
Epigallocatechin-3-Gallate Prevents Acute Gout by Suppressing NLRP3 Inflammasome Activation and Mitochondrial DNA Synthesis

 ,
, 
 and
and 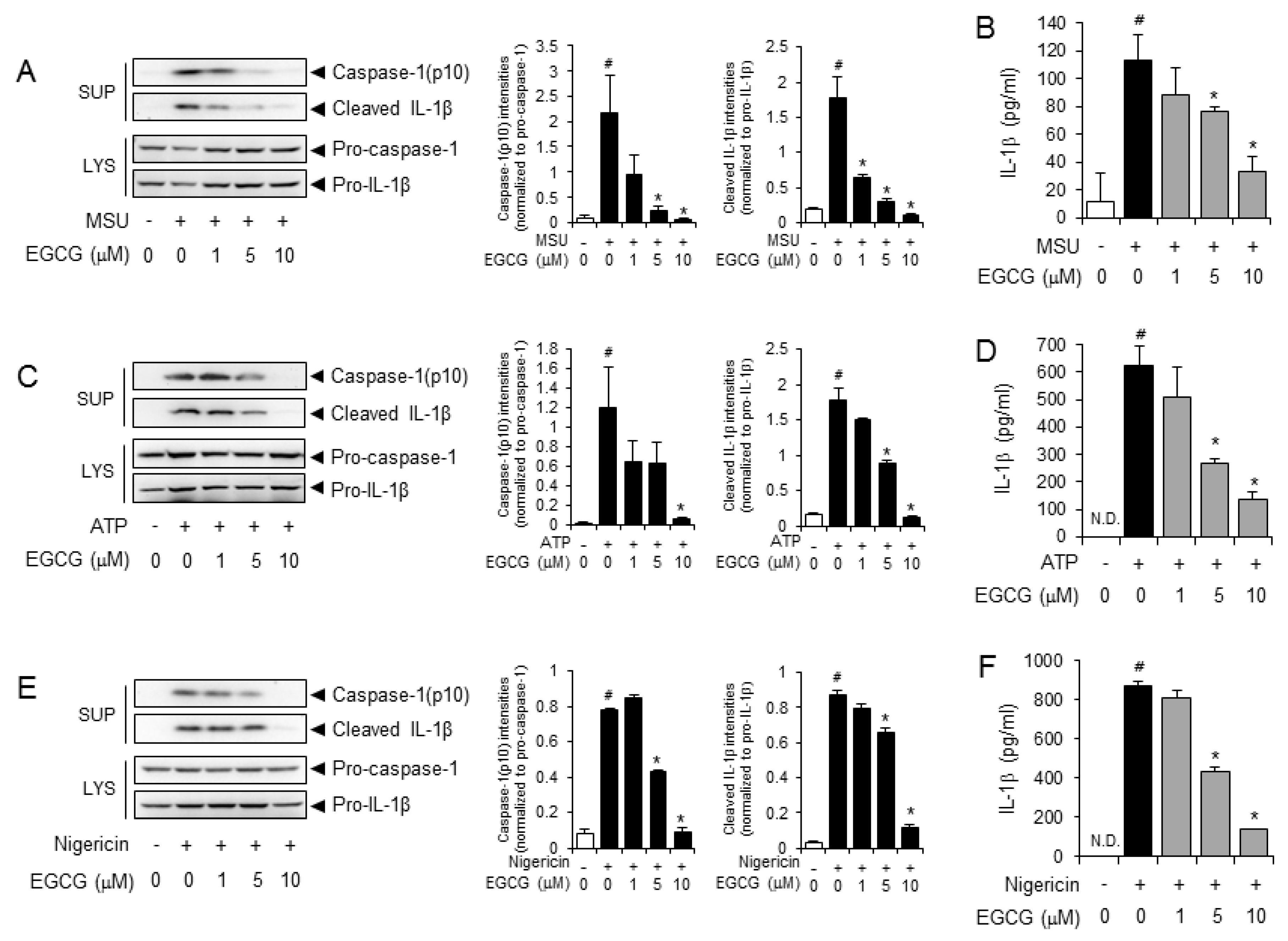{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. EGCG Suppresses NLRP3 Inflammasome Activation in Primary Macrophages
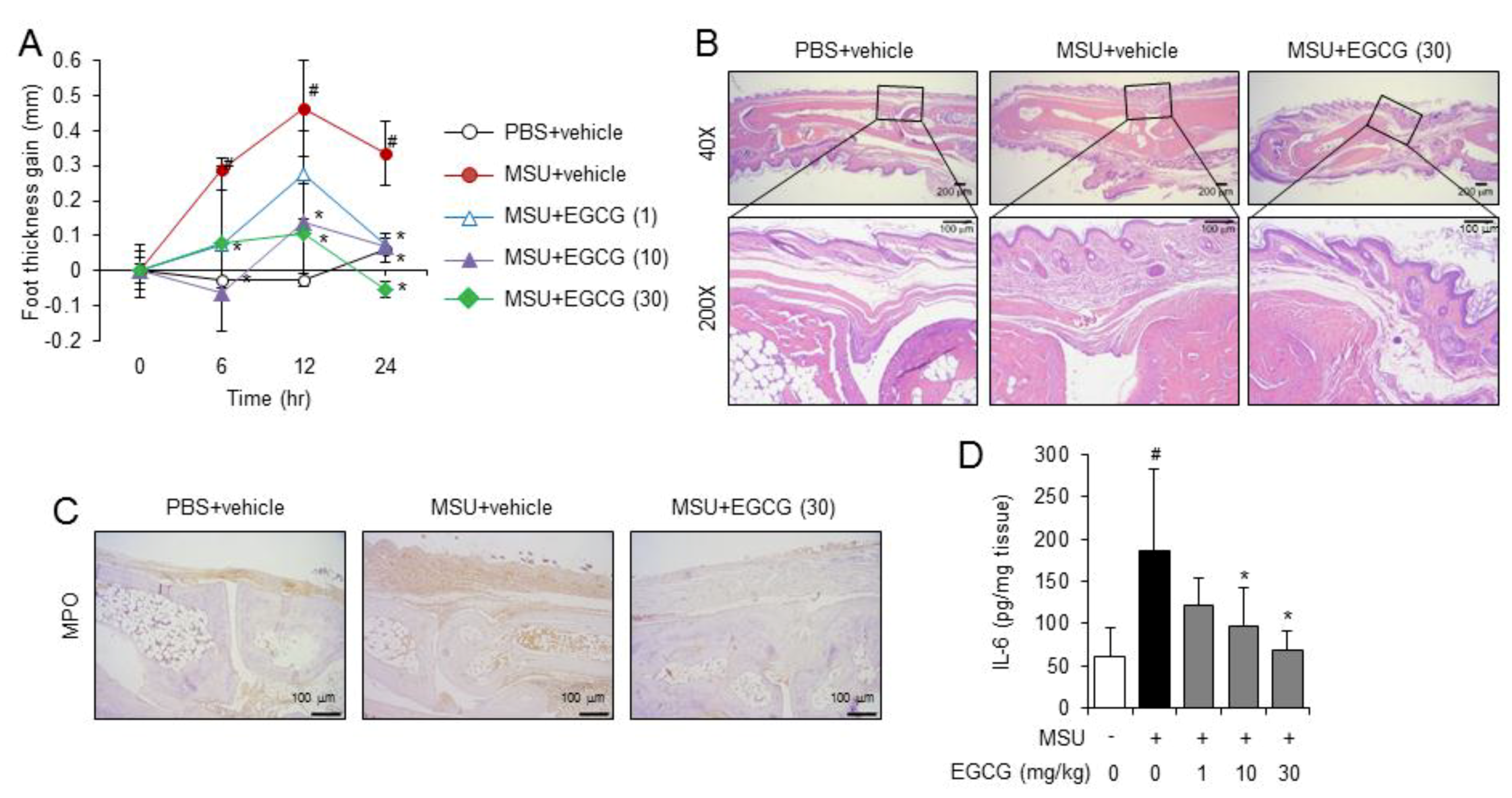
2.2. Oral Administration of EGCG Prevents Acute Gouty Inflammation in Mice by Blocking NLRP3 Inflammasome Activation
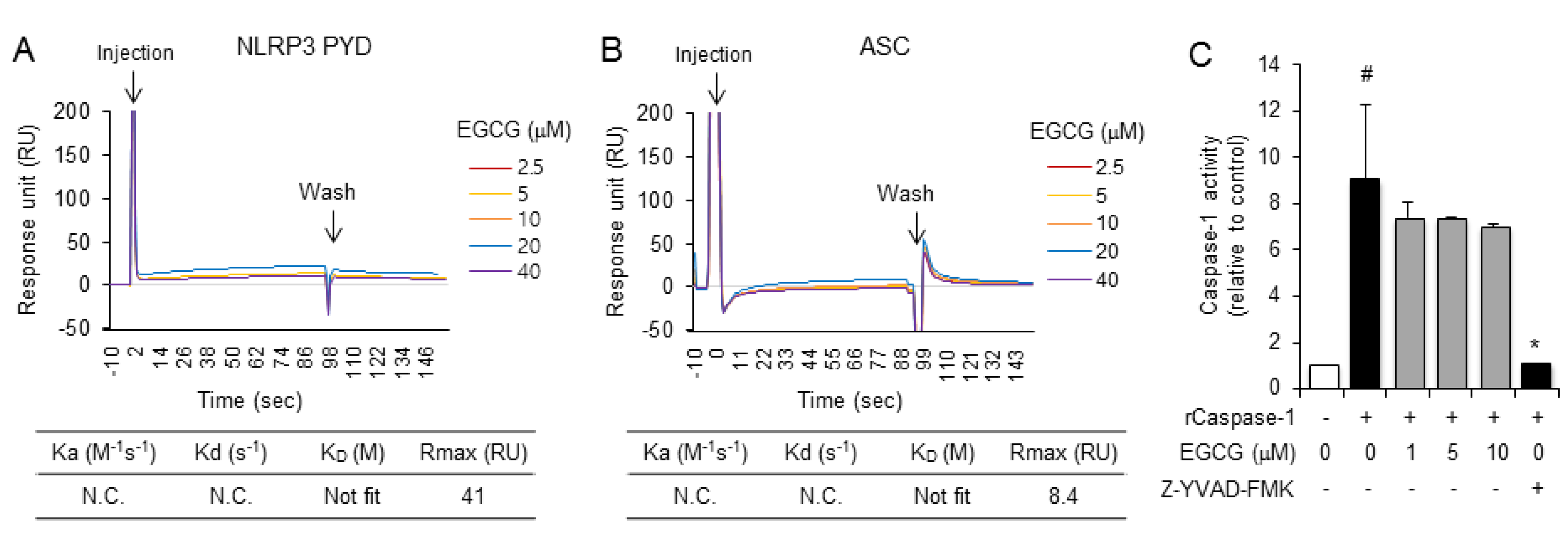
2.3. EGCG Inhibits the De Novo Synthesis of Mitochondrial DNA in Macrophages
3. Discussion
4. Materials and Methods
4.1. Animals and Cell Culture
4.2. Reagents
4.3. Analysis of Inflammasome Activation
4.4. Enzyme-Linked Immunosorbent Assays
4.5. An Acute Gout Mouse Model
4.6. Caspase-1 Enzyme Activity Assay
4.7. Reactive Oxygen Species (ROS) Measurement
4.8. Quantitative Real-Time Polymerase Chain Reaction (qPCR) Analysis for Total Mitochondrial DNA
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Wallace, K.L.; Riedel, A.A.; Joseph-Ridge, N.; Wortmann, R. Increasing prevalence of gout and hyperuricemia over 10 years among older adults in a managed care population. J. Rheumatol. 2004, 31, 1582–1587. [Google Scholar] [PubMed]
- Shi, Y.; Mucsi, A.D.; Ng, G. Monosodium urate crystals in inflammation and immunity. Immunol. Rev. 2010, 233, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Petrilli, V.; Mayor, A.; Tardivel, A.; Tschopp, J. Gout-associated uric acid crystals activate the NALP3 inflammasome. Nature 2006, 440, 237–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinon, F.; Burns, K.; Tschopp, J. The inflammasome: A molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol. Cell 2002, 10, 417–426. [Google Scholar] [CrossRef]
- Lee, H.E.; Yang, G.; Kim, N.D.; Jeong, S.; Jung, Y.; Choi, J.Y.; Park, H.H.; Lee, J.Y. Targeting ASC in NLRP3 inflammasome by caffeic acid phenethyl ester: A novel strategy to treat acute gout. Sci. Rep. 2016, 6, 38622. [Google Scholar] [CrossRef]
- McGonagle, D.; Tan, A.L.; Shankaranarayana, S.; Madden, J.; Emery, P.; McDermott, M.F. Management of treatment resistant inflammation of acute on chronic tophaceous gout with anakinra. Ann. Rheum. Dis. 2007, 66, 1683–1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlesinger, N.; Mysler, E.; Lin, H.Y.; De Meulemeester, M.; Rovensky, J.; Arulmani, U.; Balfour, A.; Krammer, G.; Sallstig, P.; So, A. Canakinumab reduces the risk of acute gouty arthritis flares during initiation of allopurinol treatment: Results of a double-blind, randomised study. Ann. Rheum. Dis. 2011, 70, 1264–1271. [Google Scholar] [CrossRef]
- Schumacher, H.R., Jr.; Sundy, J.S.; Terkeltaub, R.; Knapp, H.R.; Mellis, S.J.; Stahl, N.; Yancopoulos, G.D.; Soo, Y.; King-Davis, S.; Weinstein, S.P.; et al. Rilonacept (interleukin-1 trap) in the prevention of acute gout flares during initiation of urate-lowering therapy: Results of a phase II randomized, double-blind, placebo-controlled trial. Arthritis Rheum. 2012, 64, 876–884. [Google Scholar] [CrossRef]
- Schlesinger, N. Anti-interleukin-1 therapy in the management of gout. Curr. Rheumatol. Rep. 2014, 16, 398. [Google Scholar] [CrossRef]
- Tsai, P.Y.; Ka, S.M.; Chang, J.M.; Chen, H.C.; Shui, H.A.; Li, C.Y.; Hua, K.F.; Chang, W.L.; Huang, J.J.; Yang, S.S.; et al. Epigallocatechin-3-gallate prevents lupus nephritis development in mice via enhancing the Nrf2 antioxidant pathway and inhibiting NLRP3 inflammasome activation. Free Radic. Biol. Med. 2011, 51, 744–754. [Google Scholar] [CrossRef]
- Gao, Z.; Han, Y.; Hu, Y.; Wu, X.; Wang, Y.; Zhang, X.; Fu, J.; Zou, X.; Zhang, J.; Chen, X.; et al. Targeting HO-1 by Epigallocatechin-3-Gallate Reduces Contrast-Induced Renal Injury via Anti-Oxidative Stress and Anti-Inflammation Pathways. PLoS ONE 2016, 11, e0149032. [Google Scholar] [CrossRef] [PubMed]
- Jo, E.K.; Kim, J.K.; Shin, D.M.; Sasakawa, C. Molecular mechanisms regulating NLRP3 inflammasome activation. Cell. Mol. Immunol. 2016, 13, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Liang, S.; Sanchez-Lopez, E.; He, F.; Shalapour, S.; Lin, X.J.; Wong, J.; Ding, S.; Seki, E.; Schnabl, B.; et al. New mitochondrial DNA synthesis enables NLRP3 inflammasome activation. Nature 2018, 560, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Jhang, J.J.; Lu, C.C.; Yen, G.C. Epigallocatechin gallate inhibits urate crystals-induced peritoneal inflammation in C57BL/6 mice. Mol. Nutr. Food Res. 2016, 60, 2297–2303. [Google Scholar] [CrossRef] [PubMed]
- L’Allemain, G. Multiple actions of EGCG, the main component of green tea. Bulletin du Cancer 1999, 86, 721–724. [Google Scholar] [PubMed]
- Martinon, F. Mechanisms of uric acid crystal-mediated autoinflammation. Immunol. Rev. 2010, 233, 218–232. [Google Scholar] [CrossRef] [PubMed]
- Shimada, K.; Crother, T.R.; Karlin, J.; Dagvadorj, J.; Chiba, N.; Chen, S.; Ramanujan, V.K.; Wolf, A.J.; Vergnes, L.; Ojcius, D.M.; et al. Oxidized mitochondrial DNA activates the NLRP3 inflammasome during apoptosis. Immunity 2012, 36, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Joung, S.M.; Park, Z.Y.; Rani, S.; Takeuchi, O.; Akira, S.; Lee, J.Y. Akt contributes to activation of the TRIF-dependent signaling pathways of TLRs by interacting with TANK-binding kinase 1. J. Immunol. 2011, 186, 499–507. [Google Scholar] [CrossRef]
- Yang, G.; Oh, J.W.; Lee, H.E.; Lee, B.H.; Lim, K.M.; Lee, J.Y. Topical Application of Dieckol Ameliorates Atopic Dermatitis in NC/Nga Mice by Suppressing Thymic Stromal Lymphopoietin Production. J. Investig. Dermatol. 2016, 136, 1062–1066. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Lee, H.E.; Lee, J.Y. A pharmacological inhibitor of NLRP3 inflammasome prevents non-alcoholic fatty liver disease in a mouse model induced by high fat diet. Sci. Rep. 2016, 6, 24399. [Google Scholar] [CrossRef]
- Yeon, S.H.; Yang, G.; Lee, H.E.; Lee, J.Y. Oxidized phosphatidylcholine induces the activation of NLRP3 inflammasome in macrophages. J. Leukoc. Biol. 2017, 101, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Lee, H.E.; Shin, S.W.; Um, S.H.; Lee, J.D.; Kim, K.B.; Kang, H.C.; Cho, Y.Y.; Lee, H.S.; Lee, J.Y. Efficient Transdermal Delivery of DNA Nanostructures Alleviates Atopic Dermatitis Symptoms in NC/Nga Mice. Adv. Funct. Mater. 2018, 28, 1801918. [Google Scholar] [CrossRef]
Sample Availability: Not available. |





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.E.; Yang, G.; Park, Y.B.; Kang, H.C.; Cho, Y.-Y.; Lee, H.S.; Lee, J.Y. Epigallocatechin-3-Gallate Prevents Acute Gout by Suppressing NLRP3 Inflammasome Activation and Mitochondrial DNA Synthesis. Molecules 2019, 24, 2138. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24112138
Lee HE, Yang G, Park YB, Kang HC, Cho Y-Y, Lee HS, Lee JY. Epigallocatechin-3-Gallate Prevents Acute Gout by Suppressing NLRP3 Inflammasome Activation and Mitochondrial DNA Synthesis. Molecules. 2019; 24(11):2138. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24112138
Chicago/Turabian StyleLee, Hye Eun, Gabsik Yang, Youn Bum Park, Han Chang Kang, Yong-Yeon Cho, Hye Suk Lee, and Joo Young Lee. 2019. "Epigallocatechin-3-Gallate Prevents Acute Gout by Suppressing NLRP3 Inflammasome Activation and Mitochondrial DNA Synthesis" Molecules 24, no. 11: 2138. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24112138