Identification of Constituents Affecting the Secretion of Pro-Inflammatory Cytokines in LPS-Induced U937 Cells by UHPLC-HRMS-Based Metabolic Profiling of the Traditional Chinese Medicine Formulation Huangqi Jianzhong Tang


Abstract
:1. Introduction
2. Results
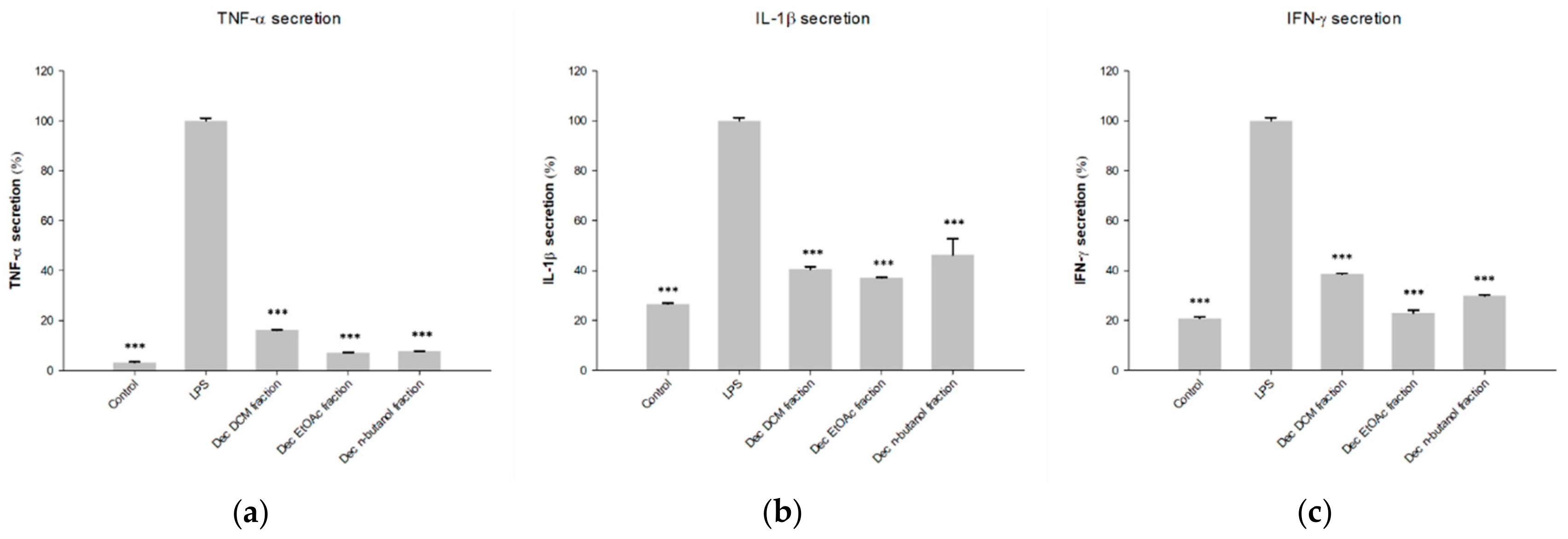
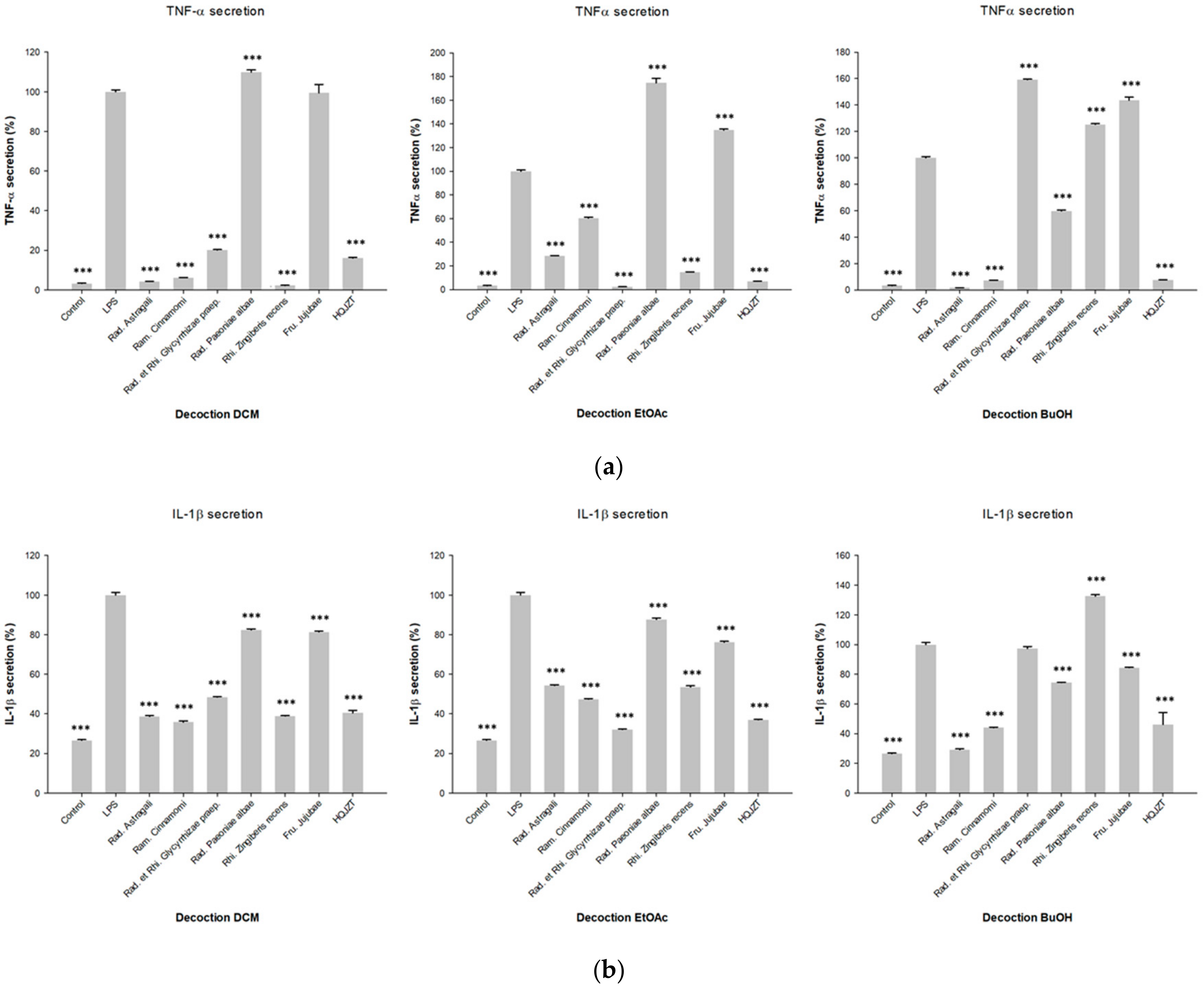
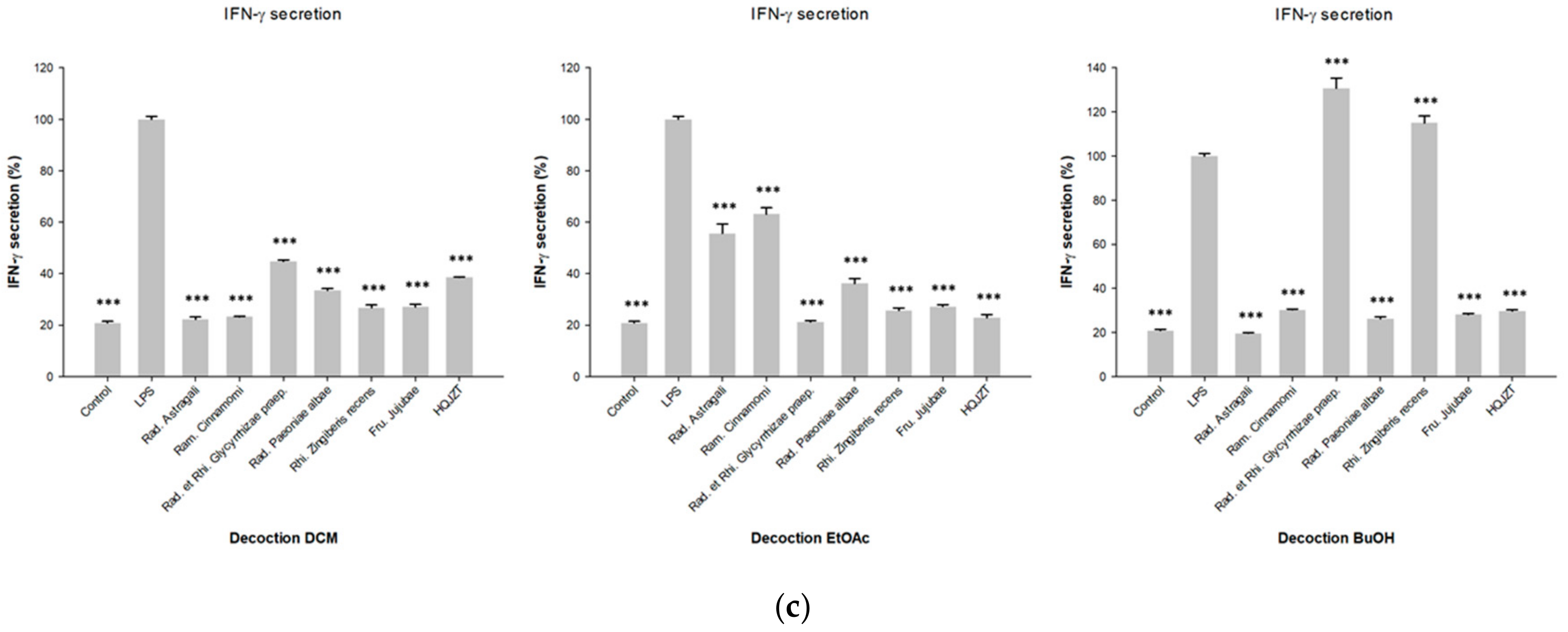
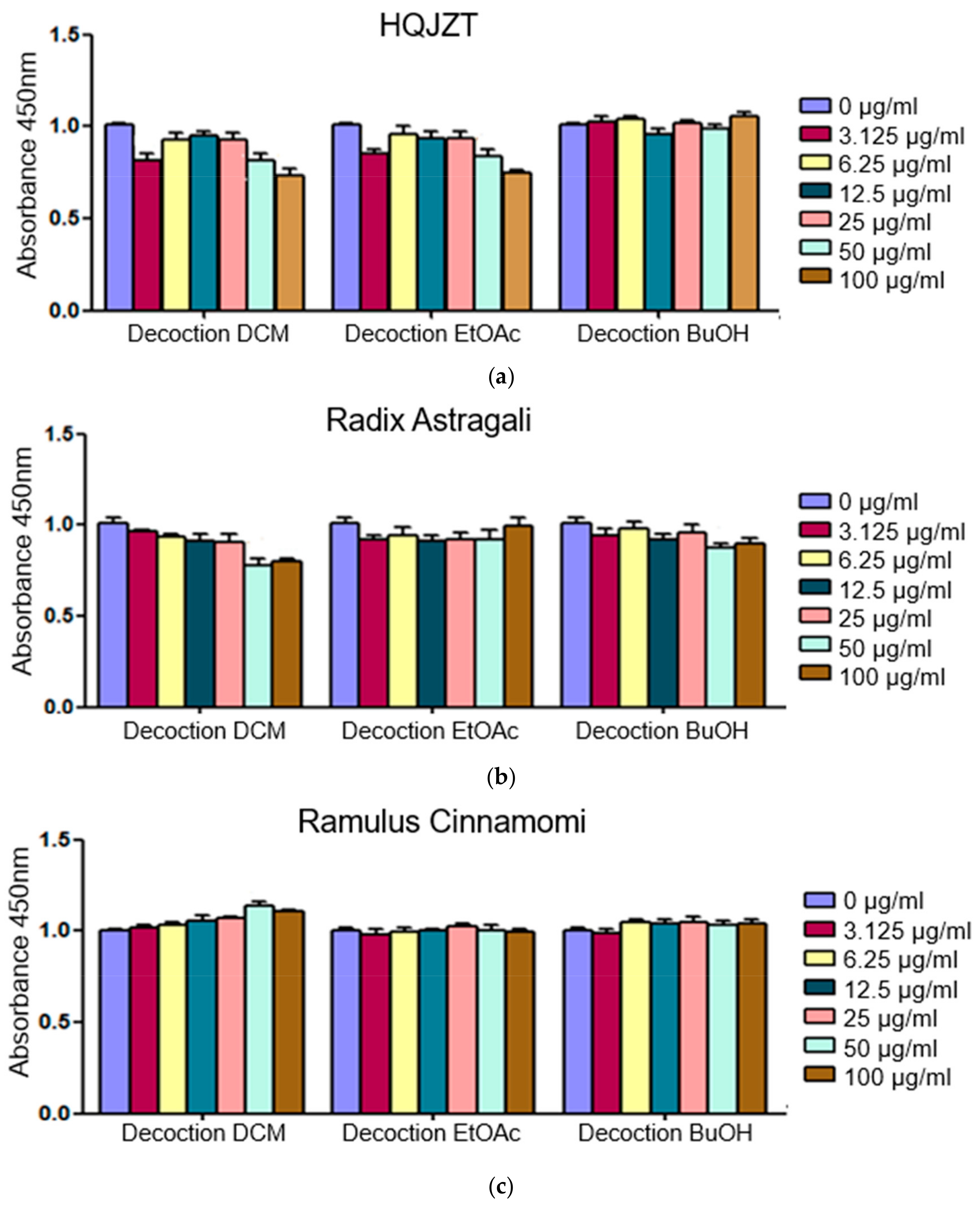
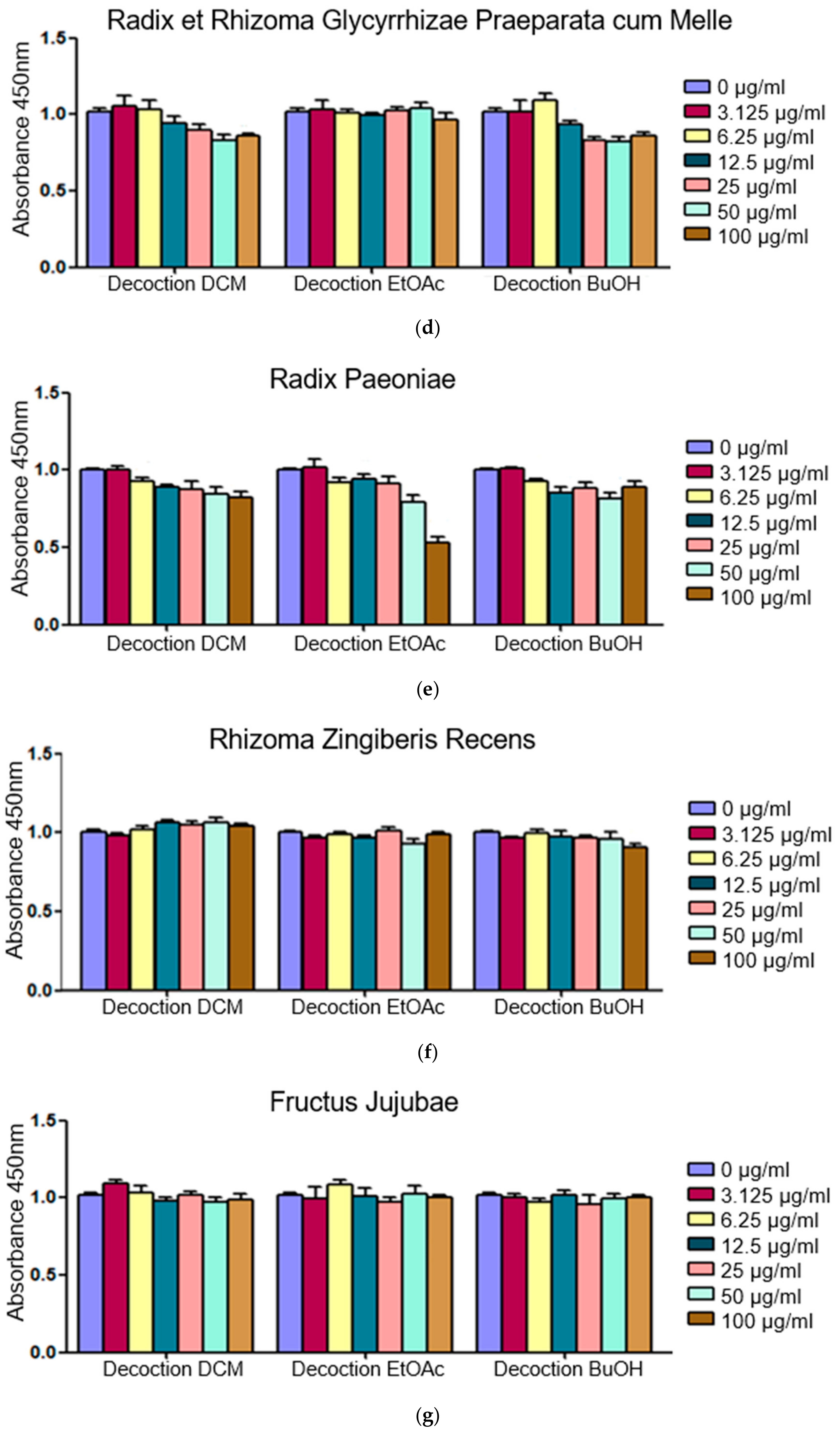
2.1. Fractions of HQZT Decoction and Its Single Herbs Exert Distinct Effects on Pro-Inflammatory Cytokines
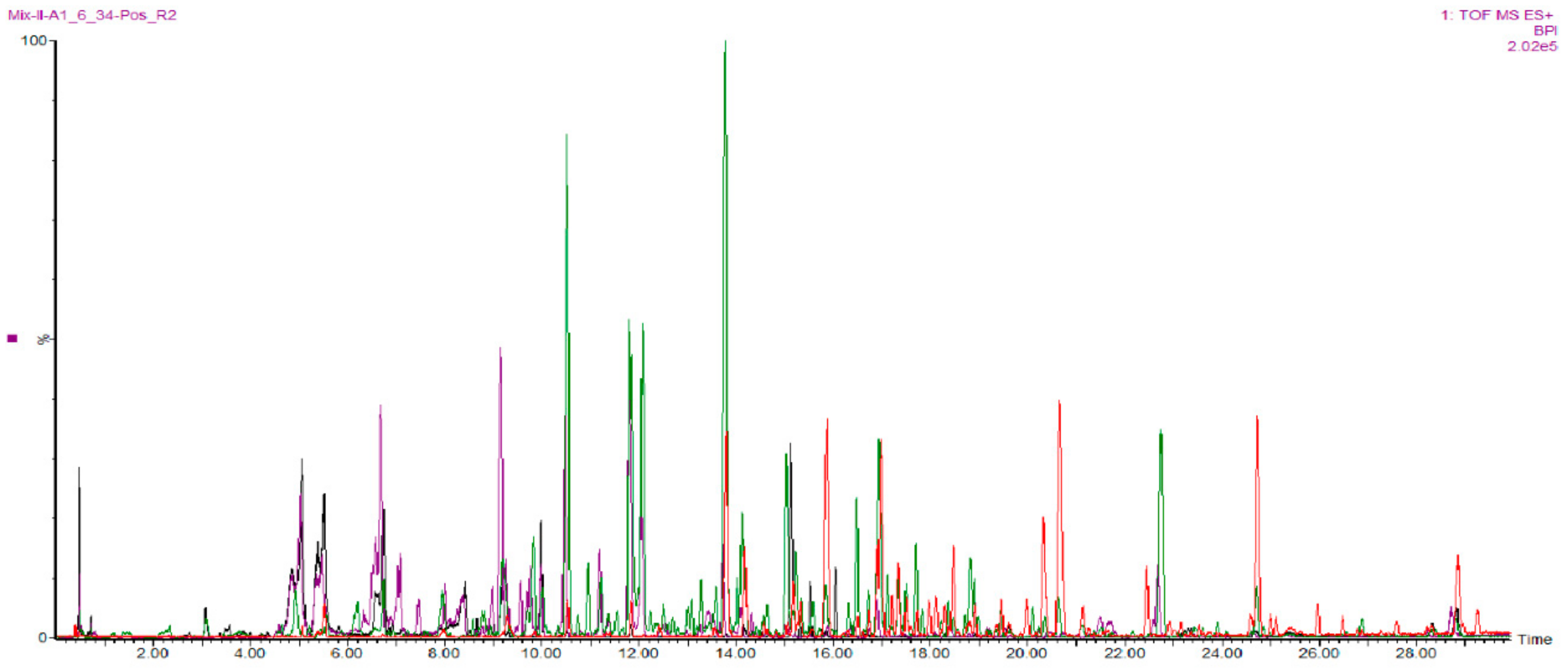
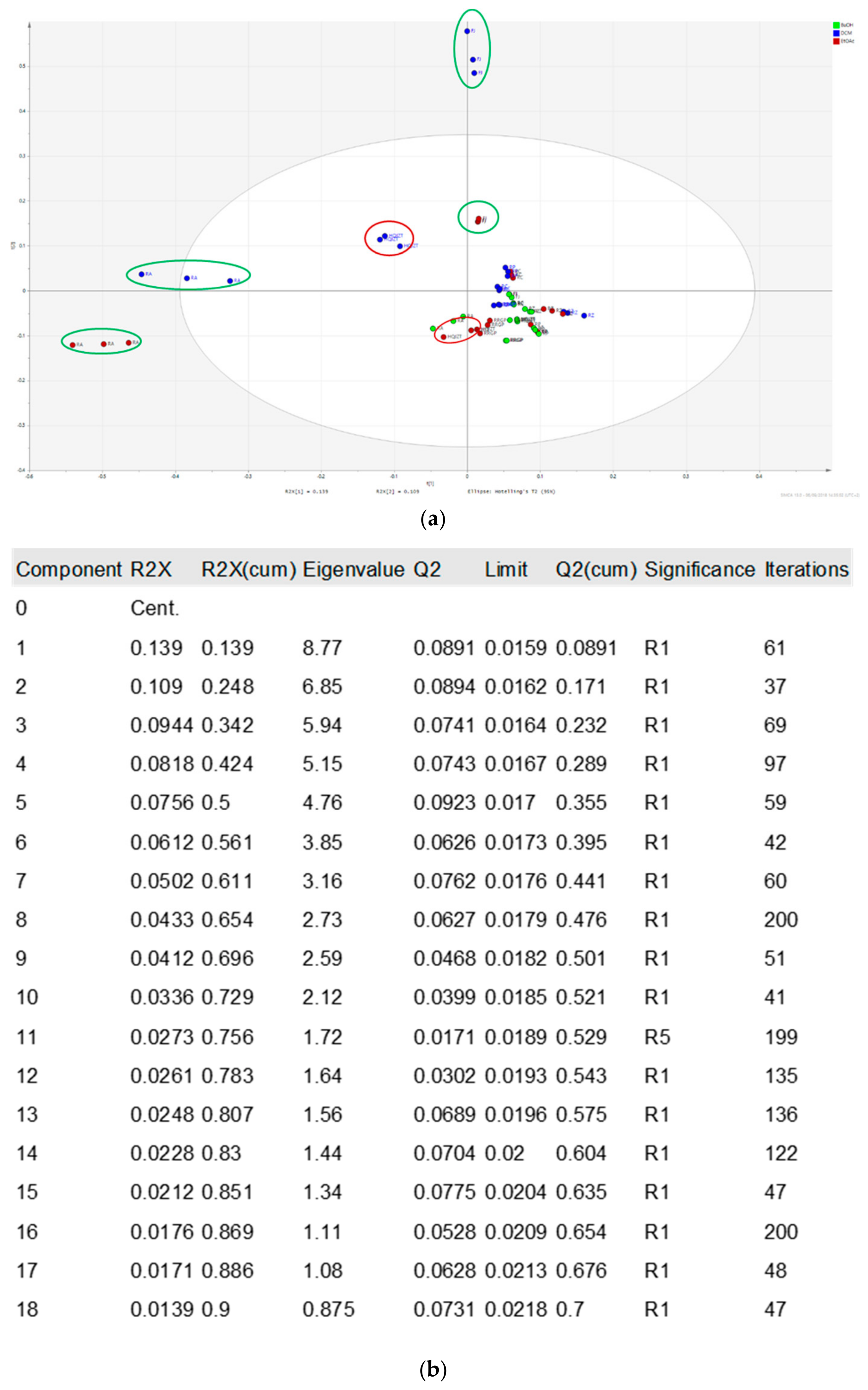
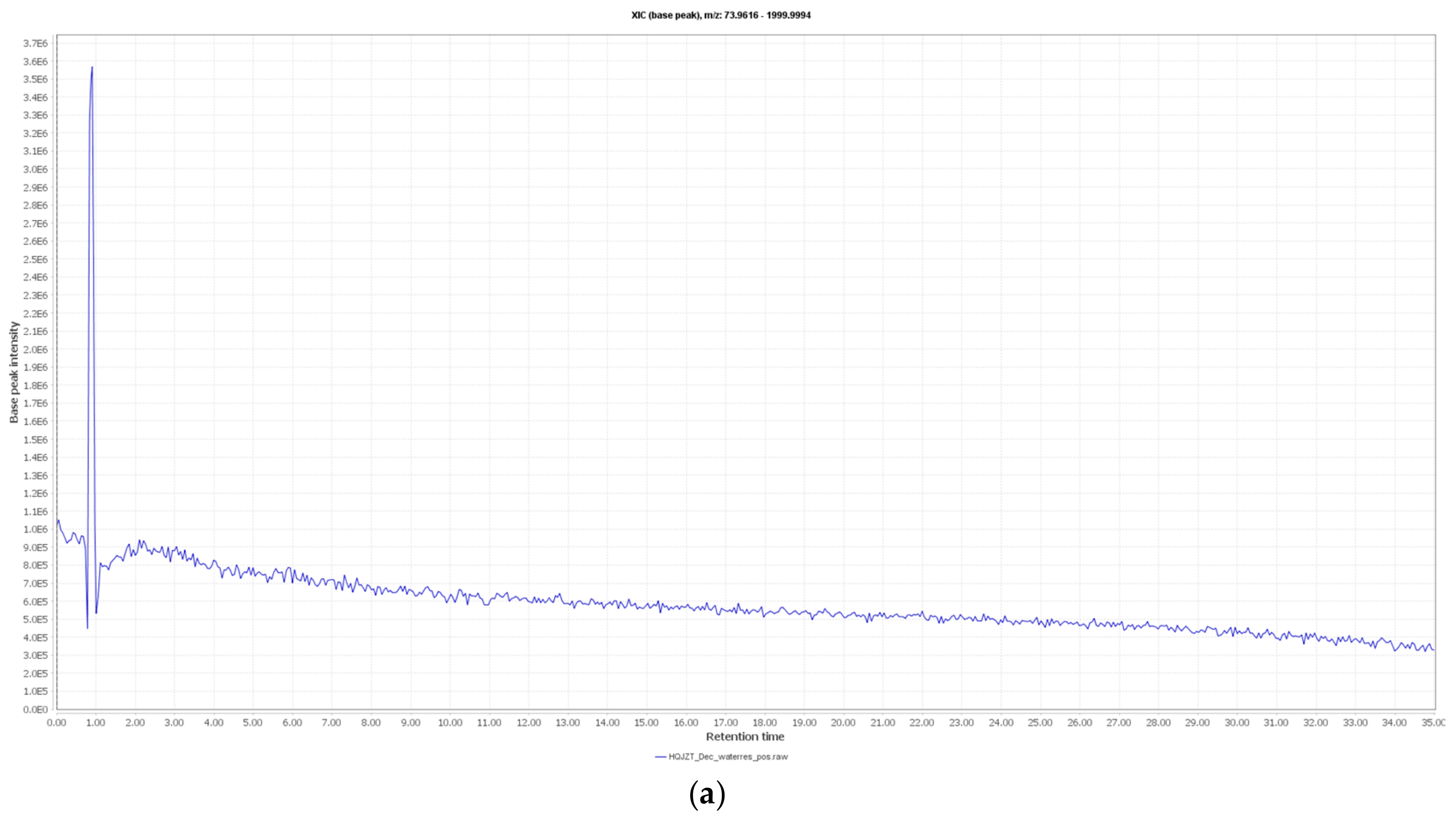
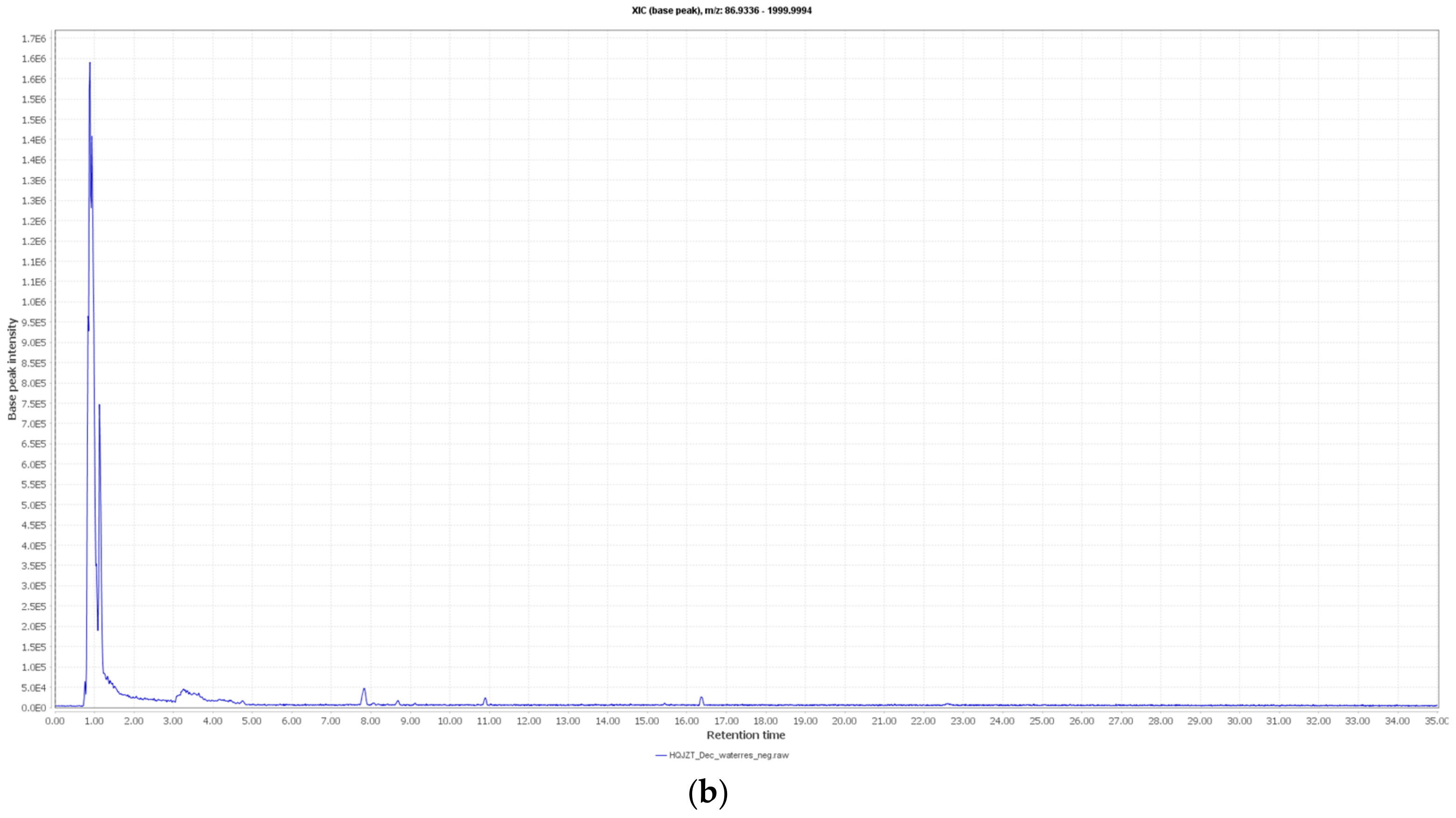
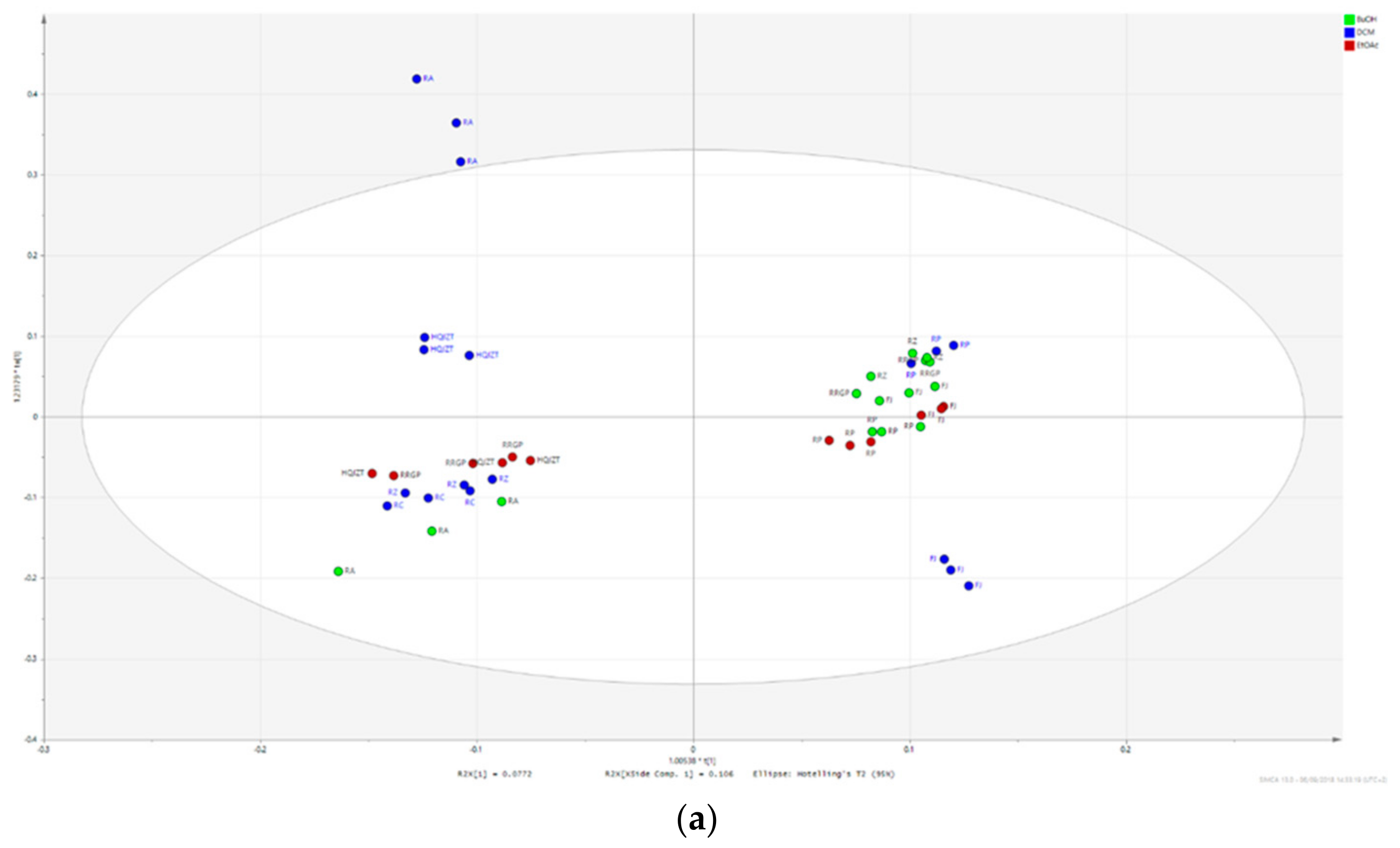
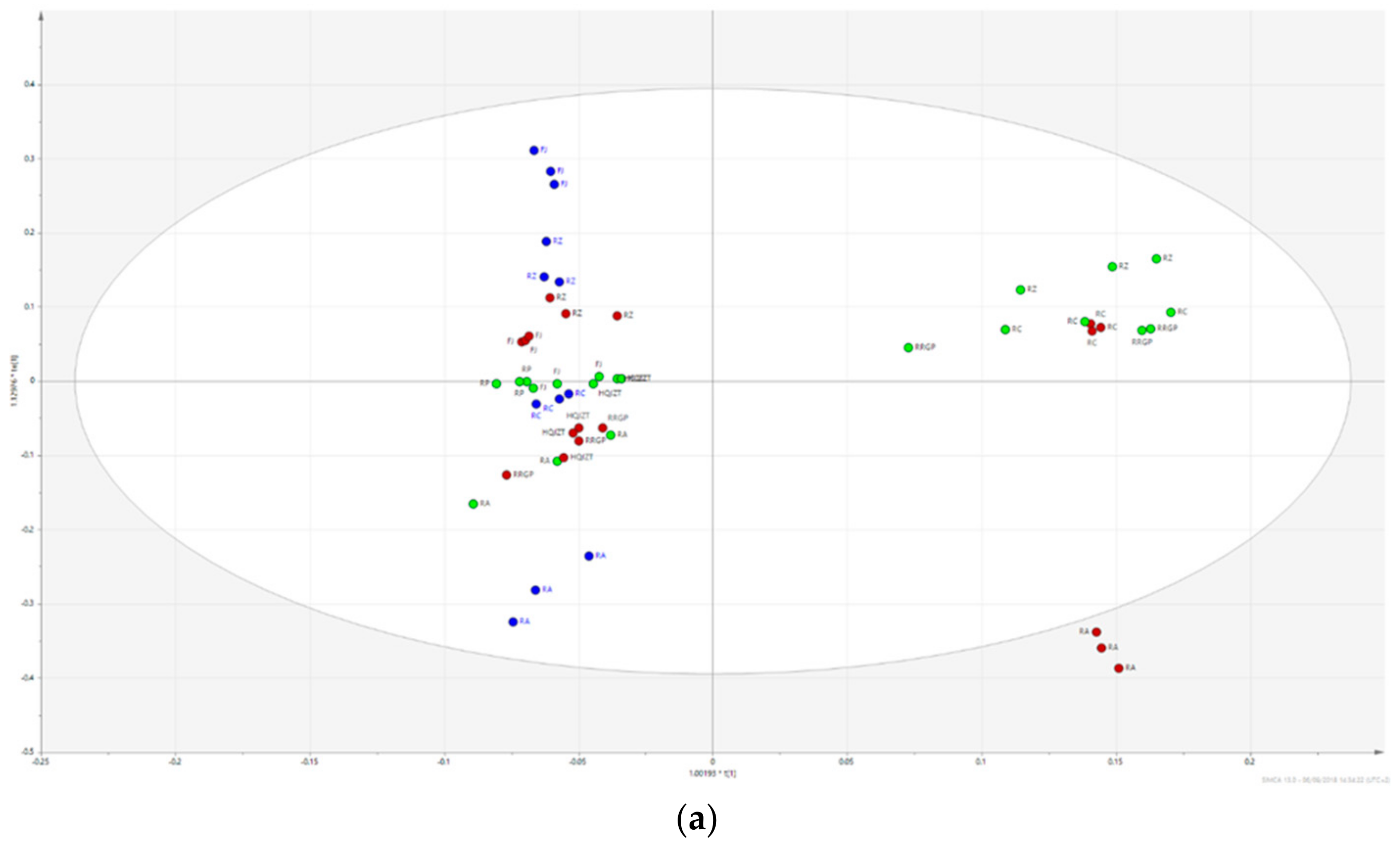
2.2. Fractions of HQZT Decoction and Its Single Herbs Possess Distinct UPLC-HRMS Profiles
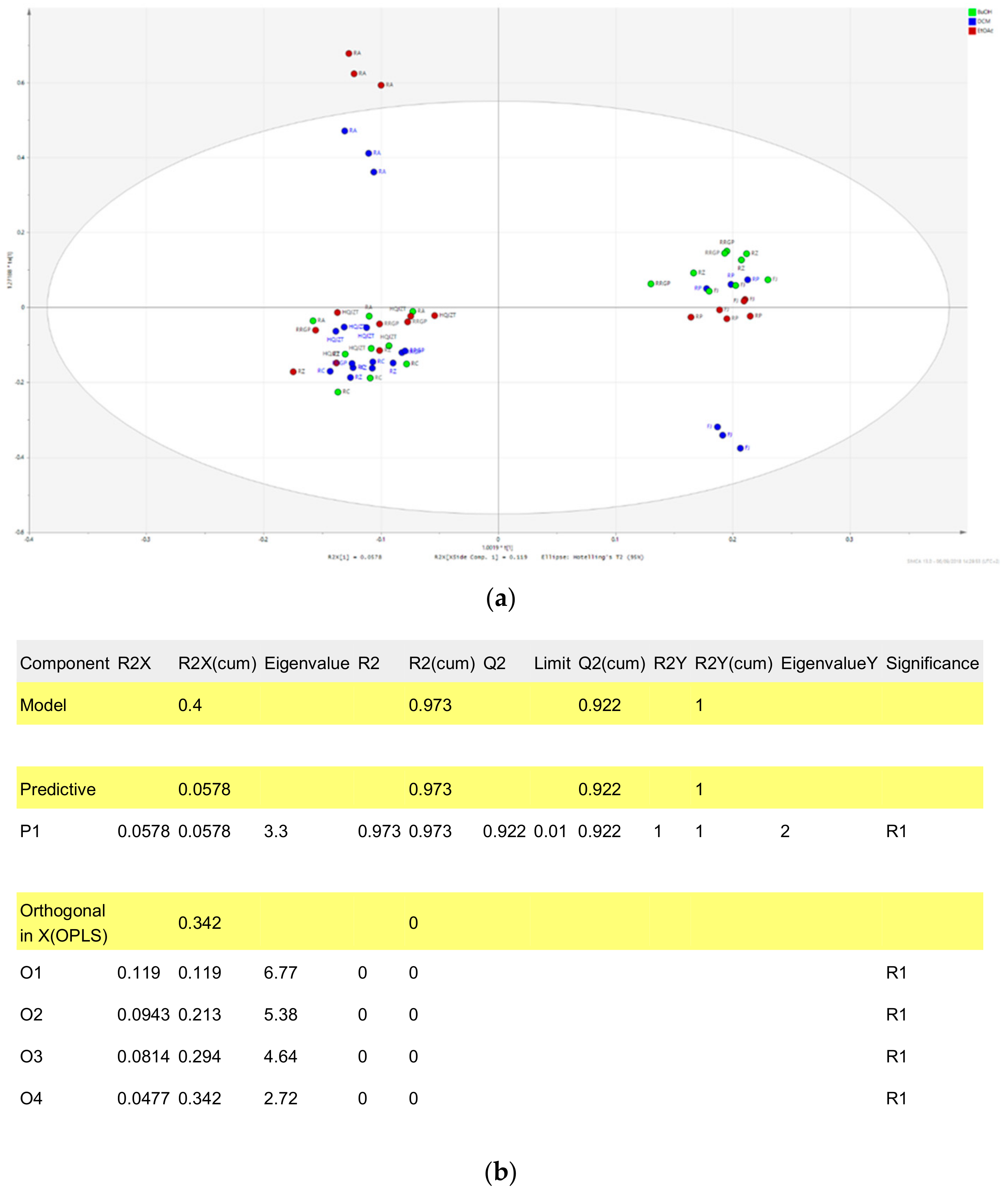
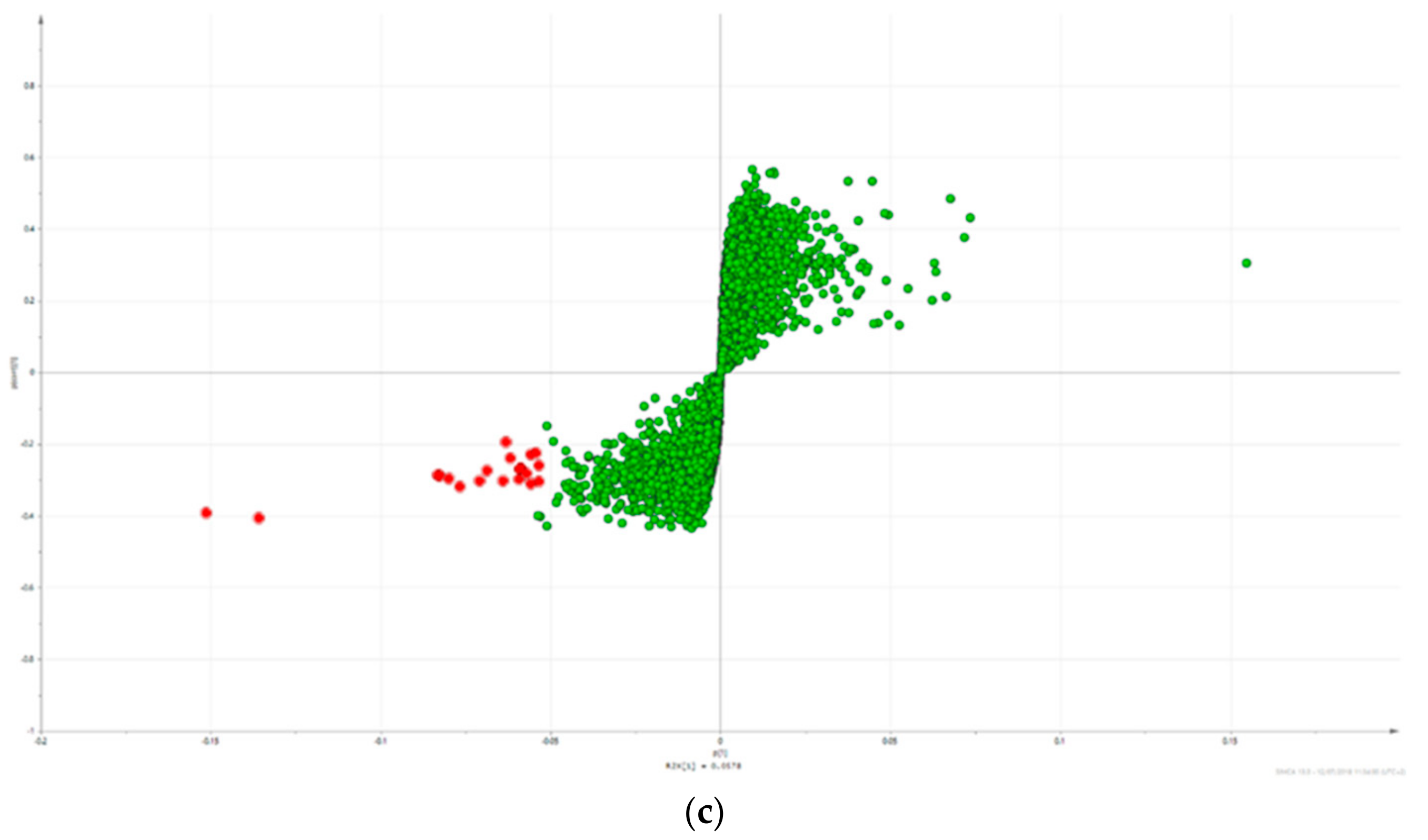
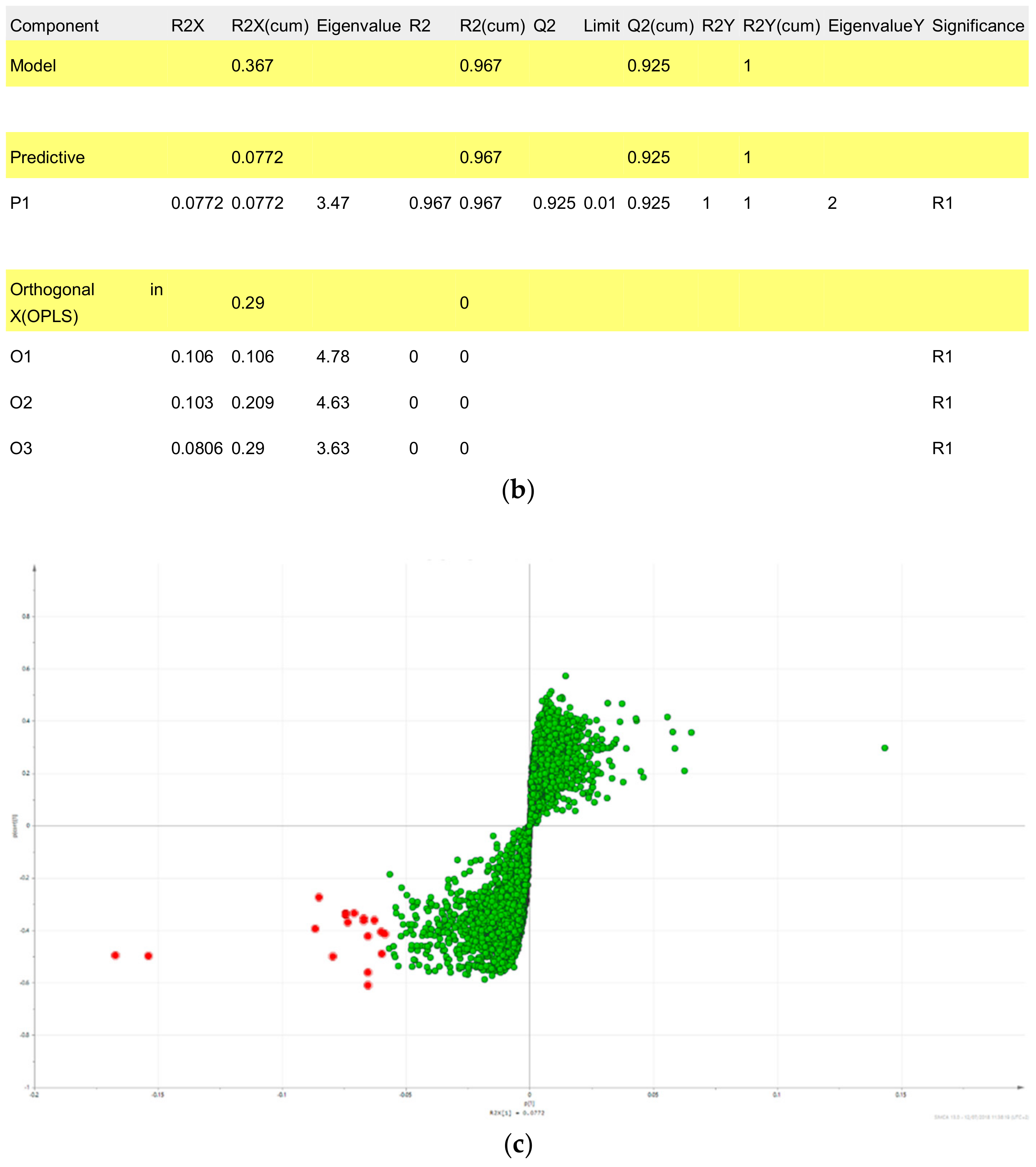
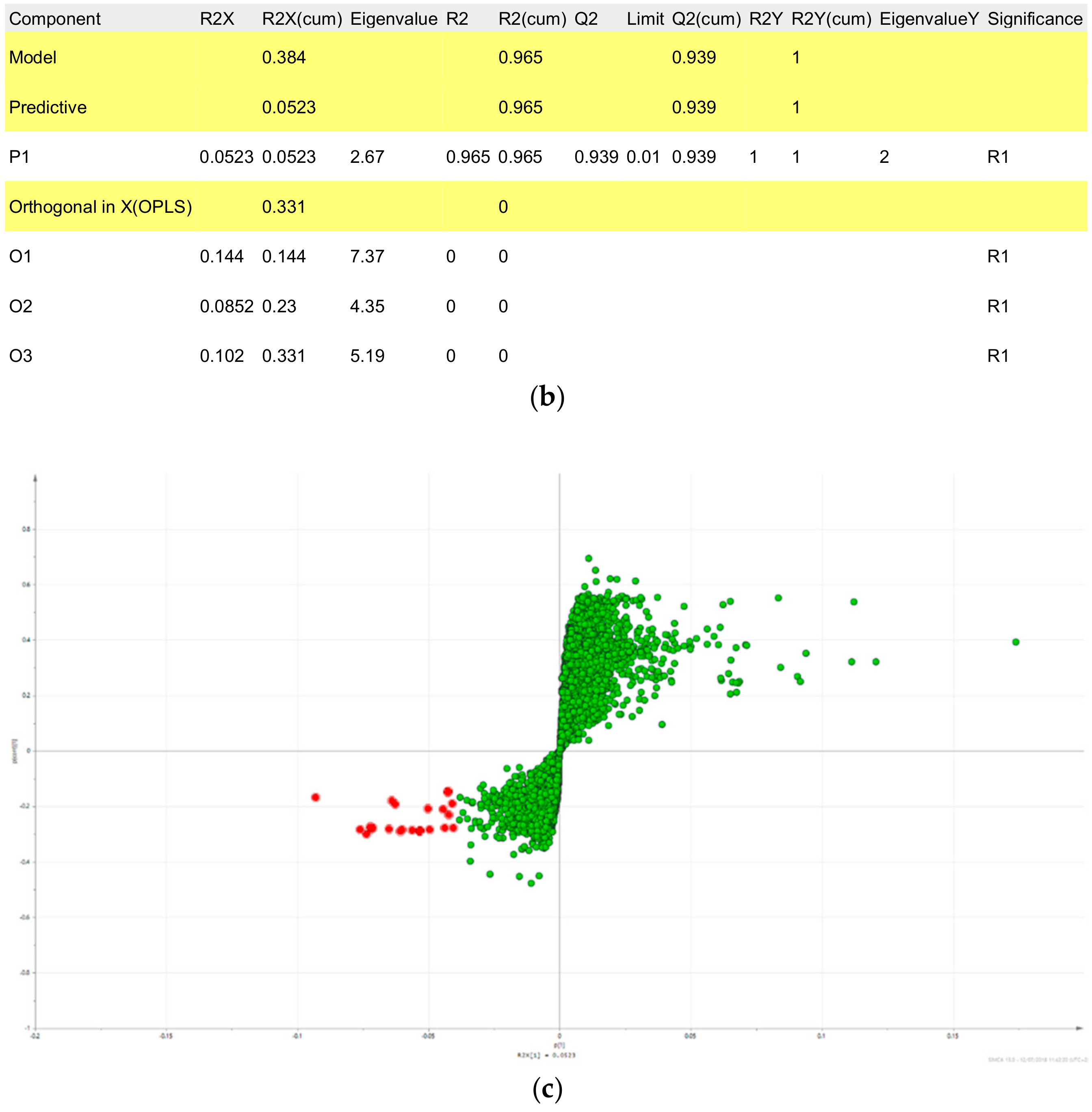
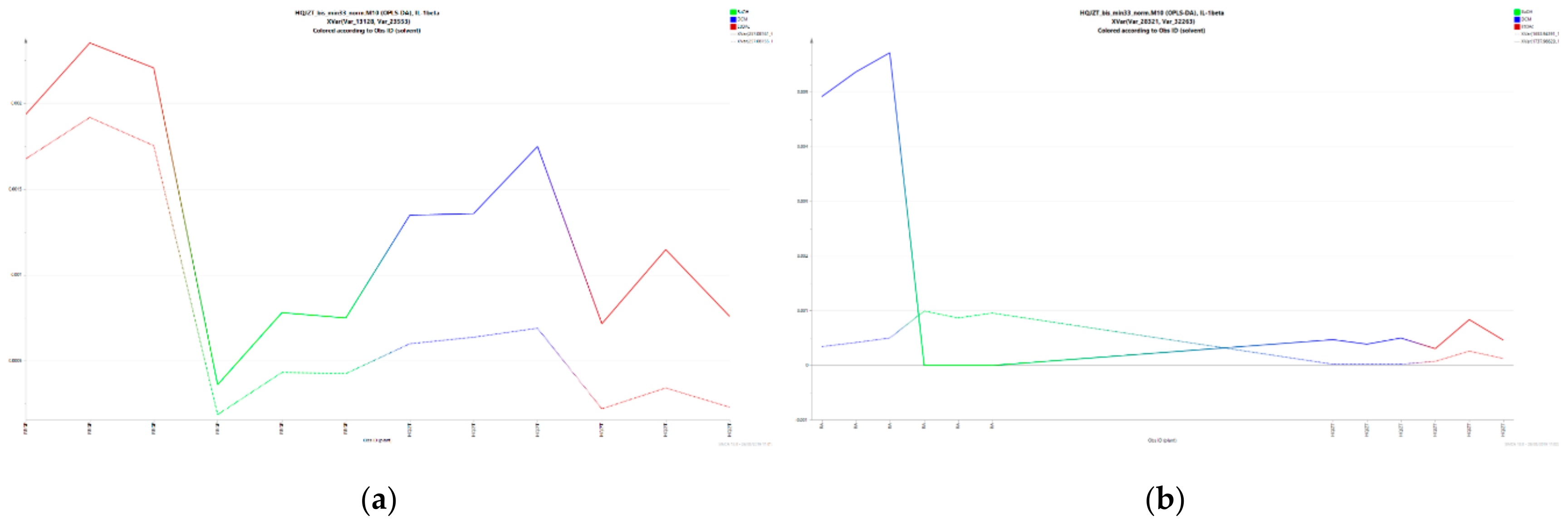
2.3. Triterpene Glycosides, Flavonoids and Monoterpene Glycosides are Most Strongly Correlated with Inhibitory Activity on Production of Pro-Inflammatory Cytokines
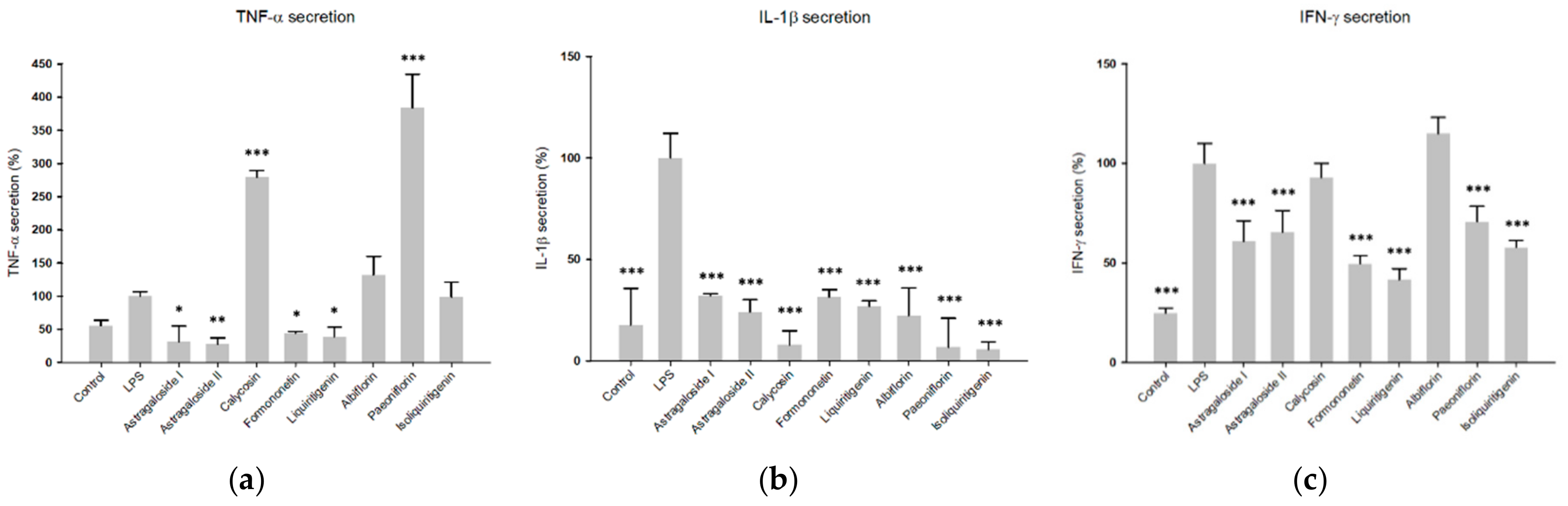
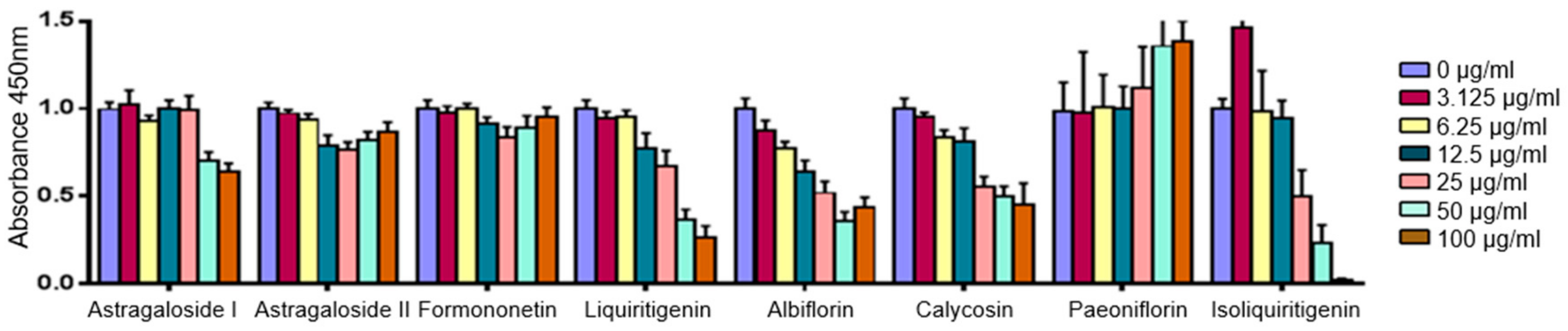
2.4. Predicted Pharmacological Activities Can Be Partially Verified for the Identified OPLS-DA Hit Compounds
3. Discussion
4. Materials and Methods
4.1. Plant Material, Chemicals and Reagents
4.2. Sample Preparation
4.3. UPLC-MS Analysis
4.4. Pharmalogical Studies
4.5. Data Processing and Multivariate Data Analysis
4.6. Metabolite Identification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant | Initial Weight Crude Drug (g) | Yield n-Hexane Extract (mg) | Yield DCM Extract (mg) | Yield EtOAc Extract (mg) | Yield n-Butanol Extract (mg) | Yield Water Residue (g) |
|---|---|---|---|---|---|---|
| Huangqin Jianzhong Tang | 45 | 34.1 | 63.6 | 209.0 | 962.9 | 7.8 |
| Radix Astragali | 45 | 12.3 | 93.6 | 104.6 | 803.6 | 13.7 |
| Ramulus Cinnamomi | 20 | 33.7 | 56.1 | 51.3 | 161.6 | 1.0 |
| Radix et Rhizoma Glycyrrhizae praeparata cum melle | 20 | 22.4 | 46.0 | 147.3 | 609.2 | 6.8 |
| Fructus Jujubae | 45 | 19.4 | 28.2 | 45.1 | 178.9 | 8.6 |
| Radix Paeoniae albae | 20 | 19.8 | 43.2 | 245.9 | 723.7 | 3.7 |
| Rhizoma Zingiberis recens | 45 | 39.3 | 17.0 | 6.3 | 55.4 | 1.0 |






| m/z_RT | p[1] | po[1] | Origin | Compound | Fragment Ions | Literature |
|---|---|---|---|---|---|---|
| 285.07656_10.514 | −0.151342 | 0.247407 | RA | Calycosin * | 253, 225, 137 | [24,25] |
| 269.08165_13.771 | −0.135887 | 0.108774 | RA | Formononetin * | 269, 253, 237, 226, 197, 181, 169, 137, 118 | Metlin, [26] |
| 1737.96629_18.398 | −0.0831685 | 0.219044 | RA | Astragaloside I * (dimer) | 689, 671, 653, 491, 473, 455, 437, 419, 297 | [24,26] |
| 1737.96335_19.162 | −0.0827036 | 0.216071 | RA | Astragaloside I isomer (dimer) | 689, 671, 653, 491, 473, 455, 437, 419, 297 | [24,26] |
| 355.11759_16.953 | −0.0798714 | −0.0373342 | RRGP | n.i. | ||
| 301.10751_10.011 | −0.0767608 | 0.176659 | RA | 3-Hydroxy-9,10-dimethoxypterocarpan or isomer | 269, 191, 167, 152, 133, 147, 123, 105 | [25] |
| 371.10109_32.627 | −0.0708791 | 0.069922 | All | n.i. | ||
| 447.12852_6.654 | −0.0687443 | 0.132943 | RA | Calycosin-7-O-β-d-glucoside or isomer | 270, 253, 225, 137 | [24] |
| 355.06981_32.607 | −0.0640716 | 0.124033 | RA | n.i. | ||
| 714.41099_18.603 | −0.0632131 | −0.0573972 | RZ | n.i. | ||
| 411.22785_13.841 | −0.0617407 | −0.0464029 | RZ | n.i. | ||
| 508.25367_16.894 | −0.0592386 | −0.0425658 | RZ | n.i. | ||
| 438.23858_13.465 | −0.0591264 | −0.0471358 | RZ | n.i. | ||
| 447.12854_6.484 | −0.0589284 | 0.116497 | RA | Calycosin-7-O-β-d-glucoside or isomer | 270, 253, 225, 137 | [24] |
| 1737.96324_20.222 | −0.0571481 | 0.146308 | RA | Astragaloside I isomer (dimer) | 851, 833, 689, 671, 653, 491, 473, 455, 437, 419, 297 | [24,26] |
| 1653.94391_15.879 | −0.0558838 | 0.154538 | RA | Astragaloside II or isomer (dimer) | 647, 629, 491, 473, 455, 437, 419 | [24,26] |
| 353.10204_16.488 | −0.0557885 | −0.0258396 | RRGP | n.i. | ||
| 471.34640_24.718 | −0.0542538 | −0.0348534 | RRGP | 18β-Glycyrrhetinic acid * | 453, 235, 217, 189, 175189 | Metlin |
| 301.10773_14.151 | −0.0535515 | 0.0891372 | RA | 3-Hydroxy-9,10-dimethoxypterocarpan or isomer | 269, 191, 167, 152, 147, 133, 123, 105 | [25] |
| 257.08161_9.822 | −0.0535154 | −0.0175027 | RRGP | Liquiritigenin * | 261, 215, 159, 147, 137, 119, 91, 81 |
| m/z RT | p[1] | po[1] | Origin | Compound | Fragment Ions | Literature |
|---|---|---|---|---|---|---|
| 285.07656_10.514 | −0.167255 | 0.258227 | RA | Calycosin * | 270, 253, 241, 229, 225, 137 | [24,25] |
| 269.08165_13.771 | −0.153853 | 0.218006 | RA | Formononetin * | 237, 225, 213, 209, 136 | Metlin, [24] |
| 355.11759_16.953 | −0.0865657 | −0.002337 | RRGP | n.i. | ||
| 714.41099_18.603 | −0.0851547 | −0.0579323 | RZ | n.i. | ||
| 301.10751_10.011 | −0.0794242 | 0.113695 | RA | 3-Hydroxy-9,10-dimethoxypterocarpan | 269, 191, 167, 152, 147, 133, 123, 105 | [25] |
| 1737.96335_19.162 | −0.0743328 | 0.178967 | RA | Astragaloside I isomer (dimer) | 851, 833, 689, 671, 653, 491, 473, 455, 437, 419 | [24,26] |
| 1737.96629_18.398 | −0.0743063 | 0.1784 | RA | Astragaloside I * (dimer) | 851, 833, 689, 671, 653, 491, 473, 455, 437, 419 | [24,26] |
| 301.10773_14.151 | −0.0733961 | 0.166495 | RA | 3-Hydroxy-9,10-dimethoxypterocarpan | 269, 191, 179, 167, 152, 133, 147, 123, 105 | [25] |
| 371.10109_32.627 | −0.0708126 | 0.0542131 | All | n.i. | ||
| 508.25367_16.894 | −0.0670202 | −0.0193137 | RZ | n.i. | ||
| 355.06981_32.607 | −0.0669833 | 0.135838 | RA | n.i. | ||
| 891.46950_19.156 | −0.0653422 | 0.0481451 | RA | Astragaloside I isomer | 671, 653, 491, 473, 455, 437, 419, 297 | [24,26] |
| 257.08161_9.822 | −0.0652777 | −0.0002341 | RRGP | Liquiritigenin * | 261, 215, 159, 147, 137, 119 | |
| 891.47006_18.395 | −0.0652695 | 0.0662892 | RA | Astragaloside I * [M + Na]+ | 689, 671, 653, 491, 473, 455, 437, 419, 297 | [24,26] |
| 447.12852_6.654 | −0.0627192 | −0.0564573 | RA | Calycosin-7-O-β-d-glucoside | 270, 253, 225, 137 | [24] |
| 353.10204_16.488 | −0.059812 | 0.0144688 | All | n.i. | ||
| 473.14387_12.038 | −0.0597473 | 0.0753214 | RA | n.i. | ||
| 315.08661_10.947 | −0.0583033 | 0.116079 | RA | Odoratin | 300, 283, 259, 255, 244, 167 | Metlin |
| m/z RT | p[1] | po[1] | Origin | Compound | Fragment Ions | Literature |
|---|---|---|---|---|---|---|
| 265.12345_8.988 | −0.0930574 | 0.175785 | FJ | n.i. | ||
| 481.17020_5.056 | −0.0759404 | −0.013529 | RP | Albiflorin * | 503 [M + Na], 197, 151, 133 | [22,23] |
| 415.21121_18.824 | −0.0736124 | 0.0616802 | FJ | n.i. | ||
| 503.15230_5.509 | −0.0722717 | −0.0109856 | RP | Paeoniflorin * [M + Na]+ | 498 [M + NH4], 179, 151, 133 | [22,23] |
| 503.15227_5.348 | −0.0719782 | −0.0112909 | RP | Paeoniflorin * [M + Na]+ | 498 [M + NH4], 179, 151, 133 | [22,23] |
| 498.19688_5.504 | −0.0712826 | −0.0135778 | RP | Paeoniflorin * [M + NH4]+ | 503 [M + Na], 179, 151, 133 | [22,23] |
| 481.17023_4.821 | −0.0650068 | −0.0111313 | RP | Albiflorin * | 503 [M + Na], 197, 151, 133 | [22,23] |
| 714.41099_18.603 | −0.0639971 | 0.0657522 | RZ | n.i. | ||
| 452.32137_22.770 | −0.0626265 | 0.0403574 | FJ | n.i. | ||
| 503.15215_5.060 | −0.0608256 | −0.0112778 | RP | Albiflorin * [M + Na]+ | 498 [M + NH4], 197, 151, 133 | [22,23] |
| 498.19685_5.348 | −0.0600451 | −0.0104305 | RP | Paeoniflorin * | 503 [M + Na], 179, 151, 133 | [22,23] |
| 503.15220_8.397 | −0.0562873 | −0.0109413 | RP | Albiflorin isomer | 219, 197, 133, 105 | [22,23] |
| 282.14928_9.203 | −0.0533495 | 0.0556512 | FJ | n.i. | ||
| 503.15228_4.813 | −0.0531328 | −0.0098537 | RP | Albiflorin * [M + Na]+ | 498 [M + NH4], 197, 151, 133 | [22,23] |
| 438.23858_13.465 | −0.0501564 | 0.0505945 | RZ | n.i. | ||
| 983.31365_5.058 | −0.0496619 | −0.0078355 | RP | Albiflorin * | 503 [M + Na], 481, 197, 151, 133, 105 | [22,23] |
| 379.24752_22.864 | −0.0443945 | 0.0688323 | FJ | n.i. | ||
| 464.24899_9.989 | −0.0438006 | −0.005821 | RP | Pinen-10-yl-ß-vicianoside | 133, 127, 115 | |
| 265.12281_9.203 | −0.0422487 | 0.010489 | RZ | n.i. | ||
| 983.31391_4.812 | −0.040604 | −0.0057421 | RP | Albiflorin * | 503 [M + Na], 481, 197, 151, 133, 105 | [22,23] |

| Compound | Chemical Structure | Predicted Activity | Verified Activity |
|---|---|---|---|
| Astragaloside I |  | TNF-α, IL-1β | TNF-α, IL-1β, IFN-γ |
| Astragaloside II |  | TNF-α | TNF-α, IL-1β, IFN-γ |
| Calycosin |  | TNF-α, IL-1β | IL-1β |
| Formononetin |  | TNF-α, IL-1β | TNF-α, IL-1β, IFN-γ |
| Liquiritigenin |  | TNF-α, IL-1β | TNF-α, IL-1β, IFN-γ |
| Albiflorin |  | IFN-γ | IL-1β, |
| Paeoniflorin |  | IFN-γ | IL-1β, IFN-γ |
| Isoliquiritigenin |  | IL-1β, IFN-γ |

References
- Bosma-den Boer, M.M.; van Wetten, M.-L.; Pruimboom, L. Chronic inflammatory diseases are stimulated by current lifestyle: How diet, stress levels and medication prevent our body from recovering. Nutr. Metab. 2012, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S. Chronic Diseases Caused by Chronic Inflammation Require Chronic Treatment: Anti-inflammatory Role of Dietary Spices. J. Clin. Cell Immunol. 2014, 5. [Google Scholar] [CrossRef]
- Hou, J.P.; Jin, Y. The Healing Power of Chinese Herbs and Medicinal Recipes; Haworth Integrative Healing Pr: New York, NY, USA; London, UK; Oxford, UK, 2005; ISBN 978-0789022028. [Google Scholar]
- Nikles, S. Metabolic and Pharmacological Profiling of Three Classical TCM Formulations Used for Chronic Inflammatory Diseases with Immune Dysfunction. Ph.D. Dissertation, Karl-Franzens-Universität, Graz, Austria, 2016. [Google Scholar]
- Zhang, Z.; Luo, X. Synopsis of Prescriptions of the Golden Chamber with 300 Cases. A Classic of Traditional Chinese Medicine with Ancient and Contemporary Case Studies; New World Press: Beijing, China, 1995; ISBN 978-7800052910. [Google Scholar]
- Chen, K.-T.; Su, C.-H.; Hsin, L.-H.; Su, Y.-C.; Su, Y.-P.; Lin, J.-G. Reducing fatigue of athletes following oral administration of huangqi jianzhong tang. Acta Pharmacol. Sin. 2002, 23, 757–761. [Google Scholar] [PubMed]
- Nikles, S.; Monschein, M.; Zou, H.; Liu, Y.; He, X.; Fan, D.; Lu, A.; Yu, K.; Isaac, G.; Bauer, R. Metabolic profiling of the traditional Chinese medicine formulation Yu Ping Feng San for the identification of constituents relevant for effects on expression of TNF-α, IFN-γ, IL-1β and IL-4 in U937 cells. J. Pharm. Biomed. Anal. 2017, 145, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.-Y.; Pan, C.-S.; Liu, Y.-Y.; Wei, X.-H.; Zhou, C.-M.; Sun, K.; He, K.; Li, C.; Yan, L.; Fan, J.-Y.; et al. Huang Qi Jian Zhong pellet attenuates TNBS-induced colitis in rats via mechanisms involving improvement of energy metabolism. Evidence-Based. Evid. Based Complement. Alternat. Med. 2013, 2013. [Google Scholar] [CrossRef]
- Wei, Y.; Ma, L.X.; Yin, S.J.; An, J.; Wei, Q.; Yang, J.X. Huangqi Jianzhong Tang for Treatment of Chronic Gastritis: A Systematic Review of Randomized Clinical Trials. Evid. Based Complement. Alternat. Med. eCAM 2015, 878164. [Google Scholar] [CrossRef]
- Chung, K.F. Cytokines. Asthma and COPD; Elsevier: Amsterdam, The Netherlands, 2009; pp. 327–341. ISBN 9780123740014. [Google Scholar]
- Udalova, I.; Monaco, C.; Nanchahal, J.; Feldmann, M. Anti-TNF Therapy. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.-M.; An, J. Cytokines, Inflammation and Pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef]
- Yuliana, N.D.; Khatib, A.; Choi, Y.H.; Verpoorte, R. Metabolomics for bioactivity assessment of natural products. Phytother. Res. 2011, 25, 157–169. [Google Scholar] [CrossRef]
- Commisso, M.; Strazzer, P.; Toffali, K.; Stocchero, M.; Guzzo, F. Untargeted metabolomics: An emerging approach to determine the composition of herbal products. Comput. Struct. Biotechnol. J. 2013, 4, e201301007. [Google Scholar] [CrossRef]
- Alonso, A.; Marsal, S.; Julià, A. Analytical methods in untargeted metabolomics: State of the art in 2015. Front. Bioeng. Biotechnol. 2015, 3, 23. [Google Scholar] [CrossRef] [PubMed]
- Wolfender, J.-L.; Marti, G.; Thomas, A.; Bertrand, S. Current approaches and challenges for the metabolite profiling of complex natural extracts: Editors’ Choice IX. J. Chromatogr. A 2015, 1382, 136–164. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Khan, I.A.; Combrinck, S.; Sandasi, M.; Chen, W.; Viljoen, A.M. 1H-NMR and UPLC-MS metabolomics: Functional tools for exploring chemotypic variation in Sceletium tortuosum from two provinces in South Africa. Phytochemistry 2018, 152, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Viljoen, A.M.; Zhao, J.; Sandasi, M.; Chen, W.; Khan, I.A. Phytochemical distinction between Pelargonium sidoides (“Umckaloabo”) and P. reniforme through 1H-NMR and UHPLC–MS metabolomic profiling. Metabolomics 2015, 11, 594–602. [Google Scholar] [CrossRef]
- Afzan, A.; Kasim, N.; Ismail, N.H.; Azmi, N.; Ali, A.M.; Mat, N.; Wolfender, J.-L. Differentiation of Ficus deltoidea varieties and chemical marker determination by UHPLC-TOFMS metabolomics for establishing quality control criteria of this popular Malaysian medicinal herb. Metabolomics 2019, 15, 35. [Google Scholar] [CrossRef] [PubMed]
- Cardoso-Taketa, A.T.; Pereda-Miranda, R.; Choi, Y.H.; Verpoorte, R.; Villarreal, M.L. Metabolic profiling of the Mexican anxiolytic and sedative plant Galphimia glauca using nuclear magnetic resonance spectroscopy and multivariate data analysis. Planta Med. 2008, 74, 1295–1301. [Google Scholar] [CrossRef] [PubMed]
- Bauermeister, A.; Pereira, F.; Grilo, I.R.; Godinho, C.C.; Paulino, M.; Almeida, V.; Gobbo-Neto, L.; Prieto-Davo, A.; Sobral, R.G.; Lopes, N.P.; et al. Intra-clade metabolomic profiling of MAR4 Streptomyces from the Macaronesia Atlantic region reveals a source of anti-biofilm metabolites. Environ. Microbiol. 2019, 21, 1099–1112. [Google Scholar] [CrossRef]
- Gong, C.; Yang, H.; Wei, H.; Qi, C.; Wang, C.-H. Pharmacokinetic comparisons by UPLC-MS/MS of isomer paeoniflorin and albiflorin after oral administration decoctions of single-herb Radix Paeoniae Alba and Zengmian Yiliu prescription to rats. Biomed. Chromatogr. 2015, 29, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Wan, M.; Zhou, D.; Gao, J.; Zhu, Y.; Bi, K. LC-MS/MS determination and pharmacokinetic study of albiflorin and paeoniflorin in rat plasma after oral administration of Radix Paeoniae Alba extract and Tang-Min-Ling-Wan. Biomed. Chromatogr. 2010, 24, 1324–1331. [Google Scholar] [CrossRef]
- Huang, X.; Liu, Y.; Song, F.; Liu, Z.; Liu, S. Studies on principal components and antioxidant activity of different Radix Astragali samples using high-performance liquid chromatography/electrospray ionization multiple-stage tandem mass spectrometry. Talanta 2009, 78, 1090–1101. [Google Scholar] [CrossRef]
- Xiao, H.B.; Krucker, M.; Albert, K.; Liang, X.M. Determination and identification of isoflavonoids in Radix astragali by matrix solid-phase dispersion extraction and high-performance liquid chromatography with photodiode array and mass spectrometric detection. J. Chromatogr. A 2004, 1032, 117–124. [Google Scholar] [CrossRef]
- Xiao, M.; Chen, H.; Shi, Z.; Feng, Y.; Rui, W. Rapid and reliable method for analysis of raw and honey-processed astragalus by UPLC/ESI-Q-TOF-MS using HSS T3 columns. Anal. Methods 2014, 6, 8045–8054. [Google Scholar] [CrossRef]
- Esposito, E.; Cuzzocrea, S. TNF-Alpha as a Therapeutic Target in Inflammatory Diseases, Ischemia- Reperfusion Injury and Trauma. CMC 2009, 16, 3152–3167. [Google Scholar] [CrossRef] [PubMed]
- Van Horssen, R.; ten Hagen, T.L.M.; Eggermont, A.M.M. TNF-alpha in cancer treatment: Molecular insights, antitumor effects, and clinical utility. Oncologist 2006, 11, 397–408. [Google Scholar] [CrossRef]
- Horiuchi, T.; Mitoma, H.; Harashima, S.-I.; Tsukamoto, H.; Shimoda, T. Transmembrane TNF-α: Structure, function and interaction with anti-TNF agents. Rheumatology 2010, 49, 1215–1228. [Google Scholar] [CrossRef] [PubMed]
- Bazzoni, F.; Beutler, B. The Tumor Necrosis Factor Ligand and Receptor Families. N. Engl. J. Med. 1996, 334, 1717–1725. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Castejon, G.; Brough, D. Understanding the mechanism of IL-1β secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef]
- Akdis, M.; Burgler, S.; Crameri, R.; Eiwegger, T.; Fujita, H.; Gomez, E.; Klunker, S.; Meyer, N.; O’Mahony, L.; Palomares, O.; et al. Interleukins, from 1 to 37, and interferon-γ: Receptors, functions, and roles in diseases. J. Allergy Clin. Immunol. 2011, 127, 701–721. [Google Scholar] [CrossRef]
- Payne, S. Chapter 6-Immunity and Resistance to Viruses. In Viruses; Payne, S., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 61–71. ISBN 978-0-12-803109-4. [Google Scholar]
- The Autoimmune Diseases, 5th ed.; Rose, N.R.; Mackay, I.R. (Eds.) Academic Press: Boston, MA, USA, 2014; ISBN 978-0-12-384929-8. [Google Scholar]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-gamma: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [CrossRef]
- Zhang, J. Yin and yang interplay of IFN-γ in inflammation and autoimmune disease. J. Clin. Investig. 2007, 117, 871–873. [Google Scholar] [CrossRef]
- Sang, A.; Yin, Y.; Zheng, Y.-Y.; Morel, L. Animal Models of Molecular Pathology: Systemic Lupus Erythematosus. In Progress in Molecular Biology and Translational Science: Animal Models of Molecular Pathology; Conn, P.M., Ed.; Academic Press: Cambridge, MA, USA, 2012; pp. 321–370. ISBN 1877-1173. [Google Scholar]
- Lin, F.-C.; Young, H.A. The talented interferon-gamma. ABB 2013, 4, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Xu, H.-Y.; Xu, L.; Wang, S.-S.; Zhang, X.-M. In vivo and in vitro immunomodulatory and anti-inflammatory effects of total flavonoids of astragalus. Afr. J. Tradit. Complement. Altern. Med. 2016, 13, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Huang, Q.; Chen, J.; Wang, M.; Pan, H.; Wang, R.; Zhou, H.; Zhou, Z.; Liu, J.; Yang, F.; et al. Calycosin suppresses expression of pro-inflammatory cytokines via the activation of p62/Nrf2-linked heme oxygenase 1 in rheumatoid arthritis synovial fibroblasts. Pharmacol. Res. 2016, 113, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sun, Y.N.; Yan, X.T.; Yang, S.Y.; Kim, S.; Lee, Y.M.; Koh, Y.-S.; Kim, Y.H. Flavonoids from Astragalus membranaceus and their inhibitory effects on LPS-stimulated pro-inflammatory cytokine production in bone marrow-derived dendritic cells. Arch. Pharmacal. Res. 2014, 37, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Yesilada, E.; Bedir, E.; Çalış, İ.; Takaishi, Y.; Ohmoto, Y. Effects of triterpene saponins from Astragalus species on in vitro cytokine release. J. Ethnopharmacol. 2005, 96, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.-Y.; Ha, Y.J.; Kim, K.-M.; Jung, Y.-S.; Jung, J.-C.; Oh, S. Anti-Inflammatory Activities of Licorice Extract and Its Active Compounds, Glycyrrhizic Acid, Liquiritin and Liquiritigenin, in BV2 Cells and Mice Liver. Molecules 2015, 20, 13041–13054. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.W.; Zhao, R.J.; Park, S.J.; Lee, J.R.; Cho, I.J.; Yang, C.H.; Kim, S.G.; Kim, S.C. Anti-inflammatory effects of liquiritigenin as a consequence of the inhibition of NF-kappaB-dependent iNOS and proinflammatory cytokines production. Br. J. Pharm. 2008, 154, 165–173. [Google Scholar] [CrossRef]
- Wang, Y.N.; Zhang, Y.; Wang, Y.; Zhu, D.X.; Xu, L.Q.; Fang, H.; Wu, W. The Beneficial Effect of Total Glucosides of Paeony on Psoriatic Arthritis Links to Circulating Tregs and Th1 Cell Function. Phytother. Res. 2014, 28, 372–381. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, R.; Zhou, F.; Cheng, H.; Xia, B. Total Glucosides of Peony Attenuates 2,4,6-Trinitrobenzene Sulfonic Acid/Ethanol-Induced Colitis in Rats Through Adjustment of Th1/Th2 Cytokines Polarization. Cell Biochem. Biophys. 2014, 68, 83–95. [Google Scholar] [CrossRef]
- Chang, Y.; Wei, W.; Zhang, L.; Xu, H.-M. Effects and mechanisms of total glucosides of paeony on synoviocytes activities in rat collagen-induced arthritis. J. Ethnopharmacol. 2009, 121, 43–48. [Google Scholar] [CrossRef]
- Zheng, Y.-Q.; Wei, W. Total glucosides of paeony suppresses adjuvant arthritis in rats and intervenes cytokine-signaling between different types of synoviocytes. Int. Immunopharmacol. 2005, 5, 1560–1573. [Google Scholar] [CrossRef]
- Bi, X.; Han, L.; Qu, T.; Mu, Y.; Guan, P.; Qu, X.; Wang, Z.; Huang, X. Anti-Inflammatory Effects, SAR, and Action Mechanism of Monoterpenoids from Radix Paeoniae Alba on LPS-Stimulated RAW 264.7 Cells. Molecules 2017, 22, 715. [Google Scholar] [CrossRef]
Sample Availability: All cited extracts and samples of the compounds formononetin, calycosin, astragaloside I, astragaloside II, liquiritigenin, isoliquiritigenin, 18ß-glycyrrhetinic acid, paeoniflorin and albiflorin are available from the authors. |








| Activity Class | TNF-α Inhibition (%) | IL-1β Inhibition (%) | IFN-γ Inhibition (%) |
|---|---|---|---|
| 1 (active) | 70–100 | 60–100 | 60–100 |
| 2 (moderately active) | 50–69 | 45–59 | 45–59 |
| 3 (inactive) | 0–49 | 0–44 | 0–44 |
| RT (min) | Detected m/z (ESI+) | (Tentative) Identification | Origin | Mono-Isotopic Mass (g/mol) | Molecular Formula | MS/MS Fragment Ions | Literature for Identification | Pharmacological Activity Predicted by OPLS-DA | p[1] | po[1] |
|---|---|---|---|---|---|---|---|---|---|---|
| 4.82 | 481.17023 | Albiflorin * | RP | 480.163 | C23H28O11 | 197, 151, 133 | [22,23] | IFN-γ | −0.0650068 | −0.0111313 |
| 5.34 | 503.15227 | Paeoniflorin * (Na-Adduct) | RP | 480.163 | C23H28O11 | 179, 151, 133 | [22,23] | IFN-γ | −0.0719782 | −0.0112909 |
| 6.48 | 447.1285 | Calycosin-7-O-β-d-glucoside or isomer | RA | 446.121 | C22H22O10 | 270, 253, 225, 137 | [24] | TNF-α | −0.0687443 | 0.132943 |
| 8.39 | 503.1522 | Albiflorin isomer (Na-Adduct) | RP | 480.163 | C23H28O11 | 219, 197, 133, 105 | [22,23] | IFN-γ | −0.0562873 | −0.0109413 |
| 9.82 | 257.08161 | Liquiritigenin * | RRGP | 256.074 | C15H12O4 | 137, 147, 119, 261, 91, 81, 215, 159 | TNF-α IL-1β | −0.0535154 −0.0652777 | −0.0175027 −0.0002341 | |
| 9.98 | 464.2489 | Pinen-10-yl-ß-vicianoside (NH4-Adduct) | RP | 446.215 | C21H34O10 | 133, 127, 115 | IFN-γ | −0.0438006 | −0.005821 | |
| 10.01 | 301.1075 | 3-Hydroxy-9,10-dimethoxypterocarpan or isomer | RA | 300.1 | C17H16O5 | 269, 191, 167, 152, 147, 133, 123, 105 | [25] | TNF-α IL-1β | −0.0767608 −0.0794242 | 0.176659 0.113695 |
| 10.51 | 285.07656 | Calycosin * | RA | 284.068 | C16H12O5 | 270, 253, 225, 229, 241, 137 | [24,25] | TNF-α, IL-1β | −0.151342 −0.167255 | 0.247407 0.258227 |
| 10.94 | 315.0866 | Odoratin | RA | 314.079 | C17H14O6 | 300, 283, 259, 255, 244, 167 | Metlin | IL-1β | −0.0583033 | 0.116079 |
| 13.77 | 269.08165 | Formononetin * | RA | 268.074 | C16H12O4 | 237, 209, 213, 225, 136 | Metlin, [24] | TNF-α IL-1β | −0.135887 −0.153853 | 0.108774 0.218006 |
| 14.15 | 301.1077 | 3-Hydroxy-9,10-dimethoxypterocarpan or isomer | RA | 300.1 | C17H16O5 | 269, 191, 167, 152, 147, 133, 123, 105 | [25] | TNF-α IL-1β | −0.0535515 −0.0733961 | 0.0891372 0.166495 |
| 15.87 | 1653.9439 | Astragaloside II (dimer) | RA | 798.44 | C41H66O15 | 647, 629, 491, 473, 455, 437, 419 | [24,26] | TNF-α | −0.0558838 | 0.154538 |
| 18.39 | 1737.9663 | Astragaloside I * (dimer) | RA | 868.482 | C45H72O16 | 689, 671, 653, 491, 473, 455, 437, 419, 297 | [24,26] | TNF-α IL-1β | −0.0831685 −0.0743063 | 0.219044 0.1784 |
| 19.15 | 1737.9633 | Astragaloside I isomer (dimer) | RA | 868.482 | C45H72O16 | 671, 653, 491, 473, 455, 437, 419, 297 | [24,26] | TNF-α IL-1β | −0.0827036 −0.0743328 | 0.216071 0.178967 |
| 20.22 | 1737.9632 | Astragaloside I isomer (dimer) | RA | 868.482 | C45H72O16 | 851, 833, 689, 671, 653, 491, 473, 455, 437, 419, 297 | [24,26] | TNF-α | −0.0571481 | 0.146308 |
| 24.71 | 471.3464 | 18β-Glycyrrhetinic acid * | RRGP | 470.34 | C30H46O4 | 453, 235, 217, 189, 175, 189 | Metlin | TNF-α | −0.0542538 | −0.0348534 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nöst, X.; Pferschy-Wenzig, E.-M.; Nikles, S.; He, X.; Fan, D.; Lu, A.; Yuk, J.; Yu, K.; Isaac, G.; Bauer, R. Identification of Constituents Affecting the Secretion of Pro-Inflammatory Cytokines in LPS-Induced U937 Cells by UHPLC-HRMS-Based Metabolic Profiling of the Traditional Chinese Medicine Formulation Huangqi Jianzhong Tang. Molecules 2019, 24, 3116. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24173116
Nöst X, Pferschy-Wenzig E-M, Nikles S, He X, Fan D, Lu A, Yuk J, Yu K, Isaac G, Bauer R. Identification of Constituents Affecting the Secretion of Pro-Inflammatory Cytokines in LPS-Induced U937 Cells by UHPLC-HRMS-Based Metabolic Profiling of the Traditional Chinese Medicine Formulation Huangqi Jianzhong Tang. Molecules. 2019; 24(17):3116. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24173116
Chicago/Turabian StyleNöst, Xuehong, Eva-Maria Pferschy-Wenzig, Stefanie Nikles, Xiaojuan He, Danping Fan, Aiping Lu, Jimmy Yuk, Kate Yu, Giorgis Isaac, and Rudolf Bauer. 2019. "Identification of Constituents Affecting the Secretion of Pro-Inflammatory Cytokines in LPS-Induced U937 Cells by UHPLC-HRMS-Based Metabolic Profiling of the Traditional Chinese Medicine Formulation Huangqi Jianzhong Tang" Molecules 24, no. 17: 3116. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24173116