
Reactive Oxygen Species Signaling Pathways: Arbiters of Evolutionary Conflict?


Department of Biological Sciences, Northern Illinois University, DeKalb, IL 60115, USA
Oxygen 2022, 2(3), 269-285; https://0-doi-org.brum.beds.ac.uk/10.3390/oxygen2030019
Submission received: 31 May 2022
/
Revised: 5 July 2022
/
Accepted: 14 July 2022
/
Published: 17 July 2022
(This article belongs to the Special Issue Reactive Oxygen Species (ROS))
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:In the history of life, cooperation between biological units has led to increased complexity, e.g., eukaryotic cells and multicellular organisms. Cooperation requires limiting the gains of “defectors” in favor of the cooperative higher-level unit. Early in an evolutionary transition, bioenergetics and reactive oxygen species (ROS) may play a large role in managing these evolutionary conflicts. Chemiosmosis can be thought of as a poorly insulated wire—when supply exceeds demand, electrons are cast off and can form ROS. ROS signaling may thus lead to the dispersal of the excess products into the environment. These products may lead to groups and the formation of higher-level units that can subsequently be targeted by selection. Examining modern symbioses such as those between corals and photosynthetic dinoflagellates provides useful insight in this context. While ROS are an important factor in coral bleaching, little is known of the function of ROS under other circumstances, although some data suggest that ROS may modulate cooperation. ROS may have functioned similarly in the origin of eukaryotes, involving chemiosmotic mitochondria and chloroplasts. ROS may act as “arbiters” of evolutionary conflict, leading to cooperation via signaling pathways that favor the emergence of the higher-level unit.
1. Introduction
In evolutionary biology, studies of cooperation have a lengthy and perhaps contentious history (e.g., [1,2,3]). While initially regarded as of less than general interest [4], as evolutionary biologists examined the history of life, cooperation was deemed to be more-and-more essential [2]. Remarkably, throughout this history, higher-level units (e.g., eukaryotic cells and multicellular organisms) emerged by the banding together of lower-level units (e.g., prokaryotic and eukaryotic cells) [5,6,7]. Parallels to these ancient events can be seen in a number of modern symbioses as well as simple multicellular organisms. Both symbiosis and multicellularity, whether leading to minor or major transitions, are fraught with evolutionary conflict, and these conflicts are a major obstacle for the higher-level unit to emerge.
Fundamental evolutionary conflicts can be described rather simply. As individuals form groups, some will contribute to group-level tasks (cooperators), while some will not (defectors) [8]. By shirking group-level contributions, defectors allocate more energy into their own replication and can outcompete cooperators. Mechanisms of conflict mediation have been credited with alleviating these evolutionary conflicts and preventing the competitive exclusion of cooperators [9,10]. Conflict mediation thus allows for the emergence of the higher-level unit (Figure 1).
Mechanisms of conflict mediation typically decrease the variation of lower-level units, making the evolution of defectors less likely, or increase the variation of the higher-level units, thus allowing selection to favor cooperative groups, or both. As described by Michod [9], in the context of the evolution of multicellularity:
“…we define a conflict mediator as a feature of the cell group that restricts the opportunity for fitness variation at the lower level (cells) and/or enhances the variation in fitness at the higher level (the cell group or organism). Accordingly, one can think of two general classes of conflict mediators: those that restrict within-group change and those that increase the variation in fitness between groups, although both have the effect of increasingly the heritability of fitness at the group level.”
In eukaryotes, numerous examples of such conflict mediation are apparent, e.g., the nearly complete loss of organellar genomes constraining variation at the lower level, and the omnipresence of sex increasing variation at the higher level. In multicellular eukaryotes, sex functions similarly, and lower-level variation is held in check by programmed cell death, among other mechanisms. Thus, such conflict mediators clearly have a role in major transitions such as the origin of eukaryotes and multicellularity, and parallels exist in more minor transitions. Nevertheless, the early stages of an evolutionary transition differ from later stages, and it remains an open question as to the extent that conflict mediators were present at the initiation of such transitions.
At the beginning of a major transition, only lower-level units are present within a larger population [11]. Evolutionary conflicts are likely to be rife, but there is no higher-level unit (except for the entire population) for selection to target. Of course, selection on a large population is likely to be much weaker than individual-level selection. More promising for the evolution of cooperation is a population structured into many, small groups. Thus, early in a transition the accepted definition of conflict mediation may not easily apply, e.g., since there is no higher-level unit that selection can effectively target, conflict mediation cannot increase the variation at this level. Further, constraining the variation of free-living, lower-level units may be more challenging than if these lower-level units were part of a higher-level unit that was subject to selection. While the term “individuation” has been suggested in this context [11], the wide usage of this term in other fields may promote misunderstandings. Hence, the more explicit term of “conflict management” is used here, based on the premise that conflicts at the lower level must be managed for a major transition to ensue [11]. Putative advantages of higher-level units are thus not relevant to these initial steps, since evolution cannot work toward a fitness advantage that is not immediately conferred.
When considered in this context, sub-group formation within a large population may be a crucial step in the evolution of cooperation. Small groups of individuals can facilitate kin selection and reciprocal altruism [12,13,14]. Groups also constitute a higher-level unit for selection to act on [15]. With sub-groups of individuals, selection can under some circumstances favor cooperation rather than competition. Group formation, for whatever reason, can automatically potentiate the emergence of cooperation.
In this regard, bioenergetics may have a central role. Chemiosmosis, which is integral to the bioenergetics of most cells and organisms, rapidly converts energy, and once storage capacity is exceeded, an overabundance of product has various negative consequences. Chemiosmosis can be thought of as a poorly insulated wire. When energetic inputs exceed metabolic demand, the wire does not melt, but it will cast off electrons, which typically form potentially detrimental reactive oxygen species (ROS), here considered to be partially reduced forms of oxygen. While to some extent chemiosmotic processes can be modulated, under some circumstances it may be advantageous to simply disperse the products into the environment. In biological systems in which energy is frequently limiting, such largesse will not go unnoticed—the free lunch will attract a crowd [16]. In other words, chemiosmosis may be a key factor that produces sub-groups of individuals within larger populations.
Dispersing excess products of chemiosmosis into the environment for purely selfish reasons can thus be the first step to sub-group formation and ultimately cooperation [16,17,18]. ROS may be crucial to managing evolutionary conflicts in favor of the higher-level unit. Strong selection favors individuals that allocate chemiosmotic products to maximize replication. Nevertheless, during favorable times chemiosmotic production may overwhelm a cell’s or an organism’s capacity to absorb and usefully allocate these products. At such times, signaling pathways activated by moderately excessive ROS formation can lead to a simple solution: dispersing these products into the environment before damage to the cell or organism occurs. The cost of emitting these products is assumed to be small compared to the cost of the potential damage [19]. The result is the “free lunch you are forced to make”. With the conflicts inherent in forming a symbiosis thus diminished, cooperation is favored. Nevertheless, ROS do not fit the usual definition of conflict mediators—increasing the variation at the higher level or constraining the variation at the lower level [9,10]. Rather, they would appear to act via more of a signaling role, e.g., when ROS reach a moderate to high level, they produce a signal to release end products into the environment, which can then lead to group formation. The signaling pathway, activated by ROS, thus produces an effect that leads a structured population. Further, such signaling likely evolved very early in the history of life, perhaps before LUCA, the last universal common ancestor. Indeed, ROS signaling may have complemented and supplanted signaling with reactive sulfur species, which may extend to the earliest forms of life prior to the oxygenation of the atmosphere [20,21]. Subsequently, and likely repeatedly in incipient symbioses, ROS signaling pathways may have been co-opted into a role in managing conflicts among lower-level units. To better elucidate the actions of ROS as “arbiters” of evolutionary conflict, the connections between ROS and chemiosmotic metabolism will be outlined, followed by the review of a modern example (the coral-dinoflagellate symbiosis) and an ancient one (the symbiosis that gave rise to eukaryotic cells) in which ROS signaling is hypothesized to have played a role. Finally, the topic of how evolutionary conflicts are managed very early in evolutionary transitions will be further discussed.
2. ROS as Indicators of Metabolic State
Chemiosmosis refers to a process of energy conversion used by virtually all living cells and organisms. Large, metal-containing protein complexes (the electron transport chain), embedded in a membrane that is impermeable to protons, pass electrons from one to another and in the process, extrude protons. These protons then return through the membrane via ATP synthase and trigger the formation of ATP. The electron transport chain in its entirety serves as a mechanism that chemiosmotic cells and organisms can use to sense their environment. This environmental sensing in turn adjusts a variety of cellular and organismal responses. Foremost in this regard, the electron transport chain links what are ultimately environmental sources and sinks of electrons (e.g., reduced carbon and oxygen). In mitochondria, for instance, reduced carbon is oxidized and coenzymes (e.g., NAD+ and FAD) are reduced. These coenzymes are then oxidized by components of the electron transport chain which in turn become reduced. As electrons are passed between complexes, this process of oxidation and reduction continues. Finally, at least in most mitochondria, the electrons are deposited on molecular oxygen, which is reduced to water. Essentially the same process occurs in chloroplasts, although beginning with removing electrons from oxygen (which requires light energy) and ending with depositing these electrons on carbon dioxide. Bioenergetic metabolism thus links external electron sources and sinks through a series of living redox couples [22].
This has been a common theme since before the last common ancestor of all life, and it highlights the crucial mechanism that organisms use to detect features of the environment. For instance, if the coenzymes are oxidized (e.g., NAD+, FAD, and NADP+), the organism is running out of substrate, and this will be indicated in various ways, e.g., the ATP/ADP ratio approaches zero. Successful organisms have quickly responded to such signals for thousands of millions of years (e.g., by feeding or initiating light capture). On the other hand, if most of the coenzymes remain reduced (e.g., NADH, FADH2, NADPH), various other responses may be indicated. The number of electron transport chains may be insufficient for the available food or light and transcription and translation may ensue. Alternatively, the terminal electron acceptor (e.g., oxygen, NADP+) might be scarce. Several organismal responses might ensue, involving behavior and locomotion (e.g., movement), physiology (e.g., respiration), or development (e.g., angiogenesis). In some cells, a switch to an alternative electron acceptor may be initiated. In the case of photosynthesis, end-product inhibition may be occurring, provoking a variety of responses. In these and many other circumstances, living things employ this sort of “redox” signaling as a rapid and effective mechanism to adjust bioenergetic metabolism to environmental conditions. Throughout the history of life, the “payoff” has been a larger dividend of energy. Since this dividend can be spent on a higher rate of replication, the Darwinian imperative is clear.
In eukaryotes, such redox signaling is often accomplished by intermediaries such as ROS (broadly defined here as partially reduced forms of molecular oxygen such as hydrogen peroxide and superoxide [19]). For example, if the environment becomes stressful and thus unfavorable for cellular replication and other energy-demanding activities, yet plenty of substrate remains available, cellular metabolic demand may diminish, and the ATP/ADP ratio may approach one. Nevertheless, oxidation of substrate may continue until the trans-membrane electrochemical gradient reaches a maximum, and the membrane-bound electron carriers become highly reduced. In such circumstances, these electrons carriers may donate electrons to molecules (e.g., molecular oxygen) whose partially reduced products can serve as messengers (e.g., ROS) [23,24,25,26]. These messengers can then trigger the appropriate adaptive response at the cellular level [27,28,29,30,31,32,33]. On the other hand, during starvation the ATP/ADP ratio may approach zero, and the trans-membrane electrochemical gradient may become minimal. The electron carriers may now be relatively oxidized and formation of, for example, ROS also becomes minimal. Again, appropriate responses can subsequently be triggered. Conveniently, ROS are ephemeral so their signaling can be rapidly tuned to changing environmental conditions.
In this context, note that cells have a number of specialized redox proteins to modulate signals generated with ROS (e.g., thioredoxins, glutaredoxins, and peroxiredoxins [34]). Energy metabolism itself can be adjusted to modulate the signaling microenvironment. For example, glycolysis can follow the canonical path to pyruvate or divert substrate in a number of ways, including most notably the pentose phosphate pathway, generating NADPH to counter oxidative stress [35]. The latter may have dramatic effects on organismal disease and longevity [36,37]. While the signaling pathways may have a variety of subtleties and complexities, it is the ROS that activate the pathways and lead to the outcomes.
Much early research on ROS focused on the toxicity of high concentrations of these molecules [38]. It has become increasingly clear, however, that ROS also have important functions in within- and between-cell signaling. It would nevertheless be misleading to imply that ROS are always harmless. As described by Brownlee [39]:
“Diabetes-specific microvascular disease is a leading cause of blindness, renal failure and nerve damage, and diabetes-accelerated atherosclerosis leads to increased risk of myocardial infarction, stroke and limb amputation. Four main molecular mechanisms have been implicated in glucose-mediated vascular damage. All seem to reflect a single hyperglycaemia-induced process of overproduction of superoxide by the mitochondrial electron-transport chain. This integrating paradigm provides a new conceptual framework for future research and drug discovery.”
Under most physiological conditions the signal encoded in ROS is read and reacted to before significant damage can occur. It is in this context that ROS signaling pathways may function as arbiters of evolutionary conflict. This hypothetical relationship is elucidated by the examples that follow.
3. Corals and Dinoflagellates: ROS Signaling in a Modern Symbiosis
Numerous marine animals form symbiotic associations, particularly clonal and colonial representatives of taxa such as sponges, ascidians, bryozoans, and cnidarians [40,41,42,43]. Remarkably, all modern reef-building cnidarians contain endosymbiotic dinoflagellates [44], formerly referred to as Symbiodinium and now classified as the family Symbiodiniaceae [45]. Many other colonial cnidarians, whether part of coral reef communities or not, also exhibit symbioses with various forms of Symbiodiniaceae. The coral-dinoflagellate symbiosis has attracted considerable attention because its breakdown seems to be integral to coral bleaching. When environmental stress (e.g., light and heat) becomes extreme, corals bleach, thus indicating that these dinoflagellates have been lost [46]. As elaborated below, ROS signaling likely alleviates evolutionary conflicts, but also contributes to the process of coral bleaching in which the cooperative symbiosis breaks down.
While taxa included in the Symbiodiniaceae form symbioses with corals and many other metazoans, they also remain capable of free-living existence. Given the intense competition for space in marine benthic communities, symbiosis appears to be a path by which Symbiodiniaceae can become larger and thus more effective competitors. In parallel, size increase also seems to have been central to the origin of eukaryotes and multicellularity (18). For the metazoan host of Symbiodiniaceae, the symbiosis provides at least partial autotrophy, since these dinoflagellates are photosynthetic and actively export various forms of reduced carbon to the host [46].
Despite these mutual benefits, robust mechanisms of conflict mediation are needed to produce a durable symbiosis and to hold defectors in check. For instance, a structured population consisting of many, small groups can mediate conflict. Even if defectors are strongly selected at the individual level, with many, small groups, purely by chance (i.e., genetic drift) some groups will comprise only cooperators. At the higher level, these groups of cooperators will be strongly selected for and outcompete groups that include defectors. This sort of scenario likely contributed to numerous evolutionary transitions, including the secondary symbioses that gave rise to dinoflagellates [11]. While some stages of the life cycle of colonial cnidarians may comprise many small groups (e.g., when small, sexually produced colonies first take up symbionts [47]), overall populations of colonial cnidarians are structured entirely unfavorably in this respect. Indeed, these populations can be characterized as relatively few very large and very long-lasting groups. In other words, cnidarian colonies are large, long-lived, and relatively scarce, and a single colony contains many trillions of symbionts. Within a colony, under these conditions defecting symbionts are strongly selected for. A stable symbiosis cannot be achieved without additional mechanisms to mediate evolutionary conflicts.
Housing a small number of symbionts in each of many host cells is a way to create many small groups within a single colony. Maintaining symbionts in small groups within cells potentiates other mechanisms of conflict mediation as elaborated below (Figure 2). Crucially, replication of symbionts is limited by the host, likely via limits on provisioning with inorganic nutrients [46]. Limited replication of symbionts in turn limits metabolic demand. Under these conditions, the biophysics of chemiosmosis dictate that excess product must be exported by the symbiont, i.e., shared with the higher-level unit. Defecting symbionts, with the transporter pathways inactivated by mutation, will shift their redox state in the direction of reduction because of end-product inhibition or other mechanisms [16,17,18,47]. High levels of ROS will then form likely triggering cell death. As noted above, programmed cell death plays an important role in several evolutionary transitions [48,49]. In a cooperating symbiont, on the other hand, these ROS may trigger the export of end products before they reach levels that impact viability. An ROS signaling pathway that triggers cell death clearly constrains the variation at the lower level. On the other hand, a pathway that leads to photosynthate export does not do this.
Features of the coral-dinoflagellate natural history that otherwise are difficult to explain can be better illuminated by this hypothesis. On sunny days, symbiotic corals release large quantities of reduced carbon [50,51]. This material includes lipids and polysaccharides, which are released in the form of “mucus”. Many other organisms utilize this material as substrate [47]. Coral workers have struggled to explain how this is adaptive. As suggested above, it may simply be a mechanism to disperse excess product into the environment. Indeed, as described by Crossland et al. [50]: “Lipid production may provide an alternative to zooxanthellar photorespiratory processes…in utilizing excess photosynthetically produced ATP and reducing power [NAD(P)H]…A variety of mechanisms for dispersal of reducing power may be an important feature in maintaining chloroplastic integrity of zooxanthellae contained by the sessile coral in high light environments (e.g., reef flats, shallow reefs)”. Using terminology that was commonplace at that time, Crossland et al. [50] refer to the Symbiodiniaceae as zooxanthellae.
Generally, oxygenic photosynthesis is a risky process, involving the separation of hydrogen atoms into electrons and protons, often in the presence of high concentrations of molecular oxygen. These risks become particularly acute if there is an excess of products, resulting in a backup of electrons and highly reduced electron carriers [52]. Production of ROS can be enhanced under these conditions [53], and ROS may lead to programmed cell death [54,55]. If the rate of CO2 fixation slows with the accumulation of photosynthate, while water splitting proceeds apace, then electrons will back up on photosystems I and II and other electron carriers. ROS formation will consequently increase, and individual symbionts risk serious damage and death. Such end-product inhibition is widely discussed in the literature of plant photosynthesis [56,57,58] and may be relevant to corals and their symbionts as Crossland et al. [50] suggest. Symbionts must export photosynthate for their own survival since their growth and replication remain limited. Rather than reaping benefits as defectors, those symbionts that have lost by mutation the capacity to translocate photosynthate may instead precipitate their own demise. Adaptations of corals to expose symbionts to optimal light levels for photosynthesis [59,60] may thus have a dual function. Not only does symbiont photosynthesis occur at a rapid pace, but also symbionts that fail to share the bounty succumb even more quickly. Note that the effectiveness of end-product inhibition is inversely proportional to the replication rate of the symbionts. In other words, rapid symbiont replication if allowed to proceed could potentially consume all available photosynthate [47,61].
While ROS are considered central to coral bleaching, most discussions of ROS in this context focus on damage and death. As the larger field has matured, so too must discussions of coral bleaching. In other words, the possibility that ROS may have a signaling role beyond triggering cell death, i.e., possibly modulating the export of photosynthate or other processes, should be considered. Little is known in this regard, likely for two reasons. First, beyond damage and death there has been little interest in exploring a signaling role for ROS in corals. Second, slow-growing, calcareous hexacorals may be poor laboratory models to study this signaling. Octocorals, which comprise fast-growing, “soft” corals may have a useful role in this context. For instance, recently the role of ROS in triggering symbiont migration in stressed octocorals was examined [62]. While both migration and ROS increase rapidly when colonies were stressed, a dose–response relationship remained elusive. In part, this may be because of the very different temporal dynamics of the two phenomena. ROS are ephemeral, but once begun, symbiont migration may continue for hours.
Other discussions of bleaching have taken a somewhat different perspective, centering on carbon-concentrating mechanisms (CCM’s). Here, the biochemical substrate for the products of photosynthesis is the focus. Carbon-concentrating mechanisms in corals serve to take up HCO3− from seawater and deliver it as CO2 to symbiont-containing host cells [63,64,65]. Plausibly, individual symbiont-containing host cells rely on energy derived from the oxidation of photosynthates to power their CCM’s [63,64]. Within a host cell, symbiont-produced photosynthate is transported to host coral mitochondria where it is oxidized to form ATP, thus powering the host CCM’s that are responsible for providing symbionts with an adequate supply of CO2. As Wooldridge [63,64] suggests, a defecting symbiont that has mutationally lost the ability to translocate photosynthates will quickly impair the ATP supply to its associated host CCM’s. The defecting symbiont will thus limit its own CO2 supply, with the same detrimental consequences as end-product inhibition. The effects of collapse of CCM’s does require that the symbiont reside inside a host cell that has functional mitochondria and transporters and not directly contact seawater, as Symbiodiniaceae have bicarbonate transporters of their own [66,67]. ROS may have an important role in this context. A shortage of CO2 would lead to high ATP/ADP and NADPH/NADP+ ratios. These in turn would lead to a backup of electrons on the photosystems and other electron carriers, resulting in high levels of ROS and cell death.
While other mechanisms of conflict mediation may have a role in maintaining the coral-dinoflagellate symbioses [47], the ones that have been co-opted from features of photosynthesis connect most directly to ROS signaling. These mechanisms are particularly effective because they require only the presence of light to distinguish between cooperating and defecting symbionts. As pointed out above, symbiont replication is suppressed within the coral. Some replication nevertheless occurs. Replication inevitably produces mutational variation [7,68], and typically loss-of-function variants will be the most common. Many loss-of-function variants will be uniformly selected against. On the other hand, those that inactivate mechanisms of photosynthate translocation may be favored at the symbiont level. Given the constraints on replication, these defecting symbionts may only gain a slight replicatory advantage. Over many symbiont generations in a long-lived coral, however, even a small advantage in replication rate can translate into large numerical gains. Meanwhile, the multiplication of these defectors represents an existential threat to the higher-level community, i.e., the cooperative symbionts and the host. How can these defectors be controlled?
As defectors fail to export photosynthate, both product inhibition and the collapse of CCM’s result in the back up of electrons onto photosystems I and II and other electron carriers. As the metabolic state of the electron carriers shifts in the direction of reduction, electrons are more likely to be cast off. Molecular oxygen is particularly likely to gain these electrons, forming various ROS. The well-known “Mehler reactions” involve ROS that are formed by photosystem I and ultimately converted by antioxidants into fully reduced oxygen, i.e., water [54,55]. However, if ROS are produced above the amounts that can be reduced by antioxidants, they can trigger damage and programmed cell death [54,55,69,70]. Under conditions in which symbionts carry out photosynthesis, a fairly constant background level of cell death likely eliminates those host cells containing defectors (Figure 2). The biophysics of photosynthesis thus functions to mediate conflict in favor of the higher-level community.
Under stressful environmental conditions, however, the calculus changes somewhat. In the presence of excess heat and light normal processes of symbiont photosynthesis fail, and many more symbionts become highly reduced and emit high levels of ROS, initiating bleaching [52,53,54]. At this stage, end-product inhibition is highly unlikely, although CCM collapse may occur [63,64]. Alternatively, bleaching may initiate with other mechanisms that trigger reduction in electron carriers, casting off of electrons, and ROS. For example, RuBisCO or other Calvin Cycle enzymes may be involved in the initiating steps in bleaching [71,72]. A diversity of initiating mechanisms may be found, as suggested by studies of different coral species and symbiont taxa [73]. Nevertheless, as long as the initiating mechanisms trigger reduction in electron carriers and ROS, cooperators will not be distinguishable from defectors, and higher levels of programmed cell death may ensue (Figure 3). During these times, corals such as Galaxea fascicularis and other scleractinians may expel intact symbionts, suggesting that many cooperators may thus be destroyed [74,75]. In these taxa under more favorable conditions, expelled symbionts were usually functionally impaired. This, however, was not the case under bleaching conditions.
Several testable predictions follow from this hypothesis. For example, do stress-resistant symbionts (Figure 3A) emit more ROS when colonies are not stressed? A recent study [62] shows that unstressed colonies of Sarcothelia sp. produce greater amounts of ROS than unstressed colonies of Sympodium sp. Both species are xeniid octocorals belonging to the Holaxonia-Alcyoniina clade [76]. However, the symbiont types of these species differ in their stress resistance. Symbionts of Durusdinium sp. (found in Sarcothelia sp.) seem to be more resistant to stress, while those of Cladocopium sp. (found in Sympodium sp.) are more productive under stress-free circumstances [45]. Less cooperative symbionts of Durusdinium sp. are hypothesized to release less reduced carbon and produce more ROS. Colonies of Sarcothelia sp. may have mechanisms to cope with these ROS. More cooperative Cladocopium sp. are hypothesized to release more reduced carbon and produce less ROS. Colonies of Sympodium sp. may thus have fewer antioxidant resources to cope with ROS produced during the stress response. Colonies that are tolerant of ROS during bleaching may have been conditioned to them under normal circumstances.
While formulated in terms of non-photosynthetic eukaryotic cells, the concept of physiological oxidative stress or oxidative eustress [77] is highly relevant here. Depending on the symbiont type hosted, the steady-state redox set point of a coral colony can vary greatly (Figure 3). More cooperative symbionts allow a less reduced set point, while less cooperative symbionts require a more reduced set point. The consequences of this difference extend to the pathophysiology of coral bleaching, in which the less cooperative symbionts paradoxically prove beneficial.
4. Eukaryogenesis: The Role of ROS in a Major Transition
The most challenging of all the major evolutionary transitions appears to be the origin of eukaryotes [78]. Certainly, all evidence suggests a long period in which life on earth was dominated by prokaryotes. What were the obstacles that hindered this transition for 2 billion years? While we may never completely comprehend the answers to this question, there can be no doubt that evolutionary conflict was a serious hurdle. Metabolism in general and chemiosmosis in particular likely had crucial roles in mediating these conflicts.
The advantages of eukaryotic cells are straightforward. Not only are they more complex, but they are also larger than their prokaryotic forebearers. As Bonner [79] points out, “…the reason for non-stop selection for organisms of increased size is that the top of the size scale is an ever-present open niche and has been open during the entire course of organic evolution”. Larger size provides a number of ecological advantages including the exploitation of more and different food resources, more efficient dispersal (e.g., escaping the constraints of low Reynolds numbers), producing more offspring, and escaping predators [80]. Prokaryotes, however, face a conundrum that greatly limits their options for size increase: their energy-converting complexes are found on the cell membrane [81]. If a prokaryotic cell gets larger, there is less surface to convert energy and more volume requiring energy conversion. Despite some exotic exceptions, by-and-large prokaryotes never transcended these surface-to-volume constraints.
One way to circumvent these constraints is to move small energy-converting cells inside a larger complex cell, thus freeing the external membrane from duties related to energy conversion [81,82]. This arrangement allows for the complex cell to increase in size. From this perspective, endosymbiosis is integral to the evolution of eukaryotes [83]. A clever engineering solution for surface-to-volume constraints, however, results in a levels-of-selection nightmare.
This can be better understood by recognizing the fundamental duality of life. All life takes up energy from the environment and typically converts this energy into more useful forms. At the same time, life involves information and replication. There remains a constant tension between these two attributes in all forms of life. In a structured population, a biological unit that relies on its sister units for energy conversion and specializes in replication may be favored. The success of such a unit, however, is frequency dependent [7]. As the proportion of units specializing in replication increases within a group, the availability of the products of energy conversion inexorably decreases. At some point, the lack of these products threatens the entire community. For the community to persist, mechanisms of conflict mediation must evolve.
While these issues prevail throughout the entire history of life, they were particularly acute in the origin of the eukaryotic cell. Two great symbioses—that of the mitochondrion and the plastid—figure prominently in the rise of eukaryotes. Both of these symbioses involve energy-converting lower-level units, with groups of these lower-level units and host cells constituting the higher-level units. In both cases, evolutionary conflict no doubt involved the usual pattern of lower-level units specializing in replication at the expense of distributing the products of energy conversion to the higher-level community.
Mitochondria may have been attendant to the very early stages in eukaryotic origins and were very likely a feature of the last eukaryotic common ancestor (LECA) [84]. Plastids were acquired by some eukaryotes not long after LECA [85,86]. The origin of eukaryotes continues to be one of the most debated topics in all of biology. Considerations have turned to the nature of the original host and metabolic relationships that may have led to the endosymbiosis, topics that continue to attract considerable attention and sometimes sharp disagreements [87,88,89,90,91,92].
In modern eukaryotes, evolutionary conflict is mediated by a number of mechanisms (e.g., the nearly complete loss of organellar genomes). Nevertheless, the evolution of many of these mechanisms required strong selection on the higher-level units. Such selection cannot have molded the initial steps in the symbiosis, which by definition involved only lower-level units. Notably, mitochondria, chloroplasts, and their bacterial relatives convert energy using chemiosmosis, which involves electron flow and proton extrusion as described above.
The discovery of quantum electron transfer in biological systems [93] occurred contemporaneously with the development the chemiosmotic theory [94]. Later work showed that membrane-bound electron carriers could form “super-complexes.” Both quantum electron transfer and super-complex formation result in extremely rapid electron transfer in chemiosmotic processes [95,96]. When chemiosmosis is linked to the soluble reactions that store energy, this rapidity poses problems. For example, the ATP and NADPH produced by chemiosmosis in chloroplasts is stored by RuBisCO and other Calvin Cycle enzymes by reducing carbon dioxide. RuBisCO is thus the most abundant protein on Earth. The channeling of chemiosmosis into slower soluble reactions with potentially limited storage capacity has other effects. As pointed out above, if products cannot be consumed or stored and their accumulation inhibits electron flow, electron carriers become highly reduced, and these electrons may divert to molecular oxygen forming ROS. The chemiosmotic process itself, by separating protons and electrons, is the cause of ROS formation.
Cooperation is not an automatic outcome of evolutionary interactions because it often involves costs. Selection may thus favor incompatible features of hosts and symbionts [97]. A symbiont may be selected to defect, e.g., by hoarding resources obtained from the host and symbiont community and using these resources for its own replication. While cooperative symbionts at least in part forgo reproduction and share resources with the larger community, defectors may gain a replicatory advantage by sequestering these resources. Yet, by sharing resources with the host, the cooperative symbiont community may establish a durable environment and insure its long-term persistence. Even with these long-term advantages, the higher-level unit (the host and the larger symbiont community) may be exploited by lower-level defectors. While symbiosis can be classified as mutualistic or parasitic (i.e., in bilateral terms), mechanistically these evolutionary interactions are multilateral and multilevel. Defecting symbionts can still arise and flourish even when a host-symbiont community appears to be dominated by mutualistic interactions. Mechanisms of conflict mediation are always necessary to control lower-level defectors. A structured population may mediate conflicts, e.g., if a population is subdivided into many small groups. These groups can lead to kin selection and reciprocity. Further, groups of cooperators can arise purely by chance. In this way, even if cooperation is selected against at the level of the individual, it can still arise and be favored at the level of the group [11].
Remarkably, ROS signaling pathways may serve as arbiters of evolutionary conflict. Chemiosmosis proceeds rapidly and conserves a large proportion of the energetic input, quickly generating products. The products of chemiosmosis may be stored, but storage mechanisms can be slow relative to chemiosmosis. Additionally, storage capacity is not unlimited. When conditions are favorable, chemiosmotic cells and organisms are often confronted with “end-product inhibition” [56,57,58,98], which can have detrimental consequences. As discussed above, an overabundance of product can inhibit electron flow. In the presence of molecular oxygen, this enhances the formation of ROS. Such partially reduced forms of oxygen can have a variety of deleterious effects. To avoid blocking electron flow, the abundant products of chemiosmotic energy conversion must be consumed, stored, or simply gotten rid of. While mechanisms that modulate chemiosmosis are available [99,100], alternatively the products of chemiosmosis can be dispersed into the environment. Because ROS are ephemeral, they are particularly well suited for this role. With the onset of end-product inhibition, ROS would increase, possibly signaling the initiation of the export of chemiosmotic products. As these products were exported, end-product inhibition would lessen, and ROS would rapidly subside.
Crucially, chemiosmosis and its consequential ROS formation can thus under some circumstances favor sharing rather than hoarding the products of energy conversion. Sharing can lead to the formation of groups, whether symbiotic associations or multicellular aggregations. These can be thought of as a by-product of sharing, which in turn is a by-product of chemiosmosis. Thus formed, these groups may be the key to the evolution of cooperation, potentiating kin selection, reciprocity, or group selection. Indeed, the biophysical constraints of chemiosmosis may have provided the initial mediation of conflicts associated with origins of eukaryotic cells and thus allowed the first steps in the formation of complex life on Earth.
Mechanisms that mediate evolutionary conflict typically constrain the variation at the lower level, so that defectors are less likely to evolve, or increase the variation at the higher level, so that cooperative groups are favored by selection [9,10]. ROS signaling pathways may or may not fit this paradigm. If a pathway leads to cell death, it constrains variation at the lower level. On the other hand, if a pathway leads to nutrient export and group formation, this clearly does not constrain lower-level variation. By leading to group formation, it may, however, be viewed as increasing the variation at the higher level. Depending on which pathway(s) are activated, ROS could lead to one or the other. Hence, as further discussed below, ROS are termed “arbiters” of evolutionary conflict to better represent their putative role in initiating a signaling process that resolves conflict and leads to the emergence of a higher-level unit.
As endosymbioses became established and higher-level units emerged, energy-converting lower-level units remained an obstacle to cooperation. Numerous additional mechanisms of conflict mediation subsequently evolved, facilitated by the large populations of higher-level units, which each contained relatively small populations of lower-level units. Purely on the basis of chance, some higher-level units could be formed from cooperative lower-level units. These groups of cooperators could then outcompete cells that included one or many defectors. Nevertheless, under conditions that lead to end-product inhibition, defectors that arise via loss-of-function mutations could still be eliminated from cooperative groups. In this way, unicellular eukaryotes developed robust and stable symbioses.
The biophysics of chemiosmosis, particularly the formation of ROS as a by-product, were likely central to the initial steps of eukaryogenesis, when only lower-level selection was operating. Both quantum electron transfer and super-complex formation contribute to high rates of chemiosmotic processes. Large quantities of products may thus be produced. While several mechanisms can modulate chemiosmosis, alternatively excess product can be released, thus providing a way to protect against an accumulation of end-products and the consequent formation of dangerous by-products such as ROS. Sharing may thus occur with little cost and considerable benefit—"the free lunch you are forced to make” [16]—thus facilitating groups. Such groups can lead to cooperative symbioses. Even after such groups have formed, however, cooperators are always vulnerable to exploitation by defectors, so additional mechanisms of conflict mediation are usually necessary.
While there are examples of prokaryotic aggregations (e.g., biofilms), these pale in comparison to eukaryotic multicellularity (e.g., Conway Morris [101]: “The history of life shouts ‘Look! Once there was bacteria, now there is New York’”). Eukaryotes seems particularly able to form complex, multicellular organisms. Possibly, the mechanisms of cooperation established by the eukaryotic cell may have been repeatedly co-opted during the emergence of eukaryotic multicellularity [102]. In multicellular organisms, the result may be fundamental relationships between nutrient supply, metabolism, and cooperation that are only now beginning to come into focus [16,17,18].
5. “Mediators” and “Arbiters” of Evolutionary Conflict
As suggested in the introduction, at the beginning of a symbiosis or a major transition, only lower-level units are present in an unstructured population [11]. Evolutionary conflicts are likely to be rife, but there is no higher-level unit that selection can effectively target. Thus, the accepted definition of conflict mediation may not easily apply, e.g., since there is no higher-level unit, conflict mediation cannot increase the variation at this level. Further, constraining the variation of free-living, lower-level units may be more challenging than if these lower-level units were part of a higher-level unit. Putative advantages of higher-level units are thus not relevant to these initial steps, since evolution cannot work toward a fitness advantage that is not immediately conferred. Evolutionary conflicts, nevertheless, remain.
In such circumstances, conflicts may overwhelm incipient cooperation, and the association of lower-level units may dissolve. If these conflicts are to be managed, an existing mechanism must be co-opted into a new role related to evolutionary conflict. Such a mechanism is expected to have evolved prior to the initiation of the transition and to function in an entirely different context.
In this regard, bioenergetics may have a role (Figure 4). Chemiosmosis, which is integral to the bioenergetics of most cells and organisms, remains a potentially fraught process. It rapidly converts energy, but once storage capacity is exceeded, an overabundance of product can have various negative consequences. In a sense, chemiosmosis behaves like a poorly insulated wire. When supply exceeds demand, it will cast off electrons, which typically form potentially detrimental ROS. Mechanisms have evolved to modulate chemiosmotic processes [99,100]. On the other hand, under certain circumstances it is also possible to leave these processes more-or-less intact and simply disperse the products into the environment. Chemiosmosis and its regulation (or lack thereof) evolved purely on the basis of lower-level selection and unrelated to an evolutionary transition.
Nevertheless, dispersing excess products of chemiosmosis into the environment for purely selfish reasons can be the first step to group formation and ultimately cooperation [16,17,18]. ROS signaling pathways may be crucial to managing evolutionary conflicts in favor of the higher-level unit (Figure 4). Strong selection favors individuals that allocate chemiosmotic products to maximize replication. Nevertheless, during favorable times chemiosmotic production may overwhelm a cell’s or an organism’s capacity to absorb and usefully allocate these products. At such times, signaling from moderately excessive ROS formation can lead to a simple solution: dispersing these products into the environment before damage to the cell or organism occurs. The cost of emitting these products is assumed to be small compared to the cost of the potential damage [19]. The result is the “free lunch you are forced to make” [16]. With the conflicts inherent in forming a symbiosis thus diminished, cooperation is favored. ROS signaling early in an evolutionary transition thus can lead to sharing and sharing can lead to groups. In this sense, this hypothetical ROS signaling arbitrates conflict by creating the higher-level unit that can then function as a target of selection. These initial steps occur largely as a by-product of individual-level selection. As suggested by Figure 4, in a symbiosis or major transition, the initial steps that lead to cooperative groups may differ substantially from later mechanisms of conflict mediation.
As pointed out above, however, the role of ROS signaling is complex. High levels of ROS can activate pathways that lead to programmed cell death. Such a result fits easily into the framework of conflict mediation, since it constrains the variation at the lower level. More moderate levels of ROS, on the other hand, may trigger pathways that lead to the export of end products, thus facilitating the formation of cooperative sub-groups and a higher-level unit. While this may appear to be outside the framework of conflict mediation, in a broader sense it may indeed fit within this framework. Creation of a higher-level unit could be viewed as the first step to increasing the variation at the higher level. Nevertheless, creation of higher-level units differs from increasing the variation in a population of established higher-level units. More considerations of the early stages of evolutionary transitions should be illuminating.
Funding
This research received no external funding.
Acknowledgments
Two reviewers provided very helpful comments and suggestions on an earlier draft of this manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- Frank, S.A. Repression of competition and the evolution of cooperation. Evolution 2003, 57, 693–705. [Google Scholar] [CrossRef]
- Okasha, S. Evolution and the Levels of Selection; Oxford University Press: Oxford, UK, 2006; pp. 1–263. [Google Scholar]
- Leigh, E.G., Jr. The group selection controversy. J. Evol. Biol. 2010, 23, 6–19. [Google Scholar] [CrossRef] [PubMed]
- Michod, R.E.; Herron, M.D. Cooperation and conflict during evolutionary transitions in individuality. J. Evol. Biol. 2006, 19, 1406–1409. [Google Scholar] [CrossRef] [PubMed]
- Buss, L.W. The Evolution of Individuality; Princeton University Press: Princeton, NJ, USA, 1987. [Google Scholar]
- Maynard Smith, J.; Szathmáry, E. The Major Transitions in Evolution; Oxford University Press: Oxford, UK, 1995; pp. 1–346. [Google Scholar]
- Michod, R.E. Darwinian Dynamics: Evolutionary Transitions in Fitness and Individuality; Princeton University Press: Princeton, NJ, USA, 1999. [Google Scholar]
- Blackstone, N.W. An Evolutionary Framework for Understanding the Origin of Eukaryotes. Biology 2016, 5, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michod, R.E. Cooperation and conflict mediation during the origin of multicellularity. In Genetic and Cultural Evolution of Cooperation; Hammerstein, P., Ed.; MIT Press: Cambridge, UK, 2003; pp. 291–307. [Google Scholar]
- Michod, R.E.; Nedelcu, A.M. Cooperation and conflict during the unicellular-multicellular and prokaryotic-eukaryotic transitions. In Evolution: From Molecules to Ecosystems; Moya, A., Font, E., Eds.; Oxford University Press: Oxford, UK, 2004; pp. 195–208. [Google Scholar]
- Radzvilavicius, A.L.; Blackstone, N.W. The evolution of individuality, revisited. Biol. Rev. 2018, 93, 1620–1633. [Google Scholar] [CrossRef]
- Hamilton, W.D. The genetical evolution of social behaviour. I. J. Theor. Biol. 1964, 7, 1–16. [Google Scholar] [CrossRef]
- Hamilton, W.D. The genetical evolution of social behaviour. II. J. Theor. Biol. 1964, 7, 17–52. [Google Scholar] [CrossRef]
- Trivers, R.L. The Evolution of Reciprocal Altruism. Q. Rev. Biol. 1971, 46, 35–57. [Google Scholar] [CrossRef]
- Wilson, D.S. A theory of group selection. Proc. Natl. Acad. Sci. USA 1975, 72, 143–146. [Google Scholar] [CrossRef] [Green Version]
- Blackstone, N.W. Chemiosmosis, evolutionary conflict, and eukaryotic symbiosis. In Symbiosis: Cellular, Molecular, Medical, and Evolutionary Aspects; Kloc, M., Ed.; Springer Nature: Cham, Switzerland, 2020; pp. 237–252. [Google Scholar]
- Blackstone, N.W.; Gutterman, J.U. Can natural selection and druggable targets synergize? Of nutrient scarcity, cancer, and the evolution of cooperation. BioEssays 2021, 43, 2000160. [Google Scholar] [CrossRef]
- Blackstone, N.W. Energy and Evolutionary Conflict: The Metabolic Roots of Cooperation; Springer Nature: Cham, Switzerland, 2022. [Google Scholar]
- Juan, C.; de la Lastra, J.P.; Plou, F.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef] [PubMed]
- Olson, K.R. Hydrogen sulfide, reactive sulfur species and coping with reactive oxygen species. Free Radic. Biol. Med. 2019, 140, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Olson, K.R. The biological legacy of sulfur: A roadmap to the future. Comp. Biochem. Physiol. Part A 2020, 252, 110824. [Google Scholar] [CrossRef] [PubMed]
- Allen, J. Control of Gene Expression by Redox Potential and the Requirement for Chloroplast and Mitochondrial Genomes. J. Theor. Biol. 1993, 165, 609–631. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, J.S.; Whiteman, M.; Yang, H.; Jones, D.P. The redox regulation of intermediary metabolism by a superox-ide-aconitase rheostat. BioEssays 2004, 26, 895–900. [Google Scholar] [CrossRef]
- Salmeen, A.; Andersen, J.N.; Meyers, M.P.; Meng, T.-C.; Hinks, J.A.; Tonks, N.K.; Barford, D. Redox regulation of protein tyrosine phosphatase 1B involves a suphenyl-amide intermediate. Nature 2003, 423, 769–773. [Google Scholar] [CrossRef]
- Van Montfort, R.L.M.; Congreve, M.; Tisi, D.; Carr, R.; Jhoti, H. Oxidation state of the active-site cysteine in protein tyrosine phosphatase 1B. Nature 2003, 423, 773–777. [Google Scholar] [CrossRef]
- Filomeni, G.; Rotilio, G.; Ciriolo, M.R. Disulfide relays and phosphorylation cascades: Partners in redox-mediated signaling pathways. Cell Death Differ. 2005, 12, 1555–1563. [Google Scholar] [CrossRef] [Green Version]
- Coloff, J.L.; Rathmell, J.C. Metabolic regulation of Akt: Roles reversed. J. Cell Biol. 2006, 175, 945–947. [Google Scholar] [CrossRef] [Green Version]
- Kondoh, H.; Lleonart, M.E.; Bernard, D.; Gil, J. Protection from oxidative stress by enhanced glycolysis; a possible mechanism of cellular immortalization. Histol. Histopathol. 2007, 22, 85–90. [Google Scholar]
- Ladurner, A.G. Rheostat Control of Gene Expression by Metabolites. Mol. Cell 2006, 24, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Coffman, J.A.; Davidson, E.H. Oral-aboral Axis Specification in the Sea Urchin Embryo. Dev. Biol. 2001, 230, 18–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coffman, J.A.; McCarthy, J.J.; Dickey-Sims, C.; Robertson, A.J. Oral-aboral axis specification in the sea urchin embryo: II. Mitochondrial distribution and redox state contribute to establishing polarity in Strongylocentrotus purpuratus. Dev. Biol. 2004, 273, 160–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fomenko, D.E.; Xing, W.; Adair, B.M.; Thomas, D.J.; Gladyshev, V.N. High-Throughput Identification of Catalytic Redox-Active Cysteine Residues. Science 2007, 315, 387–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkel, T. Reactive Oxygen Species and Signal Transduction. IUBMB Life 2001, 52, 3–6. [Google Scholar] [CrossRef]
- Hanschmann, E.-M.; Godoy, J.R.; Berndt, C.; Hudemann, C.; Lillig, C.H. Thioredoxins, Glutaredoxins, and Peroxiredox-ins—Molecular Mechanisms and Health Significance: From Cofactors to Antioxidants to Redox Signaling. Antioxid. Redox Signal. 2013, 19, 1539–1605. [Google Scholar] [CrossRef]
- Kuehne, A.; Emmert, H.; Soehle, J.; Winnefeld, M.; Fischer, F.; Wenck, H.; Gallinat, S.; Terstegen, L.; Lucius, R.; Hildebrand, J.; et al. Acute Activation of Oxidative Pentose Phosphate Pathway as First-Line Response to Oxidative Stress in Human Skin Cells. Mol. Cell 2015, 59, 359–371. [Google Scholar] [CrossRef] [Green Version]
- Legan, S.K.; Rebrin, I.; Mockett, R.J.; Radyuk, S.; Klichko, V.I.; Sohal, R.S.; Orr, W.C. Overexpression of Glucose-6-phosphate Dehydrogenase Extends the Life Span of Drosophila melanogaster. J. Biol. Chem. 2008, 283, 32492–32499. [Google Scholar] [CrossRef] [Green Version]
- Nobrega-Pereira, S.; Fernandez-Marcos, P.J.; Brioche, T.; Gomez-Cabrera, M.C.; Salvador-Pascual, A.; Flores, J.M.; Vina, J.; Serrano, M. G6PD protects from oxidative damage and improves healthspan in mice. Nat. Commun. 2016, 7, 10894. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, D.L. Fifty years of radical ideas. In Reactive Oxygen Species: From Radiation to Molecular Biology; Chiueh, C.C., Ed.; New York Academy of Sciences: New York, NY, USA, 2000; Annals 899; pp. 1–14. [Google Scholar]
- Brownlee, M. Biochemistry and molecular cell biology of diabetic complications. Nature 2001, 414, 813–819. [Google Scholar] [CrossRef]
- Wulff, J.L. Clonal organisms and the evolution of mutualism. In Population Biology and Evolution of Clonal Organisms; Jackson, J.B.C., Buss, L.W., Cook, R.E., Eds.; Yale University Press: New Haven, CT, USA, 1985; pp. 437–466. [Google Scholar]
- Burgess, S.C.; Ryan, W.H.; Blackstone, N.W.; Edmunds, P.J.; Hoogenboom, M.O.; Levitan, D.R.; Wulff, J.L. Metabolic scaling in modular animals. Invertebr. Biol. 2017, 136, 456–472. [Google Scholar] [CrossRef] [Green Version]
- Bang, C.; Dagan, T.; Deines, P.; Dubilier, N.; Duschl, W.J.; Fraune, S.; Hentschel, U.; Hirt, H.; Hülter, N.F.; Lachnit, T.; et al. Metaorganisms in extreme environments: Do microbes play a role in organismal adaptation? Zoology 2018, 127, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Blackstone, N.W. Evolutionary conflict and coloniality in animals. J. Exp. Zool. Part B Mol. Dev. Evol. 2020, 336, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Hoegh-Guldberg, O. Climate change, coral bleaching and the future of the world’s coral reefs. Mar. Freshw. Res. 1999, 50, 839–866. [Google Scholar] [CrossRef] [Green Version]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic Revision of Symbiodiniaceae Highlights the Antiquity and Diversity of Coral Endosymbionts. Curr. Biol. 2018, 28, 2570–2580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davy, S.K.; Allemand, D.; Weis, V.M. Cell Biology of Cnidarian-Dinoflagellate Symbiosis. Microbiol. Mol. Biol. Rev. 2012, 76, 229–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackstone, N.W.; Golladay, J.M. Why Do Corals Bleach? Conflict and Conflict Mediation in a Host/Symbiont Community. BioEssays 2018, 40, 1800021. [Google Scholar] [CrossRef]
- Durand, P.M.; Filho, M.M.B.; Michod, R.E. Cell Death in Evolutionary Transitions in Individuality. Yale J. Biol. Med. 2019, 92, 651–662. [Google Scholar]
- Durand, P.M. The Evolutionary Origins of Life and Death; University of Chicago Press: Chicago, IL, USA, 2021. [Google Scholar]
- Crossland, C.J.; Barnes, D.J.; Borowitzka, M. Diurnal lipid and mucus production in the staghorn coral Acropora acuminata. Mar. Biol. 1980, 60, 81–90. [Google Scholar] [CrossRef]
- Muscatine, L.; Falkowski, P.G.; Porter, J.W.; Dubinsky, Z. Fate of photosynthetic fixed carbon in light- and shade-adapted colonies of the symbiotic coral Stylophora pistillata. Proc. R. Soc. Lond. B 1984, 222, 181–202. [Google Scholar]
- Lutz, A.; Raina, J.-B.; Motti, C.A.; Miller, D.J.; van Oppen, M.J.H. Host Coenzyme Q Redox State Is an Early Biomarker of Thermal Stress in the Coral Acropora millepora. PLoS ONE 2015, 10, e0139290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrin, A.P.; Somova, E.L.; Kern, P.M.; Millet, T.A.; Bross, L.S.; Blackstone, N.W. The use of in vivo microscopy to image the cnidarian stress response. Invertebr. Biol. 2017, 136, 330–344. [Google Scholar] [CrossRef]
- Weis, V.M. Cellular mechanisms of Cnidarian bleaching: Stress causes the collapse of symbiosis. J. Exp. Biol. 2008, 211, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Paxton, C.W.; Davy, S.; Weis, V. Stress and death of cnidarian host cells play a role in cnidarian bleaching. J. Exp. Biol. 2013, 216, 2813–2820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azcon-Bieto, J. Inhibition of Photosynthesis by Carbohydrates in Wheat Leaves. Plant Physiol. 1983, 73, 681–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldschmidt, E.E.; Huber, S.C. Regulation of photosynthesis by end-product accumulation in leaves of plants storing starch, sucrose, and hexose sugars. Plant Physiol. 1992, 99, 1443–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawada, S.; Kuninaka, M.; Watanabe, K.; Sato, A.; Kawamura, H.; Komine, K.; Sakamoto, T.; Kasai, M. The Mechanism to Suppress Photosynthesis Through End-Product Inhibition in Single-Rooted Soybean Leaves during Acclimation to CO2 Enrichment. Plant Cell Physiol. 2001, 42, 1093–1102. [Google Scholar] [CrossRef] [Green Version]
- Anthony, K.R.N.; Hoogenboom, M.O.; Connolly, S.R. Adaptive variation in coral geometry and the optimization of internal colony light climates. Funct. Ecol. 2005, 19, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Klaus, J.S.; Budd, A.F.; Heikoop, J.M.; Fouke, B.W. Environmental controls on corallite morphology in the reef coral Montastraea annularis. Mar. Sci. 2007, 28, 233–260. [Google Scholar]
- Baker, D.M.; Freeman, C.J.; Wong, J.C.; Fogel, M.L.; Knowlton, N. Climate change promotes parasitism in a coral symbiosis. ISME J. 2018, 12, 921–930. [Google Scholar] [CrossRef] [Green Version]
- Strohecker, J.; Golladay, J.; Paramo, M.; Paramo, M.; El Rahmany, W.; Blackstone, N.W. Reactive Oxygen Species and the Stress Response in Octocorals. Physiol. Biochem. Zool. 2021, 94, 395–410. [Google Scholar] [CrossRef] [PubMed]
- Wooldridge, S.A. Is the coral-algae symbiosis really ‘mutually beneficial’ for the partners? BioEssays 2010, 32, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Wooldridge, S.A. Breakdown of the coral-algae symbiosis: Towards formalising a linkage between warm-water bleaching thresholds and the growth rate of the intracellular zooxanthellae. Biogeosciences 2013, 10, 1647–1658. [Google Scholar] [CrossRef] [Green Version]
- Goiran, C.; Al-Moghrabi, S.; Allemand, D.; Jaubert, J. Inorganic carbon uptake for photosynthesis by the symbiotic coral/dinoflagellate association I. Photosynthetic performances of symbionts and dependence on sea water bicarbonate. J. Exp. Mar. Biol. Ecol. 1996, 199, 207–225. [Google Scholar] [CrossRef]
- Lin, S.; Cheng, S.; Song, B.; Zhong, X.; Lin, X.; Li, W.; Li, L.; Zhang, Y.; Zhang, H.; Ji, Z.; et al. The Symbiodinium kawagutii genome illuminates dinoflagellate gene expression and coral symbiosis. Science 2015, 350, 691–694. [Google Scholar] [CrossRef] [Green Version]
- Aranda, M.; Li, Y.; Liew, Y.J.; Baumgartner, S.; Simakov, O.; Wilson, M.C.; Piel, J.; Ashoor, H.; Bougouffa, S.; Bajic, V.B.; et al. Genomes of coral dinoflagellate symbionts highlight evo-lutionary adaptations conducive to a symbiotic lifestyle. Sci. Rep. 2016, 6, 39734. [Google Scholar] [CrossRef] [Green Version]
- van Oppen, M.J.H.; Souter, P.; Howells, E.J.; Heyward, A.; Berkelmans, R. Novel genetic diversity through somatic mutations: Fuel for adaptation of reef corals? Diversity 2011, 3, 405–423. [Google Scholar] [CrossRef] [Green Version]
- Downs, C.A.; Kramarsky-Winter, E.; Martinez, J.; Kushmaro, A.; Woodley, C.M.; Loya, Y.; Ostrander, G.K. Symbiophagy as a cellular mechanism for coral bleaching. Autophagy 2009, 5, 211–216. [Google Scholar] [CrossRef] [Green Version]
- Dunn, S.R.; Schnitzler, C.E.; Weis, V.M. Apoptosis and autophagy as mechanisms of dinoflagellate symbiont release during cnidarian bleaching: Every which way you lose. Proc. R. Soc. B 2007, 274, 3079–3085. [Google Scholar] [CrossRef] [Green Version]
- Lesser, M.P. Elevated temperatures and ultraviolet radiation cause oxidative stress and inhibit photosynthesis in sym-biotic dinoflagellates. Limnol. Oceanogr. 1996, 41, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.J.; Hoegh-Guldberg, O.; Larkum, A.W.D.; Schreiber, U. Temperature-induced bleaching of corals begins with im-pairment of the CO2 fixation mechanism in zooxanthellae. Plant Cell Environ. 1998, 21, 1219–1230. [Google Scholar] [CrossRef]
- Buxton, L.; Takahashi, S.; Hill, R.; Ralph, P.J. Variability in the primary site of photosynthetic damage in Symbiodinium sp. (Dinophyceae) exposed to thermal stress. J. Phycol. 2012, 48, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Bhagooli, R.; Hidaka, M. Release of zooxanthellae with intact photosynthetic activity by the coral Galaxea fascicularis in response to high temperature stress. Mar. Biol. 2004, 145, 329–337. [Google Scholar] [CrossRef]
- Fujise, L.; Yamashita, H.; Suzuki, G.; Koike, L. Expulsion of zooxanthellae (Symbiodinium) from several species of scleractinian corals: Comparison under non-stress conditions and thermal stress conditions. Galax. J. Coral Reef Stud. 2013, 15, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Parrin, A.P.; Goulet, T.L.; Yaeger, M.A.; Bross, L.S.; McFadden, C.S.; Blackstone, N.W. Symbiodinium migration mitigates bleaching in three octocoral species. J. Exp. Mar. Biol. Ecol. 2016, 474, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Sies, H. Oxidative eustress: On constant alert for redox homeostasis. Redox Biol. 2021, 41, 101867. [Google Scholar] [CrossRef] [PubMed]
- Lane, N. The Vital Question: Energy, Evolution, and the Origin of Complex Life; Norton: New York, NY, USA, 2015. [Google Scholar]
- Bonner, J.T. The origins of multicellularity. Integr. Biol. 1998, 1, 27–36. [Google Scholar] [CrossRef]
- Lachmann, M.; Blackstone, N.W.; Haig, D.; Kowald, A.; Michod, R.E.; Szathmáry, E.; Werren, J.H.; Wolpert, L. Group 3: Cooperation and conflict in the evolution of genomes, cells, and multicellular organisms. In Genetic and Cultural Evolution of Cooperation; Hammerstein, P., Ed.; MIT Press: Cambridge, UK, 2003; pp. 327–356. [Google Scholar]
- Lane, N. Power, Sex, Suicide: Mitochondria and the Meaning of Life; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Lane, N.; Martin, W. The energetics of genome complexity. Nature 2010, 467, 929–934. [Google Scholar] [CrossRef]
- Martin, W.F. Physiology, anaerobes, and the origin of mitosing cells 50 years on. J. Theor. Biol. 2017, 434, 2–10. [Google Scholar] [CrossRef]
- Skejo, J.; Garg, S.G.; Sven, B.; Gould, S.B.; Hendriksen, M.; Tria, F.D.K.; Bremer, N.; Franjević, D.; Blackstone, N.W.; Martin, W.F. Evidence for a Syncytial Origin of Eukaryotes from Ancestral State Reconstruction. Genome Biol. Evol. 2021, 13, 1–14. [Google Scholar] [CrossRef]
- Gould, S.B.; Waller, R.F.; McFadden, G.I. Plastid Evolution. Annu. Rev. Plant Biol. 2008, 59, 491–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Baracaldo, P.; Raven, J.A.; Pisani, D.; Knoll, A.D. Early photosynthetic eukaryotes inhabited low-salinity habitats. Proc. Natl. Acad. Sci. USA 2017, 114, E7737–E7745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Embley, T.M.; Martin, W. Eukaryotic evolution, changes and challenges. Nature 2006, 440, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.F.; Garg, S.G.; Zimorski, V. Endosymbiotic theories for eukaryote origin. Philos. Trans. R. Soc. B 2015, 370, 20140330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spang, A.; Saw, J.H.; Jørgensen, S.L.; Zaremba-Niedzwiedzka, K.; Martijn, J.; Lind, A.E.; van Eijk, R.; Schleper, C.; Guy, L.; Ettema, T.J.G. Complex archaea that bridge the gap between prokaryotes and eukaryotes. Nature 2015, 521, 173–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speijer, D. Alternating terminal electron-acceptors at the basis of symbiogenesis: How oxygen ignited eukaryotic evolution. BioEssays 2017, 39, 1600174. [Google Scholar] [CrossRef] [Green Version]
- Dance, A. The mysterious microbes at the root of complex life. Nature 2021, 593, 328–330. [Google Scholar] [CrossRef]
- Liu, Y.; Makarova, K.S.; Huang, W.-C.; Wolf, Y.I.; Nikolskaya, A.N.; Zhang, X.; Cai, M.; Zhang, C.-J.; Xu, W.; Luo, Z.; et al. Expanded diversity of Asgard archaea and their relationships with eukaryotes. Nature 2021, 593, 553–557. [Google Scholar] [CrossRef]
- Chance, B.; Nishimura, M. On the mechanism of chlorophyll-cytochrome interaction: The temperature insensitivity of light-induced cytochrome oxidation in chromatium. Proc. Natl. Acad. Sci. USA 1960, 46, 19–24. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, P. Coupling of Phosphorylation to Electron and Hydrogen Transfer by a Chemi-Osmotic type of Mechanism. Nature 1961, 191, 144–148. [Google Scholar] [CrossRef]
- Dudkina, N.V.; Eubel, H.; Keegstra, W.; Boekema, E.J.; Braun, H.-P. Structure of a mitochondrial supercomplex formed by respiratory-chain complexes I and III. Proc. Natl. Acad. Sci. USA 2005, 102, 3225–3229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moser, C.C.; Keske, J.M.; Warncke, K.; Farid, R.S.; Dutton, P.L. Nature of biological electron transfer. Nature 1992, 355, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Bronstein, J.L. Mutualism; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Chance, B.; Williams, G.R. The respiratory chain and oxidative phosphorylation. Adv. Enzymol. Relat. Subj. Biochem. 1956, 17, 65–134. [Google Scholar] [PubMed]
- Allen, J.F.; Santabarbara, S.; Allen, C.A.; Puthiyaveetil, S. Discrete Redox Signaling Pathways Regulate Photosynthetic Light-Harvesting and Chloroplast Gene Transcription. PLoS ONE 2011, 6, e26372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malone, L.A.; Qian, P.; Mayneord, G.E.; Hitchcock, A.; Farmer, D.A.; Thompson, R.F.; Swainsbury, D.J.K.; Ranson, N.A.; Hunter, N.A.; Johnson, M.P. Cryo-EM structure of the spinach cytochrome b6f complex at 3.6 Å resolution. Nature 2019, 575, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Conway Morris, S. Complexity: The ultimate frontier? EMBO Rep. 2011, 12, 481–482. [Google Scholar] [CrossRef] [Green Version]
- Blackstone, N.W. Why did eukaryotes evolve only once? Genetic and energetic aspects of conflict and conflict mediation. Philos. Trans. R. Soc. B 2013, 368, 20120266. [Google Scholar] [CrossRef] [Green Version]
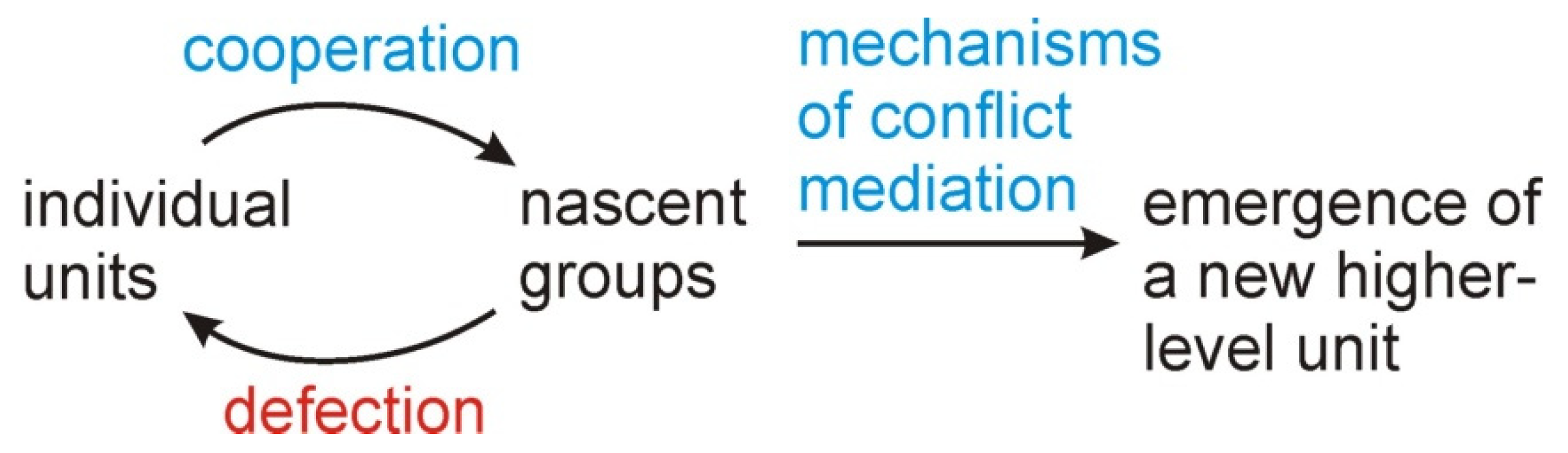
Figure 1.
In this overview of an evolutionary transition, initially only individual units are present within a larger population. Cooperation is more likely to be favored by selection when individual biological units band together to form sub-groups, resulting in a structured population. Defecting individuals, however, weaken group-level cooperation causing nascent groups to collapse. For a higher-level biological unit to fully emerge, mechanisms that mediate these conflicts must be derived (from Blackstone [8]).
Figure 1.
In this overview of an evolutionary transition, initially only individual units are present within a larger population. Cooperation is more likely to be favored by selection when individual biological units band together to form sub-groups, resulting in a structured population. Defecting individuals, however, weaken group-level cooperation causing nascent groups to collapse. For a higher-level biological unit to fully emerge, mechanisms that mediate these conflicts must be derived (from Blackstone [8]).

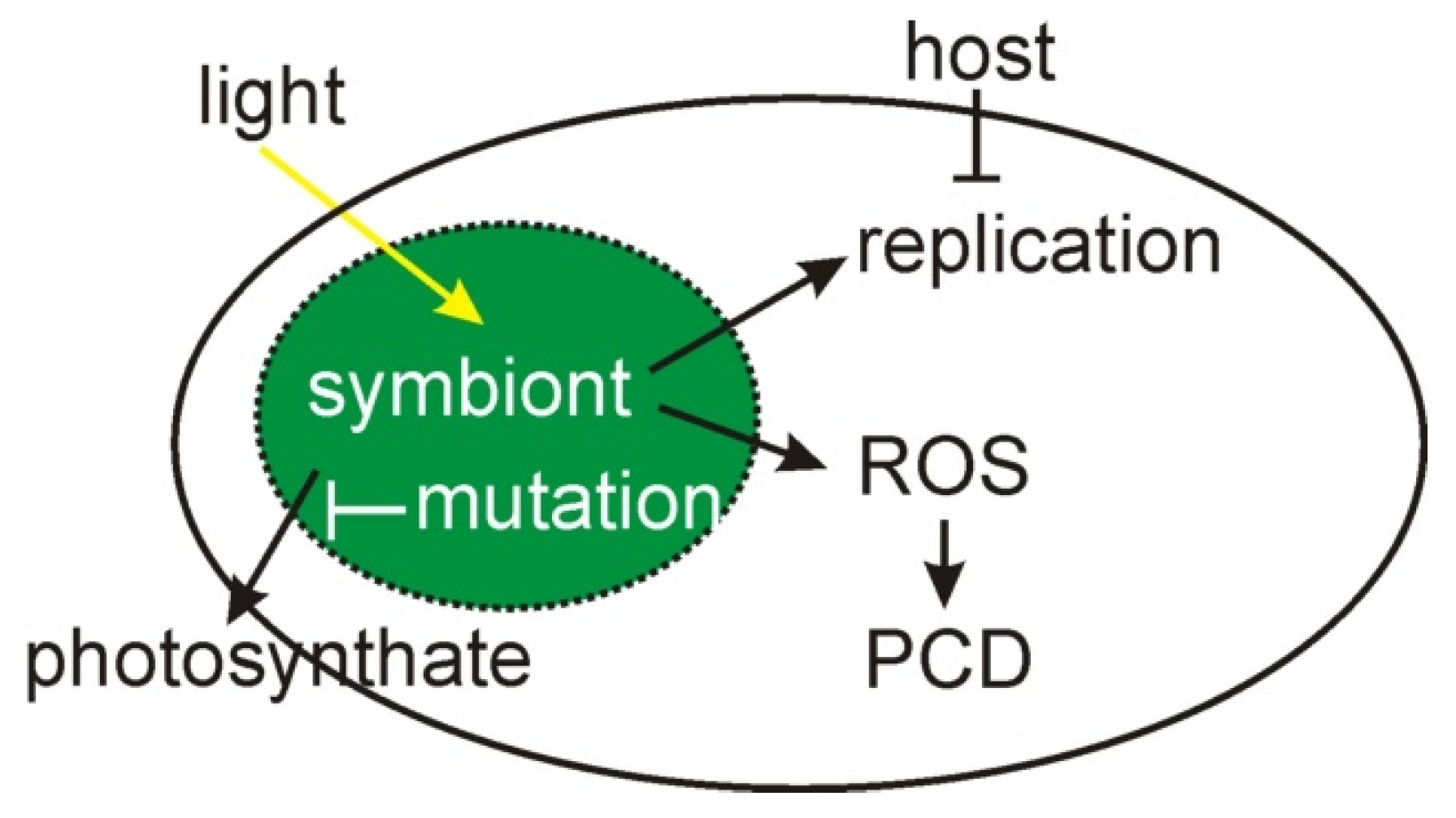
Figure 2.
A schematic of cnidarian photosymbioses and the factors that lead to evolutionary conflict and its mediation. Replication of the symbiont is constrained by the host cell; hence, to avoid product inhibition, the symbiont usually exports photosynthate. This export can be inactivated by mutations to transporter pathway genes. In the presence of light, product inhibition and other mechanisms result in reactive oxygen species (ROS) formation, which might otherwise lead to the export of photosynthate, but under these conditions trigger programmed cell death (PCD) of the host cell (Reprinted with permission from Ref. [16]. 2020, Springer Nature).
Figure 2.
A schematic of cnidarian photosymbioses and the factors that lead to evolutionary conflict and its mediation. Replication of the symbiont is constrained by the host cell; hence, to avoid product inhibition, the symbiont usually exports photosynthate. This export can be inactivated by mutations to transporter pathway genes. In the presence of light, product inhibition and other mechanisms result in reactive oxygen species (ROS) formation, which might otherwise lead to the export of photosynthate, but under these conditions trigger programmed cell death (PCD) of the host cell (Reprinted with permission from Ref. [16]. 2020, Springer Nature).

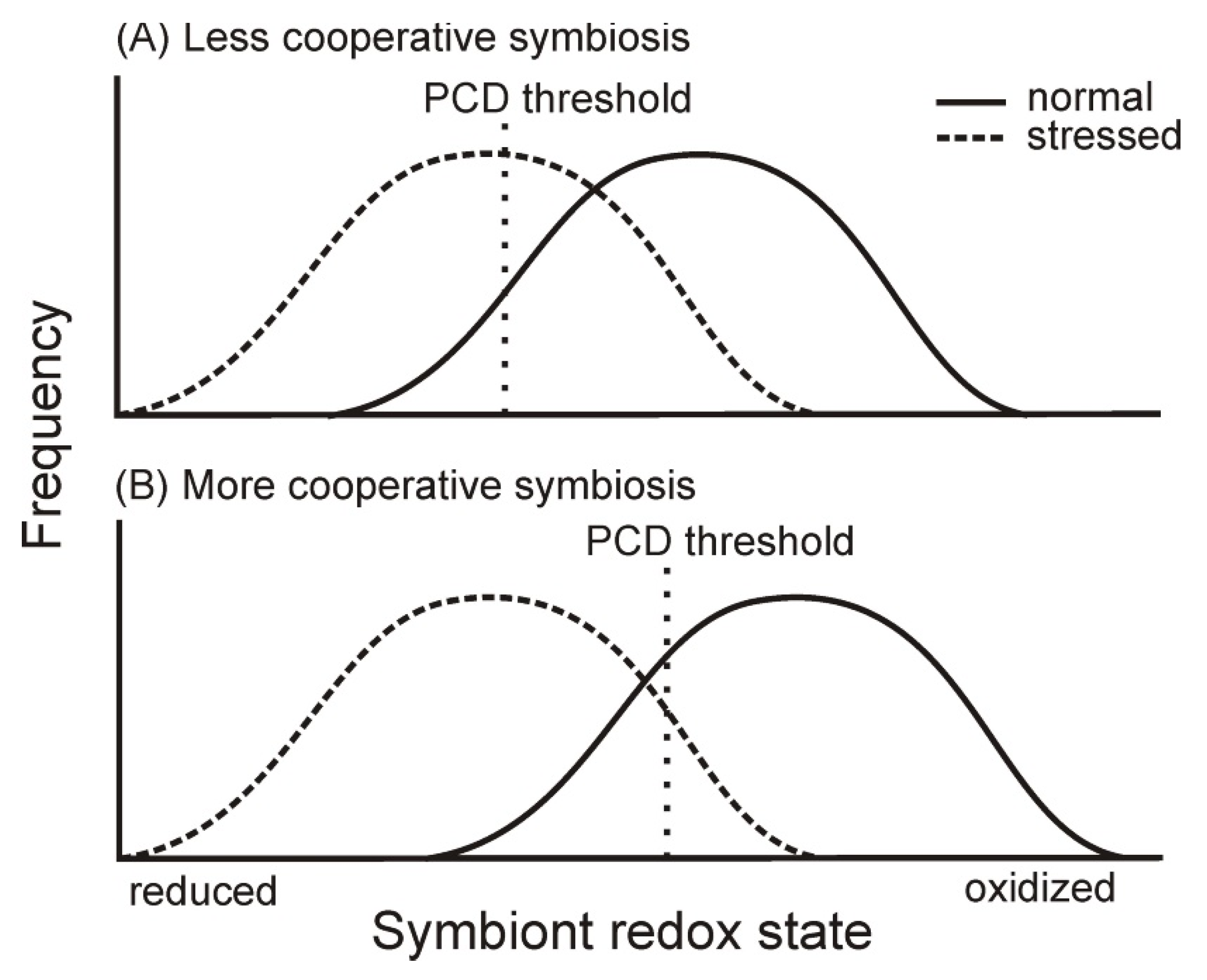
Figure 3.
For (A) less cooperative and (B) more cooperative symbioses, hypothetical frequency distributions are shown for the redox state of symbiont electron carriers in the presence of light (from reduced to oxidized) and hence reactive oxygen species (ROS) formation. The more cooperative symbioses supply substrate (e.g., CO2) and remove photosynthate more effectively, so the redox state of symbiont electron carriers is shifted toward oxidation resulting in low levels of ROS. Under favorable conditions for photosynthesis, in less cooperative symbioses (A) only highly reduced symbionts are destroyed by programmed cell death (PCD), while in more cooperative symbioses (B) the threshold is higher because of lower investment in antioxidant defenses. Under stressful conditions, however, photosynthesis fails, and the advantages of more cooperative symbioses (B) disappear. Distributions of redox state and ROS then shift toward similar distributions, but investment in antioxidants remains the same as do the thresholds for cell death. Symbiont mortality and bleaching is thus greater in more cooperative symbioses (B) than in less cooperative ones (A) (Adapted with permission from Ref. [47]. 2018, Wiley).
Figure 3.
For (A) less cooperative and (B) more cooperative symbioses, hypothetical frequency distributions are shown for the redox state of symbiont electron carriers in the presence of light (from reduced to oxidized) and hence reactive oxygen species (ROS) formation. The more cooperative symbioses supply substrate (e.g., CO2) and remove photosynthate more effectively, so the redox state of symbiont electron carriers is shifted toward oxidation resulting in low levels of ROS. Under favorable conditions for photosynthesis, in less cooperative symbioses (A) only highly reduced symbionts are destroyed by programmed cell death (PCD), while in more cooperative symbioses (B) the threshold is higher because of lower investment in antioxidant defenses. Under stressful conditions, however, photosynthesis fails, and the advantages of more cooperative symbioses (B) disappear. Distributions of redox state and ROS then shift toward similar distributions, but investment in antioxidants remains the same as do the thresholds for cell death. Symbiont mortality and bleaching is thus greater in more cooperative symbioses (B) than in less cooperative ones (A) (Adapted with permission from Ref. [47]. 2018, Wiley).

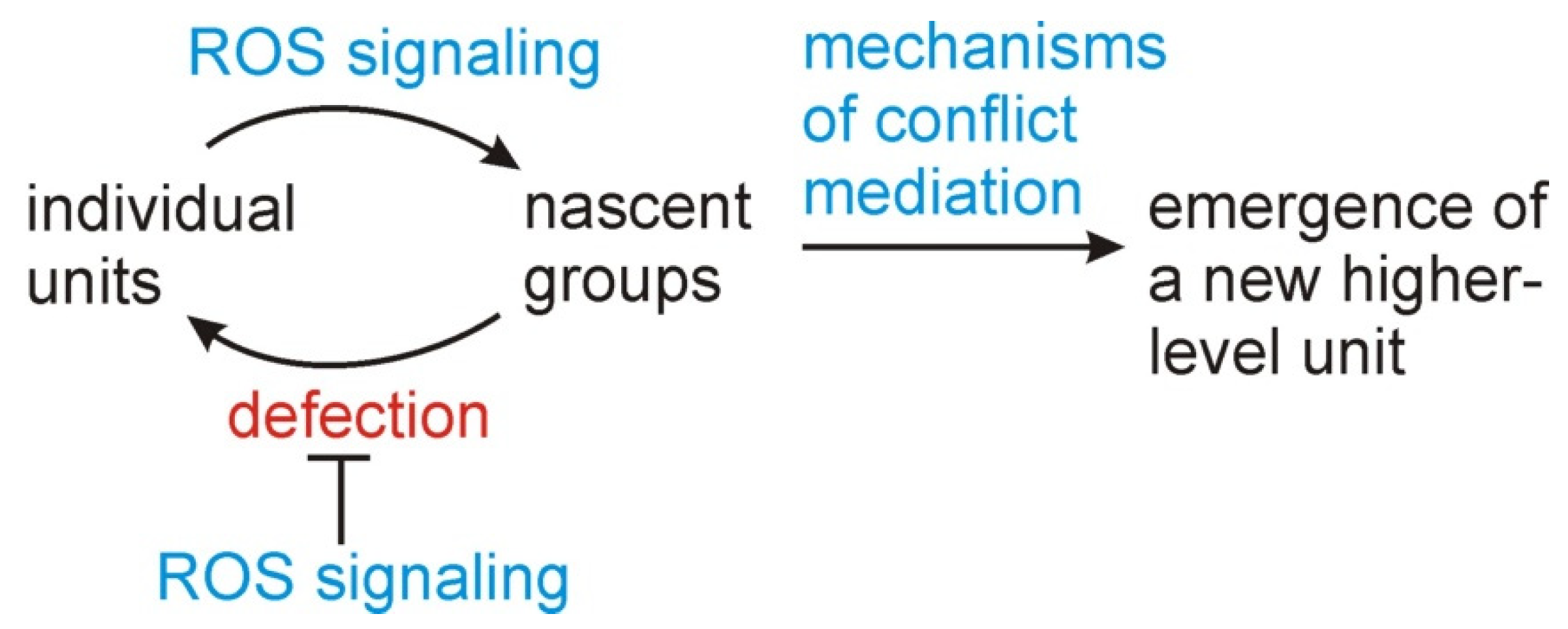
Figure 4.
A modified version of Figure 1 recognizing the role of conflict management early in an evolutionary transition before the existence of higher-level units and a structured population. ROS signaling serves as an arbiter of evolutionary conflict by leading to the formation of nascent groups and selecting against defection from these groups. These groups can then serve as a target for selection, leading to higher-level biological units and mechanisms of conflict mediation. The early stages of a symbiosis or evolutionary transition, when only lower-level selection is operating, may thus be markedly different from later stages, when a higher-level unit has emerged.
Figure 4.
A modified version of Figure 1 recognizing the role of conflict management early in an evolutionary transition before the existence of higher-level units and a structured population. ROS signaling serves as an arbiter of evolutionary conflict by leading to the formation of nascent groups and selecting against defection from these groups. These groups can then serve as a target for selection, leading to higher-level biological units and mechanisms of conflict mediation. The early stages of a symbiosis or evolutionary transition, when only lower-level selection is operating, may thus be markedly different from later stages, when a higher-level unit has emerged.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Blackstone, N.W. Reactive Oxygen Species Signaling Pathways: Arbiters of Evolutionary Conflict? Oxygen 2022, 2, 269-285. https://0-doi-org.brum.beds.ac.uk/10.3390/oxygen2030019
AMA Style
Blackstone NW. Reactive Oxygen Species Signaling Pathways: Arbiters of Evolutionary Conflict? Oxygen. 2022; 2(3):269-285. https://0-doi-org.brum.beds.ac.uk/10.3390/oxygen2030019
Chicago/Turabian StyleBlackstone, Neil W. 2022. "Reactive Oxygen Species Signaling Pathways: Arbiters of Evolutionary Conflict?" Oxygen 2, no. 3: 269-285. https://0-doi-org.brum.beds.ac.uk/10.3390/oxygen2030019