
In Vitro and In Vivo Biological Activity of Berberine Chloride against Uropathogenic E. coli Strains Using Galleria mellonella as a Host Model

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Berberine MIC Determination
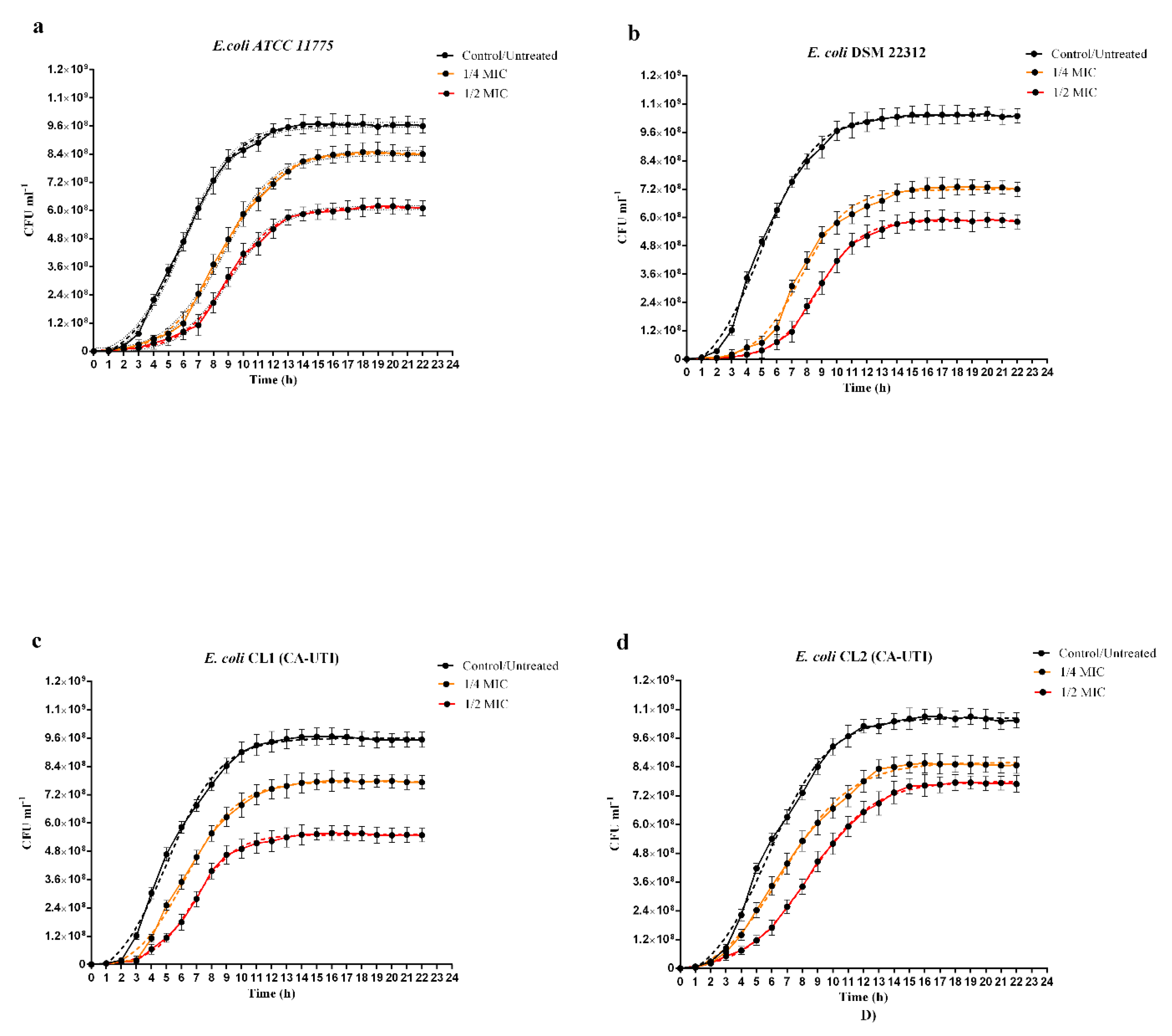
2.2. Influence of Berberine on In Vitro Bacterial Growth
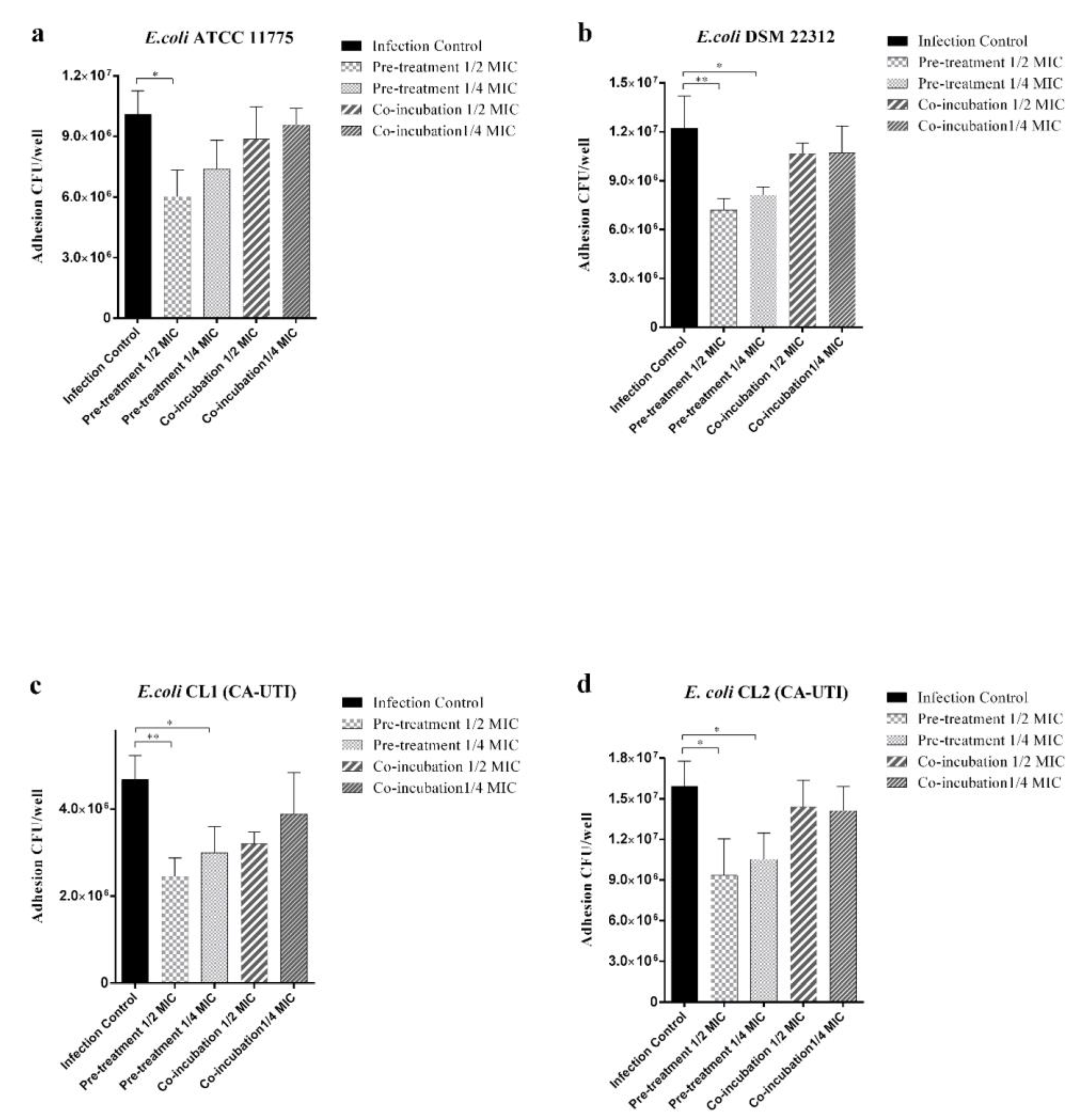
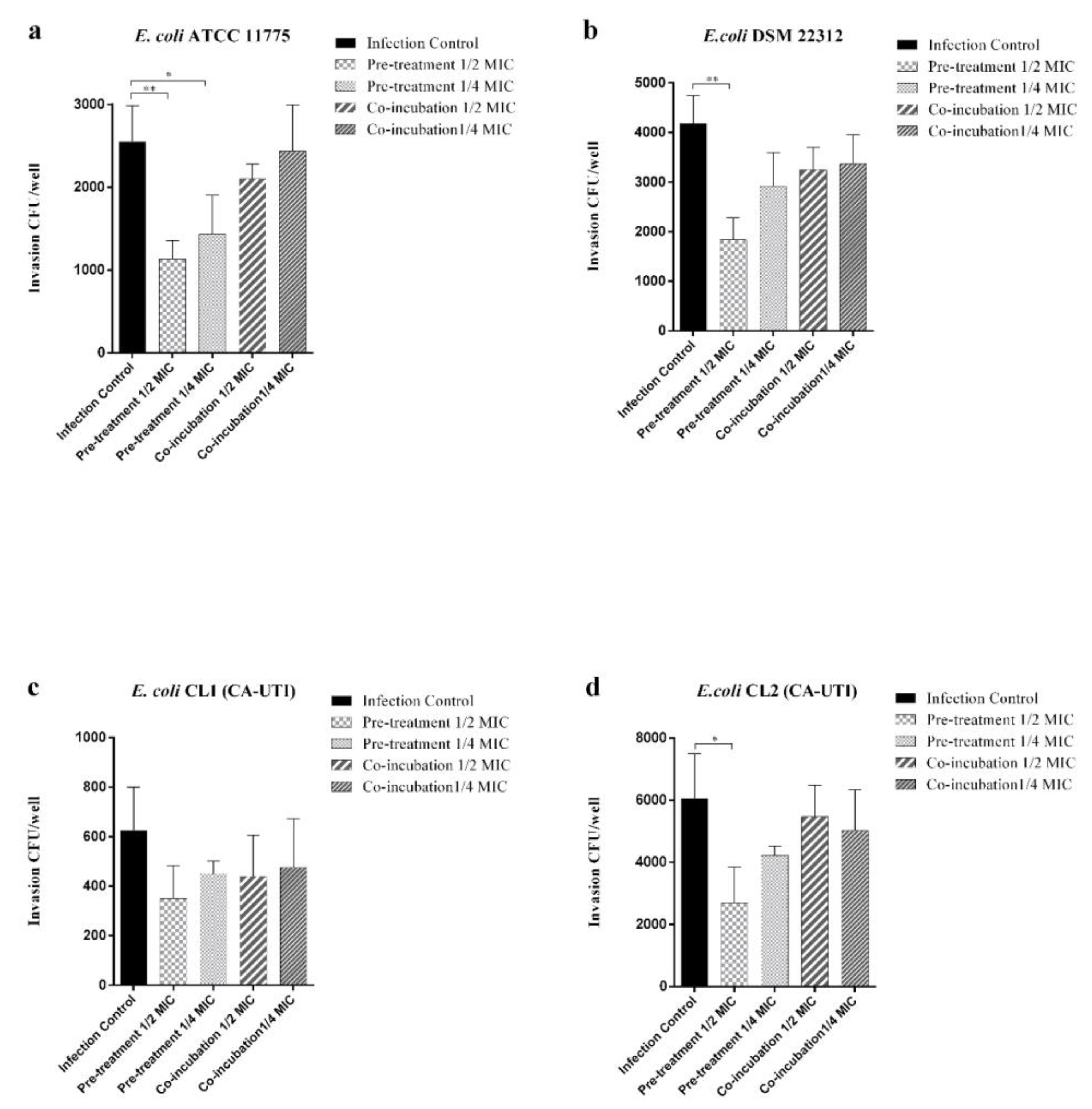
2.3. Effect of Berberine on the Adhesion and Invasion of E. coli Strains to Human 5637 ATCC (HTB-9) Cell Line
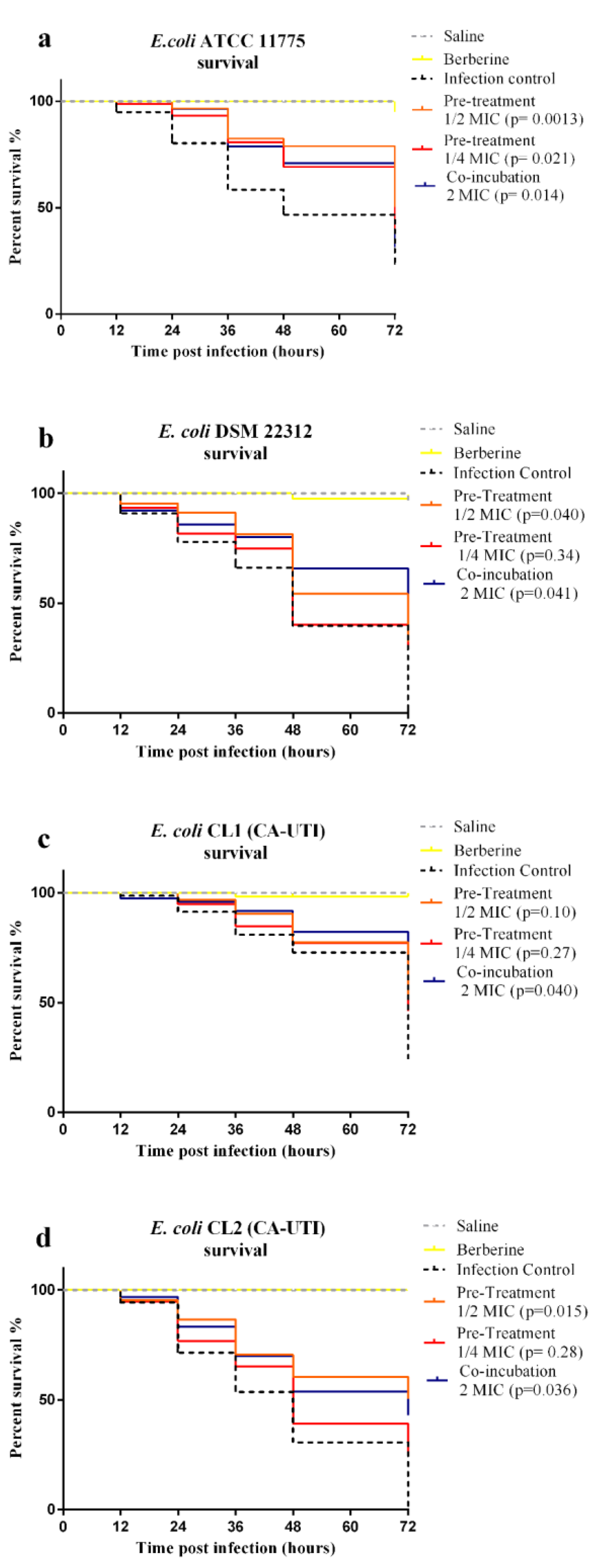
2.4. Infection Assays
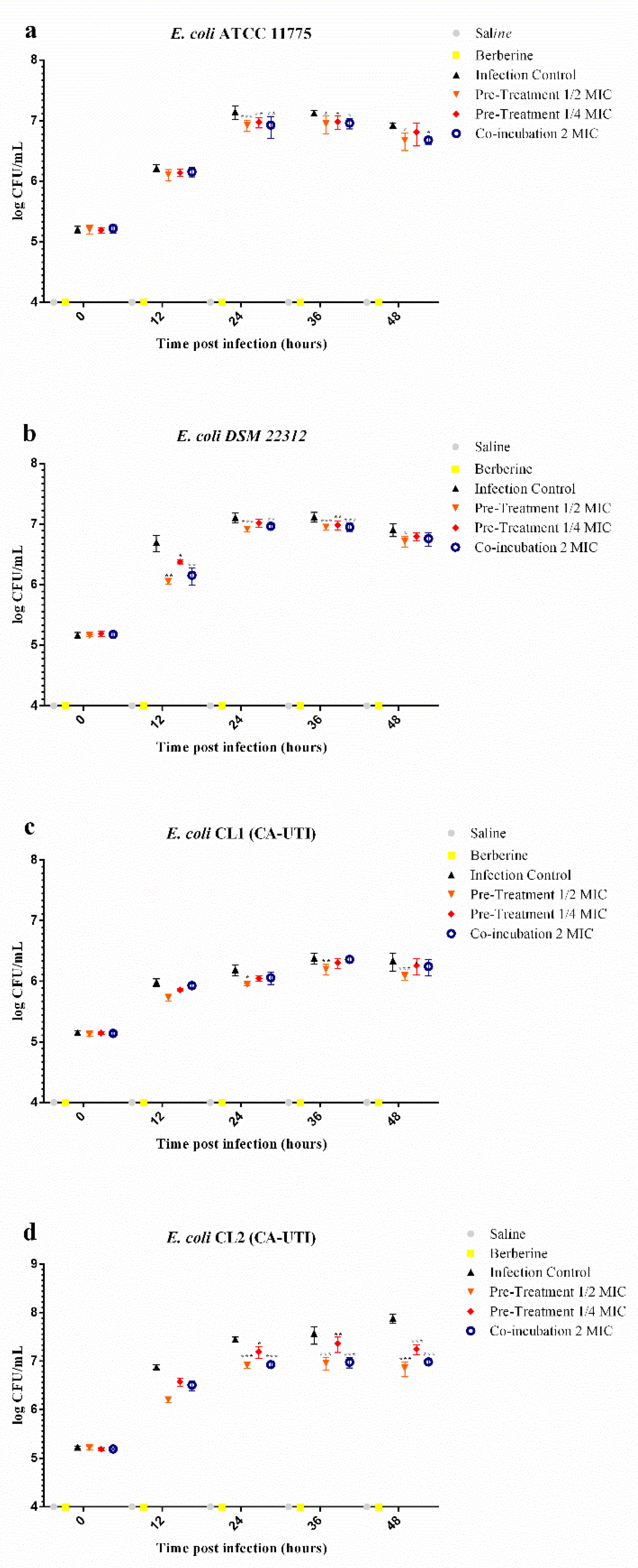
2.5. Effects of Berberine Chloride on E. coli Recovery (CFU/mL) from G. mellonella Haemolymph
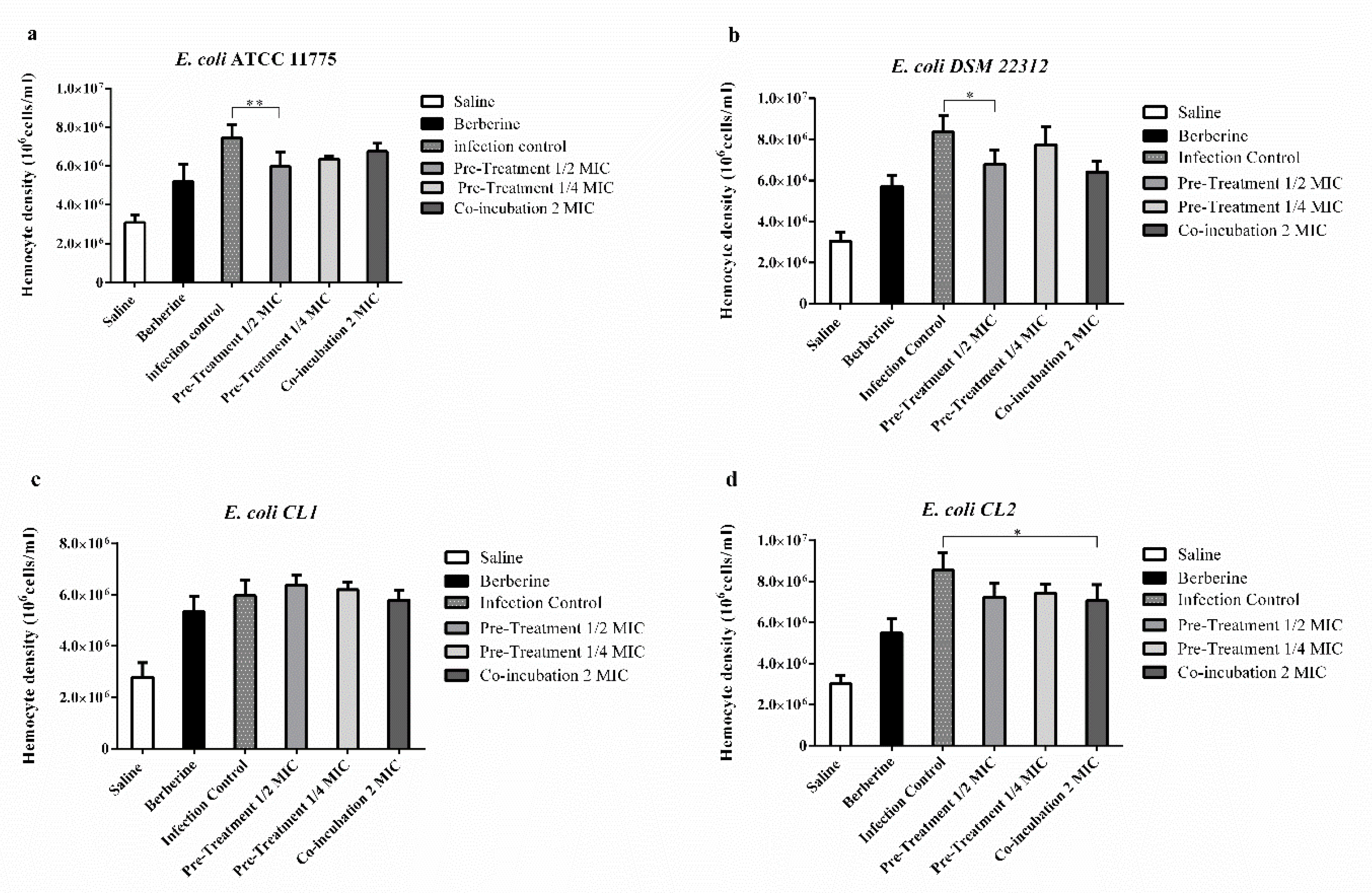
2.6. Effects of Berberine Chloride on the Enumeration of G. mellonella Hemocytes
3. Discussion
4. Materials and Methods
4.1. Chemicals and Culture Medium
4.2. Bacterial Strains, Cell Line and Larvae
4.3. Berberine MIC Determination
4.4. Influence of Berberine on Bacterial In Vitro Growth
4.5. In Vitro Bacterial Grown Curve Analysis
4.6. Effect of Berberine on E. coli Strains Adhesion and Invasion to Human Bladder Cells ATCC 5637 HTB-9: Co-Incubation and Pre-Treatment Experiments
4.7. G. mellonella Infection: Co-Incubation and Pre-Treatment Assays
4.8. G. mellonella Haemolymph Collection, E. coli Strains Recovery (CFU/mL) and Haemocytes Enumeration
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Muralidharan, S.; Kumar, J.R.; Dhanaraj, S.A. Estimation of berberine in berberis extracts by RP-HPLC. J. Pharm. Res. 2012, 5, 2065–2067. [Google Scholar]
- Dulić, M.; Ciganović, P.; Vujić, L.; Končić, M.Z. Antidiabetic and Cosmeceutical Potential of Common Barbery (Berberis vulgaris L.) Root Bark Extracts Obtained by Optimization of ‘Green’Ultrasound-Assisted Extraction. Molecules 2019, 24, 3613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Khalki, L.; Maire, V.; Dubois, T.; Zyad, A. Berberine Impairs the Survival of Triple Negative Breast Cancer Cells: Cellular and Molecular Analyses. Molecules 2020, 25, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habtemariam, S. Recent Advances in Berberine Inspired Anticancer Approaches: From Drug Combination to Novel Formulation Technology and Derivatization. Molecules 2020, 25, 1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imenshahidi, M.; Hosseinzadeh, H. Berberine and barberry (Berberis vulgaris): A clinical review. Phytother. Res. 2019, 33, 50–523. [Google Scholar] [CrossRef]
- Pasqual, M.S.; Lauer, C.P.; Moyna, P.; Henriques, J.A.P. Genotoxicity of the isoquinoline alkaloid berberine in prokaryotic and eukaryotic organisms. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 1993, 286, 243–252. [Google Scholar] [CrossRef]
- Guamán Ortiz, L.M.; Lombardi, P.; Tillhon, M.; Scovassi, A.I. Berberine, an epiphany against cancer. Molecules 2014, 19, 12349–12367. [Google Scholar] [CrossRef]
- Čerňáková, M.; Košťálová, D. Antimicrobial activity of berberine—A constituent ofMahonia aquifolium. Folia Microbiol. 2002, 47, 375–378. [Google Scholar] [CrossRef]
- Yan, D.; Jin, C.; Xiao, X.-H.; Dong, X.-P. Antimicrobial properties of berberines alkaloids in Coptis chinensis Franch by microcalorimetry. J. Biochem. Biophys. Methods 2008, 70, 845–849. [Google Scholar] [CrossRef]
- Chu, M.; Zhang, M.-B.; Liu, Y.-C.; Kang, J.-R.; Chu, Z.-Y.; Yin, K.-L.; Ding, L.-Y.; Ding, R.; Xiao, R.-X.; Yin, Y.-N. Role of berberine in the treatment of methicillin-resistant Staphylococcus aureus infections. Sci. Rep. 2016, 6, 24748. [Google Scholar] [CrossRef] [Green Version]
- Guo, N.; Zhao, X.; Li, W.; Shi, C.; Meng, R.; Liu, Z.; Yu, L. The synergy of berberine chloride and totarol against Staphylococcus aureus grown in planktonic and biofilm cultures. J. Med. Microbiol. 2015, 64, 891–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesh, R.; Shrestha, D.; Bhattachan, B.; Rai, G. Epidemiology of urinary tract infection and antimicrobial resistance in a pediatric hospital in Nepal. BMC Infect. Dis. 2019, 19, 420. [Google Scholar] [CrossRef] [PubMed]
- Karam, M.R.A.; Habibi, M.; Bouzari, S. Urinary tract infection: Pathogenicity, antibiotic resistance and development of effective vaccines against Uropathogenic Escherichia coli. Mol. Immunol. 2019, 108, 56–67. [Google Scholar] [CrossRef]
- Öztürk, R.; Murt, A. Epidemiology of urological infections: A global burden. World J. Urol. 2020, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Stamm, W.E.; Hooton, T.M. Management of urinary tract infections in adults. N. Engl. J. Med. 1993, 329, 1328–1334. [Google Scholar]
- Foxman, B. Epidemiology of urinary tract infections: Incidence, morbidity, and economic costs. Am. J. Med. 2002, 113, 5–13. [Google Scholar] [CrossRef]
- Linhares, I.; Raposo, T.; Rodrigues, A.; Almeida, A. Frequency and antimicrobial resistance patterns of bacteria implicated in community urinary tract infections: A ten-year surveillance study (2000–2009). BMC Infect. Dis. 2013, 13, 19. [Google Scholar] [CrossRef] [Green Version]
- Khoshnood, S.; Heidary, M.; Mirnejad, R.; Bahramian, A.; Sedighi, M.; Mirzaei, H. Drug-resistant gram-negative uropathogens: A review. Biomed. Pharmacother. 2017, 94, 982–994. [Google Scholar] [CrossRef]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef]
- Wojciuk, B.; Salabura, A.; Grygorcewicz, B.; Kędzierska, K.; Ciechanowski, K.; Dołęgowska, B. Urobiome: In Sickness and in Health. Microorganisms 2019, 7, 548. [Google Scholar] [CrossRef] [Green Version]
- Laupland, K.; Ross, T.; Pitout, J.; Church, D.; Gregson, D. Community-onset urinary tract infections: A population-based assessment. Infection 2007, 35, 150. [Google Scholar] [CrossRef]
- Bien, J.; Sokolova, O.; Bozko, P. Role of uropathogenic Escherichia coli virulence factors in development of urinary tract infection and kidney damage. Int. J. Nephrol. 2012, 2012. [Google Scholar]
- Shah, C.; Baral, R.; Bartaula, B.; Shrestha, L.B. Virulence factors of uropathogenic Escherichia coli (UPEC) and correlation with antimicrobial resistance. BMC Microbiol. 2019, 19, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bue, A.M.L.; Blandino, G.; Milazzo, I.; Cali, G.; Rossetti, B.; Marco, R.D. Microbiological and clinical periodontal effects of fixed orthodontic appliances in pediatric patients. Microbiol. Q. J. Microbiol. Sci. 2008, 31, 299. [Google Scholar]
- Rijavec, M.; Müller-Premru, M.; Zakotnik, B.; Žgur-Bertok, D. Virulence factors and biofilm production among Escherichia coli strains causing bacteraemia of urinary tract origin. J. Med. Microbiol. 2008, 57, 1329–1334. [Google Scholar] [CrossRef] [Green Version]
- Lewis, A.J.; Richards, A.C.; Mulvey, M.A. Invasion of host cells and tissues by uropathogenic bacteria. Urin. Tract Infect. Mol. Pathog. Clin. Manag. 2017, 4, 359–381. [Google Scholar]
- Mulvey, M.A. Adhesion and entry of uropathogenic Escherichia coli. Cell. Microbiol. 2002, 4, 257–271. [Google Scholar] [CrossRef]
- Vergalito, F.; Pietrangelo, L.; Petronio, G.P.; Colitto, F.; Cutuli, M.A.; Magnifico, I.; Venditti, N.; Guerra, G.; Di Marco, R. Vitamin E for prevention of biofilm-caused Healthcare-associated infections. Open Med. 2019, 15, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Bonkat, G.; Bartoletti, R.; Bruyère, F.; Cai, T.; Geerlings, S.E.; Koves, B.; Schubert, S.; Wagenlehner, F.; Mezei, T.; Pilatz, A.; et al. EAU Guidelines on Urological Infections; European Association of Urology: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Adamus-Białek, W.; Baraniak, A.; Wawszczak, M.; Głuszek, S.; Gad, B.; Wróbel, K.; Bator, P.; Majchrzak, M.; Parniewski, P. The genetic background of antibiotic resistance among clinical uropathogenic Escherichia coli strains. Mol. Biol. Rep. 2018, 45, 1055–1065. [Google Scholar] [CrossRef] [Green Version]
- Adwan, K.; Jarrar, N.; Abu-Hijleh, A.; Adwan, G.; Awwad, E. Molecular characterization of Escherichia coli isolates from patients with urinary tract infections in Palestine. J. Med. Microbiol. 2014, 63, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Kostakioti, M.; Hultgren, S.J.; Hadjifrangiskou, M. Molecular blueprint of uropathogenic Escherichia coli virulence provides clues toward the development of anti-virulence therapeutics. Virulence 2012, 3, 592–593. [Google Scholar] [CrossRef] [Green Version]
- Bader, M.S.; Loeb, M.; Leto, D.; Brooks, A.A. Treatment of urinary tract infections in the era of antimicrobial resistance and new antimicrobial agents. Postgrad. Med. 2019, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Del Fatti, C.; Jafari, N.; Hubbard, A.; Khasriya, R.; Roberts, A.; Rohn, J. The molecular basis of antibiotic treatment failure in chronic urinary tract infections. Access Microbiol. 2020, 2, 43. [Google Scholar]
- Nørgaard, S.M.; Jensen, C.S.; Aalestrup, J.; Vandenbroucke-Grauls, C.M.; De Boer, M.G.; Pedersen, A.B. Choice of therapeutic interventions and outcomes for the treatment of infections caused by multidrug-resistant gram-negative pathogens: A systematic review. Antimicrob. Resist. Infect. Control 2019, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Terlizzi, M.E.; Gribaudo, G.; Maffei, M.E. UroPathogenic Escherichia coli (UPEC) infections: Virulence factors, bladder responses, antibiotic, and non-antibiotic antimicrobial strategies. Front. Microbiol. 2017, 8, 1566. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, G.; Akram, M.; Jabeen, F.; Ali-Shah, S.M.; Munir, N.; Daniyal, M.; Riaz, M.; Tahir, I.M.; Ghauri, A.O.; Sultana, S. Therapeutic potential of medicinal plants for the management of urinary tract infection: A systematic review. Clin. Exp. Pharmacol. Physiol. 2019, 46, 613–624. [Google Scholar] [CrossRef]
- Blandino, G.; Fazio, D.; Di Marco, R. Probiotics: Overview of microbiological and immunological characteristics. Expert Rev. Anti-Infect. Ther. 2008, 6, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Cavallaro, V.; Catania, V.; Bonaccorso, R.; Mazzone, S.; Speciale, A.; Di Marco, R.; Blandino, G.; Caccamo, F. Effect of a broad-spectrum cephalosporin on the oral and intestinal microflora in patients undergoing colorectal surgery. J. Chemother. 1992, 4, 82–87. [Google Scholar] [CrossRef]
- Genovese, C.; Davinelli, S.; Mangano, K.; Tempera, G.; Nicolosi, D.; Corsello, S.; Vergalito, F.; Tartaglia, E.; Scapagnini, G.; Di Marco, R. Effects of a new combination of plant extracts plus d-mannose for the management of uncomplicated recurrent urinary tract infections. J. Chemother. 2018, 30, 107–114. [Google Scholar] [CrossRef]
- Geetha, R.; Roy, A.; Lakshmi, T. Nature’s weapon against urinary tract infections. Int. J. Drug Dev. Res 2011, 3, 85–100. [Google Scholar]
- Head, K.A. Natural approaches to prevention and treatment of infections of the lower urinary tract. Altern. Med. Rev. 2008, 13, 227–244. [Google Scholar] [PubMed]
- Furneri, P.M.; Fuochi, V.; Lissandrello, E.; Petroniox, G.P.; Petronio, P.; Fresta, M.; Paolino, D. The Antimicrobial Activity of Essential Oils Against Multi-Drug-Resistance Microorganisms: A Review. Front. Anti-Infect. Drug Discov. 2017, 5, 23–54. [Google Scholar]
- Ebani, V.V.; Nardoni, S.; Bertelloni, F.; Pistelli, L.; Mancianti, F. Antimicrobial activity of five essential oils against bacteria and fungi responsible for urinary tract infections. Molecules 2018, 23, 1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsson, B.; Jonasson, A.; Fianu, S. Prophylactic effect of UVA-E in women with recurrent cystitis: A preliminary report. Curr. Ther. Res. 1993, 53, 441–443. [Google Scholar] [CrossRef]
- Boberek, J.M.; Stach, J.; Good, L. Genetic evidence for inhibition of bacterial division protein FtsZ by berberine. PLoS ONE 2010, 5, e13745. [Google Scholar] [CrossRef]
- Sun, D.; Abraham, S.N.; Beachey, E.H. Influence of berberine sulfate on synthesis and expression of Pap fimbrial adhesin in uropathogenic Escherichia coli. Antimicrob. Agents Chemother. 1988, 32, 1274–1277. [Google Scholar] [CrossRef] [Green Version]
- Wullt, B. The Role Of P Fimbriae For Escherichia coli Establishment and Mucosal Inflammation in the Human Urinary Tract. Ph. D. Thesis, Lund University, Lund, Sweden, 2001. [Google Scholar]
- Gokgoz, N.B.; Avci, F.G.; Yoneten, K.K.; Alaybeyoglu, B.; Ozkirimli, E.; Sayar, N.A.; Kazan, D.; Sariyar Akbulut, B. Response of Escherichia coli to prolonged berberine exposure. Microb. Drug Resist. 2017, 23, 531–544. [Google Scholar] [CrossRef]
- Sun, T.; Li, X.-D.; Hong, J.; Liu, C.; Zhang, X.-L.; Zheng, J.-P.; Xu, Y.-J.; Ou, Z.-Y.; Zheng, J.-L.; Yu, D.-J. Inhibitory Effect of Two Traditional Chinese Medicine Monomers, Berberine and Matrine, on the Quorum Sensing System of Antimicrobial-Resistant Escherichia coli. Front. Microbiol. 2019, 10, 2584. [Google Scholar] [CrossRef]
- Guerrieri, C.G.; Pereira, M.F.; Galdino, A.C.M.; Dos Santos, A.L.S.; Elias, W.P.; Schuenck, R.P.; Spano, L.C. Typical and atypical enteroaggregative Escherichia coli are both virulent in the Galleria mellonella model. Front. Microbiol. 2019, 10, 1791. [Google Scholar] [CrossRef] [Green Version]
- Cutuli, M.A.; Petronio, G.; Vergalito, F.; Magnifico, I.; Pietrangelo, L.; Venditti, N.; Di Marco, R. Galleria mellonella as a consolidated in vivo model hosts: New developments in antibacterial strategies and novel drug testing. Virulence 2019, 10, 527–541. [Google Scholar] [CrossRef] [Green Version]
- Regeimbal, J.M.; Jacobs, A.C.; Corey, B.W.; Henry, M.S.; Thompson, M.G.; Pavlicek, R.L.; Quinones, J.; Hannah, R.M.; Ghebremedhin, M.; Crane, N.J. Personalized therapeutic cocktail of wild environmental phages rescues mice from Acinetobacter baumannii wound infections. Antimicrob. Agents Chemother. 2016, 60, 5806–5816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betts, J.W.; Hornsey, M.; Wareham, D.W.; La Ragione, R.M. In vitro and in vivo activity of theaflavin–epicatechin combinations versus multidrug-resistant acinetobacter baumannii. Infect. Dis. Ther. 2017, 6, 435–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojda, I. Immunity of the greater wax moth Galleria mellonella. Insect Sci. 2017, 24, 342–357. [Google Scholar] [CrossRef]
- Freires, I.A.; Sardi, J.d.C.O.; de Castro, R.D.; Rosalen, P.L. Alternative animal and non-animal models for drug discovery and development: Bonus or burden? Pharm. Res. 2017, 34, 681–686. [Google Scholar] [CrossRef]
- Santos, A.C.M.; Silva, R.M.; Valiatti, T.B.; Santos, F.F.; Santos-Neto, J.F.; Cayô, R.; Streling, A.P.; Nodari, C.S.; Gales, A.C.; Carvalho, E. Virulence potential of a multidrug-resistant Escherichia coli strain belonging to the emerging clonal group ST101-B1 isolated from bloodstream infection. Microorganisms 2020, 8, 827. [Google Scholar] [CrossRef]
- Klein, R.D.; Hultgren, S.J. Urinary tract infections: Microbial pathogenesis, host–pathogen interactions and new treatment strategies. Nat. Rev. Microbiol. 2020, 18, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Karaosmanoglu, K.; Sayar, N.A.; Kurnaz, I.A.; Akbulut, B.S. Assessment of Berberine as a multi-target antimicrobial: A multi-omics study for drug discovery and repositioning. Omics J. Integr. Biol. 2014, 18, 42–53. [Google Scholar] [CrossRef]
- Jin, J.; Hua, G.; Meng, Z.; Gao, P. Antibacterial mechanisms of berberine and reasons for little resistance of bacteria. Chin. Herb. Med 2010, 3, 27–35. [Google Scholar]
- Tabasi, M.; Karam, M.R.A.; Habibi, M.; Yekaninejad, M.S.; Bouzari, S. Phenotypic assays to determine virulence factors of uropathogenic Escherichia coli (UPEC) isolates and their correlation with antibiotic resistance pattern. Osong Public health Res. Perspect. 2015, 6, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Domadia, P.N.; Bhunia, A.; Sivaraman, J.; Swarup, S.; Dasgupta, D. Berberine targets assembly of Escherichia coli cell division protein FtsZ. Biochemistry 2008, 47, 3225–3234. [Google Scholar] [CrossRef]
- Ozbalci, Ç.; Unsal, Ç.; Kazan, D.; Sariyar-Akbulut, B. Proteomic response of Escherichia coli to the alkaloid extract of Papaver polychaetum. Ann. Microbiol. 2010, 60, 709–717. [Google Scholar] [CrossRef]
- Pierpaoli, E.; Cirioni, O.; Simonetti, O.; Orlando, F.; Giacometti, A.; Lombardi, P.; Provinciali, M. Potential application of berberine in the treatment of Escherichia coli sepsis. Nat. Prod. Res. 2020, 1–6. [Google Scholar] [CrossRef]
- Zhang, H.; Shan, Y.; Wu, Y.; Xu, C.; Yu, X.; Zhao, J.; Yan, J.; Shang, W. Berberine suppresses LPS-induced inflammation through modulating Sirt1/NF-κB signaling pathway in RAW264. 7 cells. Int. Immunopharmacol. 2017, 52, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Mahdavi Siuki, M.; Fazel Nasab, N.; Barati, E.; Nezafat Firizi, M.; Jalilvand, T.; Namdar Ahmadabad, H. The protective effect of berberine against lipopolysaccharide-induced abortion by modulation of inflammatory/immune responses. Immunopharmacol. Immunotoxicol. 2018, 40, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.; Ding, R.; Chu, Z.-Y.; Zhang, M.-B.; Liu, X.-Y.; Xie, S.-H.; Zhai, Y.-J.; Wang, Y.-D. Role of berberine in anti-bacterial as a high-affinity LPS antagonist binding to TLR4/MD-2 receptor. BMC Complement. Altern. Med. 2014, 14, 89. [Google Scholar] [CrossRef] [Green Version]
- Rowan, R.; Moran, C.; McCann, M.; Kavanagh, K. Use of Galleria mellonella larvae to evaluate the in vivo anti-fungal activity of [Ag 2 (mal)(phen) 3]. Biometals 2009, 22, 461. [Google Scholar] [CrossRef] [Green Version]
- Imanshahidi, M.; Hosseinzadeh, H. Pharmacological and therapeutic effects of Berberis vulgaris and its active constituent, berberine. Phytother. Res. 2008, 22, 999–1012. [Google Scholar] [CrossRef]
- Abd El-Wahab, A.E.; Ghareeb, D.A.; Sarhan, E.E.; Abu-Serie, M.M.; El Demellawy, M.A. In vitro biological assessment of Berberis vulgaris and its active constituent, berberine: Antioxidants, anti-acetylcholinesterase, anti-diabetic and anticancer effects. BMC Complement. Altern. Med. 2013, 13, 218. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Sharma, B. Toxicological effects of berberine and sanguinarine. Front. Mol. Biosci. 2018, 5, 21. [Google Scholar] [CrossRef] [Green Version]
- Wojtyczka, R.D.; Dziedzic, A.; Kępa, M.; Kubina, R.; Kabała-Dzik, A.; Mularz, T.; Idzik, D. Berberine enhances the antibacterial activity of selected antibiotics against coagulase-negative Staphylococcus strains in vitro. Molecules 2014, 19, 6583–6596. [Google Scholar] [CrossRef]
- Zhou, X.-Y.; Ye, X.-G.; He, L.-T.; Zhang, S.-R.; Wang, R.-L.; Zhou, J.; He, Z.-S. In vitro characterization and inhibition of the interaction between ciprofloxacin and berberine against multidrug-resistant Klebsiella pneumonia e. J. Antibiot. 2016, 69, 741–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wultańska, D.; Piotrowski, M.; Pituch, H. The effect of berberine chloride and/or its combination with vancomycin on the growth, biofilm formation, and motility of Clostridioides difficile. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Malavé, A. Protective effect of berberine on cyclophosphamide-induced haemorrhagic cystitis in rats. Pharmacol. Toxicol. 2001, 88, 232–237. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Nicolosi, D.; Genovese, C.; Cutuli, M.A.; D’Angeli, F.; Pietrangelo, L.; Davinelli, S.; Petronio, G.P.; Marco, R.D. Preliminary in Vitro Studies on Corynebacterium urealyticum Pathogenetic Mechanisms, a Possible Candidate for Chronic Idiopathic Prostatitis? Microorganisms 2020, 8, 463. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, S.; Kamolvit, W.; Zambrana, S.; Sandström, C.; Gonzales, E.; Östenson, C.-G.; Brauner, A. Extract of Clinopodium bolivianum protects against E. coli invasion of uroepithelial cells. J. Ethnopharmacol. 2017, 198, 214–220. [Google Scholar] [CrossRef]
- Jorjão, A.L.; de Oliveira, F.E.; Leão, M.V.P.; Jorge, A.O.C.; de Oliveira, L.D. Effect of Lactobacillus rhamnosus on the response of Galleria mellonella against Staphylococcus aureus and Escherichia coli infections. Arch. Microbiol. 2018, 200, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Heitmueller, M.; Billion, A.; Dobrindt, U.; Vilcinskas, A.; Mukherjee, K. Epigenetic mechanisms regulate innate immunity against uropathogenic and commensal-like Escherichia coli in the surrogate insect model Galleria mellonella. Infect. Immun. 2017, 85, e00336. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Berberine |
|---|---|
| E. coli ATCC 11775 | 1024 |
| E. coli DSM 22312 | 2048 |
| CL1 (CA-UTI) | 1024 |
| CL2 (CA-UTI) | 2048 |
| E. coli ATCC 1775 | E. coli DSM 22312 | E. coli CL1 | E. coli CL2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CNT | ¼ MIC | ½ MIC | CNT | ¼ MIC | ½ MIC | CNT | ¼ MIC | ½ MIC | CNT | ¼ MIC | ½ MIC | |
| Goodness of Fit R2 | 0.998 | 0.998 | 0.998 | 0.997 | 0.995 | 0.999 | 0.996 | 0.997 | 0.998 | 0.997 | 0.998 | 0.999 |
| YTop (CFU/m) | 9.66 × 108 | 8.45 × 108 | 6.12 × 108 | 1.03 × 109 | 7.19 × 108 | 5.87 × 108 | 9.59 × 108 | 7.75 × 108 | 5.49 × 108 | 1.04 × 109 | 8.58 × 108 | 7.79 × 108 |
| Ytop ± S.D (CFU/mL) | 1.98 × 107 | 1.54 × 107 | 2.11 × 107 | 3.00 × 107 | 2.92 × 107 | 9.00 × 106 | 2.95 × 107 | 2.17 × 107 | 1.08 × 107 | 3.30 × 107 | 2.36 × 107 | 1.13 × 107 |
| Inhibition (%) | / | 12.47 | 36.61 | / | 30.29 | 43.03 | / | 19.25 | 42.72 | / | 17.75 | 25.35 |
| XCFU50 (h) | 5.87 | 8.58 | 9.03 | 4.99 | 7.63 | 8.76 | 5.05 | 6.26 | 6.86 | 5.79 | 6.69 | 8.42 |
| XCFU50 ± S.D. (h) | 0.35 | 0.3 | 0.28 | 0.57 | 0.5 | 0.16 | 0.62 | 0.44 | 0.24 | 0.62 | 0.5 | 0.2 |
| ∆XCFU50 (h) | / | 2.71 | 3.16 | / | 2.64 | 3.77 | / | 1.21 | 1.81 | / | 2.63 | 0.9 |
| HillSlope | 0.24 | 0.24 | 0.27 | 0.25 | 0.28 | 0.3 | 0.24 | 0.25 | 0.31 | 0.22 | 0.19 | 0.20 |
| HillSlope ± S.D | 0.039 | 0.037 | 0.043 | 0.06 | 0.081 | 0.032 | 0.064 | 0.054 | 0.049 | 0.054 | 0.036 | 0.018 |
| ∆ HillSlope | / | 0 | 0.03 | / | 0.03 | 0.05 | / | 0.01 | 0.07 | / | −0.03 | −0.02 |
| E. coli ATCC 1175 | E. coli DSM 22318 | E. coli CL1 | E. coli CL2 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Log Units | p Value | Log Units | p Value | log Units | p Value | log Units | p Value | ||
| time post infection 12 h | Infection Control vs. Pre-Treatment 1/2 MIC | 0.1093 | >0.9999 | 0.6462 | 0.0014 | 0.2492 | 0.2989 | 0.6700 | 0.5716 |
| Infection Control vs. Pre-Treatment 1/4 MIC | 0.0767 | >0.9999 | 0.3220 | 0.0406 | 0.1255 | >0.9999 | 0.3000 | >0.9999 | |
| Infection Control vs. Co-incubation MIC | 0.0612 | >0.9999 | 0.5442 | 0.0032 | 0.0524 | >0.9999 | 0.3689 | >0.9999 | |
| time post infection 24 h | Infection Control vs. Pre-Treatment 1/2 MIC | 0.2225 | 0.001 | 0.1888 | 0.0002 | 0.2422 | 0.0329 | 0.5333 | 0.0002 |
| Infection Control vs. Pre-Treatment 1/4 MIC | 0.1706 | 0.0083 | 0.0906 | 0.0675 | 0.1411 | 0.2733 | 0.2519 | 0.0239 | |
| Infection Control vs. Co-incubation MIC | 0.2191 | 0.0011 | 0.1428 | 0.0032 | 0.1304 | 0.3411 | 0.5190 | 0.0002 | |
| time post infection 36 h | Infection Control vs. Pre-Treatment 1/2 MIC | 0.1761 | 0.0101 | 0.1708 | 0.0004 | 0.1849 | 0.005 | 0.6078 | <0.0001 |
| Infection Control vs. Pre-Treatment 1/4 MIC | 0.1462 | 0.0323 | 0.1364 | 0.0036 | 0.0807 | 0.3198 | 0.2036 | 0.0096 | |
| Infection Control vs. Co-incubation MIC | 0.1648 | 0.0156 | 0.1676 | 0.0005 | 0.0214 | >0.9999 | 0.5896 | <0.0001 | |
| time post infection 48 h | Infection Control vs. Pre-Treatment 1/2 MIC | 0.2486 | 0.0432 | 0.1964 | 0.0177 | 0.2520 | 0.0011 | 1.0291 | <0.0001 |
| Infection Control vs. Pre-Treatment 1/4 MIC | 0.1105 | 0.5842 | 0.1163 | 0.2028 | 0.0791 | 0.4485 | 0.6337 | <0.0001 | |
| Infection Control vs. Co-incubation MIC | 0.2425 | 0.0485 | 0.1491 | 0.0758 | 0.0968 | 0.2572 | 0.9015 | <0.0001 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petronio Petronio, G.; Cutuli, M.A.; Magnifico, I.; Venditti, N.; Pietrangelo, L.; Vergalito, F.; Pane, A.; Scapagnini, G.; Di Marco, R. In Vitro and In Vivo Biological Activity of Berberine Chloride against Uropathogenic E. coli Strains Using Galleria mellonella as a Host Model. Molecules 2020, 25, 5010. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25215010
Petronio Petronio G, Cutuli MA, Magnifico I, Venditti N, Pietrangelo L, Vergalito F, Pane A, Scapagnini G, Di Marco R. In Vitro and In Vivo Biological Activity of Berberine Chloride against Uropathogenic E. coli Strains Using Galleria mellonella as a Host Model. Molecules. 2020; 25(21):5010. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25215010
Chicago/Turabian StylePetronio Petronio, Giulio, Marco Alfio Cutuli, Irene Magnifico, Noemi Venditti, Laura Pietrangelo, Franca Vergalito, Antonella Pane, Giovanni Scapagnini, and Roberto Di Marco. 2020. "In Vitro and In Vivo Biological Activity of Berberine Chloride against Uropathogenic E. coli Strains Using Galleria mellonella as a Host Model" Molecules 25, no. 21: 5010. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25215010