Comparison of Free, Esterified, and Insoluble-Bound Phenolics and Their Bioactivities in Three Organs of Lonicera japonica and L. macranthoides


Abstract
:
1. Introduction
2. Results and Discussion
2.1. Differences in Contents of Three Phenolic Fractions in L. japonica and L. macranthoides
2.2. Identification and Quantification of Phenolics in Three Different Organs of L. japonica and L. macranthoides
2.3. Differences in Antioxidant Activities of the Three Phenolic Fractions in L. japonica and L. macranthoides as Well as the 10 Identified Phenolics
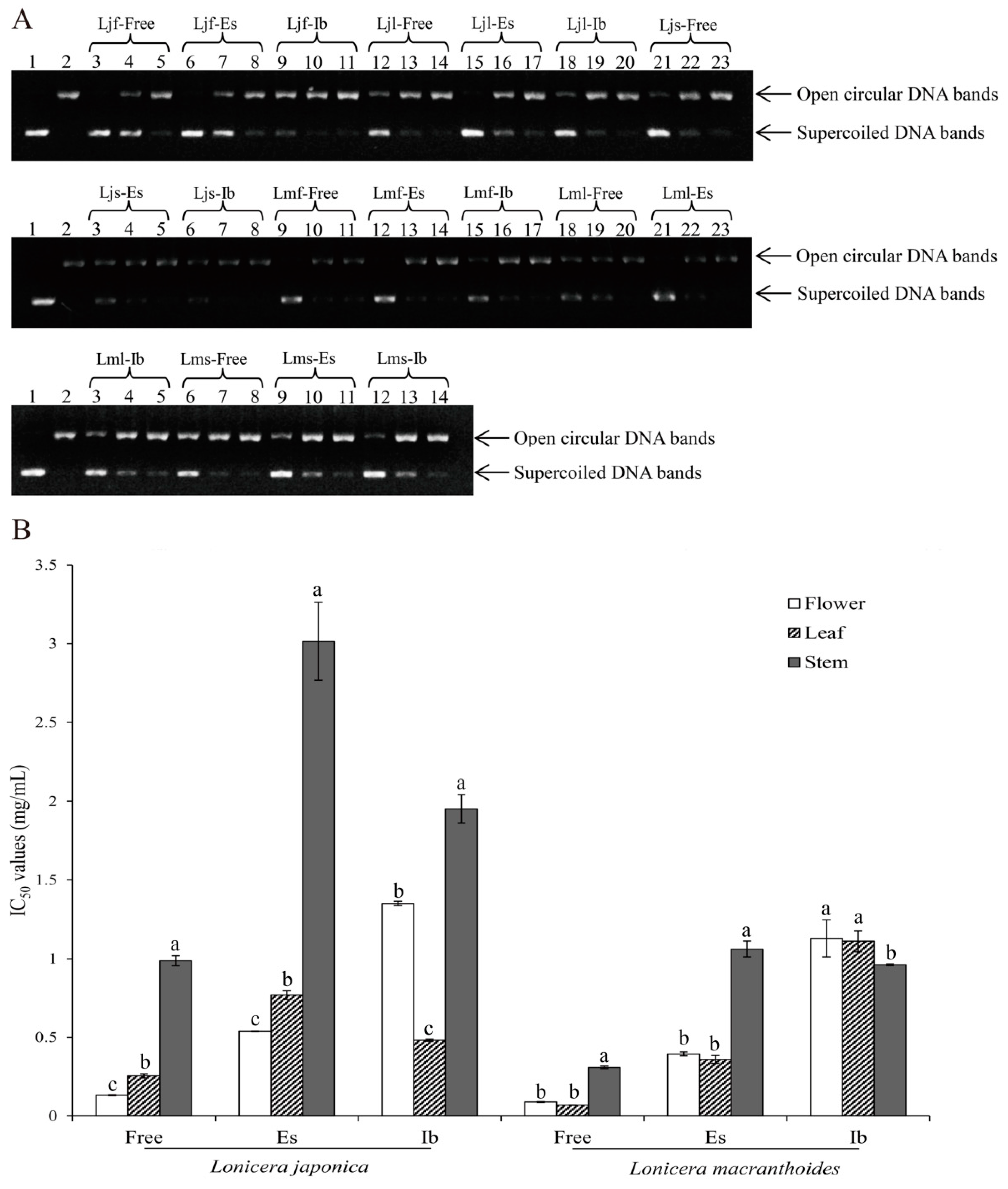
2.4. Differences in Protective Activities to DNA Damage of the Three Phenolic Fractions in L. japonica and L. macranthoides
2.5. Correlation Analysis
3. Materials and Methods
3.1. Chemical Reagents and Plant Materials
3.2. Extraction of Phenolic Compounds
3.3. Determination of Phenolic Compounds
3.4. Evaluation of Radical Scavenging Activities
3.5. Examination of Inhibition to DNA Damage
3.6. UPLC-DAD–QTOF-MS/MS Analysis
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hu, X.; Chen, L.; Shi, S.; Cai, P.; Liang, X.; Zhang, S. Antioxidant capacity and phenolic compounds of Lonicerae macranthoides by HPLC-DAD-QTOF-MS/MS. J. Pharm. Biomed. Anal. 2016, 124, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Quan, Q.; Zhou, X.; Zhu, Y.; Lan, Y.; Li, S.; Yu, Y.; Cheng, Z. A comparative study of Lonicera japonica with related species: Morphological characteristics, ITS sequences and active compounds. Biochem. Syst. Ecol. 2014, 54, 198–207. [Google Scholar] [CrossRef]
- Yan, R.; Chen, J.-B.; Sun, S.-Q.; Guo, B.-L. Rapid identification of Lonicerae japonicae Flos and Lonicerae Flos by Fourier transform infrared (FT-IR) spectroscopy and two-dimensional correlation analysis. J. Mol. Struct. 2016, 1124, 110–116. [Google Scholar] [CrossRef]
- Yuan, Y.; Song, L.; Li, M.; Liu, G.; Chu, Y.; Ma, L.; Zhou, Y.; Wang, X.; Gao, W.; Qin, S.; et al. Genetic variation and metabolic pathway intricacy govern the active compound content and quality of the Chinese medicinal plant Lonicera japonica thunb. BMC Genom. 2012, 13, 195. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.-F.; Hsiao, P.-C.; Kuo, T.-C.; Chiang, S.-T.; Chen, S.-L.; Chiou, S.-J.; Ling, X.-H.; Liang, M.-T.; Cheng, W.-Y.; Houng, J.-Y. Antioxidant and anti-inflammatory activities of Lonicera japonica Thunb. var. sempervillosa Hayata flower bud extracts prepared by water, ethanol and supercritical fluid extraction techniques. Ind. Crops Prod. 2016, 89, 543–549. [Google Scholar] [CrossRef]
- Liu, Z.-X.; Liu, C.-T.; Liu, Q.-B.; Ren, J.; Li, L.-Z.; Huang, X.-X.; Wang, Z.-Z.; Song, S.-J. Iridoid glycosides from the flower buds of Lonicera japonica and their nitric oxide production and α-glucosidase inhibitory activities. J. Funct. Foods 2015, 18, 512–519. [Google Scholar] [CrossRef]
- Park, H.S.; Park, K.I.; Lee, D.H.; Kang, S.R.; Nagappan, A.; Kim, J.A.; Kim, E.H.; Lee, W.S.; Shin, S.C.; Hah, Y.S.; et al. Polyphenolic extract isolated from Korean Lonicera japonica Thunb. induce G2/M cell cycle arrest and apoptosis in HepG2 cells: Involvements of PI3K/Akt and MAPKs. Food Chem. Toxicol. 2012, 50, 2407–2416. [Google Scholar] [CrossRef]
- Xiong, J.; Li, S.; Wang, W.; Hong, Y.; Tang, K.; Luo, Q. Screening and identification of the antibacterial bioactive compounds from Lonicera japonica Thunb. leaves. Food Chem. 2013, 138, 327–333. [Google Scholar] [CrossRef]
- Han, M.H.; Lee, W.S.; Nagappan, A.; Hong, S.H.; Jung, J.H.; Park, C.; Kim, H.J.; Kim, G.Y.; Kim, G.; Jung, J.M.; et al. Flavonoids Isolated from Flowers of Lonicera japonica Thunb. Inhibit Inflammatory Responses in BV2 Microglial Cells by Suppressing TNF-α and IL-beta Through PI3K/Akt/NF-kb Signaling Pathways. Phytother. Res. 2016, 30, 1824–1832. [Google Scholar] [CrossRef]
- Mahboob, T.; Azlan, A.M.; Tan, T.C.; Samudi, C.; Sekaran, S.D.; Nissapatorn, V.; Wiart, C. Anti-encystment and amoebicidal activity of Lonicera japonica Thunb. and its major constituent Chlorogenic acid in vitro. Asian Pac. J. Trop. Med. 2016, 9, 844–850. [Google Scholar] [CrossRef]
- Park, K.I.; Park, H.; Nagappan, A.; Hong, G.E.; Yumnam, S.; Lee, H.J.; Kim, E.H.; Lee, W.S.; Shin, S.C.; Kim, J.A.; et al. Polyphenolic compounds from Korean Lonicera japonica Thunb. induces apoptosis via AKT and caspase cascade activation in A549 cells. Oncol. Lett. 2017, 13, 2521–2530. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekara, A.; Shahidi, F. Bioactivities and antiradical properties of millet grains and hulls. J. Agric. Food Chem. 2011, 59, 9563–9571. [Google Scholar] [CrossRef] [PubMed]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Chai, X.-Y.; Li, S.-L.; Li, P. Quality evaluation of Flos Lonicerae through a simultaneous determination of seven saponins by HPLC with ELSD. J. Chromatogr. A 2005, 1070, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Estrada, B.A.; Gutierrez-Uribe, J.A.; Serna-Saldivar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Alves, G.H.; Ferreira, C.D.; Vivian, P.G.; Monks, J.L.; Elias, M.C.; Vanier, N.L.; de Oliveira, M. The revisited levels of free and bound phenolics in rice: Effects of the extraction procedure. Food Chem. 2016, 208, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ju-Dong, Y. Insoluble-bound phenolics in food. Molecules 2016, 21, 1216. [Google Scholar] [CrossRef]
- Chen, G.-L.; Zhang, X.; Chen, S.-G.; Han, M.-D.; Gao, Y.-Q. Antioxidant activities and contents of free, esterified and insoluble-bound phenolics in 14 subtropical fruit leaves collected from the south of China. J. Funct. Foods 2017, 30, 290–302. [Google Scholar] [CrossRef]
- Ayoub, M.; de Camargo, A.C.; Shahidi, F. Antioxidants and bioactivities of free, esterified and insoluble-bound phenolics from berry seed meals. Food Chem. 2016, 197, 221–232. [Google Scholar] [CrossRef]
- Gutiérrez-Uribe, J.A.; Rojas-García, C.; García-Lara, S.; Serna-Saldivar, S.O. Phytochemical analysis of wastewater (nejayote) obtained after lime-cooking of different types of maize kernels processed into masa for tortillas. J. Cereal Sci. 2010, 52, 410–416. [Google Scholar] [CrossRef]
- Irakli, M.N.; Samanidou, V.F.; Biliaderis, C.G.; Papadoyannis, I.N. Development and validation of an HPLC-method for determination of free and bound phenolic acids in cereals after solid-phase extraction. Food Chem. 2012, 134, 1624–1632. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-K.; Zhang, X.; Chen, G.-L.; Yu, J.; Yang, L.-Q.; Gao, Y.-Q. Antioxidant property and their free, soluble conjugate and insoluble-bound phenolic contents in selected beans. J. Funct. Foods 2016, 24, 359–372. [Google Scholar] [CrossRef]
- Gao, Y.; Ma, S.; Wang, M.; Feng, X.Y. Characterization of free, conjugated, and bound phenolic acids in seven commonly consumed vegetables. Molecules 2017, 22, 1878. [Google Scholar] [CrossRef] [PubMed]
- Ambigaipalan, P.; de Camargo, A.C.; Shahidi, F. Phenolic Compounds of Pomegranate Byproducts (Outer Skin, Mesocarp, Divider Membrane) and Their Antioxidant Activities. J. Agric. Food Chem. 2016, 64, 6584–6604. [Google Scholar] [CrossRef] [PubMed]
- Min, B.; Gu, L.; McClung, A.M.; Bergman, C.J.; Chen, M.-H. Free and bound total phenolic concentrations, antioxidant capacities, and profiles of proanthocyanidins and anthocyanins in whole grain rice (Oryza sativa L.) of different bran colours. Food Chem. 2012, 133, 715–722. [Google Scholar] [CrossRef]
- Alshikh, N.; de Camargo, A.C.; Shahidi, F. Phenolics of selected lentil cultivars: Antioxidant activities and inhibition of low-density lipoprotein and DNA damage. J. Funct. Foods 2015, 18, 1022–1038. [Google Scholar] [CrossRef]
- Shahidi, F.; Zhong, Y. Measurement of antioxidant activity. J. Funct. Foods 2015, 18, 757–781. [Google Scholar] [CrossRef]
- Menghini, L.; Leporini, L.; Vecchiotti, G.; Locatelli, M.; Carradori, S.; Ferrante, C.; Zengin, G.; Recinella, L.; Chiavaroli, A.; Leone, S.; et al. Crocus sativus L. stigmas and byproducts: Qualitative fingerprint, antioxidant potentials and enzyme inhibitory activities. Food Res. Int. 2018, 109, 91–98. [Google Scholar] [CrossRef]
- Seo, O.N.; Kim, G.-S.; Park, S.; Lee, J.H.; Kim, Y.-H.; Lee, W.S.; Lee, S.J.; Kim, C.Y.; Jin, J.S.; Choi, S.K.; et al. Determination of polyphenol components of Lonicera japonica Thunb. using liquid chromatography–tandem mass spectrometry: Contribution to the overall antioxidant activity. Food Chem. 2012, 134, 572–577. [Google Scholar] [CrossRef]
- Yang, L.; Yin, P.; Li, K.; Fan, H.; Xue, Q.; Li, X.; Sun, L.; Liu, Y. Seasonal dynamics of constitutive levels of phenolic components lead to alterations of antioxidant capacities in Acer truncatum leaves. Arab. J. Chem. 2017, 11, 14–25. [Google Scholar] [CrossRef]
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, antioxidants, and the degenerative diseases of aging. Proc. Natl. Acad. Sci. USA 1993, 90, 7915–7922. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Zhou, B.; Cai, Y.; Yang, L.; Liu, Z. Synergistic effect of green tea polyphenols with trolox on free radical-induced oxidative DNA damage. Food Chem. 2006, 96, 90–95. [Google Scholar] [CrossRef]
- Apea-Bah, F.B.; Serem, J.C.; Bester, M.J.; Duodu, K.G. Phenolic composition and antioxidant properties of koose, a deep-fat fried cowpea cake. Food Chem. 2017, 237, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Albishi, T.; John, J.A.; Al-Khalifa, A.S.; Shahidi, F. Antioxidant, anti-inflammatory and DNA scission inhibitory activities of phenolic compounds in selected onion and potato varieties. J. Funct. Foods 2013, 5, 930–939. [Google Scholar] [CrossRef]
- Sumczynski, D.; Bubelova, Z.; Sneyd, J.; Erb-Weber, S.; Mlcek, J. Total phenolics, flavonoids, antioxidant activity, crude fibre and digestibility in non-traditional wheat flakes and muesli. Food Chem. 2015, 174, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Alanon, M.E.; Castro-Vazquez, L.; Diaz-Maroto, M.C.; Gordon, M.H.; Perez-Coello, M.S. A study of the antioxidant capacity of oak wood used in wine ageing and the correlation with polyphenol composition. Food Chem. 2011, 128, 997–1002. [Google Scholar] [CrossRef]
- Sun, L.W.; Isaak, C.K.; Zhou, Y.L.; Petkau, J.C.; Karmin, O.; Liu, Y.J.; Siow, Y.L. Salidroside and tyrosol from Rhodiola protect H9c2 cells from ischemia/reperfusion-induced apoptosis. Life Sci. 2012, 91, 151–158. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Organs | Free | Esterified | I-B | Sum |
|---|---|---|---|---|---|
| Total Phenols (μmol Gallic Acid Equivalent per Gram of Dry Weight (μmol GAE/g DW)) | |||||
| Lonicera japonica | Flower | 150.44 ± 1.43 Ca | 38.80 ± 0.90 Cb | 6.16 ± 0.21 Ec | 195.40 |
| Leaf | 98.79 ± 0.98 Da | 21.67 ± 0.30 Dc | 29.97 ± 0.27 Ab | 150.42 | |
| Stem | 15.78 ± 0.34 Ea | 1.84 ± 0.21 Fc | 6.09 ± 0.24 Eb | 23.71 | |
| Average | 88.34 | 20.77 | 14.07 | ||
| Lonicera macranthoides | Flower | 265.13 ± 2.28 Ba | 77.04 ± 0.51 Ab | 11.07 ± 0.26 Dc | 353.25 |
| Leaf | 365.83 ± 6.32 Aa | 59.77 ± 1.04 Bb | 13.54 ± 0.17 Cc | 439.13 | |
| Stem | 97.08 ± 1.21 Da | 12.16 ± 0.52 Ec | 16.10 ± 0.59 Bb | 125.34 | |
| Average | 242.68 | 49.66 | 13.57 | ||
| Total Flavonoids (μmol Rutin Equivalent per Gram of Dry Weight (μmol RE/g DW)) | |||||
| Lonicera japonica | Flower | 60.64 ± 0.42 Ca | 16.39 ± 0.57 Cb | 3.41 ± 0.02 Ec | 80.44 |
| Leaf | 55.09 ± 0.55 Da | 11.63 ± 0.33 Dc | 12.84 ± 0.08 Ab | 79.56 | |
| Stem | 7.87 ± 0.05 Fa | 0.93 ± 0.00 Fc | 2.83 ± 0.05 Fb | 11.62 | |
| Average | 41.20 | 9.65 | 6.36 | ||
| Lonicera macranthoides | Flower | 106.46 ± 0.30 Ba | 33.96 ± 0.35 Ab | 4.87 ± 0.03 Dc | 145.29 |
| Leaf | 166.59 ± 1.87 Aa | 25.88 ± 0.31 Bb | 6.45 ± 0.27 Cc | 198.91 | |
| Stem | 45.96 ± 0.88 Ea | 5.17 ± 0.05 Ec | 7.00 ± 0.11 Bb | 58.13 | |
| Average | 106.34 | 21.67 | 6.11 | ||
| No. | Rt (min) | Error (ppm) | Measured Value (m/z) | MS/MS Fragments m/z | Molecular Formula | Identified Phenols |
|---|---|---|---|---|---|---|
| 1 | 10.10 | −2.3 | 353.09 | 191.05 [M − (caffeic acid − H2O) − H]− 179.03 [M − (quinic acid − H2O) − H]− 135.05 [M − (quinic acid − H2O) − CO2 − H]− | C16H17O9 | 5-CQA |
| 2 | 17.73 | 1.1 | 353.09 | 191.06 [M − (caffeic acid − H2O) − H]− | C16H17O9 | 3-CQA |
| 3 | 20.60 | 0.8 | 353.09 | 173.04 [M − (caffeic acid − H2O) − H2O − H]− 179.04 [M − (quinic acid − H2O) − H]− 191.06 [M − (caffeic acid − H2O) − H]− 135.04 [M − (quinic acid − H2O) − CO2 − H]− | C16H17O9 | 4-CQA |
| 4 | 22.77 | 10.6 | 179.03 | 135.05 [M − CO2 − H]− | C9H7O4 | Caffeic acid |
| 5 | 61.17 | 2.9 | 515.12 | 179.04 [M − (caffeic acid-H2O) − (quinic acid − H2O) − H]− 173.05 [M − 2(caffeic acid − H2O) − H2O − H]− 353.09 [M − (caffeic acid − H2O) − H]− 191.06 [M − 2(caffeic acid − H2O) − H]− | C25H23O12 | 3,4-diCQA |
| 6 | 62.88 | 3.1 | 447.09 | 285.04 [M − (Glu − H2O) − H]− | C21H19O11 | Luteoloside |
| 7 | 64.46 | −2.7 | 515.12 | 191.06 [M − 2(caffeic acid − H2O) − H]− 353.09 [M − (caffeic acid− H2O) − H]− 179.03 [M − (caffeic acid − H2O) − (quinic acid − H2O) − H]− 135.04 [M − (caffeic acid − H2O) − (quinic acid − H2O) − CO2 − H]− | C25H23O12 | 3,5-diCQA |
| 8 | 68.80 | 4.5 | 463.09 | 301.032 [M − (Glu − H2O) − H]− | C21H19O12 | Isoquercitrin |
| 9 | 71.40 | 0.7 | 593.15 | 285.04 [M − (rutinose − H2O) − H]− | C27H29O15 | Lonicerin |
| 10 | 85.76 | −4.1 | 515.12 | 173.05 [M − 2(caffeic acid − H2O) − H2O − H]− 353.09 [M − (caffeic acid − H2O) − H]− 179.03 [M − (caffeic acid − H2O) − (quinic acid − H2O) − H]− 191.0551 [M − 2(caffeic acid − H2O) − H]− | C25H23O12 | 4,5-diCQA |
| No. | Phenolics | Lonicera japonica | Lonicera macranthoides | ||||
|---|---|---|---|---|---|---|---|
| Free | Esterified | I-B | Free | Esterified | I-B | ||
| Flower | |||||||
| 1 | 5-CQA | 1.61 ± 0.05 c | - | - | 2.80 ± 0.12 b | - | - |
| 2 | 3-CQA | 23.49 ± 0.69 c | - | - | 42.14 ± 1.42 b | - | - |
| 3 | 4-CQA | 3.06 ± 0.08 b | - | - | 2.50 ± 0.07 c | - | - |
| 4 | Caffeic acid | 0.54 ± 0.01 i | 4.96 ± 0.11 b | 0.97 ± 0.02 g | 0.60 ± 0.10 i | 10.42 ± 0.16 a | 0.83 ± 0.01 h |
| 5 | 3,4-diCQA | 3.58 ± 0.04 a | - | 3.02 ± 0.12 c | - | - | |
| 6 | Luteoloside | 1.54 ± 0.02 c | 1.24 ± 0.04 d | - | 1.16 ± 0.08 defg | 1.15 ± 0.00 defg | - |
| 7 | 3,5-diCQA | 12.81 ± 0.57 c | - | - | 26.19 ± 1.82 a | - | - |
| 8 | Isoquercitrin | 1.19 ± 0.02 c | 1.00 ± 0.01 e | - | 1.28 ± 0.04 b | 0.99 ± 0.01 e | - |
| 9 | Lonicerin | 0.87 ± 0.05 de | - | - | 0.95 ± 0.07 d | - | - |
| 10 | 4,5-diCQA | 3.71 ± 0.21 c | - | - | 5.92 ± 0.53 b | - | - |
| Sum | 52.40 | 7.20 | 0.97 | 86.55 | 12.56 | 0.83 | |
| Leaf | |||||||
| 1 | 5-CQA | 0.85 ± 0.05 e | - | - | 11.73 ± 0.28 a | - | - |
| 2 | 3-CQA | 5.39 ± 0.14 d | - | - | 63.29 ± 2.18 a | - | - |
| 3 | 4-CQA | 0.81 ± 0.01 e | - | - | 8.82 ± 0.12 a | - | - |
| 4 | Caffeic acid | 0.50 ± 0.01 i | 1.28 ± 0.03 e | 2.68 ± 0.05 d | 0.56 ± 0.04 i | 4.72 ± 0.05 c | 1.15 ± 0.02 f |
| 5 | 3,4-diCQA | 1.68 ± 0.03 e | - | - | 3.36 ± 0.03 b | - | - |
| 6 | Luteoloside | 2.65 ± 0.08 a | 1.18 ± 0.01 def | 1.24 ± 0.01 d | 2.04 ± 0.03 b | 1.21 ± 0.00 de | 1.10 ± 0.01 fg |
| 7 | 3,5-diCQA | 11.10 ± 0.36 d | - | - | 16.25 ± 0.58 b | - | - |
| 8 | Isoquercitrin | - | - | - | 1.65 ± 0.04 a | 1.03 ± 0.01 e | 0.93 ± 0.01 f |
| 9 | Lonicerin | 2.50 ± 0.10 a | 1.51 ± 0.06 b | - | 1.07 ± 0.05 c | 0.73 ± 0.01 f | - |
| 10 | 4,5-diCQA | 2.27 ± 0.09 d | - | - | 7.20 ± 0.15 a | - | - |
| Sum | 27.74 | 3.98 | 3.92 | 115.97 | 7.69 | 3.19 | |
| Stem | |||||||
| 1 | 5-CQA | 0.51 ± 0.00 f | - | - | 1.14 ± 0.04 d | - | - |
| 2 | 3-CQA | 0.87 ± 0.02 e | - | - | 4.77 ± 0.24 d | - | - |
| 3 | 4-CQA | 0.60 ± 0.00 f | - | - | 1.22 ± 0.02 d | - | - |
| 4 | Caffeic acid | 0.51 ± 0.00 i | - | 0.59 ± 0.01 i | 0.74 ± 0.01 h | 0.84 ± 0.01 h | 1.21 ± 0.01 ef |
| 5 | 3,4-diCQA | 0.88 ± 0.01 f | - | - | 2.21 ± 0.15 d | - | - |
| 6 | Luteoloside | - | - | - | 1.08 ± 0.00 g | 1.13 ± 0.00 efg | 1.11 ± 0.01 fg |
| 7 | 3,5-diCQA | 0.60 ± 0.01 f | - | - | 6.09 ± 0.66 e | - | - |
| 8 | Isoquercitrin | - | - | - | 1.09 ± 0.02 d | - | - |
| 9 | Lonicerin | - | - | - | 0.79 ± 0.04 ef | - | - |
| 10 | 4,5-diCQA | 0.71 ± 0.02 e | - | - | 2.65 ± 0.29 d | - | - |
| Sum | 4.67 | - | 0.59 | 21.78 | 1.97 | 2.32 | |
| Species | Organs | Free | Esterified | I-B | Sum |
|---|---|---|---|---|---|
| DPPH | |||||
| Lonicera japonica | Flower | 178.60 ± 2.29 Ca | 29.91 ± 0.96 Cb | 10.58 ± 0.62 Dc | 219.08 |
| Leaf | 98.25 ± 3.19 Da | 18.36 ± 0.36 Dc | 32.86 ± 0.66 Ab | 149.46 | |
| Stem | 11.18 ± 0.40 Ea | 1.07 ± 0.70 Fc | 7.89 ± 0.27 Eb | 20.14 | |
| Average | 96.01 | 16.45 | 17.11 | ||
| Lonicera macranthoides | Flower | 279.14 ± 1.85 Ba | 88.07 ± 2.22 Ab | 12.22 ± 0.53 Cc | 379.44 |
| Leaf | 293.08 ± 5.98 Aa | 53.98 ± 2.02 Bb | 12.02 ± 0.53 Dc | 359.07 | |
| Stem | 99.77 ± 7.70 Da | 8.82 ± 0.28 Eb | 15.22 ± 0.44 Bb | 123.81 | |
| Average | 224.00 | 50.29 | 13.15 | ||
| ABTS | |||||
| Lonicera japonica | Flower | 132.96 ± 1.37 Ca | 41.04 ± 0.37 Cb | 11.70 ± 0.63 Dc | 185.70 |
| Leaf | 113.67 ± 2.41 Da | 22.72 ± 0.36 Dc | 43.28 ± 1.19 Ab | 179.67 | |
| Stem | 23.67 ± 0.64 Ea | 2.21 ± 0.07 Fc | 10.55 ± 0.22 Eb | 36.43 | |
| Average | 90.10 | 21.99 | 21.84 | ||
| Lonicera macranthoides | Flower | 238.96 ± 7.79 Ba | 95.80 ± 1.23 Ab | 19.92 ± 0.53 Cc | 354.68 |
| Leaf | 329.08 ± 5.09 Aa | 77.33 ± 0.52 Bb | 20.21 ± 0.09 Cc | 426.61 | |
| Stem | 108.28 ± 1.40 Da | 16.39 ± 0.12 Ec | 22.54 ± 0.37 Bb | 147.20 | |
| Average | 225.44 | 63.17 | 20.76 | ||
| ORAC | |||||
| Lonicera japonica | Flower | 1758.76 ± 29.04 Ca | 532.55 ± 5.87 Cb | 122.56 ± 16.85 Ec | 2413.87 |
| Leaf | 1269.89 ± 77.93 Da | 331.75 ± 10.69 Dc | 453.83 ± 7.31 Ab | 2055.47 | |
| Stem | 284.88 ± 13.82 Ea | 28.30 ± 6.78 Fc | 97.68 ± 5.11 Fb | 410.86 | |
| Average | 1104.51 | 297.53 | 224.69 | ||
| Lonicera macranthoides | Flower | 2910.83 ± 100.44 Ba | 1083.59 ± 2.65 Ab | 180.12 ± 12.08 Dc | 4174.54 |
| Leaf | 4637.35 ± 137.41 Aa | 942.67 ± 34.71 Bb | 248.16 ± 1.62 Cc | 5828.19 | |
| Stem | 1133.83 ± 80.79 Da | 211.11 ± 9.11 Eb | 272.37 ± 17.40 Bb | 1617.30 | |
| Average | 2894.00 | 745.79 | 233.55 | ||
| Phenolics * | Contribution | |||||
|---|---|---|---|---|---|---|
| Flower | Leaf | Stem | ||||
| L. japonica | L. macranthoides | L. japonica | L. macranthoides | L. japonica | L. macranthoides | |
| DPPH activity | ||||||
| Caffeic acid (4) ** | 1.90 ± 0.05 | 1.26 ± 0.04 | 3.33 ± 0.09 | 0.88 ± 0.02 | 19.93 ± 0.56 | 6.26 ± 0.18 |
| Luteoloside (6) | 4.62 ± 0.04 | 2.10 ± 0.02 | 15.00 ± 0.12 | 2.77 ± 0.02 | - | 7.79 ± 0.06 |
| Lonicerin (9) | 2.17 ± 0.04 | 1.43 ± 0.03 | 11.78 ± 0.24 | 1.20 ± 0.02 | - | 4.72 ± 0.10 |
| Isoquercitrin (8) | 2.84 ± 0.04 | 1.85 ± 0.03 | - | 1.79 ± 0.02 | - | 6.26 ± 0.09 |
| 3,4-CQA (5) | 8.84 ± 0.14 | 4.52 ± 0.07 | 7.82 ± 0.13 | 3.75 ± 0.06 | 24.52 ± 0.39 | 13.12 ± 0.21 |
| 3,5-CQA (7) | 27.95 ± 0.17 | 34.60 ± 0.21 | 45.73 ± 0.28 | 16.02 ± 0.10 | 14.59 ± 0.09 | 31.98 ± 0.19 |
| 4,5-CQA (10) | 7.49 ± 0.09 | 7.24 ± 0.09 | 8.66 ± 0.11 | 6.58 ± 0.08 | 16.10 ± 0.20 | 12.89 ± 0.16 |
| 3-CQA (2) | 30.24 ± 0.27 | 32.84 ± 0.30 | 13.09 ± 0.12 | 36.81 ± 0.33 | 12.53 ± 0.11 | 14.79 ± 0.13 |
| 5-CQA (1) | 1.85 ± 0.02 | 1.95 ± 0.03 | 1.84 ± 0.02 | 6.09 ± 0.08 | 6.51 ± 0.09 | 3.15 ± 0.04 |
| 4-CQA (3) | 2.51 ± 0.13 | 1.24 ± 0.07 | 1.26 ± 0.07 | 3.27 ± 0.17 | 5.50 ± 0.29 | 2.40 ± 0.13 |
| ABTS activity | ||||||
| Caffeic acid (4) | 1.06 ± 0.00 | 0.71 ± 0.00 | 1.86 ± 0.00 | 0.49 ± 0.00 | 11.13 ± 0.00 | 3.50 ± 0.00 |
| Luteoloside (6) | 1.84 ± 0.02 | 0.83 ± 0.01 | 5.97 ± 0.05 | 1.10 ± 0.01 | - | 3.10 ± 0.03 |
| Lonicerin (9) | 0.83 ± 0.01 | 0.55 ± 0.00 | 4.53 ± 0.04 | 0.46 ± 0.00 | - | 1.81 ± 0.02 |
| Isoquercitrin (8) | 1.13 ± 0.01 | 0.74 ± 0.01 | - | 0.71 ± 0.01 | - | 2.50 ± 0.03 |
| 3,4-CQA (5) | 6.66 ± 0.06 | 3.40 ± 0.03 | 5.89 ± 0.05 | 2.82 ± 0.03 | 18.46 ± 0.17 | 9.88 ± 0.09 |
| 3,5-CQA (7) | 21.38 ± 0.22 | 26.46 ± 0.27 | 34.98 ± 0.35 | 12.25 ± 0.12 | 11.16 ± 0.11 | 24.46 ± 0.25 |
| 4,5-CQA (10) | 7.09 ± 0.02 | 6.85 ± 0.02 | 8.19 ± 0.02 | 6.22 ± 0.02 | 15.23 ± 0.04 | 12.19 ± 0.03 |
| 3-CQA (2) | 21.75 ± 0.33 | 23.62 ± 0.36 | 9.42 ± 0.14 | 26.47 ± 0.40 | 9.01 ± 0.14 | 10.63 ± 0.16 |
| 5-CQA (1) | 1.64 ± 0.02 | 1.73 ± 0.02 | 1.63 ± 0.02 | 5.40 ± 0.07 | 5.78 ± 0.07 | 2.79 ± 0.04 |
| 4-CQA (3) | 2.83 ± 0.02 | 1.40 ± 0.01 | 1.42 ± 0.01 | 3.68 ± 0.03 | 6.20 ± 0.04 | 2.71 ± 0.02 |
| Parameters | TP | TF | DPPH | ABTS | ORAC | IDD | TQP | CA | Lu | Iq | Lo | 5-C | 3-C | 4-C | 3,4-dC | 3,5-dC | 4,5-dC |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Free Phenolics | |||||||||||||||||
| Total phenols (TP) | 1 | ||||||||||||||||
| Total flavonoids (TF) | 0.990 ** | 1 | |||||||||||||||
| DPPH | 0.967 ** | 0.930 ** | 1 | ||||||||||||||
| ABTS | 0.996 ** | 0.996 ** | 0.955 ** | 1 | |||||||||||||
| ORAC | 0.994 ** | 0.996 ** | 0.935 ** | 0.992 ** | 1 | ||||||||||||
| Inhibition to DNA damage (IDD) | −0.745 | −0.738 | −0.825 * | −0.761 | −0.711 | 1 | |||||||||||
| Total quantified phenolics (TQP) | 0.994 ** | 0.976 ** | 0.973 ** | 0.983 ** | 0.985 ** | −0.731 | 1 | ||||||||||
| caffeic acid (CA) | 0.061 | 0.040 | 0.095 | 0.087 | 0.022 | −0.234 | −0.011 | 1 | |||||||||
| Luteoloside (Lu) | 0.432 | 0.508 | 0.416 | 0.478 | 0.454 | −0.727 | 0.407 | −0.190 | 1 | ||||||||
| Isoquercitrin (Iq) | 0.818 * | 0.771 | 0.846 * | 0.799 | 0.790 | −0.725 | 0.807 | 0.470 | 0.160 | 1 | |||||||
| Lonicerin (Lo) | 0.144 | 0.223 | 0.154 | 0.203 | 0.154 | −0.525 | 0.116 | −0.222 | 0.916 * | −0.172 | 1 | ||||||
| 5-CQA (5-C) | 0.870 * | 0.906 * | 0.723 | 0.872 * | 0.917 ** | −0.453 | 0.851 * | −0.037 | 0.350 | 0.660 | 0.025 | 1 | |||||
| 3-CQA (3-C) | 0.983 ** | 0.962 ** | 0.945 ** | 0.965 ** | 0.980 ** | −0.647 | 0.992 ** | −0.035 | 0.318 | 0.802 | 0.013 | 0.882 * | 1 | ||||
| 4-CQA (4-C) | 0.882 * | 0.903 * | 0.766 | 0.872 * | 0.923 ** | −0.525 | 0.877 * | −0.055 | 0.361 | 0.737 | −0.005 | 0.978 ** | 0.906 * | 1 | |||
| 3,4-diCQA (3,4-dC) | 0.779 | 0.730 | 0.864 * | 0.753 | 0.747 | −0.859 * | 0.798 | 0.174 | 0.401 | 0.894 * | 0.082 | 0.542 | 0.766 | 0.674 | 1 | ||
| 3,5-diCQA (3,5-dC) | 0.788 | 0.732 | 0.897 * | 0.781 | 0.722 | −0.786 | 0.806 | 0.025 | 0.405 | 0.613 | 0.301 | 0.394 | 0.746 | 0.426 | 0.711 | 1 | |
| 4,5-diCQA (4,5-dC) | 0.993 ** | 0.971 ** | 0.987 ** | 0.987 ** | 0.976 ** | −0.783 | 0.990 ** | 0.122 | 0.405 | 0.859 * | 0.121 | 0.818 * | 0.973 ** | 0.843 * | 0.825 * | 0.828 * | 1 |
| Esterified Phenolics | |||||||||||||||||
| Total phenols | 1 | ||||||||||||||||
| Total flavonoids | 0.997 ** | 1 | |||||||||||||||
| DPPH | 0.982 ** | 0.983 ** | 1 | ||||||||||||||
| ABTS | 0.995 ** | 0.990 ** | 0.982 ** | 1 | |||||||||||||
| ORAC | 0.995 ** | 0.992 ** | 0.969 ** | 0.997 ** | 1 | ||||||||||||
| Inhibition to DNA damage | −0.740 | −0.749 | −0.656 | −0.712 | −0.752 | 1 | |||||||||||
| Total quantified phenolics | 0.977 ** | 0.978 ** | 0.967 ** | 0.954 ** | 0.954 ** | −0.762 | 1 | ||||||||||
| caffeic acid | 0.947 ** | 0.940 ** | 0.965 ** | 0.929 ** | 0.913 * | −0.630 | 0.977 ** | 1 | |||||||||
| Luteoloside | 0.574 | 0.584 | 0.483 | 0.543 | 0.589 | −0.974 ** | 0.611 | 0.470 | 1 | ||||||||
| Isoquercitrin | 0.880 * | 0.851 * | 0.801 | 0.858 * | 0.871 * | −0.642 | 0.860 * | 0.835 * | 0.487 | 1 | |||||||
| Lonicerin | −0.028 | 0.035 | −0.074 | −0.040 | 0.016 | −0.273 | −0.059 | −0.239 | 0.301 | 0.835 * | 1 | ||||||
| Insoluble-Bound Phenolics | |||||||||||||||||
| Total phenols | 1 | ||||||||||||||||
| Total flavonoids | 0.997 ** | 1 | |||||||||||||||
| DPPH | 0.969 ** | 0.968 ** | 1 | ||||||||||||||
| ABTS | 0.993 ** | 0.989 ** | 0.977 ** | 1 | |||||||||||||
| ORAC | 0.993 ** | 0.996 ** | 0.945 ** | 0.979 ** | 1 | ||||||||||||
| Inhibition to DNA damage | −0.883 * | −0.895 * | −0.846 * | −0.892 * | −0.911 * | 1 | |||||||||||
| Total quantified phenolics | 0.870 * | 0.895 * | 0.779 | 0.831 * | 0.914 * | −0.806 | 1 | ||||||||||
| caffeic acid | 0.958 ** | 0.968 ** | 0.987 ** | 0.958 ** | 0.948 ** | −0.849 * | 0.841 * | 1 | |||||||||
| Luteoloside | 0.795 | 0.810 | 0.656 | 0.730 | 0.850 * | −0.741 | 0.947 ** | 0.709 | 1 | ||||||||
| Isoquercitrin | −0.016 | 0.029 | −0.169 | −0.048 | 0.072 | −0.055 | 0.431 | −0.058 | 0.407 | 1 | |||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, M.; Yang, L.; Xue, Q.; Yin, P.; Sun, L.; Liu, Y. Comparison of Free, Esterified, and Insoluble-Bound Phenolics and Their Bioactivities in Three Organs of Lonicera japonica and L. macranthoides. Molecules 2019, 24, 970. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24050970
Yu M, Yang L, Xue Q, Yin P, Sun L, Liu Y. Comparison of Free, Esterified, and Insoluble-Bound Phenolics and Their Bioactivities in Three Organs of Lonicera japonica and L. macranthoides. Molecules. 2019; 24(5):970. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24050970
Chicago/Turabian StyleYu, Miao, Lingguang Yang, Qiang Xue, Peipei Yin, Liwei Sun, and Yujun Liu. 2019. "Comparison of Free, Esterified, and Insoluble-Bound Phenolics and Their Bioactivities in Three Organs of Lonicera japonica and L. macranthoides" Molecules 24, no. 5: 970. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24050970