
The Effect of Gelatine Packaging Film Containing a Spirulina platensis Protein Concentrate on Atlantic Mackerel Shelf Life


 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
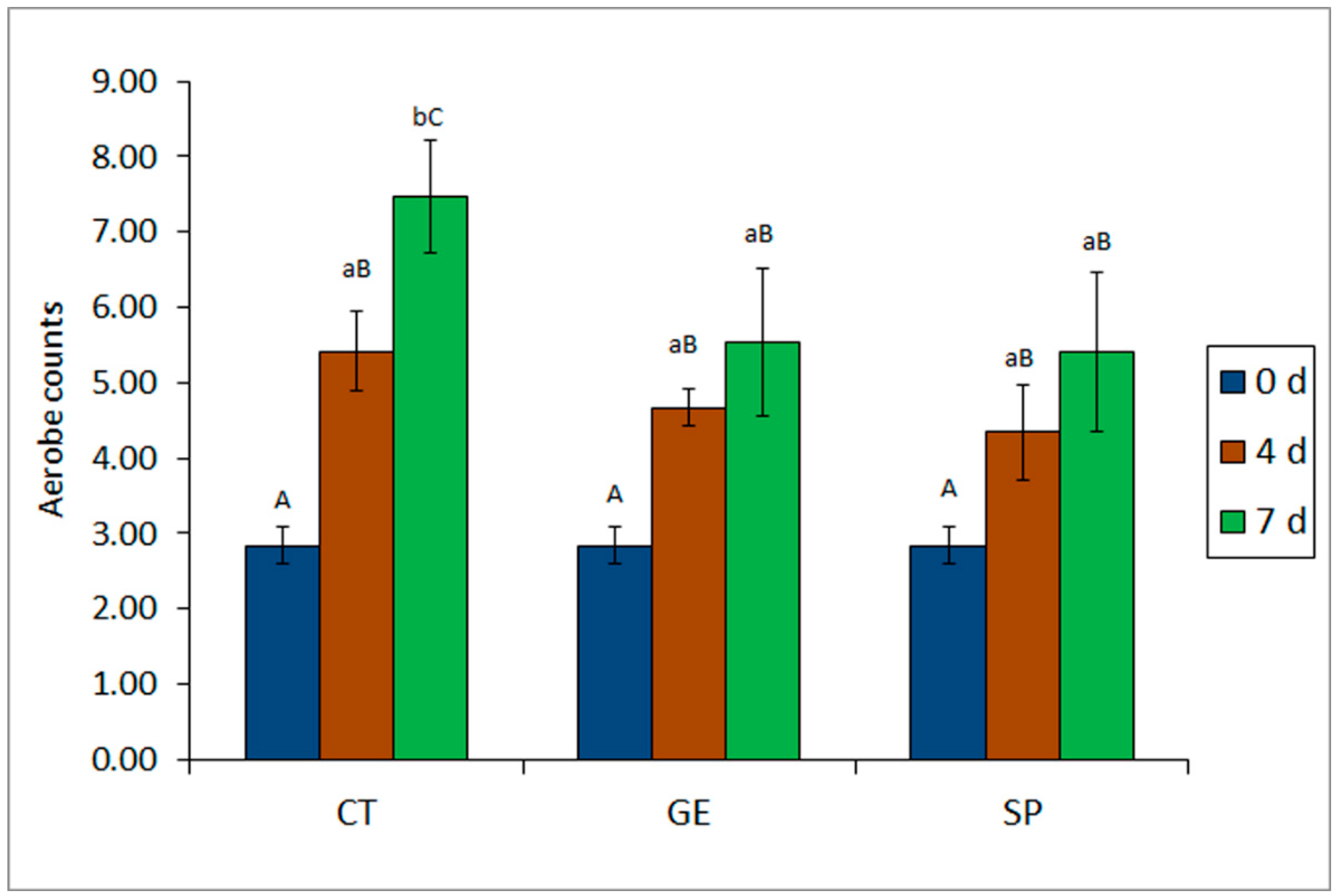
2.1. Comparative Analysis of Microbial Development in Fish Batches
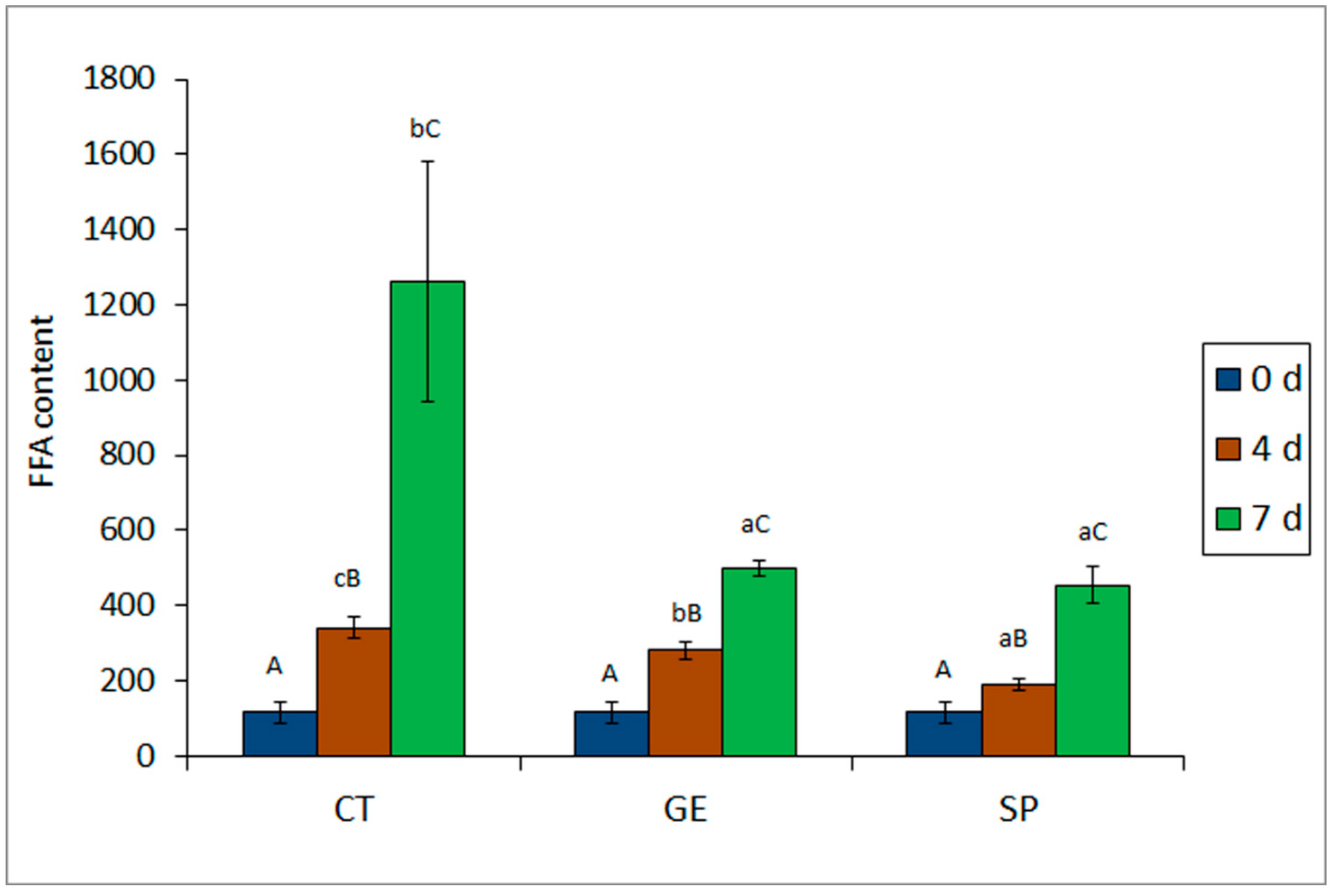
2.2. Chemical Analyses of Quality Deterioration
3. Materials and Methods
3.1. Comparative Analysis
3.2. Preparation and Physicochemical Characteristics of Film Systems
3.3. Fish Material, Processing and Sampling
3.4. Microbial Analyses
3.5. Chemical Analyses of Quality Deterioration
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tacon, A.; Metian, M. Fish Matters: Importance of Aquatic Foods in Human Nutrition and Global Food Supply. Rev. Fish. Sci. 2013, 21, 22–38. [Google Scholar] [CrossRef]
- Tilami, S.K.; Sampels, S. Nutritional Value of Fish: Lipids, Proteins, Vitamins, and Minerals. Rev. Fish. Sci. 2018, 26, 242–253. [Google Scholar]
- Dovale-Rosabal, G.; Rodríguez, A.; Contreras, E.; Ortiz-Viedma, J.; Muñoz, M.; Trigo, M.; Aubourg, S.P.; Espinosa, A. Concentration of EPA and DHA from refined salmon oil by optimizing the urea-fatty acid adduction reaction conditions using response-surface methodology. Molecules 2019, 24, 1642. [Google Scholar] [CrossRef] [Green Version]
- Ashie, I.; Smith, J.; Simpson, B. Spoilage and shelf-life extension of fresh fish and shellfish. Crit. Rev. Food Sci. Nutr. 1996, 36, 87–121. [Google Scholar] [CrossRef]
- López-Rubio, A.; Gavara, R.; Lagarón, J. Bioactive packaging: Turning foods into healthier foods through biomaterials. Trends Food Sci. Technol. 2006, 17, 567–575. [Google Scholar] [CrossRef]
- Umaraw, P.; Munekata, P.E.S.; Verma, A.K.; Barba, F.J.; Singh, V.P.; Kumar, P.; Lorenzo, J.M. Edible films/coating with tailored properties for active packaging of meat, fish and derived products. Trends Food Sci. Technol. 2020, 98, 10–24. [Google Scholar] [CrossRef]
- Giménez, B.; López de Lacey, A.; Pérez-Santín, E.; López-Caballero, M.E.; Montero, P. Release of active compounds from agar and agar gelatin films with green tea extract. Food Hydroc. 2013, 30, 264–271. [Google Scholar] [CrossRef]
- Martucci, J.; Gende, L.; Neira, L.; Ruseckaite, R. Oregano and lavender essential oils as antioxidant and antimicrobial additives of biogenic gelatin films. Ind. Crops Prod. 2015, 71, 205–213. [Google Scholar] [CrossRef]
- Jafarzadeh, S.; Jafari, S.M.; Salejhabadi, A.; Nafchi, A.M.; Kumar, S.U.; Khalil, H.P.S. Biodegradable green packaging with antimicrobial functions based on the bioactive compounds from tropical plants and their by-products. Trend Food Sci. Technol. 2020, 100, 262–277. [Google Scholar] [CrossRef]
- Sudhakar, M.P.; Ramesh Kumar, B.; Mathimani, T.; Arunkuma, K. A review on bioenergy and bioactive compounds from microalgae and macroalgae-sustainable energy perspective. J. Clean. Prod. 2019, 228, 1320–1333. [Google Scholar] [CrossRef]
- Syaichurrozi, I.; Sumardiono, S.; Budi Sasongko, S. Production of Spirulina platensis biomass using digested vinasse as cultivation medium. Trends Appl. Sci. Res. 2014, 9, 93–102. [Google Scholar]
- Ovando, C.; de Carvalho, J.; de Melo Pereira, G.; Jacques, P.; Soccol, V.; Soccol, C. Functional properties and health benefits of bioactive peptides derived from Spirulina: A review. Food Rev. Int. 2018, 34, 34–51. [Google Scholar] [CrossRef]
- Glazer, A. Phycobiliproteins-A family of valuable, widely used fluorophores. J. Appl. Phycol. 1994, 6, 105–112. [Google Scholar] [CrossRef]
- Pagels, F.; Guedes, C.; Amaro, H.; Kijjoa, A.; Vasconcelos, V. Phycobiliproteins from cyanobacteria: Chemistry and biotechnological applications. Biotechnol. Adv. 2019, 37, 422–443. [Google Scholar] [CrossRef] [PubMed]
- Hirata, T.; Tanaka, M.; Ooike, M.; Tsunomura, T.; Skaguchi, M. Antioxidant activities of phycocyanobilin prepared from Spirulina platensis. J. Appl. Phycol. 2000, 12, 439–465. [Google Scholar] [CrossRef]
- Chentir, I.; Hamdi, M.; Li, S.; Doumandji, A.; Markou, G.; Nasri, M. Stability, bio-functionality, and bio-activity of crude phycocyanin from a two-phase cultured Saharan Arthrospira sp. strain. Algal Res. 2018, 35, 395–406. [Google Scholar] [CrossRef]
- Benelhadj, S.; Gharsallaoui, A.; Degraeve, P.; Attia, H.; Ghorbel, D. Effect of pH on the functional properties of Arthrospira (Spirulina platensis) protein isolate. Food Chem. 2016, 194, 1056–1063. [Google Scholar] [CrossRef]
- Stejskal, N.; Ruseckaite, R.A.; Martucci, J.F. Spirulina platensis as potential alternative bio-feedstock of sustainable active films. In Proceedings of the Conference Compass ECO-BIO 2018, Dublin, Ireland, 4–7 March 2018. [Google Scholar]
- Stejskal, N.; Ruseckaite, R.A.; Martucci, J.F. Development and characterisation of active films based on gelatine and algal protein. In Proceedings of the XXXII Chemical Argentinian Congress, Buenos Aires, Argentina, 12–15 March 2019. [Google Scholar]
- Stejskal, N.; Miranda, J.M.; Martucci, J.F.; Ruseckaite, R.A.; Barros-Velázquez, J.; Aubourg, S.P. Quality enhancement of refrigerated hake muscle by active packaging with a protein concentrate from Spirulina platensis. Food Bioprocess Technol. 2020, 13, 1110–1118. [Google Scholar] [CrossRef]
- Leu, S.; Jhaveri, S.; Karakoltsidis, P.; Constantinides, S. Atlantic mackerel (Scomber scombrus, L.): Seasonal variation in proximate composition and distribution of chemical nutrients. J. Food Sci. 1981, 46, 1635–1638. [Google Scholar] [CrossRef]
- Martelo-Vidal, M.J.; Mesas, J.M.; Vázquez, M. Low-salt restructured fish products from Atlantic mackerel (Scomber scombrus) with texture resembling turkey breast. Food Sci. Techn. Internat. 2012, 18, 251–259. [Google Scholar] [CrossRef]
- Saeed, S.; Howell, N. 12-lipoxygenase activity in the muscle tissue of Atlantic mackerel (Scomber scombrus) and its prevention by antioxidants. J. Sci. Food Agric. 2013, 81, 745–750. [Google Scholar] [CrossRef]
- Senturk, T.; Alpas, H. Effect of high hydrostatic pressure treatment (HHPT) on quality and shelf life of Atlantic mackerel (Scomber scombrus). Food Bioprocess Technol. 2013, 6, 2306–2318. [Google Scholar] [CrossRef]
- Rodríguez, O.; Barros-Velázquez, J.; Ojea, A.; Piñeiro, C.; Aubourg, S.P. Evaluation of sensory and microbiological changes and identification of proteolytic bacteria during the iced storage of farmed turbot (Psetta maxima). J. Food Sci. 2003, 68, 2764–2771. [Google Scholar] [CrossRef]
- Campos, C.; Gliemmo, M.; Aubourg, S.P.; Barros-Velázquez, J. Novel technologies for the preservation of chilled aquatic food products. In Novel Technologies in Food Science; McElhatton, A., Amaral Sobral, P., Eds.; Springer: New York, NY, USA, 2012; Chapter 13; pp. 299–323. [Google Scholar]
- Castro-Enríquez, D.D.; Miranda, J.M.; Barros-Velázquez, J.; Aubourg, S.P. Quality increase of refrigerated fish by employment of a gelatine biofilm including a protein hydrolysate obtained from alga Fucus spiralis. Bulg. Chem. Com. 2020, 52. in press. [Google Scholar]
- Sun, Y.; Chang, R.; Li, Q.; Li, B. Isolation and characterization of an antibacterial peptide from protein hydrolysates of Spirulina platensis. Eur. Food Res. Technol. 2016, 242, 685–692. [Google Scholar] [CrossRef]
- Sadeghi, S.; Jalili, H.; Siadat, S.; Sedighi, M. Anticancer and antibacterial properties in peptide fractions from hydrolyzed spirulina protein. J. Agric. Sci. Technol. 2018, 20, 673–683. [Google Scholar]
- Ruiz-Capillas, C.; Moral, A. Correlation between biochemical and sensory quality indices in hake stored in ice. Food Res. Int. 2001, 34, 441–447. [Google Scholar] [CrossRef] [Green Version]
- García-Soto, B.; Miranda, J.M.; Rodríguez-Bernaldo de Quirós, A.; Sendón, R.; Rodríguez-Martínez, A.; Barros-Velázquez, J.; Aubourg, S.P. Effect of biodegradable film (lyophilised alga Fucus spiralis and sorbic acid) on quality properties of refrigerated megrim (Lepidorhombus whiffiagonis). Int. J. Food Sci. Technol. 2015, 50, 1891–1900. [Google Scholar] [CrossRef]
- Kolakowska, A. Lipid oxidation in food systems. In Chemical and Functional Properties of Food Lipids; Sikorski, Z., Kolakowsa, A., Eds.; CRC Press: London, UK, 2003; pp. 133–165. [Google Scholar]
- Aubourg, S.P.; Rodríguez, A.; Gallardo, J. Rancidity development during mackerel (Scomber scombrus) frozen storage: Effect of catching season and commercial presentation. Eur. J. Lipid Sci. Technol. 2005, 107, 316–323. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Hu, Y.; Xue, M.; Dun, Y.; Li, S.; Peng, N.; Liang, Y.; Zhao, S. Purification and identification of antioxidant peptides from enzymatic hydrolysate of Spirulina platensis. J. Microb. Biotechnol. 2016, 26, 1216–1223. [Google Scholar] [CrossRef] [Green Version]
- Andrade, I.; Andrade, C.; Dias, M.; Nascimento, C.; Mendes, M. Chlorella and Spirulina microalgae as sources of functional foods, nutraceuticals and food supplements; an overview. Med Crave Open J. Food Proc. Technol. 2018, 6, 45–58. [Google Scholar] [CrossRef] [Green Version]
- Bertolin, T.; Guarientio, C.; Farias, D.; Souza, F.; Gutkoski, L.; Colla, L. Antioxidant effect of phycocyanin on dried-salted fish. Ciência e Agrotecnologia Lavras 2011, 35, 751–757. [Google Scholar] [CrossRef] [Green Version]
- Chronakis, I.S. Gelation of edible blue-green algae protein isolate (Spirulina platensis strain Pacifica): Thermal transitions, rheological, properties, and molecular forces involved. J. Agric. Food Chem. 2001, 49, 888–898. [Google Scholar] [CrossRef]
- Neira, L.M.; Agustinelli, S.P.; Ruseckaite, R.A.; Martucci, J.F. Shelf life extension of refrigerated breaded hake medallions packed into active edible fish gelatin films. Pack. Technol. Sci. 2019, 32, 471–480. [Google Scholar] [CrossRef]
- Rodríguez, A.; Cruz, J.M.; Paseiro-Losada, P.; Aubourg, S.P. Effect of a polyphenol-vacuum packaging on lipid deterioration during an 18-month frozen storage of Coho salmon (Oncorhynchus kisutch). Food Bioproc. Technol. 2012, 5, 2602–2611. [Google Scholar] [CrossRef] [Green Version]
- Ben-Gigirey, B.; Vieites Baptista de Sousa, J.; Villa, T.; Barros-Velázquez, J. Changes in biogenic amines and microbiological analysis in albacore (Thunnus alalunga) muscle during frozen storage. J. Food Prot. 1998, 61, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Ben-Gigirey, B.; Vieites Baptista de Sousa, J.; Villa, T.; Barros-Velázquez, J. Histamine and cadaverine production by bacteria isolated from fresh and frozen albacore (Thunnus alalunga). J. Food Prot. 1999, 62, 933–939. [Google Scholar] [CrossRef]
- Ben-Gigirey, B.; Vieites Baptista de Sousa, J.; Villa, T.; Barros-Velázquez, J. Characterization of biogenic amine-producing Stenotrophomonas maltophilia strains isolated from white muscle of fresh and frozen albacore tuna. Int. J. Food Microb. 2000, 57, 19–31. [Google Scholar] [CrossRef]
- Tozawa, H.; Erokibara, K.; Amano, K. Proposed modification of Dyer’s method for trimethylamine determination in codfish. In Fish Inspection and Quality Control; Kreuzer, R., Ed.; Fishing News Books Ltd.: London, UK, 1971; pp. 187–190. [Google Scholar]
- Bligh, E.; Dyer, W. A rapid method of total extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Lowry, R.; Tinsley, I. Rapid colorimetric determination of free fatty acids. J. Am. Oil Chem. Soc. 1976, 53, 470–472. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |



{kind=link}
{kind=link}
| Microbial Index | Refrigeration Time (days) | Packaging Condition | ||
|---|---|---|---|---|
| CT | GE | SP | ||
| Psychrotrophs | 0 | 2.20 A (0.17) | 2.20 A (0.17) | 2.20 A (0.17) |
| 4 | 5.27 aB (1.01) | 4.28 aB (0.72) | 4.59 aB (0.43) | |
| 7 | 8.48 cC (0.16) | 6.59 bC (0.45) | 4.71 aB (0.60) | |
| Enterobacteriaceae | 0 | 1.00 A (0.05) | 1.00 A (0.05) | 1.00 A (0.05) |
| 4 | 2.60 aAB (1.47) | 1.69 aAB (0.65) | 1.79 aAB (0.75) | |
| 7 | 3.46 bB (0.15) | 1.79 aB (0.28) | 1.93 aB (0.81) | |
| Microbial Index | Refrigeration Time (days) | Packaging Condition | ||
|---|---|---|---|---|
| CT | GE | SP | ||
| Proteolytics | 0 | 2.16 A (0.28) | 2.16 A (0.28) | 2.16 A (0.28) |
| 4 | 5.11 bB (0.18) | 4.11 aB (0.35) | 4.78 abB (0.94) | |
| 7 | 7.29 bC (0.12) | 5.71 aC (0.47) | 5.63 aB (0.59) | |
| Lipolytics | 0 | 2.00 A (0.00) | 2.00 A (0.00) | 2.00 A (0.00) |
| 4 | 3.23 aB (0.83) | 2.28 aA (0.49) | 3.13 aB (0.76) | |
| 7 | 5.85 bC (0.13) | 4.26 aB (0.24) | 4.13 aB (0.49) | |
| Chemical Index | Refrigeration Time (days) | Packaging Condition | ||
|---|---|---|---|---|
| CT | GE | SP | ||
| pH | 0 | 6.53 A (0.10) | 6.53 A (0.10) | 6.53 A (0.10) |
| 4 | 6.68 aAB (0.10) | 6.63 aA (0.08) | 6.67 aA (0.07) | |
| 7 | 6.77 aB (0.08) | 6.74 aA (0.15) | 6.69 aA (0.08) | |
| Trimethylamine (TMA) (mg TMA–N kg−1 muscle) | 0 | 1.69 A (0.14) | 1.69 A (0.14) | 1.69 A (0.14) |
| 4 | 33.29 bB (18.41) | 15.64 aB (3.02) | 13.61 aB (2.61) | |
| 7 | 619.69 cC (162.83) | 98.55 bC (16.05) | 72.40 aC (4.53) | |
| FR value | 0 | 1.42 A (0.15) | 1.42 A (0.15) | 1.42 A (0.15) |
| 4 | 2.75 aB (0.23) | 2.46 aB (0.19) | 2.06 aAB (0.53) | |
| 7 | 3.22 aB (0.27) | 2.96 aC (0.14) | 2.83 aB (0.17) | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stejskal, N.; Miranda, J.M.; Martucci, J.F.; Ruseckaite, R.A.; Aubourg, S.P.; Barros-Velázquez, J. The Effect of Gelatine Packaging Film Containing a Spirulina platensis Protein Concentrate on Atlantic Mackerel Shelf Life. Molecules 2020, 25, 3209. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25143209
Stejskal N, Miranda JM, Martucci JF, Ruseckaite RA, Aubourg SP, Barros-Velázquez J. The Effect of Gelatine Packaging Film Containing a Spirulina platensis Protein Concentrate on Atlantic Mackerel Shelf Life. Molecules. 2020; 25(14):3209. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25143209
Chicago/Turabian StyleStejskal, Nadia, José M. Miranda, Josefa F. Martucci, Roxana A. Ruseckaite, Santiago P. Aubourg, and Jorge Barros-Velázquez. 2020. "The Effect of Gelatine Packaging Film Containing a Spirulina platensis Protein Concentrate on Atlantic Mackerel Shelf Life" Molecules 25, no. 14: 3209. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25143209