
Expansion of Electron Transport Chain Mutants That Cause Anesthetic-Induced Toxicity in Drosophila melanogaster

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fly Lines and Culturing
2.2. Anesthetic and O2 Exposure
2.3. Nematode Culturing and Exposures
3. Results
3.1. Combining Heterozygous Mutations in Different Subunits of Complex I Significantly Increases Isoflurane-Induced Toxicity
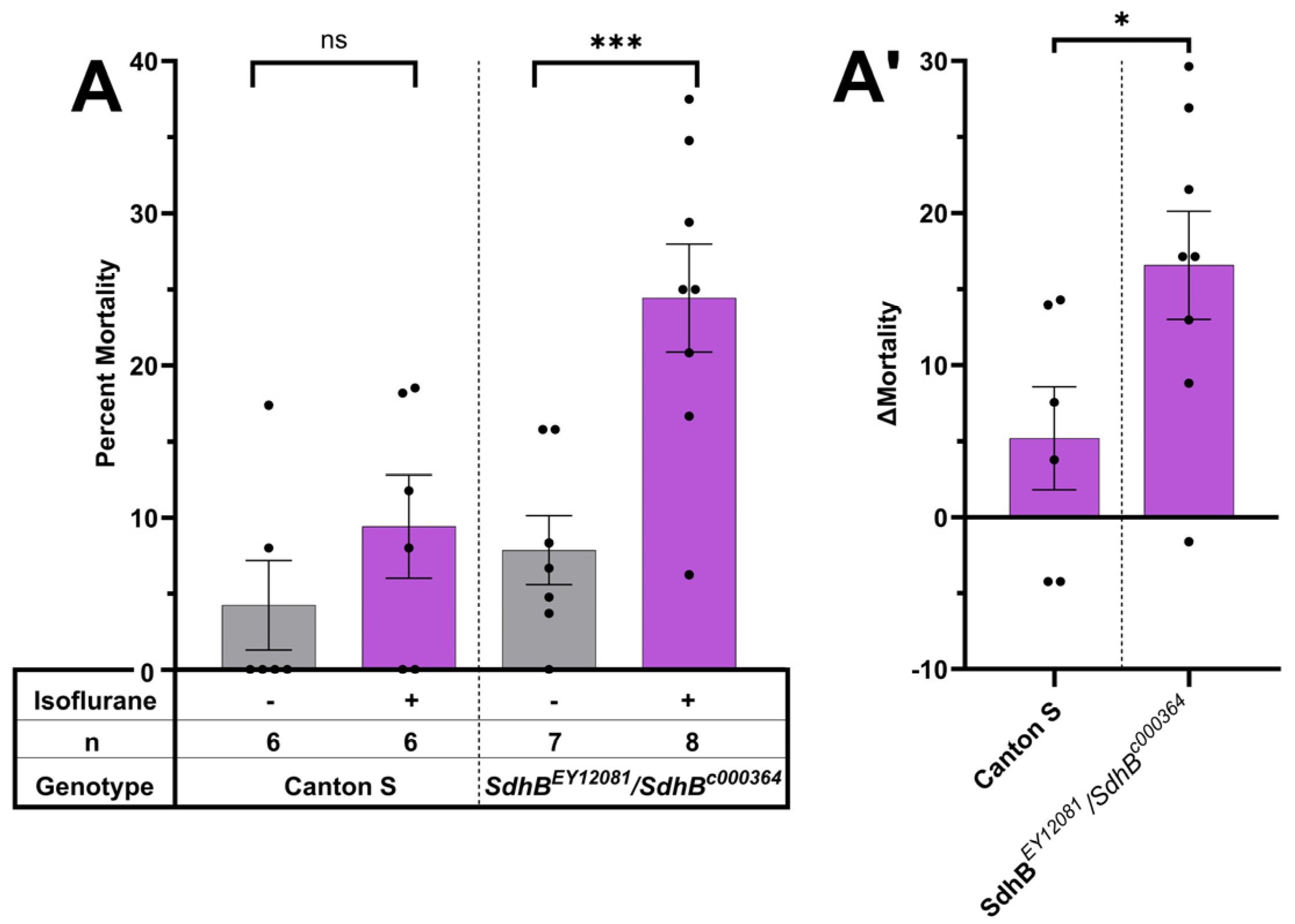
3.2. Compound Heterozygous Mutations in the SdhB Subunit of Complex II Cause Isoflurane-Induced Toxicity in Older Flies
3.3. Mutations in the SdhC subunit of Complex II (mev-1) Sensitizes C. elegans to Isoflurane-Induced Toxicity
4. Discussion
5. Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alston, C.L.; Rocha, M.C.; Lax, N.Z.; Turnbull, D.M.; Taylor, R.W. The genetics and pathology of mitochondrial disease. J. Pathol. 2017, 241, 236–250. [Google Scholar] [CrossRef]
- Morgan, P.G.; Hoppel, C.L.; Sedensky, M.M. Mitochondrial defects and anesthetic sensitivity. Anesthesiology 2002, 96, 1268–1270. [Google Scholar] [CrossRef]
- Niezgoda, J.; Morgan, P.G. Anesthetic considerations in patients with mitochondrial defects. Paediatr. Anaesth. 2013, 23, 785–793. [Google Scholar] [CrossRef]
- Kayser, E.B.; Morgan, P.G.; Sedensky, M.M. GAS-1: A mitochondrial protein controls sensitivity to volatile anesthetics in the nematode Caenorhabditis elegans. Anesthesiology 1999, 90, 545–554. [Google Scholar] [CrossRef]
- Loewen, C.A.; Ganetzky, B. Mito-Nuclear Interactions Affecting Lifespan and Neurodegeneration in a Drosophila Model of Leigh Syndrome. Genetics 2018, 208, 1535–1552. [Google Scholar] [CrossRef]
- Olufs, Z.P.G.; Loewen, C.A.; Ganetzky, B.; Wassarman, D.A.; Perouansky, M. Genetic variability affects absolute and relative potencies and kinetics of the anesthetics isoflurane and sevoflurane in Drosophila melanogaster. Sci. Rep. 2018, 8, 2348. [Google Scholar] [CrossRef]
- Olufs, Z.P.G.; Ganetzky, B.; Wassarman, D.A.; Perouansky, M. Mitochondrial Complex I Mutations Predispose Drosophila to Isoflurane Neurotoxicity. Anesthesiology 2020, 133, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Jain, I.H.; Zazzeron, L.; Goldberger, O.; Marutani, E.; Wojtkiewicz, G.R.; Ast, T.; Wang, H.; Schleifer, G.; Stepanova, A.; Brepoels, K.; et al. Leigh Syndrome Mouse Model Can Be Rescued by Interventions that Normalize Brain Hyperoxia, but Not HIF Activation. Cell Metab. 2019, 30, 824–832.e3. [Google Scholar] [CrossRef] [PubMed]
- Borchardt, L.A.; Scharenbrock, A.R.; Olufs, Z.P.G.; Wassarman, D.A.; Perouansky, M. Mutations in Complex I of the Mitochondrial Electron-Transport Chain Sensitize the Fruit Fly (Drosophila melanogaster) to Ether and Non-Ether Volatile Anesthetics. Int. J. Mol. Sci. 2023, 24, 1843. [Google Scholar] [CrossRef] [PubMed]
- Morgan, P.G.; Sedensky, M.M.; Meneely, P.M.; Cascorbi, H.F. The effect of two genes on anesthetic response in the nematode Caenorhabditis elegans. Anesthesiology 1988, 69, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Ishii, N.; Fujii, M.; Hartman, P.S.; Tsuda, M.; Yasuda, K.; Senoo-Matsuda, N.; Yanase, S.; Ayusawa, D.; Suzuki, K. A mutation in succinate dehydrogenase cytochrome b causes oxidative stress and ageing in nematodes. Nature 1998, 394, 694–697. [Google Scholar] [CrossRef]
- Cohen, P.J. Effect of anesthetics on mitochondrial function. Anesthesiology 1973, 39, 153–164. [Google Scholar] [CrossRef]
- Nahrwold, M.L.; Cohen, P.J. Additive effect of nitrous oxide and halothane on mitochondrial function. Anesthesiology 1973, 39, 534–536. [Google Scholar] [CrossRef]
- Nahrwold, M.L.; Cohen, P.J. The effects of forane and fluroxene on mitochondrial respiration: Correlation with lipid solubility and in-vivo potency. Anesthesiology 1973, 38, 437–444. [Google Scholar] [CrossRef]
- Hartman, P.S.; Ishii, N.; Kayser, E.B.; Morgan, P.G.; Sedensky, M.M. Mitochondrial mutations differentially affect aging, mutability and anesthetic sensitivity in Caenorhabditis elegans. Mech. Ageing Dev. 2001, 122, 1187–1201. [Google Scholar] [CrossRef]
- Mayr, J.A.; Haack, T.B.; Freisinger, P.; Karall, D.; Makowski, C.; Koch, J.; Feichtinger, R.G.; Zimmermann, F.A.; Rolinski, B.; Ahting, U.; et al. Spectrum of combined respiratory chain defects. J. Inherit. Metab. Dis. 2015, 38, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Bourgeron, T.; Rustin, P.; Chretien, D.; Birch-Machin, M.; Bourgeois, M.; Viegas-Pequignot, E.; Munnich, A.; Rotig, A. Mutation of a nuclear succinate dehydrogenase gene results in mitochondrial respiratory chain deficiency. Nat. Genet. 1995, 11, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Fullerton, M.; McFarland, R.; Taylor, R.W.; Alston, C.L. The genetic basis of isolated mitochondrial complex II deficiency. Mol. Genet. Metab. 2020, 131, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Ishii, N.; Takahashi, K.; Tomita, S.; Keino, T.; Honda, S.; Yoshino, K.; Suzuki, K. A methyl viologen-sensitive mutant of the nematode Caenorhabditis elegans. Mutat. Res. 1990, 237, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Randolph, T.R. Pathophysiology of compound heterozygotes involving hemoglobinopathies and thalassemias. Clin. Lab. Sci. 2008, 21, 240–248. [Google Scholar] [PubMed]
- Parfait, B.; Chretien, D.; Rötig, A.; Marsac, C.; Munnich, A.; Rustin, P. Compound heterozygous mutations in the flavoprotein gene of the respiratory chain complex II in a patient with Leigh syndrome. Hum. Genet. 2000, 106, 236–243. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borchardt, L.A.; Olufs, Z.P.G.; Morgan, P.G.; Wassarman, D.A.; Perouansky, M. Expansion of Electron Transport Chain Mutants That Cause Anesthetic-Induced Toxicity in Drosophila melanogaster. Oxygen 2024, 4, 108-116. https://0-doi-org.brum.beds.ac.uk/10.3390/oxygen4010006
Borchardt LA, Olufs ZPG, Morgan PG, Wassarman DA, Perouansky M. Expansion of Electron Transport Chain Mutants That Cause Anesthetic-Induced Toxicity in Drosophila melanogaster. Oxygen. 2024; 4(1):108-116. https://0-doi-org.brum.beds.ac.uk/10.3390/oxygen4010006
Chicago/Turabian StyleBorchardt, Luke A., Zachariah P. G. Olufs, Philip G. Morgan, David A. Wassarman, and Misha Perouansky. 2024. "Expansion of Electron Transport Chain Mutants That Cause Anesthetic-Induced Toxicity in Drosophila melanogaster" Oxygen 4, no. 1: 108-116. https://0-doi-org.brum.beds.ac.uk/10.3390/oxygen4010006