
Cuticular Hydrocarbon Profiling of Australian Gonipterini Weevils


School of Health, Medical and Applied Sciences, Central Queensland University, North Rockhampton, QLD 4701, Australia
AppliedChem 2023, 3(3), 414-427; https://0-doi-org.brum.beds.ac.uk/10.3390/appliedchem3030026
Submission received: 23 March 2023
/
Revised: 24 July 2023
/
Accepted: 11 August 2023
/
Published: 17 August 2023
Abstract
:Cuticular hydrocarbon (CHC) profiling shows promise as a chemotaxonomic tool for identifying and discriminating between closely related insect species. However, there have been limited studies using CHC profiling to differentiate between weevil species (Coleoptera: Curculionidae). This proof-of-concept study investigated the use of CHC and volatile profiling to discriminate between five weevil species from three genera in the Gonipterini tribe. A total of 56 CHCs and 41 other volatile compounds were found across the five species, with 83 of the compounds being identified through their mass fragmentation patterns. The number of CHCs from each species ranged from 20 to 43, while the proportion of CHCs unique to each species varied between 0% and 19%. The most abundant CHCs were nonacosane, 7-methylheptacosane, heptacosane, and hexacosane. Principal component analysis of the centred log-ratio transformed data revealed broad differences in CHC profiles between the two Oxyops species, with Bryachus squamicollis demonstrating the greatest divergence from the other Gonipterini species. The results suggest that CHC analysis could be used to support established taxonomic methods, including morphological features and genetic sequencing results.
1. Introduction
Traditionally, insect taxonomy has been based on morphological features [1,2]. In the last few decades, genetic techniques such as DNA sequencing and barcoding have emerged as significant taxonomic tools [3,4]. Another complementary taxonomic technique is chemotaxonomy—the use of differences in biochemical composition between species to classify and/or identify them [5,6]. Originally used for the classification of plant species [7], the technique was subsequently extended to other organisms, such as insects. The major focus has been on cuticular hydrocarbons (CHCs) [8], which are found on the cuticles of virtually all insects, act to prevent desiccation, and serve as signalling molecules for communicating with other insects. CHCs are synthesised by the insect through a number of inter-linked anabolic pathways; hence, they are reflective of the genetic diversity and metabolic pathways of the species [9,10]. For several decades, CHC profiling has been used to classify various insect species [11,12]; however, it should be noted that this method is not without its drawbacks. These include high levels of intra-specific variation in some cases, environmental variation, and the challenges of defining CHC boundaries between species [11].
There have been a limited number of studies performed on the cuticular hydrocarbon profiles of weevils (Coleoptera: Curculionidae), despite the extensive diversity and ecological significance of this family. One early study by Baker and Nelson [13] investigated the cowpea weevil (Callosobruchus maculatus), finding that mono- and dimethyl branched-chain alkanes comprised the majority of CHCs in this species, with no difference in CHC profiles between sexes. Similarly, Lapointe et al. [14] investigated the Diaprepes root weevil (Diaprepes abbreviatus) and found no significant differences by sex or maturity stage. However, observations by Martins et al. [15] suggested that males of the rice water weevil (Oryzophagus oryzae) recognise females from their CHC profiles, indicating that some differentiation must be possible.
Finally, Souza et al. [16] recently demonstrated that the cuticular hydrocarbon profiles of several species of Gonipterus weevil agreed well with molecular sequencing data, suggesting that CHC profiling could be used for the accurate classification of species from this genus. These species are from the Gonipterini tribe, which encompasses the genera Bryachus (Pascoe 1870), Gonipterus (Schoenherr 1833), Iptergonus (Lea 1908), Oxyops (Schoenherr 1826), Pantoreites (Pascoe 1870), Prophaesia (Pascoe 1870), and Syarbis (Pascoe 1865). This tribe is native to the Australo-Pacific region, although some species (particularly Gonipterus spp.) have been accidentally translocated to various locations worldwide [17]. Both adults and larvae feed on Eucalyptus leaves. Outside of their native range, several species of Gonipterus have become significantly destructive pests of commercial Eucalyptus plantations [18] due to the absence of its natural parasitoids—principally, Anaphes nitens [19]. The Gonipterus genus in particular contains a number of cryptic species [17], which has posed significant barriers to the success of biocontrol programs [18]. Identification of such species typically requires molecular analysis and dissection of male genitalia [17].
If successful, rapid CHC profiling could provide an alternative to costly and time-consuming molecular sequencing and/or dissection techniques for the identification of morphologically cryptic species from this economically important tribe. Hence, this proof-of-concept study aimed to extend the CHC profiling method of Souza et al. [16] to discriminate between different Gonipterini genera, as well as between different species in specific genera (Gonipterus and Oxyops).
2. Materials and Methods
2.1. Specimen Collection
Fifteen weevils were hand collected from Eucalyptus populnea Muell. saplings in Central Queensland (23°46′ S, 150°21′ E) on 14 February 2021. They comprised five specimens of Oxyops fasciculatus Redtenbacher, three of an undescribed Oxyops sp. only known from this location (designated throughout this manuscript as Oxyops sp. 1), one specimen tentatively identified as Gonipterus sp. n. 2, three of Gonipterus cinnamomeus Pascoe, and three of Bryachus squamicollis Pascoe. Due to the limited number of Gonipterini weevils found during the fieldwork, a larger sample size was not possible for some species. Species delineations were confirmed by Dr Rolf Oberprieler (CSIRO, Canberra, Australia).
2.2. Extraction of CHCs
The CHC extraction methods followed those of Souza et al. [16]. After being killed in a freezer (−20 °C), each weevil was placed in a 2.0 mL GC-MS vial along with 300 μL of hexane. After 4 min, they were agitated for one minute by using a vortex mixer, and the hexane extract was collected. The weevil specimens were subsequently preserved in 100% ethanol.
2.3. Analysis of CHCs
The hexane extracts were analysed via gas chromatography-mass spectrometry (GC-MS) while following the methods of Souza et al. [16]. Analysis was performed on a single-quadrupole Shimadzu QP2010 Plus system (Shimadzu, Kyoto, Japan) fitted with an autoinjector/autosampler (AOC-20i/s) and Shimadzu SH-Rxi-5Sil MS column (29 m × 0.25 mm i.d. × 0.25 µm thickness). The following conditions were used: carrier gas—helium at 1.93 mL min−1, injection temperature—250 °C, injection volume—1 µL, split ratio 5:1, ion source temperature—230 °C, interface temperature—230 °C, MS mass range—35–600 m/z, and scan rate—3.3 scans/sec. During the run, the column temperature was initially held at 40 °C for 2 min before ramping linearly at 10 °C/min to reach 260 °C, where it was held for a further 6 min. The total run time was 30 min. Higher temperatures were not used in this study, as the main focus was on low- to moderate-weight CHCs, as studied by Souza et al. [16].
Chromatogram peaks were integrated if they had a peak area of >10,000 units and slope of >1000 units/min. Linear retention indices (LRIs) were calculated from the retention times of alkane standards (C8–C40) run under the same conditions [20]. Compound identities were established through the comparison of their mass spectra and LRIs with the NIST14 and NIST14s libraries and the relevant literature [14,21,22,23].
2.4. Chemometric Analysis
Data analysis of the volatile compound abundance was conducted in R Studio running R 4.0.5. [24]. Where applicable, the results are presented as the mean ± 1 standard deviation. The CHC dataset was transformed by using the centred log-ratio (clr) method prior to principal component analysis (PCA).
3. Results
3.1. Cuticular Hydrocarbon Profiles
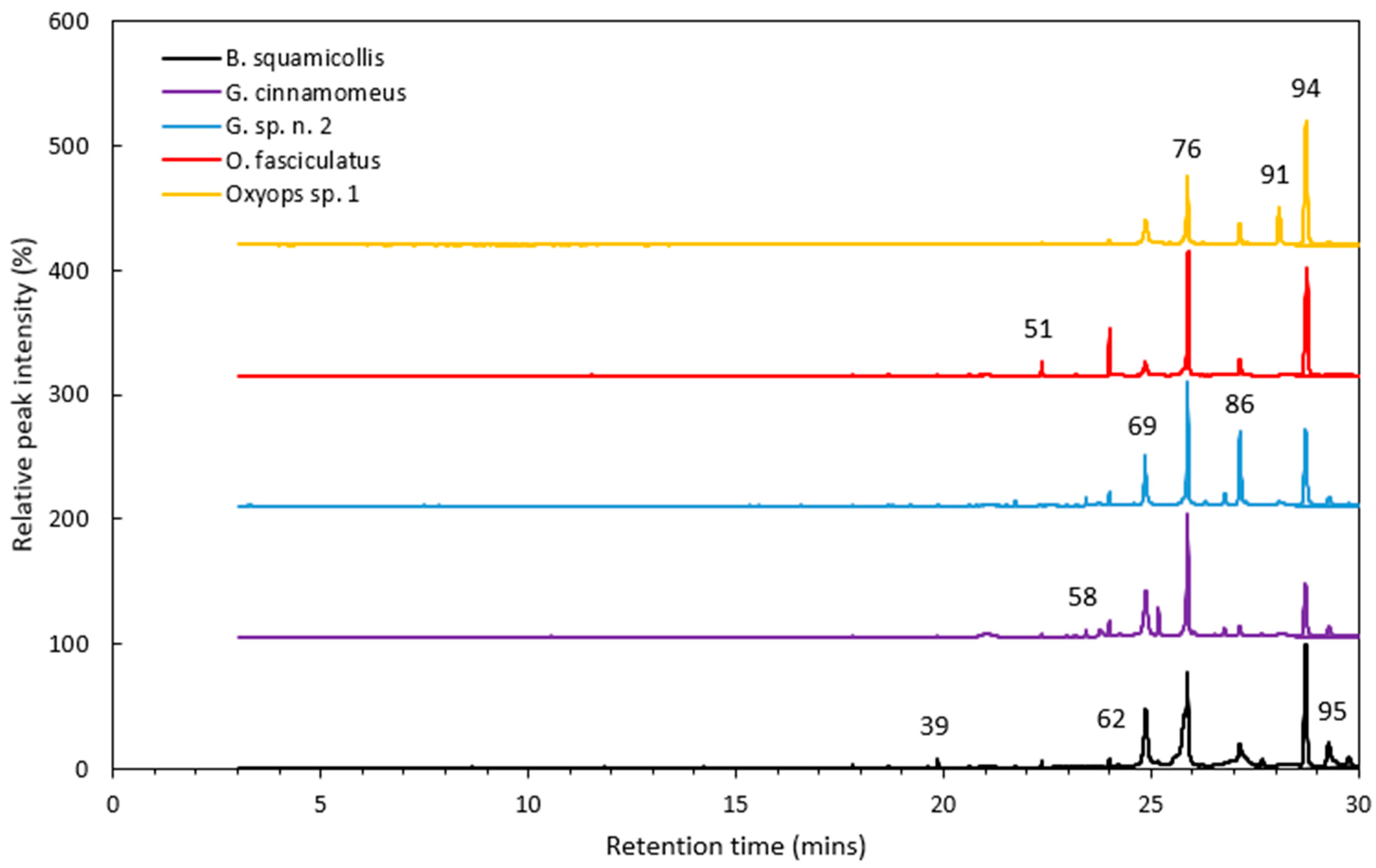
Typical GC-MS chromatograms obtained for each species are provided in Figure 1, while Table 1 shows the compounds identified across all Gonipterini species. A total of 97 peaks were found across all species, with 59 compounds being able to be positively identified from their mass fragmentation patterns and LRIs. A further 24 compounds were tentatively identified, with 14 compounds (10 alkanes, 3 ketones, and 1 aldehyde) being unable to be precisely identified. Some of the compounds identified (e.g., eucalyptol and globulol) appeared to be derived from the host plants (E. populnea), rather than being synthesised by the weevils. However, the majority of the compounds (56) could be classified as CHCs (Table 1). While all compounds are discussed in this section, only the CHC data were used in the subsequent chemometric analysis.
A total of 56 of the volatile compounds had some pheromone-type activity in one or more insect species (Table 2), with 43 being documented as having pheromone-type activity in Coleoptera [22]. Several of the compounds (aromadendrene and exo-2-hydroxycineole) have been previously identified as attractants for Gonipterus platensis [25]. Eucalyptol (1,8-cineole) is reportedly used as a defensive agent by Oxyops vitiosa larvae [26], in addition to acting as a potential attractant in adults of this species [27]. A number of 1,8-cineole metabolites have also been identified as pheromones in Gonipterus platensis [28]. No previous work was found on attractants for Bryachus.
Table 2 details the concentrations of the non-CHC volatile compounds found in each of the five Gonipterini species, while Table 3 compares the CHC contents among the species. The most abundant CHCs across all five species were nonacosane and 7-methylheptacosane. B. squamicollis also contained high levels of heptacosane, while both Gonipterus species showed high levels of hexacosane. Oxyops sp. 1 notably contained quite high concentrations (8.28 ± 6.05%) of 2-methyloctacosane, as well as a lower 7-methylheptacosane concentration than that of any other species.
The most abundant compound class was methyl alkanes (with a total of 18 compounds present), followed by aldehydes (16), n-alkanes (10), ketones (8), and dimethyl alkanes (8) (Table 2). As shown in Table 4, the greatest number of total compounds were found in B. squamicollis (71), and the greatest number of unique compounds was found only in this species (20, comprising 28.2% of the total volatile compounds found in this species). Gonipterus sp. n. 2 contained the lowest number of compounds (35), in addition to possessing only two unique compounds (henicosanal and an unidentified ketone). A total of 23 compounds were identified as being present across all five species.
3.2. Chemometric Analysis
To investigate the natural groupings in the CHC data, an unsupervised exploratory analysis was conducted on the CHC data only. Prior to the analysis, the volatile data were subjected to a centred log-ratio (clr) transformation, as recommended by Brückner and Heethoff [29] for similar datasets.
The principal component analysis (PCA) revealed a broad separation between B. squamicollis and the remaining species across the first principal component (PC 1), which explained 18.7% of the variation in the CHC dataset. The remaining species were largely separated across PC 2, which explained a further 12.9% of the variation (Figure 2). Most species were well separated across the first two PCs, although the single specimen of Gonipterus sp. n. 2 was quite close to the Oxyops sp. 1 cluster.
Examination of the PCA loadings plot (Figure 3) was used to investigate the compounds that were most strongly associated with particular Gonipterini species. For example, nonacosane was strongly associated with Oxyops sp. 1, while the large number of compounds loaded in the same direction as B. squamicollis supported previous observations about the large number of unique compounds found in this species (Table 3).
4. Discussion
Souza et al. [16] previously reported the CHC profile of Gonipterus sp. n. 2, with the major compounds present including n-heptacosane, 2-methylhexacosane, n-hexacosane, n-pentacosane, and n-octacosane. Somewhat contrasting results were found in this study, with the major compounds from this species being identified as 7-methylheptacosane, nonacosane, octacosane, hexacosane, 3-methylheptacosane, triacontane, heptacosane, and pentacosane. However, it should be noted that only one specimen from this species was analysed, so the results here may not necessarily be representative of the species as a whole. Another potential reason may be the difference in geographic locations. The present study used a specimen from central Queensland, while Souza et al. [16] collected Gonipterini specimens from a much wider region across Australia (Qld, NSW, ACT, Vic, WA). Studies have shown that CHC profiles can vary significantly with geographic location [16,30,31]. Finally, the species is part of a cryptic complex [17], so there is the possibility of misidentification, as genetic analysis was not performed in this study.
The major CHCs from G. cinnamomeus were found to be 7-methylheptacosane, nonacosane, hexacosane, and triacontane in this study, quite similarly to Gonipterus sp. n. 2. The CHC profile of this species does not appear to have been previously reported.
Souza et al. [16] also studied the CHC profiles of ten Oxyops specimens (not identified to species), reporting the major constituents as n-heptacosane, n-pentacosane, two unidentified compounds, and n-nonacosane. This largely concurred with the predominant CHCs found from O. fasciculatus in this study: nonacosane, 7-methylheptacosane, hexacosane, pentacosane, and octacosane. The CHC profile of Oxyops sp. 1 was somewhat less similar to the general Oxyops profile reported by Souza et al. [16]. The major constituents included nonacosane, 7-methylheptacosane, and hexacosane; however, it was unique in having a particularly high concentration of 2-methyloctacosane (8.28%) and the lowest concentration of pentacosane (0.97%) out of all species studied. This species (Oxyops sp. 1) has not yet been formally described yet; hence, its status in the Oxyops genus remains to be confirmed by a thorough morphological investigation and genetic study.
The results of the PCA supported B. squamicollis as the outgroup taxon. Within the remaining species, the Oxyops and Gonipterus species were loosely clustered together, but with some overlap.
Although CHC composition is primarily regulated through genetic means [9], it can be impacted by a range of factors, including diet [32,33], population age structure [34,35], locality, and climate [36,37]. However, a number of studies have found that CHC profiles are reasonably stable among different locations and ecological factors [16,38,39]. Furthermore, any impact of most of these variables would be expected to be minimal in this study, given that all specimens were collected on the same day from the same vicinity and were all collected from the same host plant species (E. populnea).
The overall results of this work support the prospect of using CHC profiles as a (relatively) rapid method of discriminating between Gonipterini genera and species. Such an approach has previously been applied across a range of insect orders to date, although the bulk of studies have been performed on Hymenoptera or Diptera [40,41,42]. CHC profiling shows particular promise when combined with other taxonomic techniques, including DNA barcoding and morphological analysis [40,43,44]. Such rapid analytical tools for discriminating between Gonipterini species could find use in a variety of applications, including identifying large numbers of specimens from field surveys or supporting the description of new species alongside DNA barcoding or morphological studies.
5. Conclusions
This study presented the cuticular hydrocarbon profiles of several Gonipterini species for the first time, including Bryachus squamicollis, Gonipterus cinnamomeus, and Oxyops fasciculatus. Principal component analysis revealed broadly differing CHC profiles between most species investigated, with B. squamicollis demonstrating the greatest divergence from the other Gonipterini genera/species. The results suggest that CHC analysis could be used to support established taxonomic methods, including the use of morphological features and genetic sequencing results.
Funding
Funding for this research was supported by a 2022 Research Grant from the Australian Entomological Society.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The full datasets supporting the findings of this research are available from the corresponding author upon request.
Conflicts of Interest
The author declares no conflict of interest.
References
- Cook, L.G.; Edwards, R.; Crisp, M.; Hardy, N. Need morphology always be required for new species descriptions? Invertebr. Syst. 2010, 24, 322–326. [Google Scholar] [CrossRef]
- Wheeler, Q.D. Undisciplined thinking: Morphology and Hennig’s unfinished revolution. Syst. Entomol. 2008, 33, 2–7. [Google Scholar] [CrossRef]
- DeSalle, R.; Goldstein, P. Review and Interpretation of Trends in DNA Barcoding. Front. Ecol. Evol. 2019, 7, 302. [Google Scholar] [CrossRef]
- Sharkey, M.J.; Janzen, D.H.; Hallwachs, W.; Chapman, E.G.; Smith, M.A.; Dapkey, T.; Brown, A.; Ratnasingham, S.; Naik, S.; Manjunath, R.; et al. Minimalist revision and description of 403 new species in 11 subfamilies of Costa Rican braconid parasitoid wasps, including host records for 219 species. ZooKeys 2021, 1013, 1–665. [Google Scholar] [CrossRef]
- Raupach, M.J.; Amann, R.; Wheeler, Q.D.; Roos, C. The application of “-omics” technologies for the classification and identification of animals. Org. Divers. Evol. 2016, 16, 774113. [Google Scholar] [CrossRef]
- Jones, O.A.; Maguire, M.L.; Griffin, J.L.; Dias, D.A.; Spurgeon, D.J.; Svendsen, C. Metabolomics and its use in ecology. Austral Ecol. 2013, 38, 713–720. [Google Scholar] [CrossRef]
- Hegnauer, R. Phytochemistry and plant taxonomy—An essay on the chemotaxonomy of higher plants. Phytochemistry 1986, 25, 1519–1535. [Google Scholar] [CrossRef]
- Martin, S.; Drijfhout, F. A Review of Ant Cuticular Hydrocarbons. J. Chem. Ecol. 2009, 35, 1151. [Google Scholar] [CrossRef]
- Holze, H.; Schrader, L.; Buellesbach, J. Advances in deciphering the genetic basis of insect cuticular hydrocarbon biosynthesis and variation. Heredity 2021, 126, 219–234. [Google Scholar] [CrossRef]
- Sprenger, P.P.; Hartke, J.; Schmitt, T.; Menzel, F.; Feldmeyer, B. Candidate genes involved in cuticular hydrocarbon differentiation between cryptic, parabiotic ant species. G3 Genes Genomes Genet. 2021, 11, jkab078. [Google Scholar] [CrossRef]
- Kather, R.; Martin, S.J. Cuticular hydrocarbon profiles as a taxonomic tool: Advantages, limitations and technical aspects. Physiol. Entomol. 2012, 37, 25–32. [Google Scholar] [CrossRef]
- Johnson, J. Near-infrared spectroscopy (NIRS) for taxonomic entomology: A brief review. J. Appl. Entomol. 2020, 144, 241–250. [Google Scholar] [CrossRef]
- Baker, J.E.; Nelson, D.R. Cuticular hydrocarbons of adults of the cowpea weevil, Callosobruchus maculatus. J. Chem. Ecol. 1981, 7, 175–182. [Google Scholar] [CrossRef]
- Lapointe, S.L.; Hunter, W.B.; Alessandro, R.T. Cuticular hydrocarbons on elytra of the Diaprepes root weevil Diaprepes abbreviatus (L.) (Coleoptera: Curculionidae). Agric. For. Entomol. 2004, 6, 251–257. [Google Scholar] [CrossRef]
- Martins, C.B.C.; Saad, E.B.; de Almeida, L.M.; Zarbin, P.H.G. Cuticular Compounds Recognition and Mating Behavior of the Rice Water Weevil Oryzophagus oryzae (Coleoptera, Curculionidae). J. Insect Behav. 2013, 26, 812–823. [Google Scholar] [CrossRef]
- Souza, N.M.; Schröder, M.L.; Hayes, R.A.; Bello, J.E.; Nahrung, H.F. Cuticular hydrocarbons of Gonipterus weevils: Are there species differences? Chemoecology 2021, 31, 159–167. [Google Scholar] [CrossRef]
- Mapondera, T.S.; Burgess, T.; Matsuki, M.; Oberprieler, R.G. Identification and molecular phylogenetics of the cryptic species of the Gonipterus scutellatus complex (Coleoptera: Curculionidae: Gonipterini). Aust. J. Entomol. 2012, 51, 175–188. [Google Scholar] [CrossRef]
- Schröder, M.L.; Slippers, B.; Wingfield, M.J.; Hurley, B.P. Invasion history and management of Eucalyptus snout beetles in the Gonipterus scutellatus species complex. J. Pest Sci. 2020, 93, 11–25. [Google Scholar] [CrossRef]
- Tooke, F. The eucalyptus snout-beetle, Gonipterus scutellatus Gyll. A study of its ecology and control by biological means. Entomol. Mem. 1953, 3, 1–282. [Google Scholar]
- van Den Dool, H.; Kratz, P.D. A generalization of the retention index system including linear temperature programmed gas—Liquid partition chromatography. J. Chromatogr. A 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Mackley, J.W.; Carlson, D.A.; Butler, J.F. Identification of the cuticular hydrocarbons of the horn fly and assays for attraction. J. Chem. Ecol. 1981, 7, 669–683. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.M. The Pherobase: Database of Insect Pheromones and Semiochemicals. Available online: https://www.pherobase.com/ (accessed on 7 July 2022).
- Carlson, D.A.; Bernier, U.R.; Sutton, B.D. Elution Patterns from Capillary GC for Methyl-Branched Alkanes. J. Chem. Ecol. 1998, 24, 1845–1865. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, version 4.0.2; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Branco, S.; Mateus, E.P.; da Silva, M.D.R.G.; Mendes, D.; Rocha, S.; Mendel, Z.; Schütz, S.; Paiva, M.R. Electrophysiological and behavioural responses of the Eucalyptus weevil, Gonipterus platensis, to host plant volatiles. J. Pest Sci. 2019, 92, 221–235. [Google Scholar] [CrossRef]
- Wheeler, G.S.; Massey, L.M.; Southwell, I.A. Antipredator Defense of Biological Control Agent Oxyops vitiosa Is Mediated by Plant Volatiles Sequestered from the Host Plant Melaleuca quinquenervia. J. Chem. Ecol. 2002, 28, 297–315. [Google Scholar] [CrossRef]
- Wheeler, G. Development of Pheromone-Based Trapping for the Melaleuca Quinquenervia Biological Control Agent, Oxyops vitiosa; University of Florida: Gainesville, FL, USA, 2016. [Google Scholar]
- Branco, S.; Mateus, E.P.; Gomes da Silva, M.D.R.; Mendes, D.; Pereira, M.M.A.; Schütz, S.; Paiva, M.R. Identification of pheromone candidates for the eucalyptus weevil, Gonipterus platensis (Coleoptera, Curculionidae). J. Appl. Entomol. 2020, 144, 41–53. [Google Scholar] [CrossRef]
- Brückner, A.; Heethoff, M. A chemo-ecologists’ practical guide to compositional data analysis. Chemoecology 2017, 27, 33–46. [Google Scholar] [CrossRef]
- Kula, C.; Amendt, J.; Drijfhout, F.P.; Moore, H.E. Geographical Variation of Cuticular Hydrocarbon Profiles of Adult Flies and Empty Puparia Amongst Three Populations of Calliphora vicina (Diptera: Calliphoridae). J. Med. Entomol. 2022, 60, 14–23. [Google Scholar] [CrossRef]
- Kota, M.V.; Heinen-Kay, J.L.; Zuk, M. Geographic variation in cuticular hydrocarbon profiles in Pacific field crickets. Ecol. Entomol. 2021, 46, 1118–1127. [Google Scholar] [CrossRef]
- Claudio-Piedras, F.; Recio-Tótoro, B.; Cime-Castillo, J.; Condé, R.; Maffei, M.; Lanz-Mendoza, H. Dietary and Plasmodium challenge effects on the cuticular hydrocarbon profile of Anopheles albimanus. Sci. Rep. 2021, 11, 11258. [Google Scholar] [CrossRef]
- Otte, T.; Hilker, M.; Geiselhardt, S. The Effect of Dietary Fatty Acids on the Cuticular Hydrocarbon Phenotype of an Herbivorous Insect and Consequences for Mate Recognition. J. Chem. Ecol. 2015, 41, 32–43. [Google Scholar] [CrossRef]
- Braga, M.V.; Pinto, Z.T.; de Carvalho Queiroz, M.M.; Blomquist, G.J. Effect of age on cuticular hydrocarbon profiles in adult Chrysomya putoria (Diptera: Calliphoridae). Forensic Sci. Int. 2016, 259, e37–e47. [Google Scholar] [CrossRef] [PubMed]
- Mpuru, S.; Blomquist, G.J.; Schal, C.; Roux, M.; Kuenzli, M.; Dusticier, G.; Clément, J.-L.; Bagnères, A.-G. Effect of age and sex on the production of internal and external hydrocarbons and pheromones in the housefly, Musca domestica. Insect Biochem. Mol. Biol. 2001, 31, 139–155. [Google Scholar] [CrossRef] [PubMed]
- Otte, T.; Hilker, M.; Geiselhardt, S. Phenotypic Plasticity of Cuticular Hydrocarbon Profiles in Insects. J. Chem. Ecol. 2018, 44, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Menzel, F.; Zumbusch, M.; Feldmeyer, B. How ants acclimate: Impact of climatic conditions on the cuticular hydrocarbon profile. Funct. Ecol. 2018, 32, 657–666. [Google Scholar] [CrossRef]
- Martin, S.J.; Helanterä, H.; Drijfhout, F.P. Evolution of species-specific cuticular hydrocarbon patterns in Formica ants. Biol. J. Linn. Soc. 2008, 95, 131–140. [Google Scholar] [CrossRef]
- van Zweden, J.S.; Dreier, S.; d’Ettorre, P. Disentangling environmental and heritable nestmate recognition cues in a carpenter ant. J. Insect Physiol. 2009, 55, 159–164. [Google Scholar] [CrossRef]
- Soon, V.; Castillo-Cajas, R.F.; Johansson, N.; Paukkunen, J.; Rosa, P.; Ødegaard, F.; Schmitt, T.; Niehuis, O. Cuticular Hydrocarbon Profile Analyses Help Clarify the Species Identity of Dry-Mounted Cuckoo Wasps (Hymenoptera: Chrysididae), Including Type Material, and Reveal Evidence for a Cryptic Species. Insect Syst. Divers. 2021, 5, 3. [Google Scholar] [CrossRef]
- Vaníčková, L.; Břízová, R.; Mendonça, A.L.; Pompeiano, A.; Do Nascimento, R.R. Intraspecific variation of cuticular hydrocarbon profiles in the Anastrepha fraterculus (Diptera: Tephritidae) species complex. J. Appl. Entomol. 2015, 139, 679–689. [Google Scholar] [CrossRef]
- Barbosa, R.R.; Braga, M.V.; Blomquist, G.J.; Queiroz, M.M.d.C. Cuticular hydrocarbon profiles as a chemotaxonomic tool for three blowfly species (Diptera: Calliphoridae) of forensic interest. J. Nat. Hist. 2017, 51, 1491–1498. [Google Scholar] [CrossRef]
- Vaníčková, L.; Virgilio, M.; Tomčala, A.; Břízová, R.; Ekesi, S.; Hoskovec, M.; Kalinová, B.; Do Nascimento, R.R.; De Meyer, M. Resolution of three cryptic agricultural pests (Ceratitis fasciventris, C. anonae, C. rosa, Diptera: Tephritidae) using cuticular hydrocarbon profiling. Bull. Entomol. Res. 2014, 104, 631–638. [Google Scholar] [CrossRef]
- Sakolsky, G.; Carlson, D.A.; Sutton, B.D.; Stoffolano, J.G., Jr. Detection of Cryptic Species in the Tabanus nigrovittatus (Diptera: Tabanidae) Complex in Massachusetts by Morphometric and Cuticular Hydrocarbon Analysis. J. Med. Entomol. 1999, 36, 610–613. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
GG-MS chromatogram of the hexane extracts from each Gonipterini species. The major peaks are marked; the compound numbers correspond to those provided in Table 1 and Table 2.

Figure 2.
Score plot showing the results of the principal component analysis performed on the clr-transformed CHC data.
Figure 2.
Score plot showing the results of the principal component analysis performed on the clr-transformed CHC data.

Figure 3.
PCA loadings plot showing the influence of individual CHCs on the principal component analysis performed on the clr-transformed CHC data.
Figure 3.
PCA loadings plot showing the influence of individual CHCs on the principal component analysis performed on the clr-transformed CHC data.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Identification details for the compounds found in the Gonipterini hexane extracts. Compounds classified as CHCs are highlighted in bold.
Table 1.
Identification details for the compounds found in the Gonipterini hexane extracts. Compounds classified as CHCs are highlighted in bold.
| No. | Compound | Class | LRI Rxi-5Sil | Lit. LRI | M+ (m/z) | Other Confirmatory MS Peaks (m/z) | Ident. ^ | Roles # (Coleoptera) | Roles # (Other Insects) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 3-hexanone | Ketone | 787 | 789 | 100 | 43, 57, 71 | MS, LRI | - | - |
| 2 | 2-hexanone | Ketone | 791 | 793 | 100 | 43, 58, 85, 71 | MS, LRI | - | - |
| 3 | 2,4-dimethylheptane | Dimethyl alkane | 820 | 822 | 128 | 43, 85, 57, 71 | MS, LRI | - | - |
| 4 | Heptanal | Aldehyde | 902 | 902 | 114 | 70, 44, 55, 57, 81, 86, 96 | MS, LRI | A, Al | A, Al, K, P |
| 5 | Octanal | Aldehyde | 1002 | 1001 | 128 | 57, 56, 84, 69, 95, 100, 110 | MS, LRI | A, Al | A, Al, K, P |
| 6 | Eucalyptol | Monoterpenoid | 1033 | 1033 | 154 | 81, 108, 139, 93 | MS, LRI | A, Al, P | A, K, P |
| 7 | 3,6-dimethyldecane | Dimethyl alkane | 1055 | 1086 | 170 | 57, 71, 85, 113, 127 | MS, LRI | - | - |
| 8 | 2,6,8-trimethyldecane | Trimethyl alkane | 1099 | 1104 | 184 | 85, 99, 127, 113, 155 | MS, LRI | - | - |
| 9 | Nonanal | Aldehyde | 1104 | 1108 | 142 | 57, 70, 82, 98, 95, 96, 114 | MS, LRI | A, Al, P | A, Al, K, P |
| 10 | Decanal | Aldehyde | 1205 | 1204 | 156 | 57, 70, 82, 95, 112, 128 | MS, LRI | A, Al, K, P | A, K, P |
| 11 | Exo-2-hydroxycineole | 1228 | 1228 | 170 | 108, 126, 93 | MS, LRI | - | P | |
| 12 | 2,6,10-trimethylundecane † | Trimethyl alkane | 1275 | 1275 | 198 | 57, 71, 85, 99, 127, 113, 155 | MS, LRI | - | - |
| 13 | 10-undecenal | Alkene aldehyde | 1282 | 1277 | 168 | 55, 67, 81, 97, 111, 135 | MS, LRI | - | - |
| 14 | Carvacrol † | Monoterpenoid | 1297 | 1298 | 150 | 81, 93, 135, 121 | MS, LRI | - | P |
| 15 | Isoascaridole † | 1312 | 1303 | 168 | 95, 110, 81, 139 | MS, LRI | - | - | |
| 16 | 4a-methyldecahydro-1-naphthalenol † | 1319 | 1363 | 168 | 95, 67, 97, 135, 121 | MS, LRI | - | - | |
| 17 | 4,6-dimethyldodecane | Dimethyl alkane | 1321 | 1325 | 198 | 85, 99, 113, 127, 155 | MS, LRI | - | - |
| 18 | cis-p-menth-1-en-3,8-diol † | 1358 | 1362 | 170 | 84, 71, 109, 138 | MS, LRI | - | - | |
| 19 | (+)-cis,trans-nepetalactone | Iridoid | 1364 | 1365 | 166 | 81, 95, 123, 109, 138 | MS, LRI | Al | A, Al |
| 20 | Dodecanal | Aldehyde | 1408 | 1407 | 184 | 57, 82, 96, 110, 140, 123 | MS, LRI | Al | K, P |
| 21 | Aromadendrene | Sesquiterpenoid | 1444 | 1440 | 204 | 161, 105, 133, 189 | MS, LRI | - | A |
| 22 | Unidentified hydrocarbon 1 | - | 1488 | - | - | 57, 71, 85, 99, 113, 127, 141, 155, 169 | MS | - | - |
| 23 | Bicyclogermacrene | Sesquiterpenoid | 1501 | 1494 | 204 | 121, 161, 136, 189 | MS, LRI | - | A, P |
| 24 | 2,6,10-trimethyltridecane † | Trimethyl alkane | 1534 | 1540 | 226 | 99, 113, 127, 155, 141, 169 | MS, LRI | - | - |
| 25 | Globulol | Sesquiterpenoid | 1592 | 1604 | 222 | 107, 109, 161, 189, 204 | MS, LRI | - | A |
| 26 | Tetradecanal | Aldehyde | 1612 | 1611 | 212 | 57, 82, 96, 124, 168 | MS, LRI | Al | A, Al, P |
| 27 | Heptadecane | n-alkane | 1699 | 1700 | 240 | 169, 183, 197 | MS, LRI | A, Al, P | A, Al, P |
| 28 | Phytane | Branched alkane | 1743 | 1753 | 282 | 127, 155, 169, 197, 211 | MS, LRI | - | - |
| 29 | cis-9-hexadecenal | Alkene aldehyde | 1795 | 1800 | 238 | 55, 69, 81, 93, 111, 121, 135, 149 | MS, LRI | - | A, P |
| 30 | Hexadecanal | Aldehyde | 1816 | 1819 | 240 | 57, 82, 96, 110, 124, 138, 165, 194, 222 | MS, LRI | Al, P | A, Al, P |
| 31 | 6,10,14-trimethyl-2-pentadecanone | Branched ketone | 1840 | 1842 | 268 | 58, 71, 85, 95, 109, 124, 137, 165, 250 | MS, LRI | - | - |
| 32 | 2-heptadecanone | Ketone | 1899 | 1886 | 254 | 58, 71, 96, 127, 166 | MS, LRI | - | - |
| 33 | 2,2-dimethyloctadecane † | Dimethyl alkane | 1910 | 1917 | 282 | 127, 155, 141, 169, 183, 197, 211, 239 | MS, LRI | - | - |
| 34 | Heptadecanal | Aldehyde | 1918 | 1920 | 254 | 138, 152, 166, 180, 194, 210, 226, 236 | MS, LRI | - | Al, P |
| 35 | 3-ethyl-3-methylheptadecane † | Branched alkane | 1953 | 1956 | 282 | 127, 141, 155, 169, 183, 197, 223 | MS, LRI | - | - |
| 36 | 9-octadecanone | Ketone | 1990 | 1980 | 268 | 71, 95, 141, 156, 169, 211, 254 | MS, LRI | - | - |
| 37 | cis-13-octadecenal | Alkene aldehyde | 1995 | 1985 | 266 | 69, 81, 83, 95, 98, 111, 121, 135, 166, 248 | MS, LRI | - | A, P |
| 38 | cis-9-octadecenal † | Alkene aldehyde | 2014 | 2007 | 266 | 55, 69, 96, 121 | MS, LRI | P | P |
| 39 | Octadecanal | Aldehyde | 2019 | 2021 | 268 | 124, 138, 152, 166, 180, 194, 222, 250 | MS, LRI | Al, P | P |
| 40 | cis-2-octadecen-1-ol acetate | Ester | 2074 | 2086 | 310 | 55, 69, 81, 97, 136 | MS, LRI | - | - |
| 41 | 2-nonadecanone | Ketone | 2098 | 2101 | 282 | 58, 71, 85, 96, 100, 127, 138, 152, 267, 282 | MS, LRI | - | - |
| 42 | Nonadecanal | Aldehyde | 2117 | 2105 | 282 | 82, 96, 109, 124, 138, 152, 166, 180 | MS, LRI | - | P |
| 43 | Unidentified hydrocarbon 2 | - | 2128 | - | - | 211, 225, 239, 253, 267, 281, 295 | MS | - | - |
| 44 | Unidentified hydrocarbon 3 | - | 2139 | - | - | 127, 155, 183, 211, 239, 267 | MS | - | - |
| 45 | Unidentified hydrocarbon 4 | - | 2148 | - | - | 225, 238, 252, 267, 295 | MS | - | - |
| 46 | Unidentified hydrocarbon 5 | - | 2160 | - | - | 155, 169, 183, 253, 197 | MS | - | - |
| 47 | Unidentified hydrocarbon 6 | - | 2168 | - | - | 99, 127, 155, 183 | MS | - | - |
| 48 | Docosane | n-alkane | 2197 | 2200 | 310 | 155, 169, 183, 196, 211, 239, 267 | MS, LRI | P | A, Al, P |
| 49 | Eicosanal | Aldehyde | 2222 | 2224 | 296 | 278, 250, 152, 124 | MS, LRI | Al | P |
| 50 | Unidentified hydrocarbon 7 a | - | 2260 | - | - | 127, 141, 155, 169, 183, 197, 211, 225, 239, 253, 267, 281 | MS | - | - |
| 51 | Tricosane | n-alkane | 2297 | 2300 | 324 | 225, 239, 253, 267, 281, 295 | MS, LRI | A, Al, P | A, Al, K, P |
| 52 | Unidentified ketone 1 | Ketone | 2304 | - | - | 58, 59, 71, 85, 96, 127 | MS | - | - |
| 53 | Henicosanal | Aldehyde | 2326 | 2329 | 310 | 82, 96, 110, 124, 209 | MS, LRI | - | - |
| 54 | 11-methyltricosane | Methyl alkane | 2331 | 2330 | 338 | 99, 113, 127, 141, 155, 169, 196, 211, 239 | MS, LRI | - | P |
| 55 | Unidentified aldehyde | Aldehyde | 2367 | - | - | 82, 97, 109, 125, 139, 180 | MS | - | - |
| 56 | 3-methyltricosane | Methyl alkane | 2374 | 2375 | 338 | 57, 71, 85, 96, 141, 183, 239 | MS, LRI | P | P |
| 57 | Tetracosane | n-alkane | 2400 | 2400 | 338 | 267, 281, 295, 309 | MS, LRI | P | A, Al, P |
| 58 | Docosanal | Aldehyde | 2430 | 2430 | 324 | 82, 96, 152, 166, 250, 278, 306 | MS, LRI | P | P |
| 59 | 9-methyltetracosane † | Methyl alkane | 2437 | 2433 | 352 | 99, 113, 127, 141, 155, 169, 183, 197 | MS, LRI | P | P |
| 60 | 2-methyltetracosane | Methyl alkane | 2473 | 2465 | 352 | 309, 267, 281, 295, 337 | MS, LRI | P | P |
| 61 | x-pentacosene † | Alkene | 2479 | 2477 | 350 | 168, 181, 197, 211, 225, 239, 253, 267 | MS, LRI | - | - |
| 62 | Pentacosane | n-alkane | 2499 | 2500 | 352 | 267, 281, 295, 309, 323 | MS, LRI | A, Al, P | A, Al, K, P |
| 63 | Unidentified ketone 2 b | Ketone | 2509 | - | - | 58, 59, 71, 85, 96, 239 | MS | - | - |
| 64 | 7-methylpentacosane † | Methyl alkane | 2522 | 2530 | 362 | 113, 127, 141, 155, 169, 183, 197, 224, 239, 253, 267, 281, 295, 309 | MS, LRI | P | - |
| 65 | 11-methylpentacosane † | Methyl alkane | 2530 | 2530 | 366 | 168, 169, 196, 224, 225 | MS, LRI | P | P |
| 66 | 13-methylpentacosane † | Methyl alkane | 2569 | 2530 | 366 | 127, 139, 141, 169, 183, 197, 225, 229, 254 | MS, LRI | P | P |
| 67 | 3-methylpentacosane | Methyl alkane | 2574 | 2574 | 366 | 337, 336, 253, 267, 281, 309 | MS, LRI | P | Al, P |
| 68 | 11,15-dimethylpentacosane † | Dimethyl alkane | 2584 | 2550 | 380 | 168, 169, 239 | MS, LRI | P | P |
| 69 | Hexacosane | n-alkane | 2600 | 2600 | 366 | 281, 295, 309, 323, 337, 351 | MS, LRI | P | A, Al, P |
| 70 | Tetracosanal | Aldehyde | 2637 | 2632 | 352 | 334, 306, 278, 264, 250 | MS, LRI | P | P |
| 71 | 2-methylhexacosane | Methyl alkane | 2663 | 2664 | 380 | 280, 337, 364, 365 | MS, LRI | P | P |
| 72 | Unidentified hydrocarbon 8 | - | 2672 | - | - | 337, 211, 225, 253, 351 | MS | - | - |
| 73 | 13-methylhexacosane † | Methyl alkane | 2682 | 2633 | 380 | 196, 197, 308, 309, 211, 280 | MS, LRI | P | P |
| 74 | Unidentified hydrocarbon 9 | - | 2690 | - | - | 99, 97, 113, 127, 169, 225, 280 | MS | - | - |
| 75 | Heptacosane | n-alkane | 2704 | 2700 | 380 | 323, 337, 351, 365 | MS, LRI | P | Al, P |
| 76 | 7-methylheptacosane † | Methyl alkane | 2712 | 2730 | 394 | 99, 97, 113, 127, 141, 155, (224), 225, 309, 337 | MS, LRI | P | - |
| 77 | Unidentified ketone 3 | Ketone | 2723 | - | - | 59, 58, 96, 111, 125, 137, 250 | MS | - | - |
| 78 | 13-methylheptacosane | Methyl alkane | 2737 | 2733 | 394 | 168, 196, 197, 224, 253 | MS, LRI | P | P |
| 79 | Unidentified hydrocarbon 10 | - | 2755 | - | - | 267, 295, 195, 197, 224 | MS | - | - |
| 80 | 11-methylheptacosane † | Methyl alkane | 2759 | 2734 | 394 | 127, 141, 155, 168, 169, 239, 252, 253 | MS, LRI | P | P |
| 81 | 2-methylheptacosane | Methyl alkane | 2764 | 2760 | 394 | 141, 183, 351 | MS, LRI | P | P |
| 82 | Docosyl pentyl ether | Ether | 2770 | 2775 | 396 | 71, 83, 97, 111, 125, 139, 153, 167 | MS, LRI | - | - |
| 83 | 3-methylheptacosane | Methyl alkane | 2774 | 2773 | 394 | 365, 267, 281, 295, 309, 337 | MS, LRI | P | P |
| 84 | 5,15- or 5,17-dimethylheptacosane † | Dimethyl alkane | 2777 | 2778 | 408 | 168, 127, 155, 211, 239 | MS, LRI | - | - |
| 85 | 5,11-dimethylheptacosane † | Dimethyl alkane | 2784 | 2783 | 408 | 99, 113, 127, 141, 155, 168, 169, 239 | MS, LRI | - | P |
| 86 | Octacosane | n-alkane | 2800 | 2800 | 394 | 337, 351, 365, 379 | MS, LRI | A, P | A, P |
| 87 | Squalene | Triterpenoid | 2811 | 2790 | 410 | 69, 81, 95, 121, 136, 137, 149 | MS, LRI | P | A, P |
| 88 | 12-methyloctacosane † | Methyl alkane | 2830 | 2829 | 408 | 224, 210, 211, 182, 183, 197 | MS, LRI | P | P |
| 89 | Hexacosanal | Aldehyde | 2837 | 2834 | 380 | 57, 71, 82, 96, 111, 124, 180, 362 | MS, LRI | - | P |
| 90 | x-methyloctacosane † | Methyl alkane | 2858 | 2864 | 408 | 365, 253, 281, 295 | MS, LRI | - | - |
| 91 | 2-methyloctacosane | Methyl alkane | 2865 | 2864 | 408 | 365, 253, 267, 281, 295, 309 | MS, LRI | P | P |
| 92 | Nonacosene | Alkene | 2881 | 2888 | 406 | 97, 83, 125, 167, 195 | MS, LRI | - | - |
| 93 | 1-hexacosanol | Fatty alcohol | 2890 | 2865 | 382 | 57, 97, 83, 69, 71, 111, 125, 153, 167, 181, 195, 209 | MS, LRI | - | P |
| 94 | Nonacosane | n-alkane | 2918 | 2900 | 408 | 337, 351, 365, 379, 393 | MS, LRI | P | A, Al, P |
| 95 | Triacontane | n-alkane | 2982 | 3000 | 422 | 168, 169, 224, 197 | MS, LRI | P | A, P |
| 96 | x,12-dimethylnonacosane † | Dimethyl alkane | 3002 | 3000 | 437 | 112, 113, 169, 182, 183, 336, 337 | MS, LRI | - | - |
| 97 | 2-methyltriacontane † | Methyl alkane | 3039 | 3058 | 437 | 239, 224, 337, 365 | MS, LRI | P | P |
Table 2.
Non-CHC volatile compounds identified in the Gonipterini hexane extracts by using GC-MS. Compounds were quantified as relative percentages of the total peak areas in the total ion chromatogram (TIC).
Table 2.
Non-CHC volatile compounds identified in the Gonipterini hexane extracts by using GC-MS. Compounds were quantified as relative percentages of the total peak areas in the total ion chromatogram (TIC).
| No. | Compound | B. squamicollis (n = 3) | G. cinnamomeus (n = 3) | G. sp. n. 2 (n = 1) | O. fasciculatus (n = 5) | Oxyops sp. 1 (n = 3) | p Value |
|---|---|---|---|---|---|---|---|
| 1 | 3-hexanone | 0.03 ± 0.01 | 0.06 ± 0.01 | 0.08 | 0.05 ± 0.04 | 0.04 ± 0.00 | NS |
| 2 | 2-hexanone | 0.02 ± 0.02 | 0.07 ± 0.01 | 0.07 | 0.06 ± 0.04 | 0.04 ± 0.01 | NS |
| 4 | Heptanal | 0.03 ± 0.01 | 0.03 ± 0.02 | 0 | 0 | 0 | * |
| 5 | Octanal | 0.01 ± 0.01 | 0 | 0 | 0 | 0 | NS |
| 6 | Eucalyptol | 0 | 0 | 0.10 | 0.01 ± 0.02 | 0 | *** |
| 9 | Nonanal | 0.13 ± 0.05 | 0 | 0 | 0 | 0 | *** |
| 10 | Decanal | 0.01 ± 0.01 | 0.02 ± 0.04 | 0 | 0 | 0 | NS |
| 11 | Exo-2-hydroxycineole | 0.04 ± 0.02 | 0.08 ± 0.04 | 0 | 0.02 ± 0.02 | 0 | * |
| 13 | 10-undecenal | 0.14 ± 0.17 | 0 | 0 | 0 | 0 | NS |
| 14 | Tentative: carvacrol | 2.42 ± 3.83 | 0.04 ± 0.07 | 0 | 0.02 ± 0.04 | 0 | NS |
| 15 | Tentative: isoascaridole | 0.01 ± 0.01 | 0 | 0 | 0 | 0 | NS |
| 16 | Tentative: 4a-methyldecahydro-1-naphthalenol | 0.21 ± 0.23 | 0 | 0 | 0 | 0 | NS |
| 18 | Tentative: cis-p-menth-1-en-3,8-diol | 0.02 ± 0.03 | 0 | 0 | 0 | 0 | NS |
| 19 | (+)-cis,trans-nepetalactone | 0.02 ± 0.03 | 0 | 0 | 0 | 0 | NS |
| 20 | Dodecanal | 0 | 0.02 ± 0.03 | 0 | 0.01 ± 0.01 | 0.01 ± 0.02 | NS |
| 21 | Aromadendrene | 0 | 0 | 0 | 0.01 ± 0.03 | 0 | NS |
| 23 | Bicyclogermacrene | 0.03 ± 0.05 | 0 | 0 | 0.02 ± 0.02 | 0 | NS |
| 25 | Globulol | 0 | 0 | 0.07 | 0.01 ± 0.03 | 0 | NS |
| 26 | Tetradecanal | 0.04 ± 0.02 | 0 | 0.05 | 0.01 ± 0.02 | 0.02 ± 0.03 | NS |
| 29 | cis-9-hexadecenal | 0.05 ± 0.01 | 0 | 0 | 0 | 0 | *** |
| 30 | Hexadecanal | 0.17 ± 0.05 | 0.09 ± 0.04 | 0.10 | 0.16 ± 0.13 | 0.13 ± 0.08 | NS |
| 31 | 6,10,14-trimethyl-2-pentadecanone | 0 | 0 | 0 | 0 | 0.01 ± 0.02 | NS |
| 32 | 2-heptadecanone | 0.09 ± 0.02 | 0.07 ± 0.03 | 0.14 | 0.04 ± 0.05 | 0.06 ± 0.02 | NS |
| 36 | 9-octadecanone | 0 | 0 | 0 | 0.1 ± 0.1 | 0 | NS |
| 37 | cis-13-octadecenal | 0.12 ± 0.03 | 0 | 0 | 0 | 0 | *** |
| 38 | Tentative: cis-9-octadecenal | 0 | 0 | 0 | 0.03 ± 0.07 | 0 | NS |
| 39 | Octadecanal | 0.61 ± 0.14 | 0.07 ± 0.04 | 0.13 | 0.19 ± 0.18 | 0.16 ± 0.13 | ** |
| 40 | cis-2-octadecen-1-ol acetate | 0 | 0 | 0 | 0.01 ± 0.01 | 0 | NS |
| 41 | 2-nonadecanone | 0.09 ± 0.02 | 0.04 ± 0.04 | 0.18 | 0.02 ± 0.05 | 0.08 ± 0.05 | NS |
| 42 | Nonadecanal | 0.06 ± 0.01 | 0 | 0 | 0 | 0 | *** |
| 49 | Eicosanal | 0.09 ± 0.04 | 0 | 0.71 | 0.03 ± 0.07 | 0 | *** |
| 52 | Unidentified ketone 1 | 0 | 0 | 0.09 | 0 | 0 | *** |
| 53 | Henicosanal | 0 | 0 | 0.50 | 0 | 0 | *** |
| 55 | Unidentified aldehyde | 0.02 ± 0.01 | 0 | 0 | 0 | 0 | * |
| 58 | Docosanal | 0.01 ± 0.01 | 0.58 ± 0.31 | 1.12 | 0 | 0 | *** |
| 63 | Unidentified ketone 2 | 0 | 0.05 ± 0.09 | 0.13 | 0 | 0 | NS |
| 70 | Tetracosanal | 1.95 ± 2.56 | 4.22 ± 0.91 | 0 | 0 | 0.16 ± 0.27 | ** |
| 77 | Unidentified ketone 3 | 0 | 0.75 ± 0.31 | 0 | 0 | 0 | *** |
| 82 | Docosyl pentyl ether | 0 | 0 | 0 | 0 | 0.27 ± 0.47 | NS |
| 87 | Squalene | 0.38 ± 0.65 | 0.36 ± 0.37 | 0.71 | 0.43 ± 0.20 | 0.27 ± 0.08 | NS |
| 89 | Hexacosanal | 0.94 ± 0.24 | 0.94 ± 0.3 | 0 | 0 | 0 | *** |
NS = not significant (p > 0.05), * p < 0.05, ** p < 0.01, *** p < 0.001.
Table 3.
Cuticular hydrocarbons (CHCs) identified in the Gonipterini hexane extracts by using GC-MS. Compounds were quantified as relative percentages of the total peak areas in the total ion chromatogram (TIC).
Table 3.
Cuticular hydrocarbons (CHCs) identified in the Gonipterini hexane extracts by using GC-MS. Compounds were quantified as relative percentages of the total peak areas in the total ion chromatogram (TIC).
| No. | Compound | B. squamicollis (n = 3) | G. cinnamomeus (n = 3) | G. sp. n. 2 (n = 1) | O. fasciculatus (n = 5) | Oxyops sp. 1 (n = 3) | p Value |
|---|---|---|---|---|---|---|---|
| 3 | 2,4-dimethylheptane | 0.02 ± 0.01 | 0.02 ± 0.02 | 0.05 | 0.03 ± 0.02 | 0.02 ± 0.00 | NS |
| 7 | 3,6-dimethyldecane | 0.03 ± 0.01 | 0.06 ± 0.02 | 0.06 | 0.05 ± 0.03 | 0.03 ± 0.01 | NS |
| 8 | 2,6,8-trimethyldecane | 0.02 ± 0.02 | 0.02 ± 0.03 | 0 | 0.01 ± 0.03 | 0.01 ± 0.01 | NS |
| 12 | Tentative: 2,6,10-trimethylundecane | 0.03 ± 0.01 | 0.06 ± 0.01 | 0.05 | 0.06 ± 0.03 | 0.03 ± 0.00 | NS |
| 17 | 4,6-dimethyldodecane | 0.03 ± 0.01 | 0.02 ± 0.03 | 0 | 0.03 ± 0.03 | 0.01 ± 0.02 | NS |
| 22 | Unidentified hydrocarbon 1 | 0.01 ± 0.01 | 0.01 ± 0.02 | 0.06 | 0.05 ± 0.02 | 0.03 ± 0.00 | NS |
| 24 | Tentative: 2,6,10-trimethyltridecane | 0.01 ± 0.01 | 0.01 ± 0.02 | 0 | 0 | 0 | NS |
| 27 | Heptadecane | 0.03 ± 0.01 | 0.05 ± 0.01 | 0.05 | 0.04 ± 0.03 | 0.02 ± 0.02 | NS |
| 28 | Phytane | 0.01 ± 0.02 | 0 | 0 | 0 | 0 | NS |
| 33 | Tentative: 2,2-dimethyloctadecane | 0.04 ± 0.02 | 0.06 ± 0.06 | 0.09 | 0.03 ± 0.03 | 0.02 ± 0.03 | NS |
| 34 | Heptadecanal | 0.05 ± 0.01 | 0 | 0 | 0 | 0 | *** |
| 35 | Tentative: 3-ethyl-3-methylheptadecane | 0.03 ± 0.02 | 0.06 ± 0.02 | 0.05 | 0.06 ± 0.03 | 0.03 ± 0.00 | NS |
| 43 | Unidentified hydrocarbon 2 | 0.23 ± 0.21 | 0.25 ± 0.43 | 0 | 0.03 ± 0.07 | 0.12 ± 0.22 | NS |
| 44 | Unidentified hydrocarbon 3 | 0 | 0.35 ± 0.60 | 0 | 0 | 0.23 ± 0.40 | NS |
| 45 | Unidentified hydrocarbon 4 | 0.48 ± 0.46 | 0.42 ± 0.73 | 0 | 0.06 ± 0.14 | 0.28 ± 0.48 | NS |
| 46 | Unidentified hydrocarbon 5 | 0.10 ± 0.08 | 0.14 ± 0.24 | 0.07 | 0.04 ± 0.02 | 0.01 ± 0.02 | NS |
| 47 | Unidentified hydrocarbon 6 | 0.08 ± 0.08 | 0.09 ± 0.15 | 0 | 0 | 0.28 ± 0.49 | NS |
| 48 | Docosane | 0.02 ± 0.02 | 0 | 0 | 0.06 ± 0.05 | 0 | NS |
| 50 | Unidentified hydrocarbon 7 | 0 | 0 | 0 | 0 | 0.03 ± 0.03 | * |
| 51 | Tricosane | 0.74 ± 0.19 | 0.37 ± 0.09 | 0 | 1.35 ± 0.54 | 0.27 ± 0.06 | ** |
| 54 | 11-methyltricosane | 0.20 ± 0.34 | 0 | 0 | 0 | 0.02 ± 0.04 | NS |
| 56 | 3-methyltricosane | 0.02 ± 0.03 | 0 | 0 | 0 | 0 | NS |
| 57 | Tetracosane | 0.03 ± 0.02 | 0.07 ± 0.01 | 0.16 | 0.09 ± 0.05 | 0.02 ± 0.02 | * |
| 59 | Tentative: 9-methyltetracosane | 0.01 ± 0.02 | 0 | 0 | 0 | 0 | NS |
| 60 | 2-methyltetracosane | 0 | 0.94 ± 1.64 | 0 | 0 | 0.69 ± 1.19 | NS |
| 61 | Tentative: x-pentacosene | 0 | 0 | 0 | 0.03 ± 0.07 | 0 | NS |
| 62 | Pentacosane | 1.67 ± 0.93 | 1.83 ± 0.37 | 1.82 | 4.26 ± 2.02 | 0.97 ± 0.88 | NS |
| 64 | Tentative: 7-methylpentacosane | 0 | 0.27 ± 0.46 | 0 | 0.48 ± 0.42 | 0 | NS |
| 65 | Tentative: 11-methylpentacosane | 0 | 0.15 ± 0.26 | 0 | 0.11 ± 0.14 | 0 | NS |
| 66 | Tentative: 13-methylpentacosane | 0.03 ± 0.05 | 0 | 0 | 0 | 0 | NS |
| 67 | 3-methylpentacosane | 0.26 ± 0.23 | 1.09 ± 0.64 | 0.27 | 0 | 0 | ** |
| 68 | Tentative: 11,15-dimethylpentacosane | 0 | 0 | 0 | 0.70 ± 1.57 | 0 | NS |
| 69 | Hexacosane | 9.04 ± 6.87 | 13.26 ± 7.6 | 14.70 | 8.30 ± 7.10 | 11.01 ± 8.76 | NS |
| 71 | 2-methylhexacosane | 0 | 0 | 0 | 0.03 ± 0.07 | 0 | NS |
| 72 | Unidentified hydrocarbon 8 | 0 | 0.51 ± 0.46 | 0 | 0 | 0.15 ± 0.15 | NS |
| 73 | Tentative: 13-methylhexacosane | 0.65 ± 1.12 | 0.02 ± 0.04 | 0 | 0.88 ± 1.37 | 0 | NS |
| 74 | Unidentified hydrocarbon 9 | 0.36 ± 0.62 | 0 | 0 | 0 | 0 | NS |
| 75 | Heptacosane | 11.32 ± 2.95 | 4.3 ± 4.94 | 2.27 | 1.54 ± 3.44 | 0.36 ± 0.62 | * |
| 76 | Tentative: 7-methylheptacosane | 21.49 ± 5.33 | 28.94 ± 7.24 | 25.45 | 27.58 ± 6.29 | 16.77 ± 6.68 | NS |
| 78 | 13-methylheptacosane | 0.32 ± 0.06 | 0.07 ± 0.13 | 0 | 0.08 ± 0.15 | 0.73 ± 0.24 | ** |
| 79 | Unidentified hydrocarbon 10 | 1.14 ± 1.2 | 0 | 0 | 0.01 ± 0.02 | 0.10 ± 0.17 | NS |
| 80 | Tentative: 11-methylheptacosane | 0 | 4.60 ± 4.16 | 0 | 1.18 ± 2.49 | 0.14 ± 0.23 | NS |
| 81 | 2-methylheptacosane | 0 | 0 | 0 | 0 | 0.08 ± 0.07 | * |
| 83 | 3-methylheptacosane | 0 | 3.14 ± 1.61 | 2.82 | 0 | 0 | *** |
| 84 | Tentative: 5,15- or 5,17-dimethylheptacosane | 1.06 ± 1.07 | 0 | 0 | 0 | 0 | NS |
| 85 | Tentative: 5,11-dimethylheptacosane | 0.36 ± 0.62 | 1.93 ± 3.35 | 0 | 3.93 ± 7.21 | 0.79 ± 1.37 | NS |
| 86 | Octacosane | 6.89 ± 2.27 | 3.55 ± 0.89 | 19.10 | 4.16 ± 0.85 | 4.43 ± 1.34 | *** |
| 88 | Tentative: 12-methyloctacosane | 0.22 ± 0.14 | 0.05 ± 0.08 | 0 | 0 | 0 | * |
| 90 | Tentative: x-methyloctacosane | 0.05 ± 0.05 | 0 | 0.11 | 0 | 0 | *** |
| 91 | 2-methyloctacosane | 0.24 ± 0.14 | 0.93 ± 0.22 | 0.96 | 0 | 8.28 ± 6.05 | ** |
| 92 | Nonacosene | 1.03 ± 0.40 | 0 | 0 | 0 | 0 | * |
| 93 | 1-hexacosanol | 0 | 0.21 ± 0.20 | 0 | 0 | 0 | *** |
| 94 | Nonacosane | 26.54 ± 5.05 | 17.34 ± 3.08 | 24.90 | 42.99 ± 14.09 | 51.41 ± 13.58 | * |
| 95 | Triacontane | 5.72 ± 0.63 | 7.31 ± 3.07 | 2.79 | 0.51 ± 0.37 | 1.38 ± 0.68 | *** |
| 96 | Tentative: x,12-dimethylnonacosane | 0.30 ± 0.50 | 0 | 0 | 0.01 ± 0.02 | 0 | NS |
| 97 | Tentative: 2-methyltriacontane | 1.38 ± 0.43 | 0 | 0 | 0 | 0 | *** |
NS = not significant (p > 0.05), * p < 0.05, ** p < 0.01, *** p < 0.001.
Table 4.
Summary of the numbers of compounds identified in each Gonipterini species.
| Category | B. squamicollis | G. cinnamomeus | G. sp. n. 2 | O. fasciculatus | Oxyops sp. 1 |
|---|---|---|---|---|---|
| Number of identified compounds | 71 | 54 | 35 | 52 | 45 |
| Number of unique compounds | 20 | 2 | 2 | 7 | 4 |
| Percentage of unique compounds | 28.2% | 3.7% | 5.7% | 13.5% | 8.9% |
| Number of identified CHCs | 43 | 37 | 20 | 34 | 33 |
| Number of unique CHCs | 8 | 1 | 0 | 3 | 2 |
| Percent of unique CHCs | 18.6% | 2.7% | 0% | 8.8% | 6.1% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Johnson, J.B. Cuticular Hydrocarbon Profiling of Australian Gonipterini Weevils. AppliedChem 2023, 3, 414-427. https://0-doi-org.brum.beds.ac.uk/10.3390/appliedchem3030026
AMA Style
Johnson JB. Cuticular Hydrocarbon Profiling of Australian Gonipterini Weevils. AppliedChem. 2023; 3(3):414-427. https://0-doi-org.brum.beds.ac.uk/10.3390/appliedchem3030026
Chicago/Turabian StyleJohnson, Joel B. 2023. "Cuticular Hydrocarbon Profiling of Australian Gonipterini Weevils" AppliedChem 3, no. 3: 414-427. https://0-doi-org.brum.beds.ac.uk/10.3390/appliedchem3030026