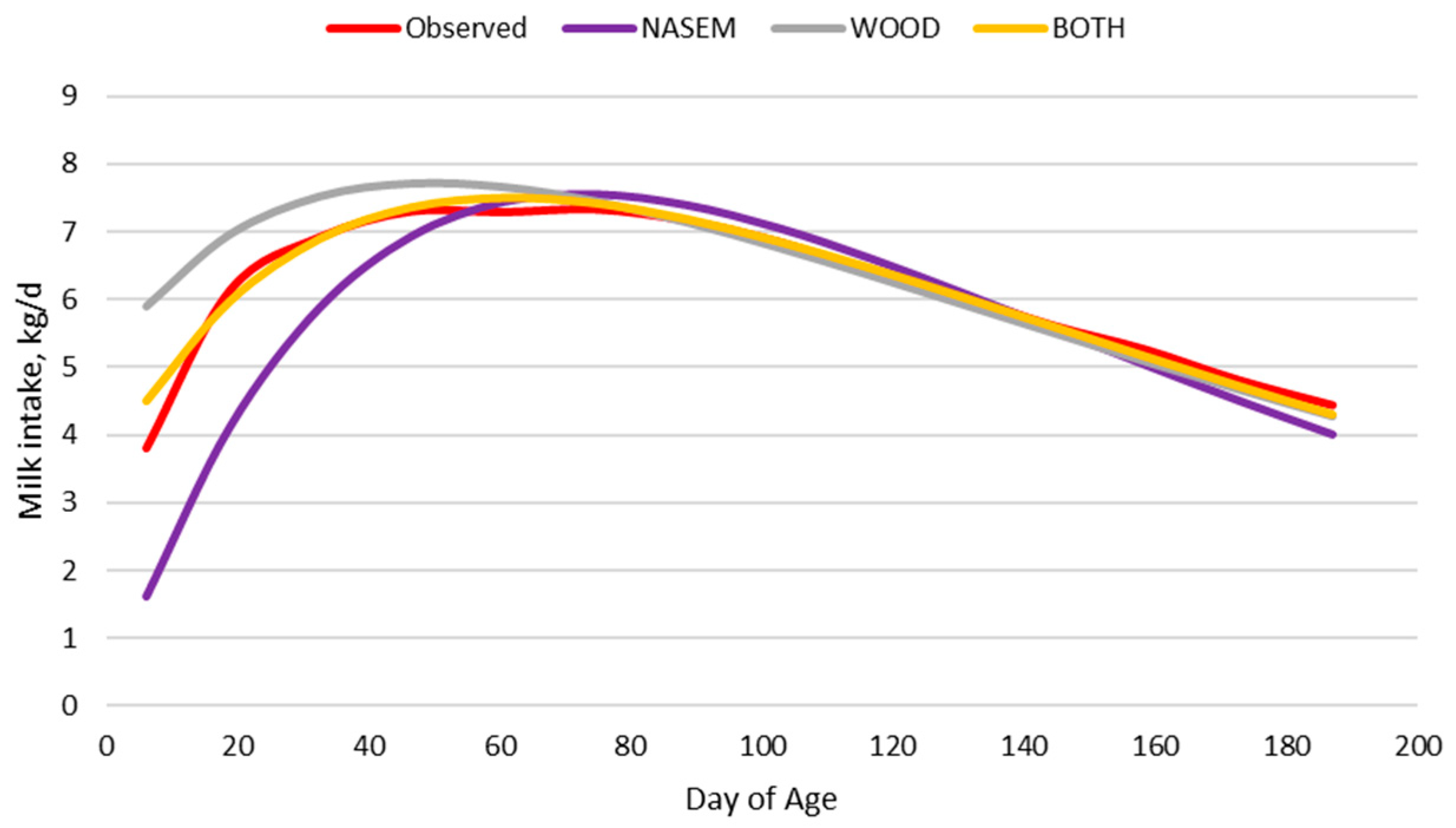
3.1.1. Milk Intake
3.1.2. Original Nutrition Model
3.1.3. Adjusted Nutrition Model
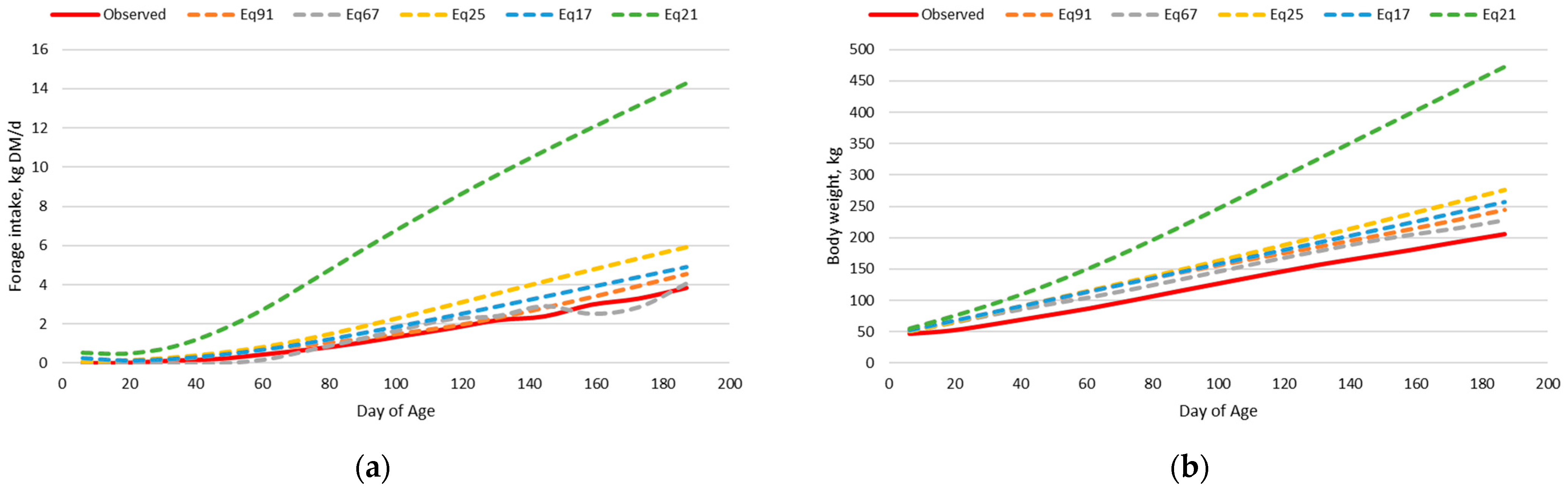
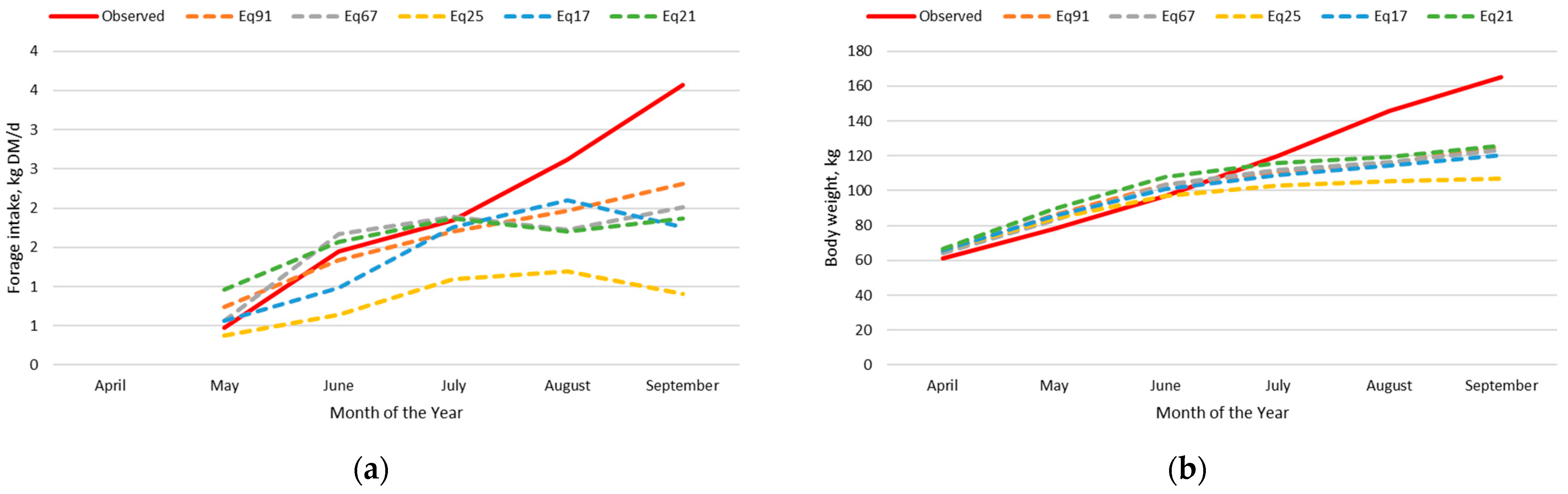
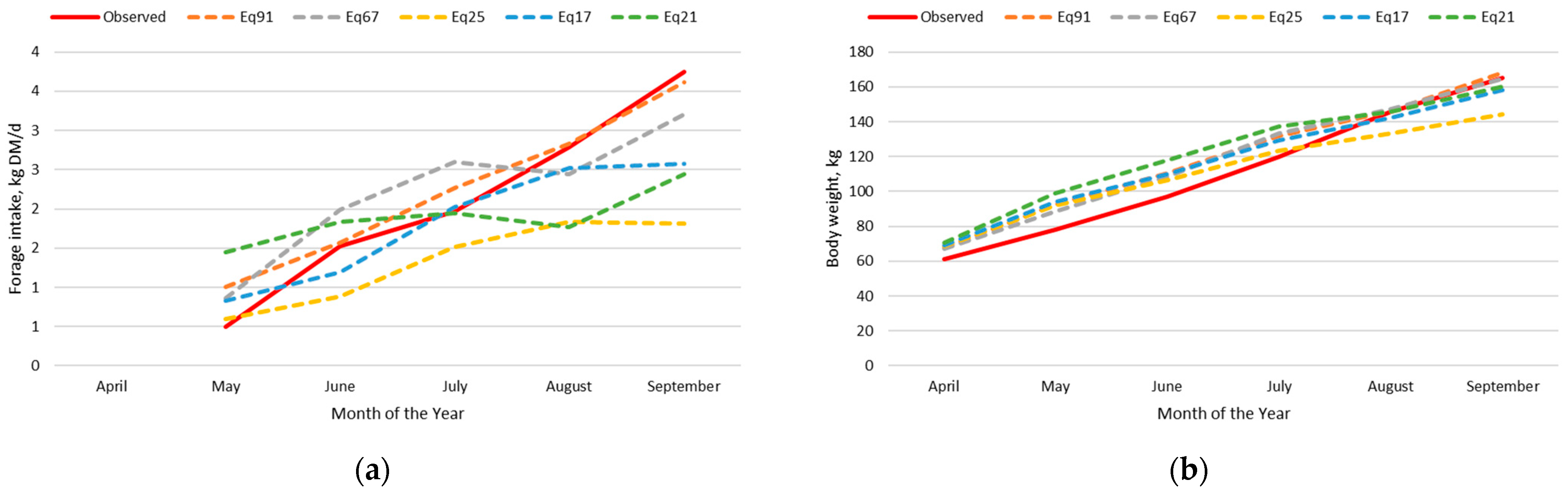
3.2. Beef Calf Intake and Body Weight Dataset
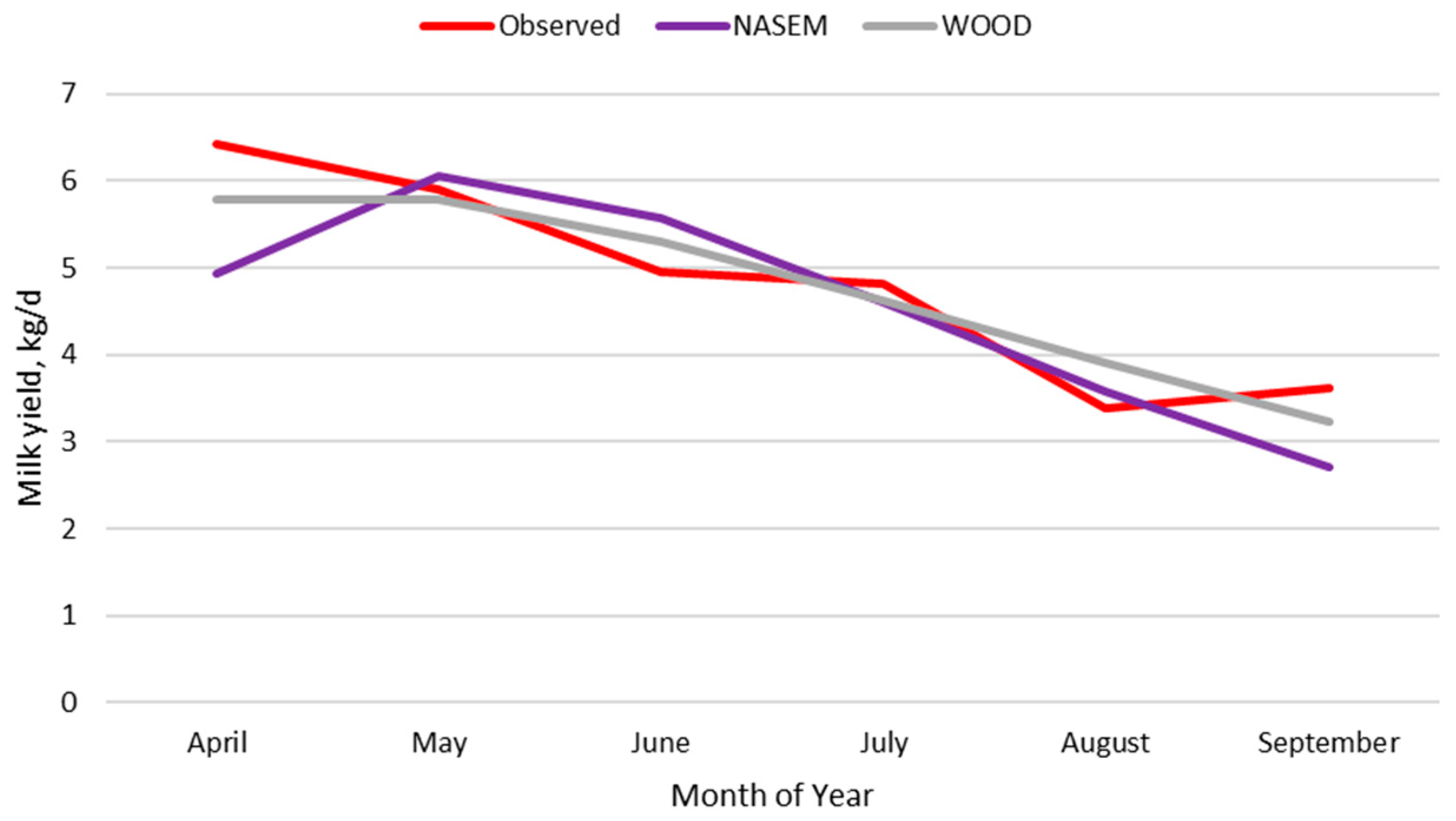
3.2.1. Milk Intake
3.2.2. Original Nutrition Model
3.2.3. Adjusted Nutrition Model
3.2.4. Exploration of Model Deviation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rotz, C.A.; Asem-Hiablie, S.; Place, S.; Thoma, G. Environmental Footprints of Beef Cattle Production in the United States. Agric. Syst. 2019, 169, 1–13. [Google Scholar] [CrossRef]
- Stanley, P.L.; Rowntree, J.E.; Beede, D.K.; DeLonge, M.S.; Hamm, M.W. Impacts of Soil Carbon Sequestration on Life Cycle Greenhouse Gas Emissions in Midwestern USA Beef Finishing Systems. Agric. Syst. 2018, 162, 249–258. [Google Scholar] [CrossRef]
- Day, M.E.; Tang, M.; Lancaster, P.A.; Presley, D.; Pendell, D.L.; Fick, W.H.; Doro, L.; Ahlers, A.; Ricketts, A. Simulation of the Impact of Rangeland Management Strategies on Soil Health, Environmental Footprint, Economic Impact, and Human-Edible Nutrient Conversion from Grasslands in the Central and Northern Great Plains of the United States. Sustainability 2023, 15, 12456. [Google Scholar] [CrossRef]
- Hirooka, H. Systems Approaches to Beef Cattle Production Systems Using Modeling and Simulation. Anim. Sci. J. 2010, 81, 411–424. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-Y.; Ale, S.; Teague, W.R.; Jeong, J. Evaluating the Ranch and Watershed Scale Impacts of Using Traditional and Adaptive Multi-Paddock Grazing on Runoff, Sediment and Nutrient Losses in North Texas, USA. Agric. Ecosyst. Environ. 2017, 240, 32–44. [Google Scholar] [CrossRef]
- Wang, T.; Teague, W.R.; Park, S.C. Evaluation of Continuous and Multipaddock Grazing on Vegetation and Livestock Performance—a Modeling Approach. Rangel. Ecol. Manag. 2016, 69, 457–464. [Google Scholar] [CrossRef]
- Fang, Q.X.; Harmel, R.D.; Ma, L.; Bartling, P.N.S.; Derner, J.D.; Jeong, J.; Williams, J.R.; Boone, R.B. Evaluating the APEX Model for Alternative Cow-Calf Grazing Management Strategies in Central Texas. Agric. Syst. 2022, 195, 103287. [Google Scholar] [CrossRef]
- Cheng, G.; Harmel, R.D.; Ma, L.; Derner, J.D.; Augustine, D.J.; Bartling, P.N.S.; Fang, Q.X.; Williams, J.R.; Zilverberg, C.J.; Boone, R.B.; et al. Evaluation of APEX Modifications to Simulate Forage Production for Grazing Management Decision-Support in the Western US Great Plains. Agric. Syst. 2021, 191, 103139. [Google Scholar] [CrossRef]
- Zilverberg, C.J.; Williams, J.; Jones, C.; Harmoney, K.; Angerer, J.; Metz, L.J.; Fox, W. Process-Based Simulation of Prairie Growth. Ecol. Model. 2017, 351, 24–35. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine. Nutrient Requirements of Beef Cattle: Eighth Revised Edition; Animal Nutrition Series; 8th ed.; The National Academies Press: Washington, DC, USA, 2016; ISBN 978-0-309-31702-3. [Google Scholar]
- Tedeschi, L.O.; Fox, D.G.; Guiroy, P.J. A Decision Support System to Improve Individual Cattle Management. 1. A Mechanistic, Dynamic Model for Animal Growth. Agric. Syst. 2004, 79, 171–204. [Google Scholar] [CrossRef]
- Aherin, D.G.; Weaber, R.L.; Pendell, D.L.; Heier Stamm, J.L.; Larson, R.L. Stochastic, Individual Animal Systems Simulation Model of Beef Cow-Calf Production: Development and Validation. Transl. Anim. Sci. 2022, 7, txac155. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, P.A.; Larson, R.L. Evaluation of Strategies to Improve the Environmental and Economic Sustainability of Cow–Calf Production Systems. Animals 2022, 12, 385. [Google Scholar] [CrossRef] [PubMed]
- Owens, F.N.; Dubeski, P.; Hanson, C.F. Factors That Alter the Growth and Development of Ruminants. J. Anim. Sci. 1993, 71, 3138–3150. [Google Scholar] [CrossRef] [PubMed]
- Owens, F.N.; Gill, D.R.; Secrist, D.S.; Coleman, S.W. Review of Some Aspects of Growth and Development of Feedlot Cattle. J. Anim. Sci. 1995, 73, 3152–3172. [Google Scholar] [CrossRef] [PubMed]
- Coleman, S.W.; Evans, B.C.; Guenther, J.J. Body and Carcass Composition of Angus and Charolais Steers as Affected by Age and Nutrition. J. Anim. Sci. 1993, 71, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Sainz, R.D.; De la Torre, F.; Oltjen, J.W. Compensatory Growth and Carcass Quality in Growth-Restricted and Refed Beef Steers. J. Anim. Sci. 1995, 73, 2971–2979. [Google Scholar] [CrossRef] [PubMed]
- Hersom, M.J.; Horn, G.W.; Krehbiel, C.R.; Phillips, W.A. Effect of Live Weight Gain of Steers during Winter Grazing: I. Feedlot Performance, Carcass Characteristics, and Body Composition of Beef Steers. J. Anim. Sci. 2004, 82, 262–272. [Google Scholar] [CrossRef]
- McCurdy, M.P.; Horn, G.W.; Wagner, J.J.; Lancaster, P.A.; Krehbiel, C.R. Effects of Winter Growing Programs on Subsequent Feedlot Performance, Carcass Characteristics, Body Composition, and Energy Requirements of Beef Steers. J. Anim. Sci. 2010, 88, 1564–1576. [Google Scholar] [CrossRef]
- Sharman, E.D.; Lancaster, P.A.; McMurphy, C.P.; Garmyn, A.J.; Pye, B.J.; Mafi, G.G.; Goad, C.L.; Phillips, W.A.; Starkey, J.D.; Krehbiel, C.R.; et al. Effect of Rate of Body Weight Gain in Steers during the Stocker Phase. I. Growth, Partitioning of Fat among Depots, and Carcass Characteristics of Growing-Finishing Beef Cattle. J. Anim. Sci. 2013, 91, 4322–4335. [Google Scholar] [CrossRef]
- Lancaster, P.A. Assessment of Equations to Predict Body Weight and Chemical Composition in Growing/Finishing Cattle and Effects of Publication Year, Sex, and Breed Type on the Deviation from Observed Values. Animals 2022, 12, 3554. [Google Scholar] [CrossRef]
- Baker, R.D.; Du, Y.L.P.L.; Barker, J.M. Milk-Fed Calves: 1. The Effect of Milk Intake upon the Herbage Intake and Performance of Grazing Calves. J. Agric. Sci. 1976, 87, 187–196. [Google Scholar] [CrossRef]
- Holloway, J.W.; Butts, W.T.; Worley, T.L. Utilization of Forage and Milk Energy by Angus Calves Grazing Fescue or Fescue-Legume Pastures. J. Anim. Sci. 1982, 55, 1214–1223. [Google Scholar] [CrossRef]
- Tedeschi, L.O.; Fox, D.G. Predicting Milk and Forage Intake of Nursing Calves. J. Anim. Sci. 2009, 87, 3380–3391. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, P.A.; Tedeschi, L.O.; Buessing, Z.; Davis, M.E. Assessment of Milk Yield and Nursing Calf Feed Intake Equations in Predicting Calf Feed Intake and Weaning Weight among Breeds. J. Anim. Sci. 2021, 99, skaa406. [Google Scholar] [CrossRef] [PubMed]
- Wood, P.D.P. Algebraic Model of the Lactation Curve in Cattle. Nature 1967, 216, 164–165. [Google Scholar] [CrossRef]
- Tedeschi, L.O.; Fox, D.G.; Baker, M.J.; Long, K.L. A Model to Evaluate Beef Cow Efficiency. In Nutrient Digestion and Utilization in Farm Animals: Modelling Approaches; Kebreab, E., Dijkstra, J., Bannink, A., Gerrits, W.J.J., France, J., Eds.; CABI Publishing: Cambridge, MA, USA, 2006; pp. 84–98. [Google Scholar]
- Abdelsamei, A.H. Forage Intake, Growth and Final Composition of Suckling Calves Fed Five Levels of Milk Preweaning. Ph.D. Disstertation, Cornell University, Ithaca, NY, USA, 1989. [Google Scholar]
- Boggs, D.L. Effects of Milk and Forage Intake on Calf Performance. Master’s Thesis, Kansas State University, Manhattan, KS, USA, 1977. [Google Scholar]
- Tilley, J.M.A.; Terry, R.A. A Two-Stage Technique for the in Vitro Digestion of Forage Crops. Grass Forage Sci. 1963, 18, 104–111. [Google Scholar] [CrossRef]
- Blum, J.K.; Schalles, R.R.; Zoellner, K.O. Factors Influencing Net Income from a Steer through Feedlot; Kansas Agricultural Experiment Station Research Reports; Kansas State University: Manhattan, KS, USA, 1976; pp. 88–90. [Google Scholar]
- Dikeman, M.E.; May, M.L.; Lipsey, R.J.; Loveday, H.D.; Allen, D.M. Performance and Carcass Characteristics of Different Cattle Types; Kansas State University: Manhattan, KS, USA, 1975. [Google Scholar]
- Garrett, W.N.; Hinman, N. Re-Evaluation of the Relationship between Carcass Density and Body Composition of Beef Steers. J. Anim. Sci. 1969, 28, 1–5. [Google Scholar] [CrossRef]
- Ferrell, C.L.; Garrett, W.N.; Hinman, N.; Grichting, G. Energy Utilization by Pregnant and Non-Pregnant Heifers. J. Anim. Sci. 1976, 42, 937–950. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- McLeod, M.N.; Minson, D.J. The Use of the in Vitro Technique in the Determination of the Digestibility of Grass/Legume Mixtures. Grass Forage Sci. 1969, 24, 296. [Google Scholar] [CrossRef]
- Terry, R.A.; Mundell, D.C.; Osbourn, D.F. Comparison of Two in Vitro Procedures Using Rumen Liquor-Pepsin or Pepsin-Cellulase for Prediction of Forage Digestibility. Grass Forage Sci. 1978, 33, 13–18. [Google Scholar] [CrossRef]
- Goldman, A.; Genizi, A.; Yulzari, A.; Seligman, N.G. Improving the Reliability of the Two-Stage in Vitro Assay for Ruminant Feed Digestibility by Calibration against in Vivo Data from a Wide Range of Sources. Anim. Feed. Sci. Technol. 1987, 18, 233–245. [Google Scholar] [CrossRef]
- Genizi, A.; Goldman, A.; Yulzari, A.; Seligman, N.G. Evaluation of Methods for Calibrating in Vitro Digestibility Estimates of Ruminant Feeds. Anim. Feed. Sci. Technol. 1990, 29, 265–278. [Google Scholar] [CrossRef]
- Carro, M.D.; López, S.; González, J.S.; Ovejero, F.J. Comparison of Laboratory Methods for Predicting Digestibility of Hay in Sheep. Small Rumin. Res. 1994, 14, 9–17. [Google Scholar] [CrossRef]
- Khazaal, K.; Dentinho, M.T.; Ribeiro, J.M.; Ørskov, E.R. Prediction of Apparent Digestibility and Voluntary Intake of Hays Fed to Sheep: Comparison between Using Fibre Components, in Vitro Digestibility or Characteristics of Gas Production or Nylon Bag Degradation. Anim. Sci. 1995, 61, 527–538. [Google Scholar] [CrossRef]
- Geisert, B.G. Development of a Set of Forage Standard to Estimate in Vivo Digestibility of Forages and Prediction of Forage Quality of Diets Consumed by Cattle Grazing Nebraska Sandhills Range Pastures. Ph.D. Thesis, The University of Nebraska, Lincoln, NE, USA, 2007. [Google Scholar]
- McLean, J.A.; Tobin, G. Animal and Human Calorimetry; Cambridge University Press: New York, NY, USA, 1987; ISBN 978-0-521-30905-9. [Google Scholar]
- Smith, W.B.; Galyean, M.L.; Kallenbach, R.L.; Greenwood, P.L.; Scholljegerdes, E.J. Understanding Intake on Pastures: How, Why, and a Way Forward. J. Anim. Sci. 2021, 99, skab062. [Google Scholar] [CrossRef]
- Burns, J.C.; Pond, K.R.; Fisher, D.S. Measurement of Forage Intake. In Forage Quality, Evaluation, and Utilization; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1994; pp. 494–532. ISBN 978-0-89118-579-6. [Google Scholar]
- Cottle, D.J. The Trials and Tribulations of Estimating the Pasture Intake of Grazing Animals. Anim. Prod. Sci. 2013, 53, 1209–1220. [Google Scholar] [CrossRef]
- Akdağ, A.; Ocak, N. Herbage Intake Determination Methods of Grazing Animals. Anim. Sci. 2019, 52, 1. [Google Scholar]
- Coleman, S.W. Challenges to Assessing Forage Intake by Grazing Ruminants. In Proceedings of the 8th World Congress on Genetics Applied to Livestock Production, Belo Horizonte, Brazil, 13–18 August 2006; pp. 6–14. [Google Scholar]
- Cordova, F.J.; Wallace, J.D.; Pieper, R.D. Forage Intake by Grazing Livestock: A Review. J. Range Manag. 1978, 31, 430–438. [Google Scholar] [CrossRef]
- Allison, C.D. Factors Affecting Forage Intake by Range Ruminants: A Review. J. Range Manag. 1985, 38, 305–311. [Google Scholar] [CrossRef]
- Ellis, W.C. Determinants of Grazed Forage Intake and Digestibility. J. Dairy Sci. 1978, 61, 1828–1840. [Google Scholar] [CrossRef]
- Cuchillo-Hilario, M.; Wrage-Mönnig, N.; Isselstein, J. Forage Selectivity by Cattle and Sheep Co-Grazing Swards Differing in Plant Species Diversity. Grass Forage Sci. 2018, 73, 320–329. [Google Scholar] [CrossRef]
- Pauler, C.M.; Isselstein, J.; Suter, M.; Berard, J.; Braunbeck, T.; Schneider, M.K. Choosy Grazers: Influence of Plant Traits on Forage Selection by Three Cattle Breeds. Funct. Ecol. 2020, 34, 980–992. [Google Scholar] [CrossRef]
- Forbes, T.D.A. Effects of Sward Structure on Forage Intake. Implications for Increasing Performance of Cattle on Wheat Pasture. In Proceedings of the National Wheat Pasture Symposium; Oklahoma State University: Stillwater, OK, USA, 1984; pp. 207–222. [Google Scholar]
- Da Trindade, J.K.; Neves, F.P.; Pinto, C.E.; Bremm, C.; Mezzalira, J.C.; Nadin, L.B.; Genro, T.C.M.; Gonda, H.L.; Carvalho, P.C.F. Daily Forage Intake by Cattle on Natural Grassland: Response to Forage Allowance and Sward Structure. Rangel. Ecol. Manag. 2016, 69, 59–67. [Google Scholar] [CrossRef]
- Griffiths, W.M.; Hodgson, J.; Arnold, G.C. The Influence of Sward Canopy Structure on Foraging Decisions by Grazing Cattle. I. Patch Selection. Grass Forage Sci. 2003, 58, 112–124. [Google Scholar] [CrossRef]
- Vicini, J.L.; Prigge, E.C.; Bryan, W.B.; Varga, G.A. Influence of Forage Species and Creep Grazing on a Cow-Calf System. I. Intake and Digestibility of Forages. J. Anim. Sci. 1982, 55, 752–758. [Google Scholar] [CrossRef]
- Cherney, D.J.R.; Mertens, D.R.; Moore, J.E. Intake and Digestibility by Wethers as Influenced by Forage Morphology at Three Levels of Forage Offering1,2. J. Anim. Sci. 1990, 68, 4387–4399. [Google Scholar] [CrossRef]
- Baker, R.D.; Barker, J.M. Milk-Fed Calves: 4. The Effect of Herbage Allowance and Milk Intake upon Herbage Intake and Performance of Grazing Calves. J. Agric. Sci. 1978, 90, 31–38. [Google Scholar] [CrossRef]
- Peischel, H.A. Factors Affecting Milk and Grass Consumption of Calves Grazing Native Range. Ph.D. Thesis, Kansas State University, Manhattan, KS, USA, 1980. [Google Scholar]
- Boggs, D.L.; Smith, E.F.; Schalles, R.R.; Brent, B.E.; Corah, L.R.; Pruitt, R.J. Effects of Milk and Forage Intake on Calf Performance. J. Anim. Sci. 1980, 51, 550–553. [Google Scholar] [CrossRef]
- Bryan, K.S. Nutritional Aspects of Suckling Beef Calves Grazing Native Rangeland in Southwestern Montana. Master’s Thesis, Montana State University, Bozeman, MT, USA, 1989. [Google Scholar]
- Broesder, J.T.; Judkins, M.B.; Krysl, L.J.; Gunter, S.A.; Barton, R.K. Thirty or Sixty Percent Milk Replacer Reduction for Calves: Effects on Alfalfa Hay Intake and Digestibility, Digestive Kinetics and Ruminal Fermentation. J. Anim. Sci. 1990, 68, 2974–2985. [Google Scholar] [CrossRef]
- Ansotegui, R.P.; Havstad, K.M.; Wallace, J.D.; Hallford, D.M. Effects of Milk Intake on Forage Intake and Performance of Suckling Range Calves. J. Anim. Sci. 1991, 69, 899–904. [Google Scholar] [CrossRef]
- Sowell, B.F.; Wallace, J.D.; Branine, M.E.; Hubbert, M.E.; Fredrickson, E.L.; Bowman, J.G.P. Effects of Restricted Suckling on Forage Intake of Range Calves. J. Range Manag. 1996, 49, 290–293. [Google Scholar] [CrossRef]
- Abdelsamei, A.H.; Fox, D.G.; Tedeschi, L.O.; Thonney, M.L.; Ketchen, D.J.; Stouffer, J.R. The Effect of Milk Intake on Forage Intake and Growth of Nursing Calves. J. Anim. Sci. 2005, 83, 940–947. [Google Scholar] [CrossRef]
- Walker, J. Determining Forage Intake of Nursing Calves Grazing Native Sandhills Rangeland. Master’s Thesis, University of Nebraska, Lincoln, NE, USA, 2014. [Google Scholar]
- Lusby, K.S.; Stephens, D.F.; Totusek, R. Effects of Milk Intake by Nursing Calves on Forage Intake on Range and Creep Intake and Digestibility in Drylot. J. Anim. Sci. 1976, 43, 1066–1071. [Google Scholar] [CrossRef]
- Webb, L.E.; Engel, B.; Berends, H.; van Reenen, C.G.; Gerrits, W.J.J.; de Boer, I.J.M.; Bokkers, E.A.M. What Do Calves Choose to Eat and How Do Preferences Affect Behaviour? Appl. Anim. Behav. Sci. 2014, 161, 7–19. [Google Scholar] [CrossRef]
- Minick, J.A.; Buchanan, D.S.; Rupert, S.D. Milk Production of Crossbred Daughters of High- and Low-Milk EPD Angus and Hereford Bulls. J. Anim. Sci. 2001, 79, 1386–1393. [Google Scholar] [CrossRef] [PubMed]
- Visker, M.H.P.W.; Wang, Y.; van Pelt, M.L.; Bovenhuis, H. Milk Production of the Mother Is Associated with the Birth Weight Rather than the Sex of the Calf; Wageningen Academic Publishers: Wageningen, The Netherlands, 2015; p. 522. [Google Scholar]
- Robison, O.W.; Yusuff, M.K.M.; Dillard, E.U. Milk Production in Hereford Cows I. Means and Correlations. J. Anim. Sci. 1978, 47, 131–136. [Google Scholar] [CrossRef]
- Jeffery, H.B.; Berg, R.T.; Hardin, R.T. Factors Influencing Milk Yield of Beef Cattle. Can. J. Anim. Sci. 1971, 51, 551–560. [Google Scholar] [CrossRef]
- Lalman, D.L.; Williams, J.E.; Hess, B.W.; Thomas, M.G.; Keisler, D.H. Effect of Dietary Energy on Milk Production and Metabolic Hormones in Thin, Primiparous Beef Heifers. J. Anim. Sci. 2000, 78, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Spencer, C.M. Relationship of Maternal Dietary Energy Intake to Milk Production, Body Composition, and Efficiency of Calf Growth. Master’s Thesis, Oklahoma State University, Stillwater, OK, USA, 2017. [Google Scholar]
- Jenkins, T.G.; Ferrell, C.L. Lactation Characteristics of Nine Breeds of Cattle Fed Various Quantities of Dietary Energy. J. Anim. Sci. 1992, 70, 1652–1660. [Google Scholar] [CrossRef] [PubMed]
- Restle, J.; Pacheco, P.S.; Moletta, J.L.; Brondani, I.L.; Cerdótes, L. Genetic Group and Postpartum Nutritional Level on the Milk Yield and Composition of Beef Cows. R. Bras. Zootec. 2003, 32, 585–597. [Google Scholar] [CrossRef]
- Friggens, N.C.; Ridder, C.; Løvendahl, P. On the Use of Milk Composition Measures to Predict the Energy Balance of Dairy Cows. J. Anim. Sci. 2007, 90, 5453–5467. [Google Scholar] [CrossRef]
- Auldist, M.J.; Walsh, B.J.; Thomson, N.A. Seasonal and Lactational Influences on Bovine Milk Composition in New Zealand. J. Dairy Res. 1998, 65, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Linn, J.G. Factors Affecting the Composition of Milk from Dairy Cows. In Designing Foods: Animal Product Options in the Marketplace; National Academies Press (US): Washington, DC, USA, 1988. [Google Scholar]
- Mondragon, I.; Wilton, J.W.; Allen, O.B.; Song, H. Stage of Lactation Effects, Repeatabilities and Influences on Weaning Weights of Yield and Composition of Milk in Beef Cattle. Can. J. Anim. Sci. 1983, 63, 751–761. [Google Scholar] [CrossRef]
- Gleddie, V.M.; Berg, R.T. Milk Production in Range Beef Cows and Its Relationship to Calf Gains. Can. J. Anim. Sci. 1968, 48, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Lawson, J.E. Milk Production and Weight of Cows of the Hereford and Highland Breeds and Their Reciprocal Crosses. Can. J. Anim. Sci. 1981, 61, 861–869. [Google Scholar] [CrossRef]
- Church, D.C. The Ruminant Animal: Digestive Physiology and Nutrition; Waveland Press: Long Grove, IL, USA, 1993; ISBN 978-1-4786-4855-0. [Google Scholar]
- National Research Council. Nutrient Requirements of Beef Cattle, 6th ed.; National Academies Press: Washington, DC, USA, 1984; ISBN 978-0-309-03447-0. [Google Scholar]
- National Research Council. Nutrient Requirements of Beef Cattle, 7th ed.; National Academy Press: Washington, DC, USA, 2000; ISBN 978-0-309-38813-9. [Google Scholar]
- Hildebrand, C.; Lancaster, P.A. Evaluating Relationships Among Empty Body, Protein, and Fat Gain and Retained Energy for Pre- and Postweaning Calves. J. Anim. Sci. 2022, 100, 190–191. [Google Scholar] [CrossRef]
- Hill, T.M.; Bateman, H.G.; Aldrich, J.M.; Schlotterbeck, R.L. Effect of Milk Replacer Program on Digestion of Nutrients in Dairy Calves. J. Anim. Sci. 2010, 93, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Hill, T.M.; Quigley, J.D.; Bateman, H.G.; Suarez-Mena, F.X.; Dennis, T.S.; Schlotterbeck, R.L. Effect of Milk Replacer Program on Calf Performance and Digestion of Nutrients in Dairy Calves to 4 Months of Age. J. Dairy Sci. 2016, 99, 8103–8110. [Google Scholar] [CrossRef]
- Dennis, T.S.; Suarez-Mena, F.X.; Hu, W.; Hill, T.M.; Quigley, J.D.; Schlotterbeck, R.L. Effects of Milk Replacer Feeding Rate and Long-Term Antibiotic Inclusion in Milk Replacer on Performance and Nutrient Digestibility of Holstein Dairy Calves up to 4 Months of Age. J. Dairy Sci. 2019, 102, 2094–2102. [Google Scholar] [CrossRef]
- Dennis, T.S.; Suarez-Mena, F.X.; Hill, T.M.; Quigley, J.D.; Schlotterbeck, R.L.; Hulbert, L. Effect of Milk Replacer Feeding Rate, Age at Weaning, and Method of Reducing Milk Replacer to Weaning on Digestion, Performance, Rumination, and Activity in Dairy Calves to 4 Months of Age. J. Dairy Sci. 2018, 101, 268–278. [Google Scholar] [CrossRef]
- Terré, M.; Devant, M.; Bach, A. Effect of Level of Milk Replacer Fed to Holstein Calves on Performance during the Preweaning Period and Starter Digestibility at Weaning. Livest. Sci. 2007, 110, 82–88. [Google Scholar] [CrossRef]
- Cabezas-Garcia, E.H.; Krizsan, S.J.; Shingfield, K.J.; Huhtanen, P. Between-Cow Variation in Digestion and Rumen Fermentation Variables Associated with Methane Production. J. Dairy Sci. 2017, 100, 4409–4424. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.C.; Herd, R.M.; Donoghue, K.A.; Bird-Gardiner, T.; Arthur, P.F.; Hegarty, R.S. Associations of Digestibility with Phenotypic and Genetic Variation in Methane Production in Angus Cattle. Anim. Prod. Sci. 2022, 62, 1749–1756. [Google Scholar] [CrossRef]
- Berry, D.P.; Horan, B.; O’Donovan, M.; Buckley, F.; Kennedy, E.; McEvoy, M.; Dillon, P. Genetics of Grass Dry Matter Intake, Energy Balance, and Digestibility in Grazing Irish Dairy Cows. J. Dairy Sci. 2007, 90, 4835–4845. [Google Scholar] [CrossRef]
- Mehtiö, T.; Mäntysaari, P.; Kokkonen, T.; Kajava, S.; Prestløkken, E.; Kidane, A.; Wallén, S.; Nyholm, L.; Negussie, E.; Mäntysaari, E.A.; et al. Genetic Parameters for Cow-Specific Digestibility Predicted by near Infrared Reflectance Spectroscopy. Livest. Sci. 2019, 226, 1–6. [Google Scholar] [CrossRef]
- Preston, T.R.; Archibald, J.D.H.; Tinkler, W. The Digestibility of Grass by Young Calves. J. Agric. Sci. 1957, 48, 259–265. [Google Scholar] [CrossRef]
- McCollough, M.E.; Sisk, L.R. Effect of Age of Young Dairy Calves on Dry-Matter Digestibility of Alfalfa Pellets. Nature 1958, 181, 57. [Google Scholar] [CrossRef]
- Godfrey, N.W. The Functional Development of the Calf II. Development of Rumen Function in the Calf. J. Agric. Sci. 1961, 57, 177–183. [Google Scholar] [CrossRef]
- Lamothe, M.; Klopfenstein, T.J.; Adams, D.C.; Musgrave, J.A.; Erickson, G.E. Microbial Protein Synthesis and Efficiency in Nursing Calves; Nebraska Beef Cattle Reports; University of Nebraska: Lincoln, NE, USA, 2003; pp. 13–15. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Mean | SD | Minimum | Maximum |
|---|---|---|---|---|
| Birth date, Julian d | 208.70 | 24.35 | 170 | 257 |
| Birth weight, kg | 44.9 | 9.0 | 36.0 | 57.0 |
| 30-d BW, kg | 61.6 | 12.3 | 35.0 | 92.0 |
| 30-d milk intake, kg/d | 6.65 | 2.75 | 2.50 | 12.08 |
| 30-d forage intake, kg DM/d | 0.08 | 0.09 | 0.00 | 0.41 |
| 60-d BW, kg | 85.0 | 17.0 | 49.0 | 117.0 |
| 60-d milk intake, kg/d | 7.25 | 3.15 | 2.50 | 12.69 |
| 60-d forage intake, kg DM/d | 0.29 | 0.30 | 0.00 | 1.33 |
| 90-d BW, kg | 115.8 | 23.1 | 71.0 | 165.0 |
| 90-d milk intake, kg/d | 7.27 | 3.51 | 2.37 | 12.83 |
| 90-d forage intake, kg DM/d | 0.88 | 0.57 | 0.16 | 2.14 |
| 115-d BW, kg | 142.6 | 26.7 | 84.0 | 184.0 |
| 115-d milk intake, kg/d | 6.65 | 3.04 | 2.14 | 11.83 |
| 115-d forage intake, kg DM/d | 1.55 | 0.64 | 0.39 | 2.58 |
| 145-d BW, kg | 168.1 | 29.8 | 104.0 | 220.0 |
| 145-d milk intake, kg/d | 5.90 | 2.99 | 1.84 | 10.46 |
| 145-d forage intake, kg DM/d | 2.26 | 0.66 | 1.08 | 3.33 |
| 165-d BW, kg | 187.2 | 33.9 | 117.0 | 239.0 |
| 165-d milk intake, kg/d | 5.27 | 2.67 | 1.60 | 9.34 |
| 165-d forage intake, kg DM/d | 2.98 | 0.71 | 1.22 | 4.79 |
| 195-d BW, kg | 212.2 | 38.0 | 127.0 | 269.0 |
| 195-d milk intake, kg/d | 4.44 | 2.27 | 1.34 | 8.45 |
| 195-d forage intake, kg DM/d | 3.83 | 0.52 | 2.55 | 4.84 |
| Item | Mean | SD | Minimum | Maximum |
|---|---|---|---|---|
| Birth date, Julian | 86.75 | 12.84 | 60 | 122 |
| Birth weight, kg | 34.8 | 3.4 | 23.1 | 41.8 |
| April BW, kg | 61.0 | 9.7 | 40.0 | 80.6 |
| April milk yield, kg/d | 6.43 | 1.49 | 3.40 | 9.99 |
| May BW, kg | 78.3 | 11.3 | 55.4 | 99.0 |
| May milk yield, kg/d | 5.90 | 1.49 | 3.13 | 10.44 |
| May forage intake, kg DM/d | 0.47 | 0.21 | 0.20 | 10.16 |
| June BW, kg | 96.8 | 13.4 | 68.5 | 119.8 |
| June milk yield, kg/d | 4.95 | 1.44 | 1.32 | 8.40 |
| June forage intake, kg DM/d | 1.45 | 0.69 | 0.47 | 3.14 |
| July BW, kg | 120.1 | 16.6 | 83.5 | 148.4 |
| July milk yield, kg/d | 4.82 | 1.45 | 2.27 | 8.40 |
| July forage intake, kg DM/d | 1.85 | 0.72 | 0.92 | 5.05 |
| Aug. BW, kg | 145.9 | 18.9 | 103.5 | 177.9 |
| Aug. milk yield, kg/d | 3.38 | 0.95 | 1.35 | 6.81 |
| Aug. forage intake, kg DM/d | 2.62 | 0.64 | 1.45 | 4.36 |
| Sept. BW, kg | 165.3 | 21.1 | 116.1 | 202.5 |
| Sept. milk yield, kg/d | 3.61 | 1.12 | 1.13 | 5.90 |
| Sept. forage intake, kg DM/d | 3.57 | 0.84 | 2.36 | 6.14 |
| Item 1 | NASEM | WOOD | Both |
|---|---|---|---|
| CCC | 0.951 | 0.954 | 0.969 |
| Cb | 0.991 | 0.996 | 1.000 |
| MB (SD), kg/d | 0.393 (0.919) | −0.253 (0.927) | −0.045 (0.771) |
| MB, % | 6.44 | −4.14 | −0.74 |
| Intercept ± SE | 0.9094 ± 0.0753 | 0.2690 ± 0.0842 | 0.1622 ± 0.0728 |
| Slope ± SE | 0.9096 ± 0.0115 | 0.9179 ± 0.0118 | 0.9662 ± 0.0106 |
| Pr > F | <0.0001 | <0.0001 | 0.0025 |
| Item | Eq91 1 | Eq67 | Eq25 | Eq17 | Eq21 |
|---|---|---|---|---|---|
| Forage Intake | |||||
| CCC 2 | 0.878 | 0.920 | 0.729 | 0.874 | 0.234 |
| Cb | 0.957 | 0.999 | 0.797 | 0.921 | 0.257 |
| MB (SD), kg/d | −0.259 (0.722) | −0.023 (0.546) | −0.954 (0.984) | −0.527 (0.580) | −5.055 (3.698) |
| MB, % | −17.49 | −1.58 | −64.4 | −35.6 | −341.16 |
| Intercept ± SE | 0.2266 ± 0.0326 | 0.1322 ± 0.0332 | 0.0135 ± 0.0360 | −0.0563 ± 0.0283 | −0.1712 ± 0.0397 |
| Slope ± SE | 0.7188 ± 0.0134 | 0.8956 ± 0.0163 | 0.6005 ± 0.0113 | 0.7639 ± 0.0109 | 0.2513 ± 0.0049 |
| Pr > F | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Body Weight | |||||
| CCC | 0.876 | 0.929 | 0.819 | 0.864 | 0.402 |
| Cb | 0.908 | 0.951 | 0.833 | 0.878 | 0.423 |
| MB (SD), kg | −26.439 (16.932) | −18.005 (15.271) | −36.952 (23.855) | −30.847 (17.077) | −122.473 (84.252) |
| MB, % | −21.54 | −14.67 | −30.1 | −25.13 | −99.78 |
| Intercept ± SE | −7.9868 ± 1.6539 | 1.8476 ± 1.2441 | 7.0021 ± 1.0362 | −0.5910 ± 1.0313 | 25.1856 ± 1.5894 |
| Slope ± SE | 0.8763 ± 0.0102 | 0.8589 ± 0.0080 | 0.7248 ± 0.0058 | 0.8030 ± 0.0061 | 0.3978 ± 0.0057 |
| Pr > F | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Item 3 | Eq91 2 | Eq67 | Eq25 | Eq17 | Eq21 |
|---|---|---|---|---|---|
| CCC | 0.935 | 0.936 | 0.958 | 0.964 | 0.640 |
| Cb | 0.998 | 0.997 | 0.99 | 0.995 | 0.690 |
| MB (SD), kg/d | −0.008 (0.503) | 0.097 (0.471) | −0.176 (0.366) | −0.131 (0.342) | −1.366 (1.171) |
| MB, % | −0.55 | 6.57 | −11.9 | −8.83 | −92.26 |
| Intercept ± SE | 0.1745 ± 0.0288 | 0.1655 ± 0.0287 | −0.0298 ± 0.0224 | −0.0328 ± 0.0216 | −0.0674 ± 0.0340 |
| Slope ± SE | 0.8763 ± 0.0140 | 0.9503 ± 0.0150 | 0.9109 ± 0.0103 | 0.9387 ± 0.0102 | 0.5416 ± 0.0093 |
| Pr > F | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Item 2 | Original EBG Equation | New EBG Equation |
|---|---|---|
| CCC | 0.926 | 0.992 |
| Cb | 0.938 | 0.999 |
| MB (SD), kg/d | −20.966 (11.365) | −0.135 (7.368) |
| MB, % | −17.08 | −0.11 |
| Intercept ± SE | −6.4356 ± 1.0076 | −1.7250 ± 0.7569 |
| Slope ± SE | 0.8989 ± 0.0064 | 1.0129 ± 0.0056 |
| Pr > F | <0.0001 | 0.0643 |
| Item | Eq91 1 | Eq67 | Eq25 | Eq17 | Eq21 |
|---|---|---|---|---|---|
| Forage Intake | |||||
| CCC 2 | 0.914 | 0.766 | 0.881 | 0.901 | 0.412 |
| Cb | 0.984 | 0.825 | 0.967 | 0.973 | 0.451 |
| MB (SD), kg/d | 0.229 (0.500) | 0.600 (0.594) | 0.268 (0.561) | 0.252 (0.513) | −2.561 (2.527) |
| MB, % | 15.47 | 40.53 | 18.11 | 17.02 | −172.88 |
| Intercept ± SE | 0.2588 ± 0.0298 | 0.3072 ± 0.0294 | 0.2037 ± 0.0343 | 0.1928 ± 0.0312 | 0.1269 ± 0.0347 |
| Slope ± SE | 0.9761 ± 0.0167 | 1.3382 ± 0.0231 | 1.0539 ± 0.0206 | 1.0489 ± 0.0184 | 0.3328 ± 0.0064 |
| Pr > F | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Body Weight | |||||
| CCC | 0.978 | 0.940 | 0.968 | 0.969 | 0.701 |
| Cb | 0.991 | 0.973 | 0.995 | 0.995 | 0.739 |
| MB (SD), kg/d | 3.472 (10.701) | 10.934 (15.392) | 4.749 (13.263) | 4.807 (13.190) | −47.540 (48.220) |
| MB, % | 2.83 | 8.91 | 3.87 | 3.92 | −38.73 |
| Intercept ± SE | −9.6804 ± 0.9940 | 0.7530 ± 1.5226 | 2.4086 ± 1.3481 | 2.7352 ± 1.3383 | 29.2959 ± 1.5428 |
| Slope ± SE | 1.1103 ± 0.0077 | 1.0911 ± 0.0124 | 1.0198 ± 0.0104 | 1.0176 ± 0.0103 | 0.5488 ± 0.0078 |
| Pr > F | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Item 1 | NASEM | WOOD |
|---|---|---|
| CCC | 0.796 | 0.820 |
| Cb | 0.993 | 0.991 |
| MB (SD), kg/d | 0.206 (1.078) | 0.063 (0.969) |
| MB, % | 4.29 | 1.31 |
| Intercept ± SE | 1.0875 ± 0.1719 | 0.3321 ± 0.1850 |
| Slope ± SE | 0.8087 ± 0.0350 | 0.9433 ± 0.0371 |
| Pr > F | <0.0001 | 0.1673 |
| Item | Eq91 1 | Eq67 | Eq25 | Eq17 | Eq21 |
|---|---|---|---|---|---|
| Forage Intake | |||||
| CCC 2 | 0.613 | 0.476 | 0.167 | 0.491 | 0.348 |
| Cb | 0.745 | 0.752 | 0.278 | 0.706 | 0.608 |
| MB (SD), kg/d | 0.389 (0.802) | 0.433 (0.963) | 1.160 (1.064) | 0.565 (0.910) | 0.409 (1.037) |
| MB, % | 19.36 | 21.51 | 57.67 | 28.07 | 20.33 |
| Intercept ± SE | −0.6318 ± 0.1251 | 0.1182 ± 0.1605 | 0.1346 ± 0.1714 | 0.0939 ± 0.1391 | −0.3867 ± 0.2296 |
| Slope ± SE | 1.6294 ± 0.0720 | 1.1992 ± 0.0940 | 2.2045 ± 0.1873 | 1.3254 ± 0.0878 | 1.4965 ± 0.1374 |
| Pr > F | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Body Weight | |||||
| CCC | 0.762 | 0.76 | 0.574 | 0.766 | 0.766 |
| Cb | 0.899 | 0.898 | 0.757 | 0.874 | 0.909 |
| MB (SD), kg/d | 10.224 (21.910) | 10.946 (21.887) | 17.594 (26.833) | 11.911 (23.166) | 7.114 (22.379) |
| MB, % | 9.18 | 9.83 | 15.79 | 10.69 | 6.39 |
| Intercept ± SE | −11.3177 ± 4.5786 | −8.3620 ± 4.5074 | −11.4078 ± 6.2286 | −10.2020 ± 4.9313 | −18.4033 ± 4.9166 |
| Slope ± SE | 1.2129 ± 0.0436 | 1.1922 ± 0.0432 | 1.3092 ± 0.0645 | 1.2223 ± 0.0479 | 1.2447 ± 0.0457 |
| Pr > F | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Item | Eq91 2 | Eq67 | Eq25 | Eq17 | Eq21 |
|---|---|---|---|---|---|
| CCC 3 | 0.834 | 0.725 | 0.630 | 0.738 | 0.528 |
| Cb | 0.983 | 0.953 | 0.830 | 0.959 | 0.727 |
| MB (SD), kg/d | 0.016 (0.653) | 0.112 (0.800) | 0.555 (0.806) | 0.196 (0.787) | 0.418 (0.894) |
| MB, % | 0.81 | 5.58 | 27.56 | 9.73 | 20.76 |
| Intercept ± SE | −0.0222 ± 0.0915 | 0.0721 ± 0.1176 | 0.4491 ± 0.1001 | 0.2499 ± 0.1061 | −0.3290 ± 0.1520 |
| Slope ± SE | 1.0193 ± 0.0408 | 1.0211 ± 0.0557 | 1.0723 ± 0.0590 | 0.9701 ± 0.0515 | 1.4684 ± 0.0891 |
| Pr > F | 0.8283 | 0.0854 | <0.0001 | 0.0005 | <0.0001 |
| Item | Original 2 | New Forage Digestibility | New Milk Energy | New EBG Equation | New Combination |
|---|---|---|---|---|---|
| CCC 3 | 0.821 | 0.883 | 0.863 | 0.901 | 0.931 |
| Cb | 0.913 | 0.957 | 0.946 | 0.981 | 0.985 |
| MB (SD), kg/d | 10.796 (18.441) | 7.260 (16.097) | 6.998 (17.324) | 1.757 (16.143) | −5.965 (13.045) |
| MB, % | 9.69 | 6.52 | 6.28 | 1.58 | −5.35 |
| Intercept ± SE | −11.0374 ± 3.5433 | −7.8853 ± 3.0152 | −13.3124 ± 3.3515 | −10.5959 ± 3.1486 | −8.3616 ± 2.5005 |
| Slope ± SE | 1.2170 ± 0.0338 | 1.1454 ± 0.0277 | 1.1945 ± 0.0308 | 1.1127 ± 0.0275 | 1.0204 ± 0.0203 |
| Pr > F | <0.0001 | < 0.0001 | <0.0001 | < 0.0001 | <0.0001 |
| Item | Eq91 1 | Eq67 | Eq25 | Eq17 | Eq21 |
|---|---|---|---|---|---|
| Forage Intake | |||||
| CCC 2 | 0.822 | 0.670 | 0.434 | 0.686 | 0.380 |
| Cb | 0.956 | 0.951 | 0.546 | 0.842 | 0.628 |
| MB (SD), kg/d | −0.149 (0.674) | −0.107 (0.879) | 0.783 (0.925) | 0.281 (0.809) | 0.233 (1.078) |
| MB, % | −7.01 | −5.08 | 39.87 | 13.24 | 10.96 |
| Intercept ± SE | −0.4464 ± 0.1063 | −0.1010 ± 0.1427 | −0.4021 ± 0.1337 | −0.4482 ± 0.1263 | −1.0047 ± 0.2712 |
| Slope ± SE | 1.1309 ± 0.0429 | 0.9969 ± 0.0588 | 1.8840 ± 0.0924 | 1.3958 ± 0.0634 | 1.6545 ± 0.1391 |
| Pr > F | <0.0001 | 0.1603 | <0.0001 | <0.0001 | <0.0001 |
| Body Weight | |||||
| CCC | 0.935 | 0.938 | 0.896 | 0.926 | 0.886 |
| Cb | 0.978 | 0.985 | 0.972 | 0.98 | 0.956 |
| MB (SD), kg/d | −8.173 (11.663) | −6.918 (12.128) | −0.104 (16.267) | −5.956 (13.310) | −10.486 (14.937) |
| MB, % | −7.34 | −6.21 | −0.09 | −5.35 | −9.41 |
| Intercept ± SE | −6.6719 ± 2.1851 | −3.2861 ± 2.2109 | −19.3587 ± 3.2651 | −13.6285 ± 2.6198 | −16.0307 ± 3.0696 |
| Slope ± SE | 0.9875 ± 0.0174 | 0.9693 ± 0.0178 | 1.1727 ± 0.0282 | 1.0654 ± 0.0214 | 1.0455 ± 0.0242 |
| Pr > F | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Birth Period 1 | p-Value 2 | |||||||
|---|---|---|---|---|---|---|---|---|
| Equation 3 | Covariate Level | 1 | 2 | 3 | SEM | BP | Cov | BP × Cov |
| Eq91 | 0.75 kg/d | −0.62 a | −0.46 a | −0.06 b | 0.09 | 0.01 | 0.01 | 0.01 |
| 1.50 kg/d | −0.37 a | −0.22 a | 0.02 b | 0.06 | ||||
| 2.25 kg/d | −0.11 | 0.02 | 0.10 | 0.06 | ||||
| 3.00 kg/d | 0.15 | 0.26 | 0.19 | 0.07 | ||||
| 3.75 kg/d | 0.40 | 0.50 | 0.27 | 0.10 | ||||
| Eq67 | 0.75 kg/d | −1.08 a | −0.75 a | −0.22 b | 0.10 | 0.01 | 0.01 | 0.01 |
| 1.50 kg/d | −0.61 a | −0.39 a | 0.02 b | 0.07 | ||||
| 2.25 kg/d | −0.13 a | −0.03 a | 0.27 b | 0.07 | ||||
| 3.00 kg/d | 0.34 | 0.34 | 0.52 | 0.08 | ||||
| 3.75 kg/d | 0.82 | 0.70 | 0.76 | 0.11 | ||||
| Eq25 | 0.75 kg/d | −0.72 a | −0.55 ab | −0.32 a | 0.10 | 0.47 | 0.01 | 0.05 |
| 1.50 kg/d | −0.37 | −0.26 | −0.13 | 0.07 | ||||
| 2.25 kg/d | −0.02 | 0.03 | 0.06 | 0.07 | ||||
| 3.00 kg/d | 0.33 | 0.32 | 0.25 | 0.08 | ||||
| 3.75 kg/d | 0.66 | 0.61 | 0.44 | 0.11 | ||||
| Eq17 | 0.75 kg/d | −0.64 a | −0.55 a | −0.24 b | 0.09 | 0.17 | 0.01 | 0.08 |
| 1.50 kg/d | −0.34 | −0.27 | −0.07 | 0.07 | ||||
| 2.25 kg/d | −0.03 | 0.01 | 0.09 | 0.07 | ||||
| 3.00 kg/d | 0.27 | 0.29 | 0.26 | 0.08 | ||||
| 3.75 kg/d | 0.58 | 0.57 | 0.43 | 0.11 | ||||
| Eq21 | 0.75 kg/d | −1.11 a | −1.02 a | −0.54 b | 0.09 | 0.01 | 0.01 | 0.01 |
| 1.50 kg/d | −0.50 a | −0.54 a | −0.20 b | 0.07 | ||||
| 2.25 kg/d | 0.10 | −0.05 | 0.14 | 0.07 | ||||
| 3.00 kg/d | 0.71 a | 0.43 b | 0.47 ab | 0.08 | ||||
| 3.75 kg/d | 1.31 a | 0.92 b | 0.81 b | 0.11 | ||||
| Birth Period 1 | p-Value 2 | |||||||
|---|---|---|---|---|---|---|---|---|
| Equation 3 | Covariate Level | 1 | 2 | 3 | SEM | BP | Cov | BP × Cov |
| Eq91 | 60 kg | −11.6 a | −6.1 a | 2.2 b | 1.7 | 0.01 | 0.01 | 0.01 |
| 90 kg | −5.8 a | −3.5 a | 2.2 b | 1.2 | ||||
| 120 kg | −0.1 | −0.8 | 2.3 | 1.1 | ||||
| 150 kg | 5.6 | 1.9 | 2.3 | 1.5 | ||||
| Eq67 | 60 kg | −11.7 a | −6.4 a | 1.6 b | 1.7 | 0.01 | 0.01 | 0.01 |
| 90 kg | −5.8 a | −3.6 a | 2.1 b | 1.2 | ||||
| 120 kg | 0.1 | −0.8 | 2.5 | 1.1 | ||||
| 150 kg | 5.9 | 1.9 | 3.0 | 1.6 | ||||
| Eq25 | 60 kg | −15.3 a | −10.5 a | 0.8 b | 2.1 | 0.01 | 0.01 | 0.01 |
| 90 kg | −6.3 a | −6.1 a | 1.4 b | 1.5 | ||||
| 120 kg | 2.7 | −1.7 | 2.1 | 1.5 | ||||
| 150 kg | 11.7 a | 2.7 b | 2.7 b | 1.9 | ||||
| Eq17 | 60 kg | −12.7 a | −7.8 a | 1.7 b | 1.9 | 0.01 | 0.01 | 0.01 |
| 90 kg | −5.6 a | −4.6 a | 1.8 b | 1.3 | ||||
| 120 kg | 1.5 | −1.4 | 1.9 | 1.2 | ||||
| 150 kg | 8.6 a | 1.8 b | 2.0 b | 1.7 | ||||
| Eq21 | 60 kg | −18.5 a | −8.2 b | 1.7 c | 2.1 | 0.01 | 0.01 | 0.01 |
| 90 kg | −8.9 a | −4.4 a | 1.9 b | 1.4 | ||||
| 120 kg | 0.7 | −0.7 | 2.1 | 1.4 | ||||
| 150 kg | 10.3 a | 3.0 b | 2.3 b | 1.8 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldin, G.C.; Hildebrand, C.; Larson, R.L.; Lancaster, P.A. Evaluation and Development of a Nutrition Model to Predict Intake and Growth of Suckling Calves. Ruminants 2024, 4, 47-78. https://0-doi-org.brum.beds.ac.uk/10.3390/ruminants4010004
Baldin GC, Hildebrand C, Larson RL, Lancaster PA. Evaluation and Development of a Nutrition Model to Predict Intake and Growth of Suckling Calves. Ruminants. 2024; 4(1):47-78. https://0-doi-org.brum.beds.ac.uk/10.3390/ruminants4010004
Chicago/Turabian StyleBaldin, Geovana Camila, Caleb Hildebrand, Robert L. Larson, and Phillip A. Lancaster. 2024. "Evaluation and Development of a Nutrition Model to Predict Intake and Growth of Suckling Calves" Ruminants 4, no. 1: 47-78. https://0-doi-org.brum.beds.ac.uk/10.3390/ruminants4010004