Investigating the Effect of Intrinsic Motivation on Alpha Desynchronization Using Sample Entropy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
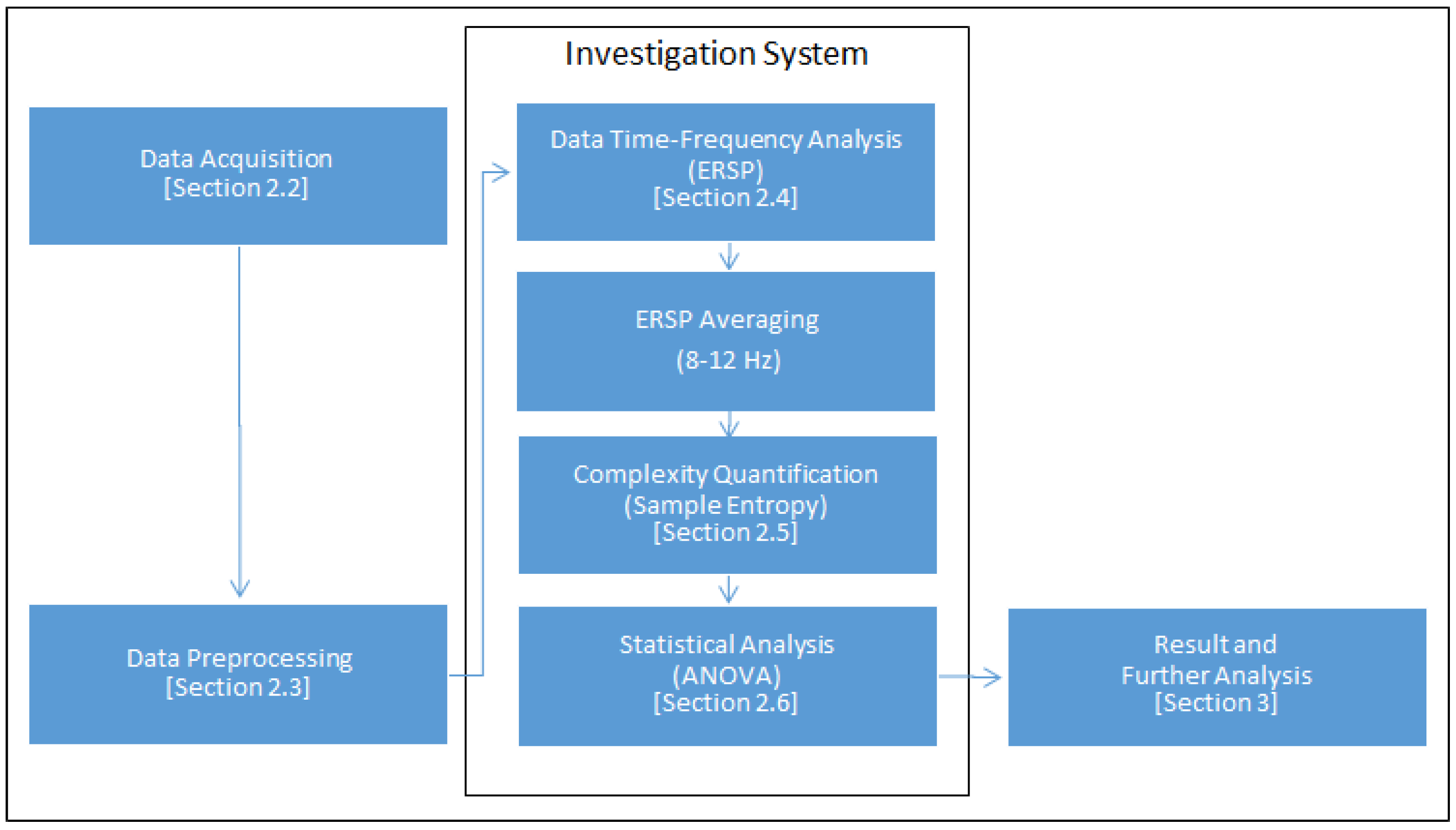
2. Materials and Methods
2.1. Participants
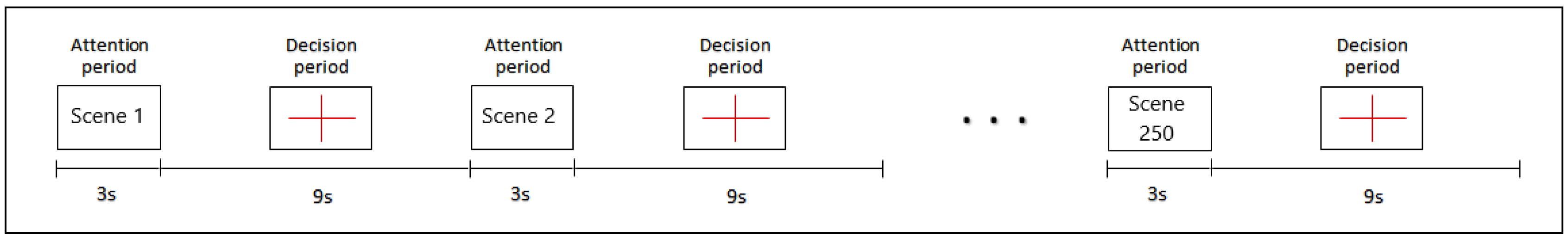
2.2. Stimuli and Procedure
2.3. Preprocessing
2.4. Event-Related Spectral Perturbation
2.5. Sample Entropy
2.6. Statistical Analysis
3. Results and Discussion
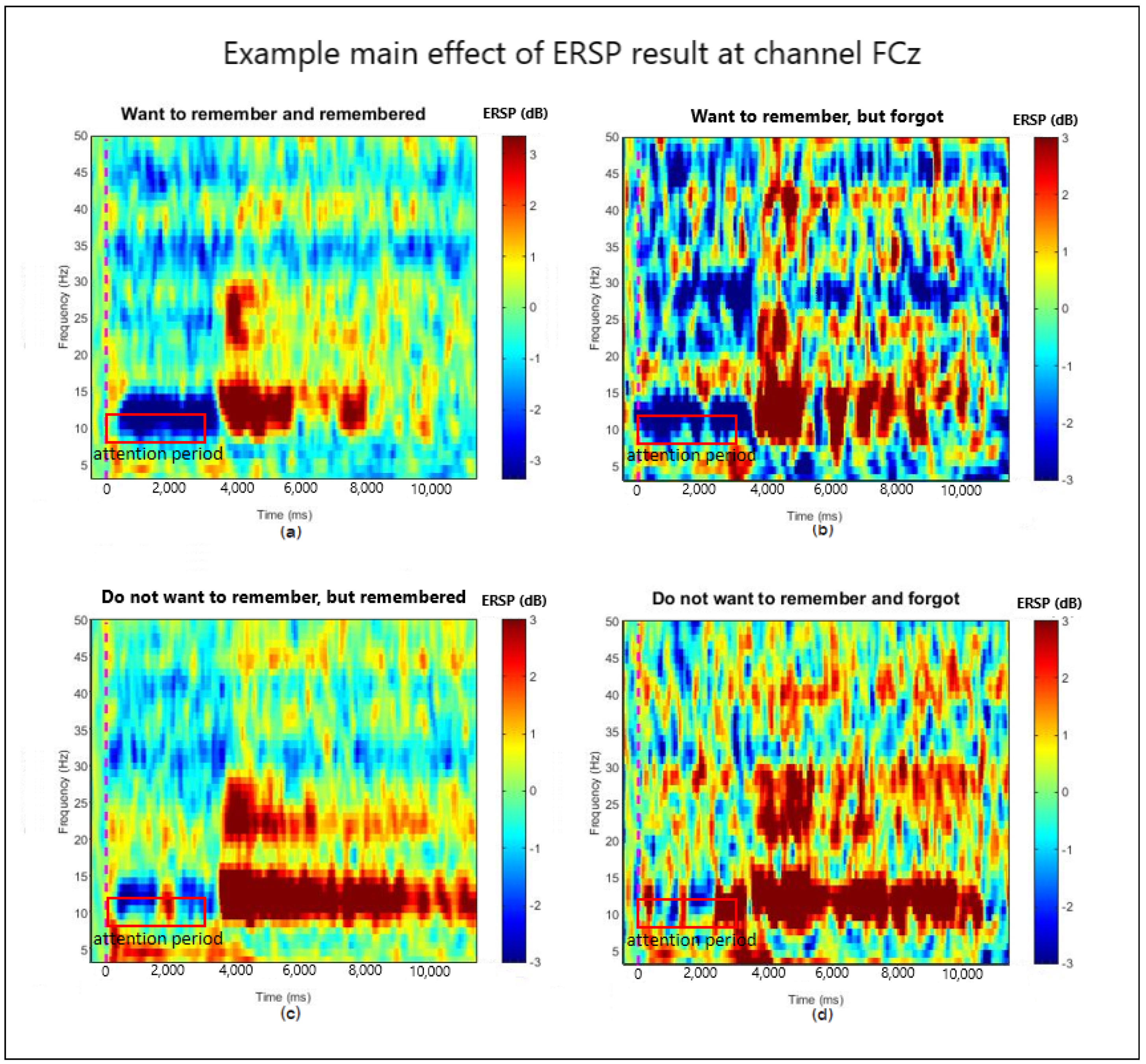
3.1. ERSP Main Effect of Motivation and Recognition
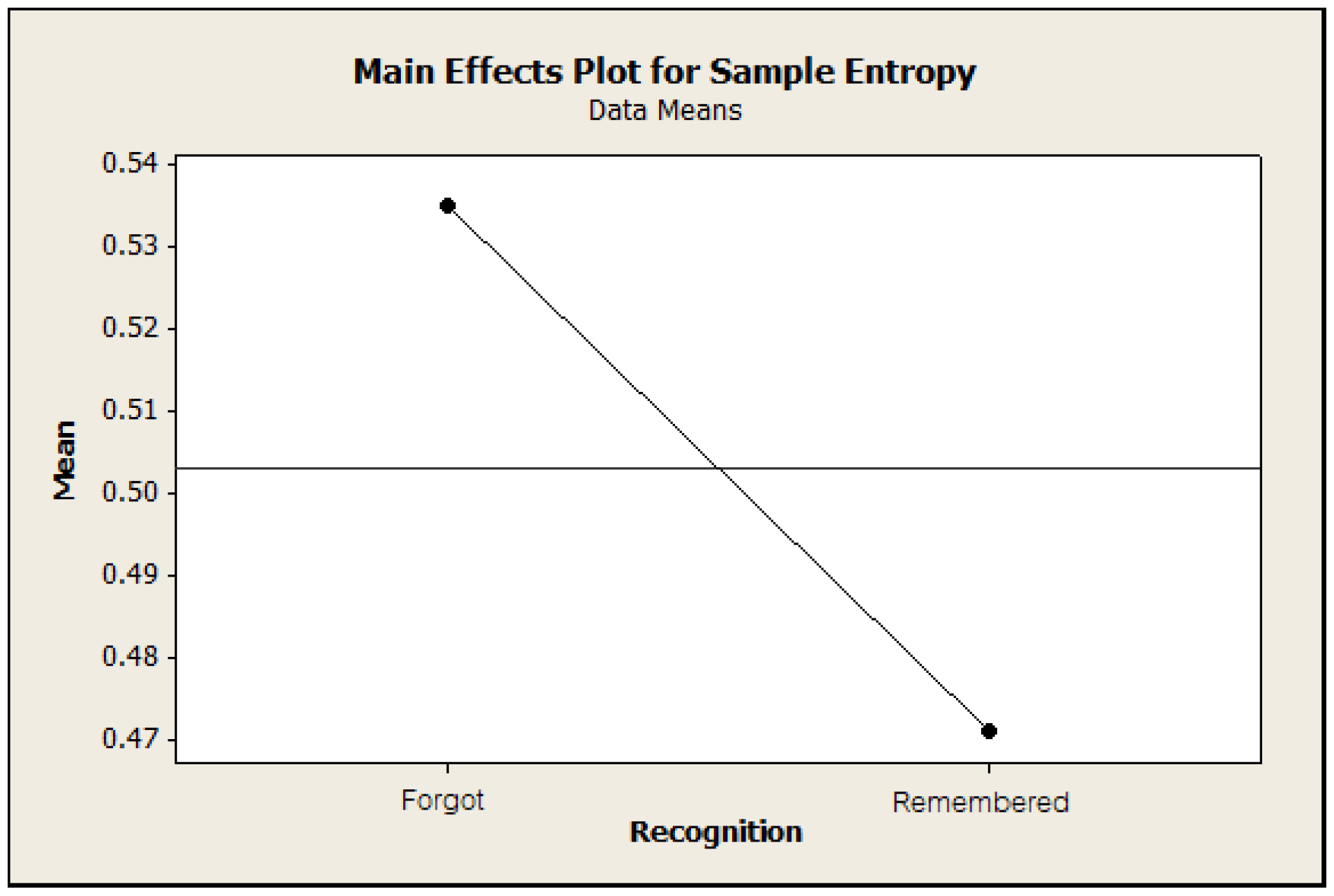
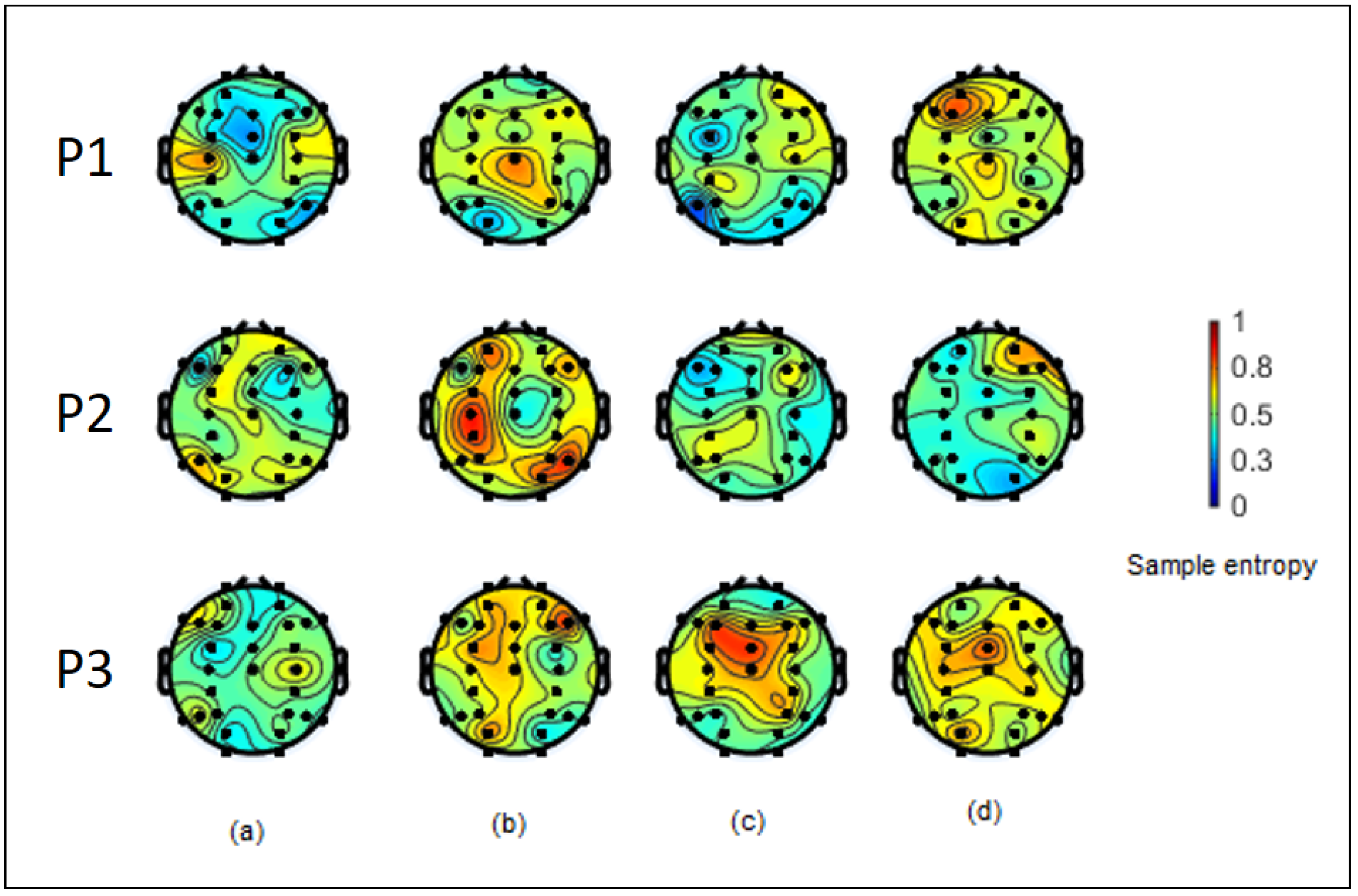
3.2. Sample Entropy Analysis of ERSP
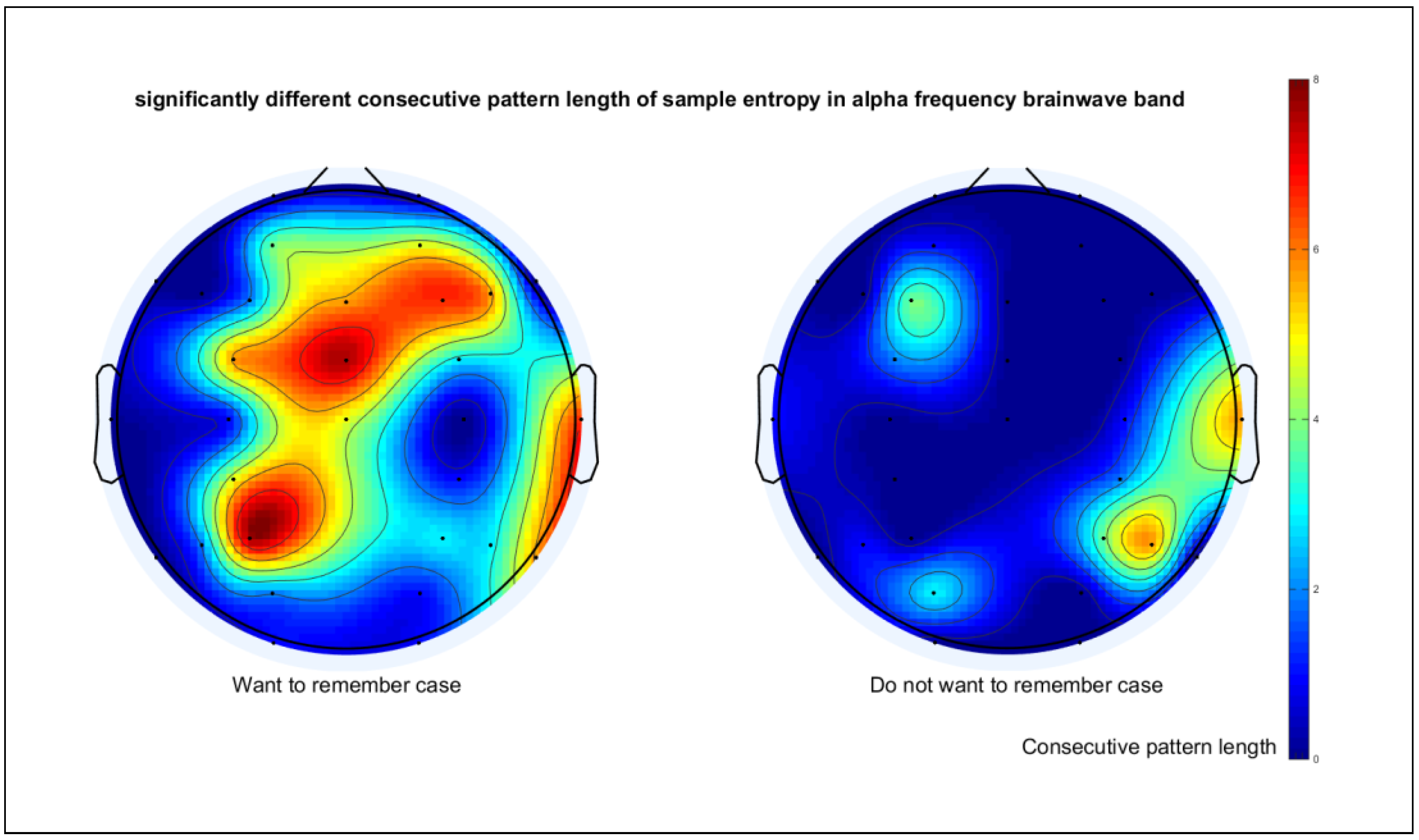
3.3. Continuous Alpha Desynchronization Pattern
| Algorithm 1 Identifying the longest continuous alpha desynchronization pattern. |
| 1: template_size = 0 2: r_remem = 0.2*SD of input_data_remem 3: SE_remem = sampEn(input_data_remem, template_size, r_rate) 4: r_forgot = 0.2*SD of input_data_forgot 5: SE_forgot = sampEn(input_data_forgot, template_size, r_rate) 6: While p-value of paired t-test between SE_remem and SE_forgot > 0.001 do 7: template_size = template_size+1 8: SE_remem = sampEn(input_data_remem, template_size, r_rate) 9: SE_forgot = sampEn(input_data_forgot, template_size, r_rate) 10: End while |
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhu, Z.T.; Yu, M.H.; Riezebos, P. A research framework of smart education. Smart Learn. Environ. 2016, 3, 4–20. [Google Scholar] [CrossRef]
- Donovan, N. The Effects of Intrinsic and Extrinsic Motivation on Cognitive Performance in Humans. Huron Univ. Coll. J. Learn. Motiv. 2015, 53, 5. [Google Scholar]
- Lieberman, H.R.; Tharion, W.J.; Shukitt-Hale, B.; Speckman, K.L.; Tulley, R. Effects of caffeine, sleep loss, and stress on cognitive performance and mood during U.S. Navy SEAL training. Psychopharmacology 2002, 164, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Babiloni, C.; Miniussi, C.; Babiloni, F.; Carducci, F.; Cincotti, F.; Percio, D.C.; Sirello, G.; Fracassi, C.; Nobre, C.A.; Rossini, P.M. Sub-second “temporal attention” modulates alpha rhythms. A high-resolution EEG study. Cogn. Brain Res. 2004, 19, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Dockree, M.P.; Kelly, P.S.; Roche, A.P.R.; Hogan, J.M.; Reilly, B.R.; Robertson, H.I. Behavioural and physiological impairments of sustained attention after traumatic brain injury. Cogn. Brain Res. 2004, 20, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Krause, M.C.; Pörn, B.; Lang, A.H.; Laine, M. Relative alpha desynchronization and synchronization during speech perception. Cogn. Brain Res. 1997, 5, 295–299. [Google Scholar] [CrossRef]
- Klimesch, W. EEG alpha and theta oscillations reflect cognitive and memory performance: A review and analysis. Brain Res. Rev. 1999, 29, 169–195. [Google Scholar] [CrossRef]
- Klimesch, W.; Sauseng, P.; Gerloff, C. Enhancing cognitive performance with repetitive transcranial magnetic stimulation at human individual alpha frequency. Eur. J. Neurosci. 2003, 17, 1129–1133. [Google Scholar] [CrossRef] [PubMed]
- Haegens, S.; Nácher, V.; Luna, R.; Romo, R.; Jensen, O. Alpha-Oscillations in the monkey sensorimotor network influence discrimination performance by rhythmical inhibition of neuronal spiking. Proc. Natl. Acad. Sci. USA 2011, 108, 19377–19382. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.P.; Lalor, E.C.; Reilly, R.B.; Foxe, J.J. Increases in alpha oscillatory power reflect an active retinotopic mechanism for distracter suppression during sustained visuospatial attention. J. Neurophysiol. 2006, 95, 3844–3851. [Google Scholar] [CrossRef] [PubMed]
- Fu, K.M.; Foxe, J.J.; Murray, M.M.; Higgins, B.A.; Javitt, D.C.; Schroeder, C.E. Attention-dependent suppression of distracter visual input can be cross-modally cued as indexed by anticipatory parieto-occipital alpha-band oscillations. Cogn. Brain Res. 2001, 12, 145–152. [Google Scholar] [CrossRef]
- Jin, J.; Yu, L.; Ma, Q. Neural Basis of Intrinsic Motivation: Evidence from Event-Related Potentials. Comput. Intell. Neurosci. 2015, 2015, 698725. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Frey, H.; Molholm, S.; Foxe, J.J. Interests shape how adolescents pay attention: the interaction of motivation and top-down attentional processes in biasing sensory activations to anticipated events. Eur. J. Neurosci. 2015, 41, 818–834. [Google Scholar] [CrossRef] [PubMed]
- Phukhachee, T.; Maneewongvatana, S.; Kaewkamnerdpong, B.; Angsuwatanakul, T.; Iramina, K. Measuring brain activation by using baseline-normalized event-related spectral perturbation in working memory task. EMBC 2017, 41, 4379–4382. [Google Scholar]
- Richman, S.J.; Moorman, R.J. Physiological time-series analysis using approximate entropy and sample entropy. Am. J. Physiol.-Heart Circ. Physiol. 2000, 278, H2039–H2049. [Google Scholar] [CrossRef] [PubMed]
- Yentes, J.M.; Hunt, N.; Schmid, K.K.; Kaipust, J.P.; McGrath, D.; Stergiou, D.N. The Appropriate Use of Approximate Entropy and Sample Entropy with Short Data Sets. Ann. Biomed. Eng. 2013, 41, 349–365. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Hays, J.; Ehinger, A.K.; Oliva, A.; Torralba, A. SUN database: Large-scale scene recognition from abbey to zoo. In Proceedings of the IEEE Computer Society Conference on Computer Vision and Pattern Recognition, San Francisco, CA, USA, 13–18 June 2010; pp. 3485–3492. [Google Scholar]
- Yoo, J.J.; Hinds, O.; Ofen, N.; Hompson, W.T.; Whitfield-Gabrieli, S.; Triantafyllou, C.; Gabrieli, D.E.J. When the brain is prepared to learn: Enhancing human learning using real-time fMRI. NeuroImage 2012, 59, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Makeig, S. Auditory event-related dynamics of the EEG spectrum and effects of exposure to tones. Electroencephalogr. Clin. Neurophysiol. 1993, 86, 283–293. [Google Scholar] [CrossRef]
- Pincus, S.M.; Gladstone, I.M.; Ehrenkranz, R.A. A regularity statistic for medical data analysis. J. Clin. Monit. Comput. 1991, 7, 335–345. [Google Scholar] [CrossRef]
- Rosanova, M.; Casali, A.; Bellina, V.; Resta, F.; Mariotti, M.; Massimini, M. Natural Frequencies of Human Corticothalamic Circuits. J. Neurosci. 2009, 29, 7679–7685. [Google Scholar] [CrossRef] [PubMed]
- Aprigio, D.; Adolfo, W.; Bittencourt, J.; Gongora, M.; Teixeira, S.; Basile, F.L.; Budde, H.; Cagy, M.; Ribeiro, P.; Velasques, B. Alpha power oscillation in the frontal cortex under Bromazepam and Modafinil effects. Arq. Neuro-Psiquiatr. 2015, 73, 918–923. [Google Scholar] [CrossRef] [PubMed]
- Parvaz, M.A.; MacNamara, A.; Goldstein, R.Z.; Hajcak, G. Event-related induced frontal alpha as a marker of lateral prefrontal cortex activation during cognitive reappraisal. Cognit. Affect. Behav. Neurosci. 2012, 12, 730–740. [Google Scholar] [CrossRef] [PubMed]
- Epstein, R.; Kanwisher, N. A cortical representation of the local visual environment. Nature 1998, 392, 598–601. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, K.G.; Zarahn, E.; D’Esposito, M. An Area within Human Ventral Cortex Sensitive to “Building” Stimuli. Neuron 2014, 21, 373–383. [Google Scholar] [CrossRef]
- Ishai, A.; Ungerleider, G.L.; Martin, A.; Schouten, L.J.; Haxby, V.J. Distributed representation of objects in the human ventral visual pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 9379–9384. [Google Scholar] [CrossRef] [PubMed]
- LeCun, Y.; Bengio, Y.; Hinton, G. Deep learning. Nature 2015, 521, 436–444. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phukhachee, T.; Maneewongvatana, S.; Angsuwatanakul, T.; Iramina, K.; Kaewkamnerdpong, B. Investigating the Effect of Intrinsic Motivation on Alpha Desynchronization Using Sample Entropy. Entropy 2019, 21, 237. https://0-doi-org.brum.beds.ac.uk/10.3390/e21030237
Phukhachee T, Maneewongvatana S, Angsuwatanakul T, Iramina K, Kaewkamnerdpong B. Investigating the Effect of Intrinsic Motivation on Alpha Desynchronization Using Sample Entropy. Entropy. 2019; 21(3):237. https://0-doi-org.brum.beds.ac.uk/10.3390/e21030237
Chicago/Turabian StylePhukhachee, Tustanah, Suthathip Maneewongvatana, Thanate Angsuwatanakul, Keiji Iramina, and Boonserm Kaewkamnerdpong. 2019. "Investigating the Effect of Intrinsic Motivation on Alpha Desynchronization Using Sample Entropy" Entropy 21, no. 3: 237. https://0-doi-org.brum.beds.ac.uk/10.3390/e21030237