
Lack of Habituation in Migraine Patients Based on High-Density EEG Analysis Using the Steady State of Visual Evoked Potential

 ,
,  and
and
Abstract
:1. Introduction
2. Related Work
3. Materials and Methods
3.1. Subjects
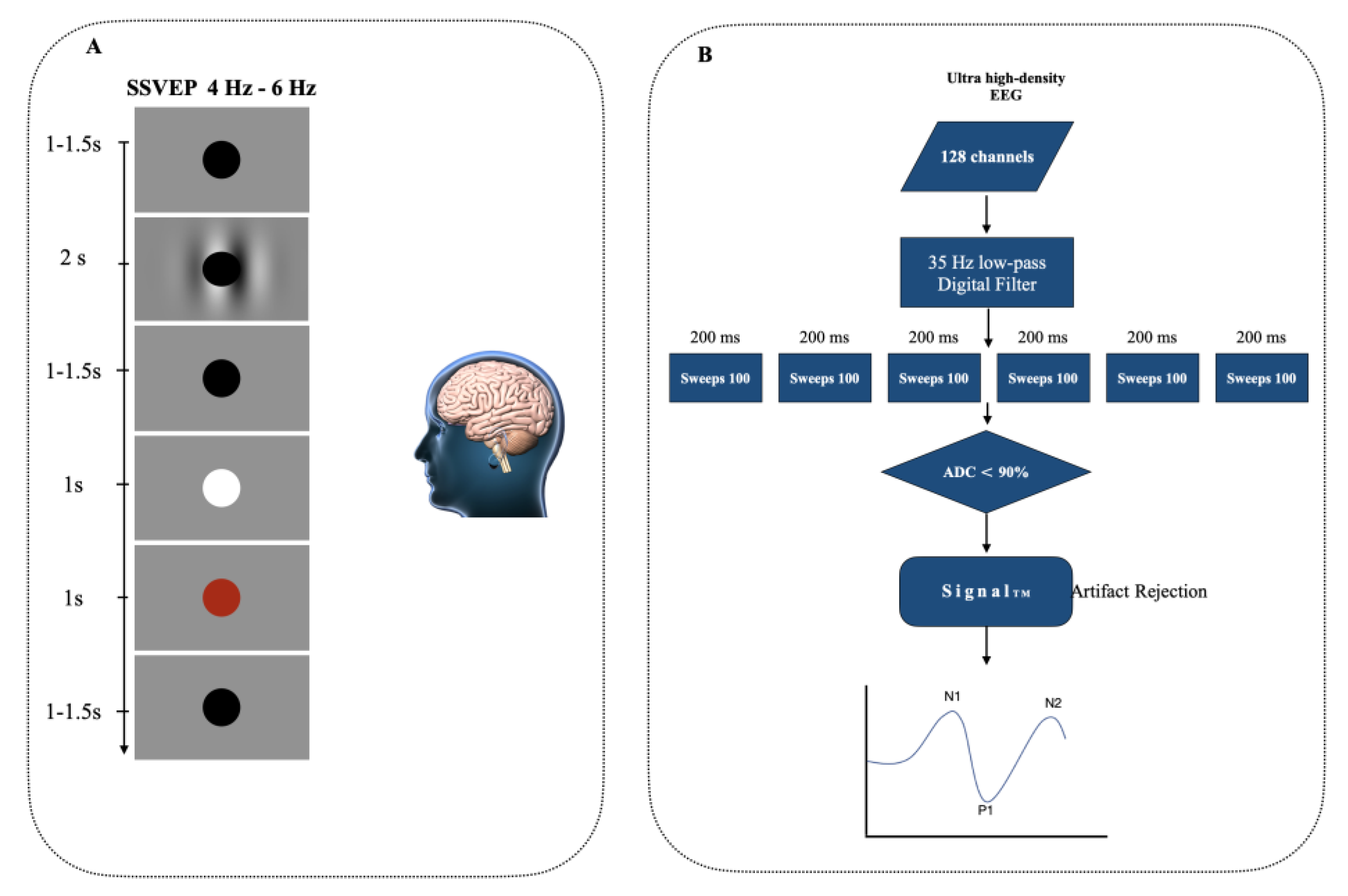
3.2. Stable State Visually Evoked Potentials (SSVEP)
3.3. EEG Capture
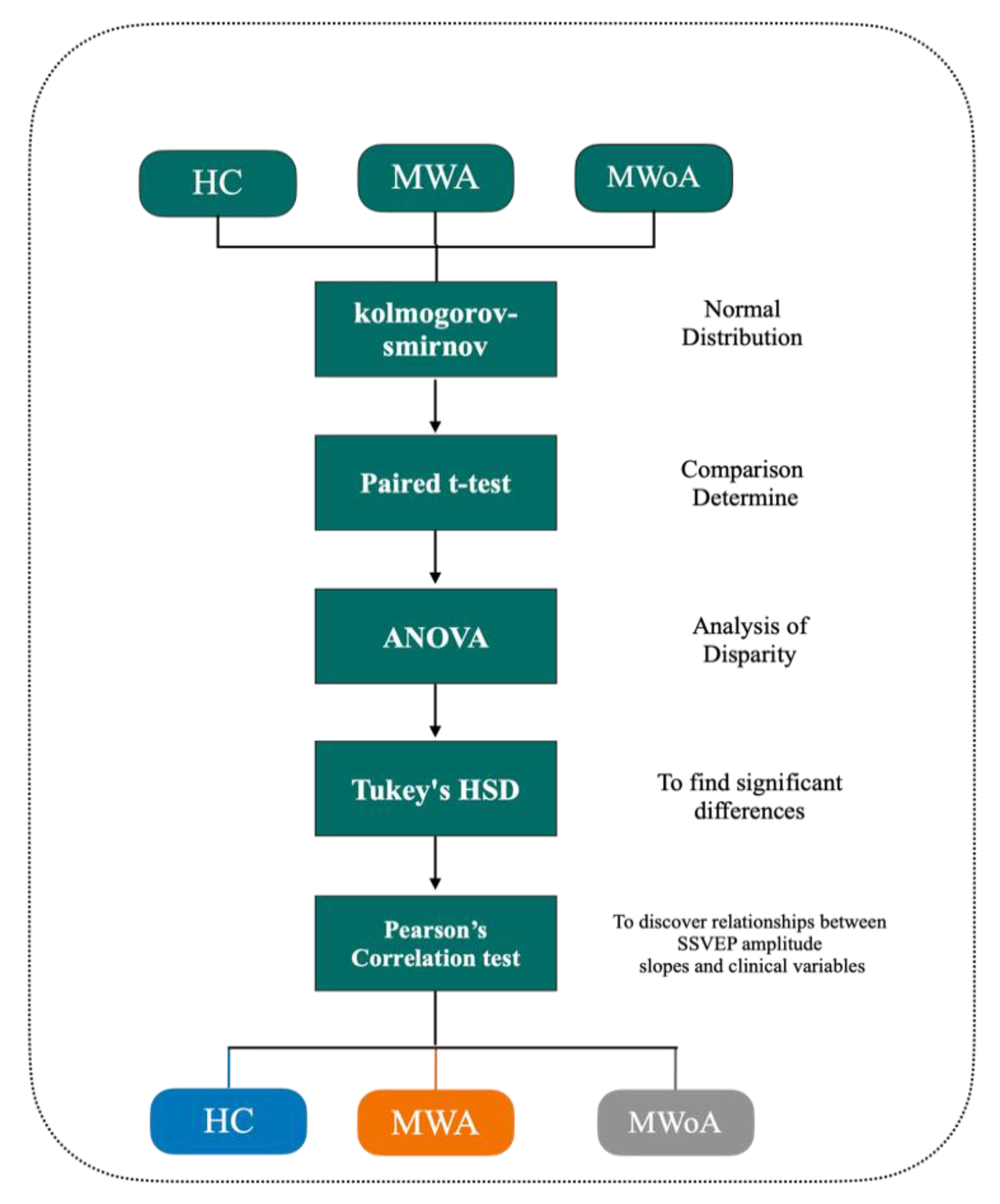
4. Statistics
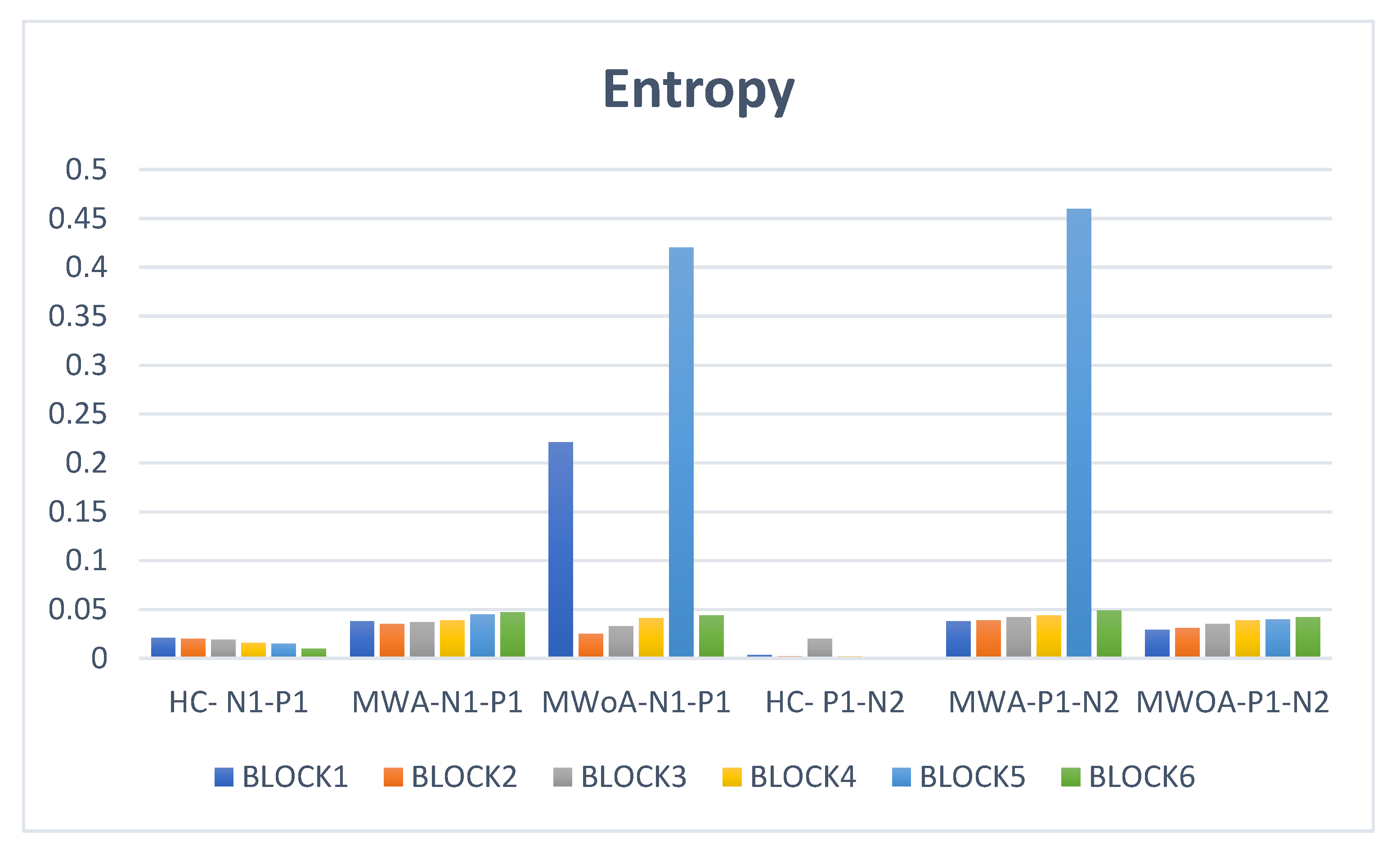
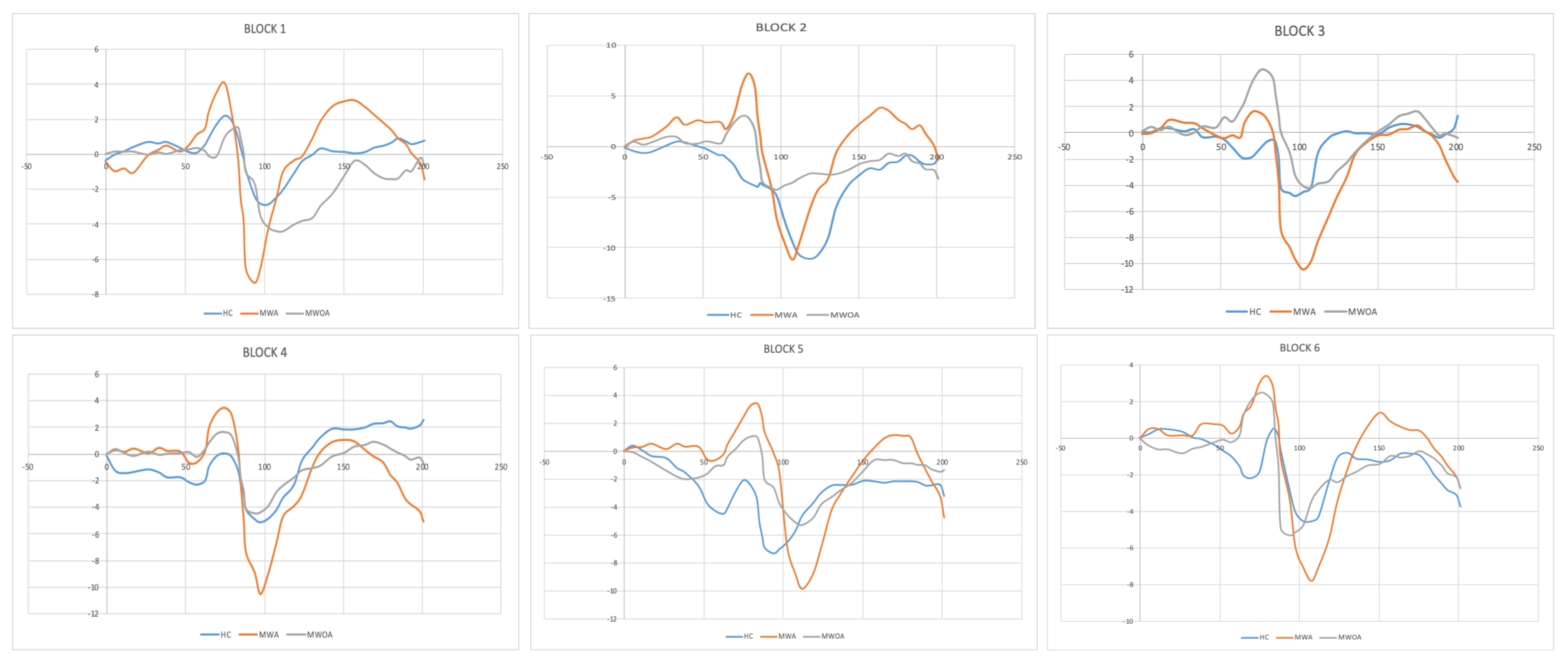
5. Results
6. According to an ANOVA Analysis, There Were Significant Differences between Groups in All Voltage Differences (N1–P1 and P1–N2)
7. Discussion
8. Conclusions and Future Work
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Estave, P.M.; Beeghly, S.; Anderson, R.; Margol, C.; Shakir, M.; George, G.; Berger, A.; O’Connell, N.; Burch, R.; Haas, N.; et al. Learning the full impact of migraine through patient voices: A qualitative study. Headache J. Head Face Pain 2021, 61, 1004–1020. [Google Scholar] [CrossRef] [PubMed]
- Maleki, N.; Szabo, E.; Becerra, L.; Moulton, E.; Scrivani, S.J.; Burstein, R.; Borsook, D. Ictal and interictal brain activation in episodic migraine: Neural basis for extent of allodynia. PLoS ONE 2021, 16, e0244320. [Google Scholar] [CrossRef] [PubMed]
- Headache Classification Committee of the International Headache Society (IHS) The International Classification of Headache Disorders, 3rd edition (beta version). Cephalalgia 2013, 33, 629–808. [CrossRef] [PubMed] [Green Version]
- Abdulhussein, M.A.; An, X.; Alsakaa, A.A.; Ming, D. Lack of habituation in migraine patients and Evoked Potential types: Analysis study from EEG signals. J. Inf. Optim. Sci. 2022, 43, 855–891. [Google Scholar] [CrossRef]
- Vgontzas, A.; Burch, R. Episodic migraine with and without aura: Key differences and implications for pathophysiology, management, and assessing risks. Curr. Pain Headache Rep. 2018, 22, 78. [Google Scholar] [CrossRef]
- Schoenen, J.; Wang, W.; Albert, A.; Delwaide, P. Potentiation instead of habituation characterizes visual evoked potentials in migraine patients between attacks. Eur. J. Neurol. 1995, 2, 115–122. [Google Scholar] [CrossRef]
- Coppola, G.; Iacovelli, E.; Bracaglia, M.; Serrao, M.; Di Lorenzo, C.; Pierelli, F. Electrophysiological correlates of episodic migraine chronification: Evidence for thalamic involvement. J. Headache Pain 2013, 14, 76. [Google Scholar] [CrossRef]
- Bjørk, M.; Hagen, K.; Stovner, L.; Sand, T. Photic EEG-driving responses related to ictal phases and trigger sensitivity in migraine: A longitudinal, controlled study. Cephalalgia 2011, 31, 444–455. [Google Scholar] [CrossRef]
- Coppola, G.; Pierelli, F.; Schoenen, J. Habituation and migraine. Neurobiol. Learn. Mem. 2009, 92, 249–259. [Google Scholar] [CrossRef]
- Bowyer, S.M.; Okada, Y.C.; Papuashvili, N.A.; Moran, J.E.; Barkley, G.L.; Welch, K.M.A.; Tepley, N. Analysis of MEG signals of spreading cortical depression with propagation constrained to a rectangular cortical strip: I. Lissencephalic rabbit model. Brain Res. 1999, 843, 71–78. [Google Scholar] [CrossRef]
- Santos, E.; Schöll, M.; Sanchez-Porras, R.; Kentar, M.; Orakcioglu, B.; Unterberg, A.; Dickhaus, H.; Sakowitz, O.W. Cortical Spreading Depression Dynamics Can Be Studied Using Intrinsic Optical Signal Imaging in Gyrencephalic Animal Cortex. Acta Neurochir. 2013, 118, 93–97. [Google Scholar] [CrossRef]
- Lauritzen, M. Pathophysiology of the migraine aura: The spreading depression theory. Brain 1994, 117, 199–210. [Google Scholar] [CrossRef] [Green Version]
- Mulleners, W.M.; Chronicle, E.P.; Palmer, J.E.; Koehler, P.J.; Vredeveld, J.-W. Visual Cortex Excitability in Migraine With and Without Aura. Headache J. Head Face Pain 2001, 41, 565–572. [Google Scholar] [CrossRef]
- Omland, P.M.; Nilsen, K.B.; Uglem, M.; Gravdahl, G.; Linde, M.; Hagen, K.; Sand, T. Visual Evoked Potentials in Interictal Migraine: No Confirmation of Abnormal Habituation. Headache J. Head Face Pain 2013, 53, 1071–1086. [Google Scholar] [CrossRef]
- Afra, J.; Cecchini, A.P.; De Pasqua, V.; Albert, A.; Schoenen, J. Visual evoked potentials during long periods of pattern-reversal stimulation in migraine. Brain 1998, 121, 233–241. [Google Scholar] [CrossRef]
- Coppola, G.; Parisi, V.; Di Lorenzo, C.; Serrao, M.; Magis, D.; Schoenen, J.; Pierelli, F. Lateral inhibition in visual cortex of migraine patients between attacks. J. Headache Pain 2013, 14, 20. [Google Scholar] [CrossRef] [Green Version]
- Coppola, G.; Di Lorenzo, C.; Schoenen, J.; Pierelli, F. Habituation and sensitization in primary headaches. J. Headache Pain 2013, 14, 65. [Google Scholar] [CrossRef]
- Rolle, C.E.; Narayan, M.; Wu, W.; Toll, R.; Johnson, N.; Caudle, T.; Yan, M.; El-Said, D.; Watts, M.; Eisenberg, M.; et al. Functional connectivity using high density EEG shows competitive reliability and agreement across test/retest sessions. J. Neurosci. Methods 2022, 367, 109424. [Google Scholar] [CrossRef]
- Bromm, B. Human brain electrophysiology. Evoked potentials and evoked magnetic fields in science and medicine. Pain 1989, 39, 371–372. [Google Scholar] [CrossRef]
- Coppola, G.; Ambrosini, A.; Di Clemente, L.; Magis, D.; Fumal, A.; Gérard, P.; Pierelli, F.; Schoenen, J. Interictal Abnormalities of Gamma Band Activity in Visual Evoked Responses in Migraine: An Indication of Thalamocortical Dysrhythmia? Cephalalgia 2007, 27, 1360–1367. [Google Scholar] [CrossRef]
- Golla, F.; Winter, A. Analysis of cerebral responses to flicker in patients complaining of episodic headache. Electroencephalogr. Clin. Neurophysiol. 1959, 11, 539–549. [Google Scholar] [CrossRef]
- Vialatte, F.-B.; Maurice, M.; Dauwels, J.; Cichocki, A. Steady-state visually evoked potentials: Focus on essential paradigms and future perspectives. Prog. Neurobiol. 2010, 90, 418–438. [Google Scholar] [CrossRef] [PubMed]
- Simon, R.H.; Zimmerman, A.W.; Tasman, A.; Hale, M.S. Spectral analysis of photic stimulation in migraine. Electroencephalogr. Clin. Neurophysiol. 1982, 53, 270–276. [Google Scholar] [CrossRef]
- Genco, S.; De Tommaso, M.; Prudenzano, A.; Savarese, M.; Puca, F.M. EEG Features in Juvenile Migraine: Topographic Analysis of Spontaneous and Visual Evoked Brain Electrical Activity: A Comparison with Adult Migraine. Cephalalgia 1994, 14, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Nyrke, T.; Kangasniemi, P.; Lang, A.H. Difference of steady-state visual evoked potentials in classic and common migraine. Electroencephalogr. Clin. Neurophysiol. 1989, 73, 285–294. [Google Scholar] [CrossRef]
- Shibata, K.; Yamane, K.; Otuka, K.; Iwata, M. Abnormal visual processing in migraine with aura: A study of steady-state visual evoked potentials. J. Neurol. Sci. 2008, 271, 119–126. [Google Scholar] [CrossRef]
- De Tommaso, M.; Sciruicchio, V.; Guido, M.; Sasanelli, G.; Specchio, L.M.; Puca, F.M. EEG Spectral Analysis in Migraine Without Aura Attacks. Cephalalgia 1998, 18, 324–328. [Google Scholar] [CrossRef]
- De Tommaso, M.; Stramaglia, S.; Schoffelen, J.M.; Guido, M.; Libro, G.; Losito, L.; Sciruicchio, V.; Sardaro, M.; Pellicoro, M.; Puca, F.M. Steady-state visual evoked potentials in the low frequency range in migraine: A study of habituation and variability phenomena. Int. J. Psychophysiol. 2003, 49, 165–174. [Google Scholar] [CrossRef]
- Shibata, K.; Yamane, K.; Nishimura, Y.; Kondo, H.; Otuka, K. Spatial frequency differentially affects habituation in migraineurs: A steady-state visual-evoked potential study. Doc. Ophthalmol. 2011, 123, 65–73. [Google Scholar] [CrossRef]
- De Tommaso, M.; Ambrosini, A.; Brighina, F.; Coppola, G.; Perrotta, A.; Pierelli, F.; Sandrini, G.; Valeriani, M.; Marinazzo, D.; Stramaglia, S.; et al. Altered processing of sensory stimuli in patients with migraine. Nat. Rev. Neurol. 2014, 10, 144–155. [Google Scholar] [CrossRef]
- Fogang, Y.; Gérard, P.; De Pasqua, V.; Pepin, J.L.; Ndiaye, M.; Schoenen, J.D. Analysis and clinical correlates of 20 Hz photic driving on routine EEG in migraine. Acta Neurol. Belg. 2015, 115, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Judit, A.; Sándor, P.; Schoenen, J. Habituation of visual and intensity dependence of auditory evoked cortical potentials tends to normalize just before and during the migraine attack. Cephalalgia 2000, 20, 714–719. [Google Scholar] [CrossRef]
- Coppola, G.; Bracaglia, M.; Di Lenola, D.; Iacovelli, E.; Di Lorenzo, C.; Serrao, M.; Evangelista, M.; Parisi, V.; Schoenen, J.; Pierelli, F. Lateral inhibition in the somatosensory cortex during and between migraine without aura attacks: Correlations with thalamocortical activity and clinical features. Cephalalgia 2016, 36, 568–578. [Google Scholar] [CrossRef]
- Chamanzar, A.; Haigh, S.M.; Grover, P.; Behrmann, M. Abnormalities in cortical pattern of coherence in interictal migraine detected using ultra high-density EEG. medRxiv 2020. [Google Scholar] [CrossRef]
- Haigh, S.M.; Ms, A.C.; Grover, P.; Behrmann, M. Cortical Hyper-Excitability in Migraine in Response to Chromatic Patterns. Headache J. Head Face Pain 2019, 59, 1773–1787. [Google Scholar] [CrossRef]
- Coppola, G.; Bracaglia, M.; Di Lenola, D.; Di Lorenzo, C.; Serrao, M.; Parisi, V.; di Renzo, A.; Martelli, F.; Fadda, A.; Schoenen, J.; et al. Visual evoked potentials in subgroups of migraine with aura patients. J. Headache Pain 2015, 16, 92. [Google Scholar] [CrossRef] [Green Version]
- Coutin-Churchman, P.; De Freytez, A.P. Vector analysis of visual evoked potentials in migraineurs with visual aura. Clin. Neurophysiol. 2003, 114, 2132–2137. [Google Scholar] [CrossRef]
- Nguyen, B.; McKendrick, A.M.; Vingrys, A. Simultaneous retinal and cortical visually evoked electrophysiological responses in between migraine attacks. Cephalalgia 2012, 32, 896–907. [Google Scholar] [CrossRef]
- Khalil, N.M.; Legg, N.J.; Anderson, D.J. Long term decline of P100 amplitude in migraine with aura. J. Neurol. Neurosurg. Psychiatry 2000, 69, 507–511. [Google Scholar] [CrossRef] [Green Version]
- Alyasseri, Z.A.A.; Khader, A.T.; Al-Betar, M.A.; Papa, J.P.; Alomari, O.A.; Makhadme, S.N. An Efficient Optimization Technique of EEG Decomposition for User Authentication System. In Proceedings of the 2018 2nd International Conference on BioSignal Analysis, Processing and Systems (ICBAPS), Kuching, Malaysia, 24–26 July 2018; pp. 1–6. [Google Scholar] [CrossRef]
- Chen, W.-T.; Wang, S.-J.; Fuh, J.-L.; Lin, C.-P.; Ko, Y.-C.; Lin, Y.-Y. Persistent ictal-like visual cortical excitability in chronic migraine. Pain 2011, 152, 254–258. [Google Scholar] [CrossRef]
- Hoffmann, J.; Baca, S.M.; Akerman, S. Neurovascular mechanisms of migraine and cluster headache. J. Cereb. Blood Flow Metab. 2019, 39, 573–594. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, T.; Tominaga, N.; Kaneko, J.; Sato, M.; Akutsu, T.; Hamada, J.; Sakai, F.; Nishiyama, K. Biphasic neurovascular changes in prolonged migraine aura in familial hemiplegic migraine type 2. J. Neurol. Neurosurg. Psychiatry 2015, 86, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Charles, A.C.; Baca, S.M. Cortical spreading depression and migraine. Nat. Rev. Neurol. 2013, 9, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Alyasseri, Z.A.A.; Khadeer, A.T.; Al-Betar, M.A.; Abasi, A.; Makhadmeh, S.; Ali, N.S. The effects of EEG feature extraction using multi-wavelet decomposition for mental tasks classification. In Proceedings of the International Conference on Information and Communication Technology—ICICT’19, Baghdad, Iraq, 15–16 April 2019; pp. 139–146. [Google Scholar] [CrossRef]
- Gordon, G.R.J.; Choi, H.B.; Rungta, R.L.; Ellis-Davies, G.C.R.; MacVicar, B.A. Brain metabolism dictates the polarity of astrocyte control over arterioles. Nature 2008, 456, 745–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Descamps, B.; Vandemaele, P.; Reyngoudt, H.; Deblaere, K.; Leybaert, L.; Paemeleire, K.; Achten, E. Absence of haemodynamic refractory effects in patients with migraine without aura—An interictal fMRI study. Cephalalgia 2011, 31, 1220–1231. [Google Scholar] [CrossRef] [PubMed]
- Griebe, M.; Flux, F.; Wolf, M.E.; Hennerici, M.G.; Szabo, K. Multimodal Assessment of Optokinetic Visual Stimulation Response in Migraine With Aura. Headache J. Head Face Pain 2014, 54, 131–141. [Google Scholar] [CrossRef]
- Zaletel, M.; Strucl, M.; Bajrović, F.F.; Pogacnik, T. Coupling between Visual Evoked Cerebral Blood Flow Velocity Responses and Visual Evoked Potentials in Migraneurs. Cephalalgia 2005, 25, 567–574. [Google Scholar] [CrossRef]
- Hamel, E.; Currents, H. Serotonin and Migraine: Biology and Clinical Implications. Cephalalgia 2007, 27, 1293–1300. [Google Scholar] [CrossRef]
- Mohamed, R.E.; Aboelsafa, A.A.; Al-Malt, A.M. Interictal alterations of thalamic metabolic concentration ratios in mi-graine without aura detected by proton magnetic resonance spectroscopy. Egypt. J. Radiol. Nucl. Med. 2013, 44, 859–870. [Google Scholar] [CrossRef] [Green Version]
- Reyngoudt, H.; Achten, E.; Paemeleire, K. Magnetic resonance spectroscopy in migraine: What have we learned so far? Cephalalgia 2012, 32, 845–859. [Google Scholar] [CrossRef] [Green Version]
- Cevoli, S.; Favoni, V.; Cortelli, P. Energy Metabolism Impairment in Migraine. Curr. Med. Chem. 2019, 26, 6253–6260. [Google Scholar] [CrossRef]
- Gross, E.C.; Putananickal, N.; Orsini, A.-L.; Vogt, D.R.; Sandor, P.S.; Schoenen, J.; Fischer, D. Mitochondrial function and oxidative stress markers in higher-frequency episodic migraine. Sci. Rep. 2021, 11, 4543. [Google Scholar] [CrossRef]
- Bron, C.; Sutherland, H.G.; Griffiths, L.R. Exploring the Hereditary Nature of Migraine. Neuropsychiatr. Dis. Treat. 2021, 17, 1183–1194. [Google Scholar] [CrossRef]
- Di Lorenzo, C.; Pierelli, F.; Coppola, G.; Grieco, G.S.; Rengo, C.; Ciccolella, M.; Magis, D.; Bolla, M.; Casali, C.; Santorelli, F.M.; et al. Mitochondrial DNA haplogroups influence the therapeutic response to riboflavin in migraineurs. Neurology 2009, 72, 1588–1594. [Google Scholar] [CrossRef]
- Rasmussen, A.H.; Olofsson, I.A.; Chalmer, M.A.; Olesen, J.; Hansen, T.F. Higher burden of rare frameshift indels in genes related to synaptic transmission separate familial hemiplegic migraine from common types of migraine. J. Med. Genet. 2020, 57, 610–616. [Google Scholar] [CrossRef] [Green Version]
- Nyholt, D.R.; Anttila, V.; Winsvold, B.S.; Kurth, T.; Stefansson, H.; Kallela, M.; Malik, R.; De Vries, B.; Terwindt, G.M.; Ikram, M.A.; et al. Concordance of genetic risk across migraine subgroups: Impact on current and future genetic association studies. Cephalalgia 2015, 35, 489–499. [Google Scholar] [CrossRef]
- Tang, C.; Unekawa, M.; Shibata, M.; Tomita, Y.; Izawa, Y.; Sugimoto, H.; Ikeda, K.; Kawakami, K.; Suzuki, N.; Nakahara, J. Characteristics of cortical spreading depression and c-Fos expression in transgenic mice having a mutation associated with familial hemiplegic migraine 2. Cephalalgia 2020, 40, 1177–1190. [Google Scholar] [CrossRef]
- De Boer, I.; Maagdenberg, A.M.V.D.; Terwindt, G.M. Advance in genetics of migraine. Curr. Opin. Neurol. 2019, 32, 413–421. [Google Scholar] [CrossRef]
- Sutherland, H.G.; Albury, C.L.; Griffiths, L.R. Advances in genetics of migraine. J. Headache Pain 2019, 20, 72. [Google Scholar] [CrossRef]
- Moeller, F.; Siebner, H.R.; Wolff, S.; Muhle, H.; Granert, O.; Jansen, O.; Stephani, U.; Siniatchkin, M. Mapping brain activity on the verge of a photically induced generalized tonic-clonic seizure. Epilepsia 2009, 50, 1632–1637. [Google Scholar] [CrossRef]
- Hayrunnisa, B.; Doga, V.; Goadsby, P.J. Aura and Head pain: Relationship and gaps in the translational models. J. Headache Pain 2019, 20, 94. [Google Scholar]
- Datta, R.; Aguirre, G.K.; Hu, S.; Detre, J.A.; Cucchiara, B. Interictal cortical hyperresponsiveness in migraine is directly related to the presence of aura. Cephalalgia 2013, 33, 365–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, H.-L.; Zhou, X.; Chen, Y.-C.; Yu, Y.-S.; Guo, X.; Zhou, G.-P.; Zhou, Q.-Q.; Qu, L.-J.; Yin, X.; Li, J.; et al. Impaired intrinsic functional connectivity between the thalamus and visual cortex in migraine without aura. J. Headache Pain 2019, 20, 116. [Google Scholar] [CrossRef] [PubMed]
- Carneiro-Nascimento, S.; Levy, D. Cortical spreading depression and meningeal nociception. Neurobiol. Pain 2022, 11, 100091. [Google Scholar] [CrossRef]
- Shepherd, A.J.; Wyatt, G.; Tibber, M.S. Visual metacontrast masking in migraine. Cephalalgia 2011, 31, 346–356. [Google Scholar] [CrossRef]
- Al-Qazzaz, N.K.; Alyasseri, Z.A.A.; Abdulkareem, K.H.; Ali, N.S.; Al-Mhiqani, M.N.; Guger, C. EEG feature fusion for motor imagery: A new robust framework towards stroke patients rehabilitation. Comput. Biol. Med. 2021, 137, 104799. [Google Scholar] [CrossRef]
- Cerda-Company, X.; Penacchio, O.; Otazu, X. Chromatic Induction in Migraine. Vision 2021, 5, 37. [Google Scholar] [CrossRef]
- Bell, T.; Stokoe, M.; Khaira, A.; Webb, M.; Noel, M.; Amoozegar, F.; Harris, A.D. GABA and glutamate in pediatric migraine. Pain 2021, 162, 300–308. [Google Scholar] [CrossRef]
- Bednář, M.; Kubová, Z.; Kremláček, J. Lack of visual evoked potentials amplitude decrement during prolonged reversal and motion stimulation in migraineurs. Clin. Neurophysiol. 2014, 125, 1223–1230. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Authors | Method | Cases | Phase | Result | |
|---|---|---|---|---|---|
| 1 | (Golla and Winter 1959) [21] | 3.5–25 | 50 MIG 113 HC | Inter-ictal | Increased amplitude and lack of habituation (SSVEPs) |
| 2 | (Simon et al., 1982) [23] | 10–40 Hz | 11 MIG 11 HC | Inter-ictal | Increased power in the alpha band at 24, 34, 38 Hz stimulation |
| 3 | (Nyrke, Kangasniemi, and Lang 1989) [25] | 10–24 Hz | 30 MWA 20 MWoA 49 HC | Inter-ictal | In MWA, increased power at 16–22 Hz, in MWoA reduced 2nd harmonic |
| 4 | (Genco et al., 1994) [24] | 15–30 HZ | 15 teen MWA (10–18) 25 teen MWoA (10–17) 11 teen HC (10–18) 20 adult MWA (19–45) 22 adult MWoA (19–45) 20 adult HC (18–45) | Inter-ictal | Increased amplitude of the SSVEP in all migraine patients |
| 5 | (de Tommaso et al., 1998) [27] | 27 Hz | 16 MWoA 20 HC | Ictal, Inter-ictal | In inter-ictal phase increased power in MWoA, In ictal phase normal power in MWoA and HC |
| 6 | (de Tommaso et al., 2003) [28] | 3 Hz, 6 Hz, 9 Hz | 15 MWoA 15 HC | Inter-ictal | No habituation phenome in MWoA and HC, increased power at 3 Hz in MWoA |
| 7 | (Shibata et al., 2008) [26] | 5–10 Hz | 10 MWoA 10 MWA 20 HC | Inter-ictal | MWA and MWoA have abnormal excitability in the primary visual cortex and significantly increased amplitude to SSVEP |
| 8 | (Shibata et al., 2011) [29] | 0.5, 1.0, 2.0, 4.0 (cpd) with a stimulus rate of 7.5 Hz | 12 MWoA 12 MWA 12 HC | Inter-ictal | MWA and MWoA showed high amplitude to SSVEPs and did not reveal a clear lack of habituation |
| 9 | (Fogang et al., 2015) [31] | 5 Hz, 10 Hz, 15 Hz, 20 Hz | 171 MWoA 61 MWA 48 C M 24 HV | Inter-ictal | The lack of habituation of cortical responses during repetitive stimulation might identify subgroups of migraine patients on spectral analysis of the EEG because of normal habituation in chronic migraine of the evoked activities and PD lower power. |
| Demographic Data and Clinical Characteristics | HC (n = 27) | MWA (n = 13) | MWoA (n = 19) |
|---|---|---|---|
| Women | 8 | 7 | 5 |
| Age (years) | 27 ± 1 | 28 ± 1 | 30 ± 1 |
| Duration of migraine history (years) | 15.2 ± 8.1 | 14.1 ± 2.3 | |
| Attack frequency/month (n) | 2.8 ± 2.1 | 2.1 ± 1.2 | |
| Attack duration (hours) | 28.8 ± 19.7 | 24.6 ± 20.3 | |
| Days since the last migraine attack | 14.7 ± 18.1 | 19.9 ± 17.1 |
| EEG Parameters (ms) | HC | MWoA | MWA |
|---|---|---|---|
| N1 (75) | 74.7 ± 1.2 | 74.7 ± 6.3 | 74.7 ± 5.5 |
| P1 (100) | 102.1 ± 0.1 | 101.7 ± 7.4 | 101.1 ± 3.2 |
| N2 (145) | 135.4 ± 3.2 | 156.8 ± 6.5 | 150.3 ± 7.3 |
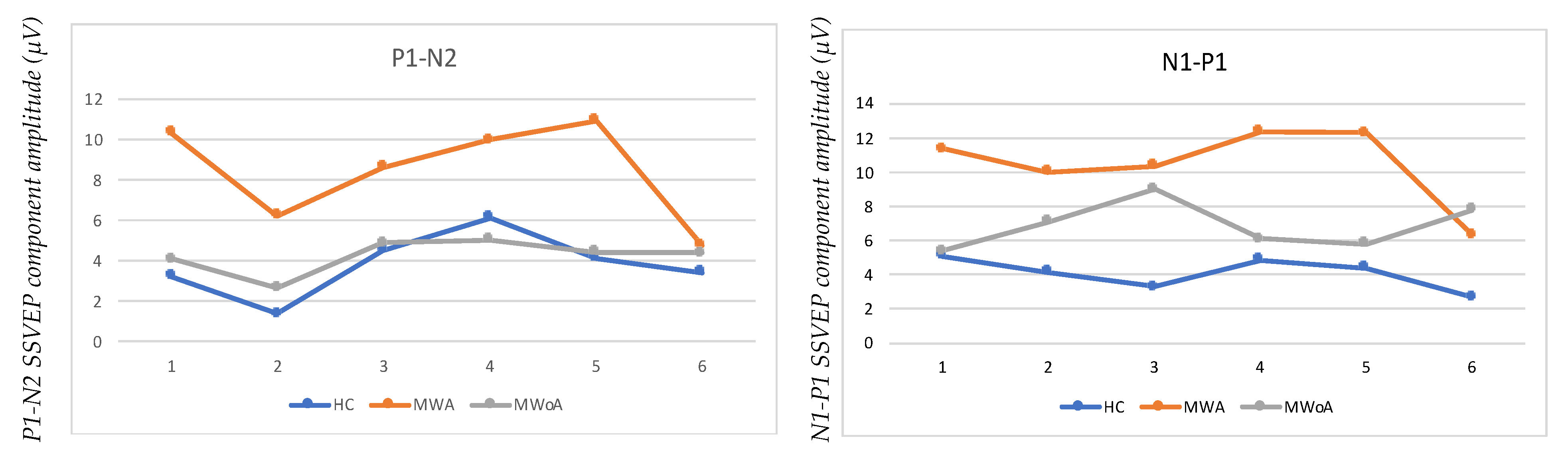
| HC-N1–P1 | MWA-N1–P1 | MWoA-N1–P1 | HC-P1–N2 | MWA-P1–N2 | MWOA-P1–N2 | |
|---|---|---|---|---|---|---|
| BLOCK1 | 5.0942227 | 11.3879 | 5.426988 | 3.2195488 | 10.34833 | 4.062311 |
| BLOCK2 | 4.2105263 | 10 | 7.122642 | 1.3815789 | 6.231884 | 2.641509 |
| BLOCK3 | 3.3333333 | 10.38043 | 9.027778 | 4.5075758 | 8.641304 | 4.861111 |
| BLOCK4 | 4.9017821 | 12.35472 | 6.097765 | 6.1074421 | 9.960602 | 5.011181 |
| BLOCK5 | 4.4410654 | 12.27959 | 5.841923 | 4.1069709 | 10.91944 | 4.440718 |
| BLOCK6 | 2.7193086 | 6.375129 | 7.819883 | 3.4170887 | 4.753607 | 4.360018 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbas Abdulhussein, M.; Alyasseri, Z.A.A.; Mohammed, H.J.; An, X. Lack of Habituation in Migraine Patients Based on High-Density EEG Analysis Using the Steady State of Visual Evoked Potential. Entropy 2022, 24, 1688. https://0-doi-org.brum.beds.ac.uk/10.3390/e24111688
Abbas Abdulhussein M, Alyasseri ZAA, Mohammed HJ, An X. Lack of Habituation in Migraine Patients Based on High-Density EEG Analysis Using the Steady State of Visual Evoked Potential. Entropy. 2022; 24(11):1688. https://0-doi-org.brum.beds.ac.uk/10.3390/e24111688
Chicago/Turabian StyleAbbas Abdulhussein, Msallam, Zaid Abdi Alkareem Alyasseri, Husam Jasim Mohammed, and Xingwei An. 2022. "Lack of Habituation in Migraine Patients Based on High-Density EEG Analysis Using the Steady State of Visual Evoked Potential" Entropy 24, no. 11: 1688. https://0-doi-org.brum.beds.ac.uk/10.3390/e24111688